
Abstract
Cerebellar abnormalities are commonly associated with hydrocephalus. However, the effect of hydrocephalus on the otherwise normal cerebellum has been largely neglected. This study assesses the morphological changes in the Purkinje cells in relation to cerebellar dysfunction observed in juvenile hydrocephalic rats. Fifty-five three-week old albino Wistar rats were used, hydrocephalus was induced by intracisternal injection of kaolin (n = 35) and others served as controls (n = 20). Body weight measurements, hanging wire, negative geotaxis, and open field tests were carried out at the onset and then weekly for 4 weeks, rats were killed, and their cerebella processed for Hematoxylin and Eosin, Cresyl violet and Golgi staining. Qualitative and quantitative studies were carried out; quantitative data were analyzed using two-way ANOVA and independent T tests at p < 0.05. Hydrocephalic rats weighed less than controls (p = 0.0247) but their cerebellar weights were comparable. The hydrocephalic rats had a consistently shorter latency to fall in the hanging wire test (F(4,112) = 18.63; p < 0.0001), longer latency to turn in the negative geotaxis test (F(4,112) = 22.2; p < 0.0001), and decreased horizontal (F(4,112) = 4.172, p = 0.0035) and vertical movements (F(4,112) = 4.397; p = 0.0024) in the open field test than controls throughout the 4 weeks post-induction. Cellular compression in the granular layer, swelling of Purkinje cells with vacuolations, reduced dendritic arborization and increased number of pyknotic Purkinje cells were observed in hydrocephalic rats. Hydrocephalus caused functional and morphological changes in the cerebellar cortex. Purkinje cell loss, a major pathological feature of hydrocephalus, may be responsible for some of the motor deficits observed in this condition.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Hydrocephalus is a neurological disorder characterized by dilatation of the ventricular system, usually because cerebrospinal fluid produced therein is not fully returned into the systemic circulation (Khan et al. 2006; Del Bigio 2010). Alterations in neuronal cytology have been identified at various stages of hydrocephalus (Wallace et al. 2010), for example, pyknotic and dark shrunken neurons observed in the cerebral cortex of hydrocephalic cats (Di Curzio et al. 2013); but other studies suggest that neuron degeneration or death is not a major pathological feature of hydrocephalus (Rubin et al. 1972; Jones et al. 1991; Hale et al. 1992). Attenuation of white matter and oligodendrocyte loss occur prominently in hydrocephalus and are associated with many of the neurological deficits observed (Olopade et al. 2012).
The mammalian cerebellum is a functionally diverse component of the brain which is crucial for the performance of smooth and purposeful voluntary movements, permitting postural adjustments and the learning of new skills in the process (D’Angelo and Casali 2012). The cerebellar cortex is remarkably uniform, microscopically, and is deployed in a trilaminar array of cells into the molecular, Purkinje cell and granule cell layers. The Purkinje cell layer is perhaps the most significant of these because it is the sole output cell of the cerebellar cortex (Eccles et al. 1967; Palay and Chan-Palay 1974). Purkinje cells are particularly susceptible when the cerebellar cortex is subjected to genetic and acquired insults (Herrup and Wilczynski 1982; Welsh et al 2002).
Abnormalities of the cerebellum which lead to the obstruction of CSF are commonly associated with the development of hydrocephalus. However, the effect of ventricular enlargement on the structure and function of the otherwise normal cerebellum has not been well elucidated. The clinical reports of cerebellar effects of hydrocephalus include cerebellar dysarthria with severe cerebellar ataxia of all four limbs (Singh et al. 1988), swelling of the cerebellum and brain stem with abnormal enhancement of the basal meninges and effacement of the fourth ventricle (Uyanikgil et al. 2017). Other studies have reported derangement of the normal laminar cellular organization of the cerebellum and thickening of the outer molecular layer (Del Bigio et al. 2002). Progressive Purkinje cell loss and cerebellar granular layer aplasia leading to severe cerebellar ataxia have also been reported (Wallace et al. 2010).
A study of the cerebellum in hydrocephalus is important because the motor function decline observed in hydrocephalic patients should not be attributed solely to the effects of hydrocephalus on the motor cortex. Given the central role of Purkinje cells in motor coordination, the purpose of our study is to assess the morphological changes occurring in the Purkinje cells of the cerebellum in relation to cerebellar dysfunction observed in juvenile hydrocephalic rats.
Materials and methods
A total of 55 3-week old albino Wistar rats (35–51 g) obtained from the animal holding facility of the department of Anatomy, University of Ibadan were used for the study. The rats were divided into 2 groups of 35 experimental and 20 control animals. All experiments were approved by the Ethical Review Board of the University of Ibadan and all procedures on animal handling conformed to the acceptable guidelines on the ethical use of animals in research. The rats were allowed to acclimatize for 7 days in natural light–dark cycle at room temperature and were given food and water ad libitum.
Hydrocephalus was induced in the rats according to the protocol by Di Curzio et al. (2014). Briefly, the rats in the experimental group were anaesthetized with intraperitoneal injection of ketamine/xylazine (90/10 mg/kg) and had an intracisternal injection of 0.04 ml sterile kaolin suspension (250 mg/ml in distilled water) percutaneously, with a 27-gauge needle to induce hydrocephalus. In the control rats, a sham procedure was performed in which the cisterna magna was punctured without fluid injection. They were all monitored for about 1 h after the procedure and returned to their cages.
The rats were housed in groups of 4 and given food and water ad libitum. They were weighed twice weekly and assessed for the development of hydrocephalus such as increased head circumference, abnormal gait and dull general appearance.
Neurobehavioral tests
The following neurobehavioral tests were carried out at onset of the study, and then weekly for four weeks: hanging wire test, negative geotaxis test, and open field test. A total of 15 rats per group were used for these neurobehavioral tests.
Hanging wire test
This test assesses muscular strength in animals (Jansone et al. 2016). The forelimb paws of the rat were placed on a horizontally suspended metal wire (measuring 2 mm in diameter and 1 m in length), placed one meter above a landing surface which is filled with soft bedding. The length of the time each rat was able to stay suspended before falling off the wire (latency to fall) was recorded. A maximum time of 2 min was given to each rat after which it was removed. This test was performed weekly with three trials per session. The average performance for each session is presented as the average of the three trials.
Negative geotaxis test
This test is used to assess motor and vestibular function in rodents (Lubics et al. 2005; Fan et al. 2010). The rats were placed prone on a plane which was inclined downward at 45° to the horizontal; they were expected to turn and climb upward against gravity. Time taken to make a U-turn and start to walk upwards (latency to turn) was recorded. A maximum time of 30 s was given to each rat for the test (Rodriguez-Fanjul et al. 2017).
Open field test
The open field test was performed in a white painted wooden box with a floor area measuring 72 cm by 72 cm and walls 36 cm high. Black lines were drawn on the floor of the box to divide it into 18 cm by 18 cm squares. There is a square in the center of the open field also measuring 18 cm by 18 cm (Stanford 2007).
The rats were observed individually in the open field box for a period for 5 min each and the following parameters were assessed: line crossing (frequency with which the rat crossed one of the grid lines with all four paws), rearing (frequency with which the rat stood on their hind legs only), Center square duration (duration of time the rat spent in the center square), Center square entries (frequency with which the rats crossed one of the red lines with all four paws). All the parameters were observed and recorded manually by the same set of observers, blinded to the group assignments of the rats.
Tissue processing and analysis
The rats were sacrificed four weeks post-induction of hydrocephalus after the neurobehavioral tests had been completed. They were anaesthetized with intraperitoneal injection of 90 mg/kg ketamine, after which transcardiac perfusion was carried out using 10% neutral buffered formaldehyde (NBF). The brain was removed, cerebellum excised, weighed and fixed in 10% NBF. After 48 h, coronal sections of the cerebrum were made at the level of the optic chiasm to confirm the enlargement of the ventricle. The cerebella were processed, embedded in paraffin blocks and sectioned at 5 microns. The sections were stained with Hematoxylin and Eosin, and Nissl stains and examined histologically with a Leica DM 750 light microscope (Leica Microsystems, Heerbrugg, Switzerland 2016); representative pictures were taken from the two groups.
In the Nissl-stained slides, quantitative analysis was carried out in the lateral hemisphere of the cerebellum, blind of group assignment. Stereological counts of the normal Purkinje cells, that is, Purkinje cells with a clearly defined cell body and nucleus, were performed in representative fields of the lateral cerebellar hemisphere randomly selected, in two sections per rat and five rats per group at × 20 magnification. In the same area, the number of pyknotic cells, that is, cells with dark nuclei, loss or abnormal clumping of heterochromatin, were also counted and recorded. Pyknotic index was calculated using the equation below according to Taveira et al. (2012):
A subset of the fixed cerebella (five per group), underwent a modified Golgi staining. Briefly, the fixed samples were immersed in potassium dichromate solution (3 g/100 ml of distilled water) for 5 days (solution was replaced every 24 h), then transferred into silver nitrate solution (2 g/100 ml) for 3 days (also replaced every 24 h), infiltrated and embedded in paraffin wax. The paraffin blocks were sectioned at 60 μm, dehydrated in increasing concentrations of alcohol, cleared in xylene and mounted on adhesive glass slides and cover-slipped with DPX. The sections were viewed under a Leica DM 750 light microscope (Leica Microsystems, Heerbrugg, Switzerland 2016) at × 40 and × 100 magnifications; qualitative analysis of the Purkinje cells was carried out and compared between the two groups. Isolated pyramidal cells which had their cell body and processes well delineated were singled out for qualitative assessment and pictures taken.
Statistical analysis
Quantitative data from the behavioral tests and histological examination were expressed as means ± SEM statistically evaluated by the two-way analysis of variance (ANOVA) and independent t tests respectively, using GraphPad prism version 5.0 windows software (SanDiego, Califonia USA). Statistical significance was set at p < 0.05 and the confidence interval fixed at 95% level.
Results
Macroscopic observations
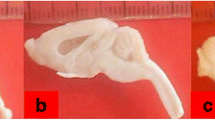
In hydrocephalic rats, the head was enlarged and dome-shaped. The eyeballs were proptosed and the hind limbs were splayed out and weak (Fig. 1). These features were evident from as early as one week after the intracisternal injection. Five of the rats that had intracisternal injection for induction of hydrocephalus died before the end of the study, giving a mortality rate of 14% of hydrocephalic rats. Three of them died immediately after induction of hydrocephalus, possibly due to brain stem injury and two died within 3 days of induction. Five of the rats that had kaolin injection failed to develop hydrocephalus and were thus removed from the study. None of the control rats died before the end of the study.

Figure showing control rat (a) and hydrocephalic rats (b and c). Blue arrow in (b) shows enlarged and dome shaped head, while yellow arrow in (c) shows protruded eyeball
Body weight The body weight of the hydrocephalic rats was found to be significantly less than that of the controls from the first to fourth weeks post induction. The hydrocephalic rats initially lost weight within the first few days of injection but later, a gradual increase was noticed. However, they had a significantly lower weight throughout the experimental period when compared to the control rats (Slopes: 1.671 ± 0.1937 vs. 2.194 ± 0.076 respectively, F = 6.329, DFn = 1, DFd = 14; p = 0.0247) (Fig. 2a).

Line graph comparing the body weight (g) over four weeks post injection (a), and bar chart showing the cerebellar to body weight index (b) of hydrocephalic rats and the controls
Cerebellar weight There was no significant difference in the cerebellar weight between the hydrocephalic rats and controls (0.395 ± 0.009 g vs 0.378 ± 0.0104 g respectively, t = 1.912, p = 0.0635). There was however a significant difference in the cerebellum to body weight index between the two groups (t = 2.362, p = 0.0227) (Fig. 2b).
Neurobehavioural studies
Hanging wire test
Before induction of hydrocephalus, both groups were comparable in latency to fall, however, after hydrocephalus induction, at each time point of measurement, the latency to fall was shorter in the hydrocephalic than in the control groups. The differences were statistically significant and were more pronounced with time (Interaction F (4, 112) = 18.63; p < 0.0001) (Fig. 3a).

Bar charts of Hanging wire test showing latency to fall (a), Negative geotaxis test showing latency to turn (b); and open field test showing line crossings (c) and frequency of rearing within 5 min (d) over four weeks post injection in hydrocephalic rats and controls. *p < 0.05, **p < 0.01, ****p < 0.0001
Negative geotaxis test Before induction of hydrocephalus, both groups were similar in latency to turn, and this decreased over time in both control and experimental rats. However, after hydrocephalus induction, the hydrocephalic rats took longer to reverse direction at every time point of measurement and the differences, which were statistically significant increased with time (Interaction F (4, 112) = 22.2; p < 0.0001) (Fig. 3b).
Open field test The following were observed while the rats were placed in the open field box.
-
a)
Line crossing: The number of lines crossed, signifying horizontal movements was significantly fewer in the hydrocephalic rats than in the control group (Interaction F (4, 112) = 4.172, p = 0.0035) (Fig. 3c).
-
b)
Rearing: Similarly, the hydrocephalic group was found to have a significantly lower frequency of rearing, that is, vertical movements when compared with the control group (Interaction F (4, 112) = 4.397; p = 0.0024) (Fig. 3d).
Histological observations
The cerebellar cortex in the control rats had Purkinje cells with a clearly defined cell body and nucleus, mostly arranged in a single layer between the granule cell and molecular layers. In the Purkinje cell layer of hydrocephalic rats, some Purkinje cells with vacuolations, irregular somatic outline, some shrunken and some swollen, with clumped heterochromatin and dark shrunken nuclei were observed. Some areas within the layer were found to be devoid of Purkinje cells; dying Purkinje cells with ill-defined cell body and no visible nucleus, were also observed (Fig. 4).

Photomicrograph of cerebellar cortex Control (a, c) and Hydrocephalic (b, d) rats; H&E × 40 and × 100 (inset) magnification. G: granule cell layer, P: Purkinje cell layer, M: molecular cell layer. In (a), yellow arrows show Purkinje cell with clearly defined cell body and nucleus. In (b), red arrows show Purkinje cells with vacuolation and black arrow shows irregular cell body of Purkinje cell, clumped heterochromatin and dark shrunken nucleus. In (c), yellow arrows show Purkinje cells with clearly defined cell body and nucleus. In (d), black arrows show dying Purkinje cells with loss of prominence of the nucleus; red arrows show gaps within the Purkinje layer depicting loss of Purkinje cells
The normal arrangement of granule cells within the granular layer observed in the control rats was distorted in the hydrocephalic ones. The granule cells were also more compacted in the hydrocephalic rats than in the controls (Fig. 5a,b). The population of normal/spared Purkinje cells was significantly less (59.6 ± 3.14 vs. 48.2 ± 2.615, t = 2.79, p = 0.0236) and the population of pyknotic cells significantly higher in the hydrocephalic group when compared to the control group. The pyknotic index was thus significantly higher in the hydrocephalic rats than in the controls (17.25 ± 1.15 vs. 29.47 ± 3.07%, t = 3.731, p = 0.0058) (Fig. 5c,d).

Photomicrograph of cerebellar cortex in control (a) and hydrocephalic (b) rats, Cresyl Violet stain × 40 magnification. G: granule cell layer, P: Purkinje cell layer, M: molecular cell layer. In (a), yellow arrow shows normal arrangement of granule cells in the granule cell layer and in (b), red arrow shows compacted granule cells in the granule cell layer. Graphical representation of the population of spared/normal Purkinje cells (c) and the pyknotic index (d) in the two groups (*p< 0.05, **p < 0.01)
In the control group, the flask-shaped Purkinje cell bodies were arranged in a single row between the outer molecular layer and the inner granule cells layer. Their extensive network of thread-like processes, dendritic arborizations, are easily seen. In the hydrocephalic rats, the Purkinje cell bodies were found to be smaller in size and had fewer dendritic arborizations than the controls (Fig. 6).

Photomicrograph of Purkinje cell (a, c) Control group, (b, d) Hydrocephalic group. Modified Golgi stain × 40 and × 100 (insets) magnificiation. In (a), red arrow shows the flask shape of the Purkinje cell, blue arrow shows dendritic aborizations of the Purkinje cell. In (b), red arrow shows a shrunken Purkinje cell body and blue arrow shows fewer dendritic aborizations of the Purkinje cell compared to the control group. Similarly, blue arrow in (c) shows the full dendritic arborizations of the Purkinje cell which is much reduced in (d)
Discussion
In this study, we have demonstrated degeneration and loss of Purkinje neurons in juvenile hydrocephalic albino rats. Surviving Purkinje neurons are smaller and with attenuation of their dendritic arbor. These changes occur in association with significant reduction in exploratory behavior, upper and lower limb strength and coordination.
Neurobehavioral deficits have been frequently described in hydrocephalus, in association with cerebral white matter damage. However, the role of injury to the Purkinje neuron is often overlooked and understated. Purkinje cell anomalies have been previously described in hydrocephalic states, but not as a direct consequence of the hydrocephalic distension of the ventricles. In human neonatal post hemorrhagic hydrocephalus, loss of these neurons occur in relation to cerebellar subarachnoid hemorrhage and olivocerebellar pathology (Fukumizu et al. 1995). In congenital hydrocephalus in H-Tx rats, Purkinje neurons show evidence of increased HMGB-1 expression and early features of apoptosis (Watanabe et al. 2013). It is however uncertain whether this was the result of the hydrocephalus or a manifestation of the genetic defect, which thus occurs prior to the development of hydrocephalus.
The cerebellum is a well-recognized neural entity in the pathophysiology of hydrocephalus. There are several well described roles of this organ in the etiology of the condition. In Dandy–Walker malformation, a condition in which there is partial or complete agenesis of the cerebellar vermis, the hydrocephalus is characterized by a disproportionate enlargement of the fourth and third ventricles (Osenbach and Menezes 1992). In hydrocephalus associated with spina bifida, the relationship is complex. However, there is substantial dysmorphology of the cerebellum with significant loss of Purkinje cells (Dennis et al. 2010). These lesions are not considered to be secondary to the associated hydrocephalic process.
Cerebellar mass lesions of vascular, neoplastic and infective nature frequently lead to hydrocephalus as a result of obstruction of the fourth ventricle. Whereas these causative relationships between hydrocephalus and the cerebellum have been well described and investigated, the impact of hydrocephalus on the structure and function of the cerebellum has not received equally prominent attention, until recently. This is surprising given the importance of the cerebellum in motor coordination, cognition and autonomic control.
The induction of hydrocephalus through intracisternal injection of kaolin has been used in several studies (Del Bigio and Zhang 1998; Olopade et al. 2012) and been found to be an effective method in experimental animals (Khan et al. 2006). It was observed that changes in ventricular size had begun in the early stage and continued until late stage. Kaolin injection into the cisterna magna of rats produced an obstruction of CSF outflow resulting in the hydrocephalic features such as enlarged dome shaped head, unsteady gait and occasionally, proptosis. We hypothesize that the initial reduction in body weight observed in hydrocephalic rats was caused by loss of appetite due to a rapid increase in intracranial pressure shortly after kaolin injection. However, the difference in weight between the two groups decreased over time, which coincided with the period of intracranial pressure equilibration reported by other studies (Williams et al. 2014). Gross cerebellar weight between the groups was comparable, but there was an increased cerebellar to body weight index in the hydrocephalic rats. This may be associated with the reduced general body weight, thus causing an apparent increase in the relative cerebellar weight.
In the hanging wire test, the reduction in the drop off time observed in the hydrocephalic rats when compared with the controls suggests that hydrocephalus resulted in muscle weakness, hypotonia being one of the features associated with cerebellar dysfunction. Negative geotaxis refers to an orienting response and movement expressed in opposition to cues of a gravitational vector (Fan et al. 2010) and thus tests motor coordination, a cerebellar function. The increased time taken by the hydrocephalic group for negative geotaxis test when compared to the control group, suggests that hydrocephalus is associated with deficits in motor coordination, balance, or vestibular input, all functions of the cerebellum.
The open field test provides simultaneous measures of locomotion, exploration, and general activity in rodents (Feyissa et al. 2017; Seibenhener and Wooten 2015). The significant decrease observed in behavioral patterns like line crossings and rearing, in the hydrocephalic group when compared to the control group, indicates some degree of motor deficits and anxiety, respectively. The motor deficits could be due to loss of Purkinje cells in many areas of the Purkinje layer and atrophy of Purkinje cells with hyperchromatic or pyknotic nuclei. Disturbance in the non-motor functions of the cerebellum (Strick et al. 2009) can also be deduced here, by the change in emotionality seen in the hydrocephalic rats (Timmann and Daum 2007). These are becoming increasingly important in recent times but were not explicitly assessed in this study.
Purkinje cells are the sole output neurons of the cerebellar cortex and play pivotal roles in coordination, control and learning of movements (Glickstein and Doron 2008). They are the most morphologically striking element in the cerebellar cortex.
Some studies have used parameters like ours to obtain pyknotic index in the hippocampus of hydrocephalic rats (Turgut et al. 2018; Taveira et al 2012). Watanabe et al. (2012) reported an increase in expression of cerebellar protein high-mobility group box-1 (HMGB-1), a protein secreted in response to cell damage, which was related to apoptosis of Purkinje cells in hydrocephalic H-Tx rats. The numbers of spared cells in the Purkinje layer were significantly less in the hydrocephalic group when compared with the control and seems to agree with an ongoing apoptotic process within the cerebellum.
The cerebellar sections in the hydrocephalic rats showed areas of vacuolations and pyknotic Purkinje cells. Vacuolations are spaces or vesicles within the cytoplasm of a cell, enclosed by a membrane and typically containing fluid. This finding was consistent with other studies (Martin et al. 1991), which reported similar damage seen in the Purkinje cell of the cerebellum in β-aminopropionitrile toxicity. Cells with dark nuclei, loss and abnormal clumping of heterochromatin were considered pyknotic. Dark shrunken neurons have been reported in light and electron microscopic studies on the cerebellar Purkinje layer of hydrocephalic rats (Uyanikgil et al. 2017).
Histological analysis of the cerebellum of hydrocephalic rats also revealed swollen Purkinje cells surrounded by empty spaces and disruption of the normal Purkinje cell architecture. Degenerating Purkinje cells appeared pale, swollen and vacuolated or as shrunken or hyper chromatic cells. Bent, elongated or folded nuclei were observed frequently in degenerating Purkinje cells. This finding was supported by Wallace et al. (2010) who reported decreased Purkinje cell density and size, due to prenatal Escherichia coli infection, was accompanied by impairments in motor coordination and balance in rats, and Tien and Ashdown (1992) suggested that this is probably related to behavioral manifestations of cerebellar alterations such as imbalance and ataxia in rats and humans. A similar loss of Purkinje cells and decrease in dendritic network of the Purkinje cell was reported by Oliveira et al. (2014) following chronic ethanol intake. These findings were consistent with findings made by Watanabe et al. (2013) on congenital hydrocephalic (HTx) rats. Purkinje cell defects in the cerebellum are also characterized by diminution of dendritic arborization and cell body size. The dendritic arborization of the Purkinje cell is critical to its functioning as Purkinje cells are at the center of the cerebellar neuronal circuit. Each Purkinje cell receives up to 200,000 synapses and transmits the integrated signal to the deep cerebellar nuclei with which it establishes functional synapses (Dusart and Flamant 2012). It is therefore pertinent to show that loss of the Purkinje cells or its dendritic arborization is fraught with problems in motor coordination, balance, muscle tone, as seen in hydrocephalus.
Conclusion
The results in this study showed that hydrocephalus caused morphological alterations and increased cell death in the Purkinje cells of the cerebellum within the hydrocephalic group when compared to control group, cerebellar dysfunctions in hydrocephalic juvenile rats have also been reported. Such alterations in Purkinje cell morphology suggest a parallel between hydrocephalus and the observed motor deficits in the condition.
Limitations of the study
This study was limited by the fact that we were unable to confirm ventricular enlargement before sacrifice of the rats due to unavailability of an MRI for animal studies in our institution. Therefore, we had to wait until sacrifice before we could detect and remove animals that did not develop any ventricular enlargement after kaolin injection.
References
D’Angelo E, Casali S (2012) Seeking a unified framework for cerebellar function and dysfunction: from circuit operations to cognition. Front Neural Circuits 6:116
Del Bigio MR, Wilson MF, Enno T (2002) Chronic hydrocephalus in rats and humans: white matter loss and behavior changes. Ann Neurol 53:337–346
Del Bigio MR, Zhang YW (1998) Cell death, axonal damage, and cell birth in the immature rat brain following induction of hydrocephalus. Exp Neurol 154:157–169
Del Bigio MR (2010) Neuropathology and structural changes in hydrocephalus. Dev Disabil Res Rev 16:16–22
Dennis M, Salman MS, Juranek JM (2010) Cerebellar motor function in spina bifida meningomyelocele. Cerebellum 9:484–498. https://doi.org/10.1007/s12311-010-0191-8
Di Curzio DL, Buist RJ, Del Bigio MR (2013) Reduced sub-ventricular zone proliferation and white matter damage in Juvenile Ferrets with Kaolin induced hydrocephalus. Exp Neurol 248:112–128
Di Curzio DL, Turner-Brannen E, Del Bigio MR (2014) Oral antioxidant therapy for juvenile rats with kaolin-induced hydrocephalus. Fluids Barriers CNS 11:23
Dusart I, Flamant F (2012) Profound morphological and functional changes of rodent Purkinje cells between the first and the second postnatal weeks: a metamorphosis? Front Neuroanat. https://doi.org/10.3389/fnana.2012.00011
Eccles JC, Ito M, Szentágothai J (1967) Purkinje cells and their responses. The cerebellum as a neuronal machine. Springer-Verlag, New York, pp 71–92
Fan LW, Tien LT, Zheng B, Pang Y, Rhodes PG, Cai Z (2010) Interleukin-1beta-induced brain injury and neurobehavioral dysfunctions in juvenile rats can be attenuated by alpha-phenyl-n-tert-butyl-nitrone. Neuroscience 168(1):240–252. https://doi.org/10.1016/j.neuroscience.2010.03.024
Feyissa DD, Aher YD, Engidawork E, Höger H, Lubec G, Korz V (2017) Individual differences in male rats in a behavioral test battery: a multivariate statistical approach. Front Behav Neurosci 11:26. https://doi.org/10.3389/fnbeh.2017.00026
Fukumizu M, Takashima S, Becker LE (1995) Neonatal posthemorrhagic hydrocephalus: neuropathologic and immunohistochemical studies. Pediatr Neurol 13(3):230–234
Glickstein M, Doron K (2008) Cerebellum: connections and functions. Cerebellum 7(4):589–594
Hale PM, McAllister JP II, Katz SD, Wright LC, Lovely TJ, Miller DW, Wolfson BJ, Salotto AG, Shroff DV (1992) Improvement of cortical morphology in infantile hydrocephalic animals after ventriculoperitoneal shunt placement. Neurosurgery 31(6):1085–1096
Herrup K, Wilczynski SL (1982) Cerebellar cell degeneration in the leaner mutant mouse. Neuroscience 7(9):2185–2196
Jansone B, Dzirkale Z, Jekabsons K, Pilipenko V, Beitnere U, Mâgure I, Skumbiòð R, Klçtnieks U, Vanaga I, Muceniece R, Kluða V (2016) Spruce needle polyprenols protect against atorvastatin-induced muscle weakness and do not influence central nervous system functions in rats. Proc Latvian Acad Sci Section B 70(1):13–20
Jones HC, Bucknall RM, Harris NG (1991) The cerebral cortex in congenital hydrocephalus in the H-Tx rat: a quantitative light microscopic study. Acta Neuropathol 82(3):217–224. https://doi.org/10.1007/BF00294448
Khan OH, Enno TL, Del Bigio MR (2006) Brain damage in neonatal rats following kaolin induction of hydrocephalus. Exp Neurol 200(2):311–320
Lubics A, Reglodi D, Tamás A, Kiss P, Szalai M, Szalontay L, Lengvári I (2005) Neurological reflexes and early motor behavior in rats subjected to neonatal hypoxic-ischemic injury. Behav Brain Res 157(1):157–165
Martin JE, Sosa-Melgarejo JA, Swash M, Mather K, Leigh PN, Berry CL (1991) Purkinje cell toxicity of β-aminopropionitrile in the rat. Virchows Archiv A 419:403–408
Oliveira SA, Chuffa LG, Fioruci-Fontanelli BA et al (2014) Apoptosis of Purkinje and granular cells of the cerebellum following chronic ethanol intake. Cerebellum 13(6):728–738. https://doi.org/10.1007/s12311-014-0591-2
Olopade FE, Shokunbi MT, Sirén A (2012) The relationship between ventricular dilatation neuropathological and neurobehavioural changes in hydrocephalic rats. Fluids Barriers CNS 9:19
Osenbach RK, Menezes AH (1992) Diagnosis and management of the Dandy-Walker malformation: 30 years of experience. Pediatr Neurosurg 18:179–189. https://doi.org/10.1159/000120660
Palay SL, Chan-Palay V (1974) The Purkinje cell. Cerebellar cortex: cytology and organization. Springer-Verlag, Berlin, pp 11–62
Rodríguez-Fanjul J, Durán Fernández-Feijóo C, Lopez-Abad M, Lopez Ramos MG, Balada Caballé R, Alcántara-Horillo S, Camprubí Camprubí M (2017) Neuroprotection with hypothermia and allopurinol in an animal model of hypoxic-ischemic injury: Is it a gender question? PLoS ONE 12(9):e0184643. https://doi.org/10.1371/journal.pone.0184643
Rubin RC, Hochwald G, Liwnicz B, Tiell M, Mizutani H, Shulman K (1972) The effect of severe hydrocephalus on size and number of brain cells. Dev Med Child Neurol Suppl 27:117–120. https://doi.org/10.1111/j.1469-8749.1972.tb09783.x
Seibenhener ML, Wooten MC (2015) Use of the Open Field Maze to measure locomotor and anxiety-like behavior in mice. J Vis Exp 96:e52434. https://doi.org/10.3791/52434
Singh R, Prasad K, Kumar A, Misra A, Padmakumar K, Malaviya A (1988) Cerebellar ataxia in systemic lupus erythematosus: three case reports. Ann Rheum Dis 47(11):954–956
Stanford SC (2007) The Open Field Test: reinventing the wheel. J Psychopharmacol 21(2):134–135. https://doi.org/10.1177/0269881107073199
Strick PL, Dum RP, Fiez JA (2009) Cerebellum and non-motor function. Annu Rev Neurosci 32:413–434
Taveira KV, Carraro KT, Catalão CH, Lopes LS (2012) Morphological and morphometric analysis of the hippocampus in Wistar rats with experimental hydrocephalus. Pediatr Neurosurg 48(3):163–167. https://doi.org/10.1159/000345959
Tien RD, Ashdown BC (1992) Crossed cerebellar diaschisis and crossed cerebellar atrophy: correlation of MR findings, clinical symptoms, and supratentorial diseases in 26 patients. AJR Am J Roentgenol 158(5):1155–1159
Timmann D, Daum I (2007) Cerebellar contributions to cognitive functions: a progress report after two decades of research. Cerebellum 6(3):159–162. https://doi.org/10.1080/14734220701496448
Turgut M, Baka M, Uyanıkgil Y (2018) Melatonin attenuates histopathological changes in the hippocampus of infantile rats with kaolin-induced hydrocephalus. Pediatr Neurosurg 53(4):229–237
Uyanıkgil Y, Turgut M, Baka M (2017) Effects of melatonin on the cerebellum of infant rat following kaolin-induced hydrocephalus: a histochemical and immunohistochemical study. Cerebellum 16:142–150
Wallace K, Veerisetty S, Paul I, May W, Miguel-Hidalgo JJ, Bennett W (2010) Prenatal infection decreases calbindin, decreases Purkinje cell volume and density and produces long-term motor deficits in Sprague-Dawley rats. Dev Neurosci 32(4):302–312
Watanabe M, Miyajima M, Nakajima M et al (2012) Expression analysis of high mobility group box-1protein (HMGB-1) in the cerebral cortex, hippocampus, and cerebellum of the congenital hydrocephalus (H-Tx) rat. Acta Neurochir Suppl 113:91–96
Watanabe M, Miyajima M, Ogino I, Nakajima M, Arai H (2013) Cerebellar Purkinje cells exhibit increased expression of HMGB-1 and apoptosis in congenital hydrocephalic H-Tx rats. Neurosurgery 72(3):459–467. https://doi.org/10.1227/NEU.0b013e31827fcd83
Welsh JP, Yuen G, Placantonakis DG, Vu TQ, Haiss F, O'Hearn E, Molliver ME, Aicher SA (2002) Why do Purkinje cells die so easily after global brain ischemia? Aldolase C, EAAT4, and the cerebellar contribution to posthypoxic myoclonus. Adv Neurol 89:331–359
Williams MT, Braun AA, Amos-Kroohs RM, McAllister JP et al (2014) Kaolin-induced ventriculomegaly at weaning produces long term learning, memory and motor deficits in rats. Int J Dev Neurosci 35:7–15. https://doi.org/10.1016/j.ijdevneu.2014.02.002
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
The authors declare that they have no conflict of interest.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
About this article

Cite this article
Olopade, F.E., Femi-Akinlosotu, O., Adekanmbi, A.J. et al. Neurobehavioural changes and morphological study of cerebellar purkinje cells in kaolin induced hydrocephalus. Anat Sci Int 96, 87–96 (2021). https://doi.org/10.1007/s12565-020-00561-z
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s12565-020-00561-z