
Abstract
Organotin compounds (OTCs) have contaminated marine environments and are known to induce imposex in many marine female snails. To evaluate the bioaccumulation of environmental OTCs in rock shell Thais clavigera we translocated some from a non-polluted area to a polluted area in Kagoshima Bay, Japan, and held them with prey (P) or without prey (NP). After 13 weeks’ exposure, the proportion of animals exhibiting imposex and their relative penis length (RPL) index in the P group were 71 and 15.1 %, respectively. Due to high mortality, keeping of the NP group was terminated after 8 weeks; the proportion of them exhibiting imposex was 0 %. While the tributyltin (TBT) concentration was 1.6 ng/g (wet weight; w.w.) at the beginning of the study and increased to 178 ng/g w.w. by week 8 in the NP group, it increased by 483 and 424 ng/g w.w. by weeks 6 and 13 in the P group, and almost reached a steady state in the P group until week 6. Furthermore, the accumulation of TBT was higher in the P group than in the NP group, suggesting that the potential for imposex induction in rock shells in the sites of Kagoshima Bay is still high due to higher OTC concentrations in their prey organisms.
Similar content being viewed by others

Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
Organotin compounds (OTCs), the active biocides found in anti-fouling paints, are a class of environmental contaminants [1, 2] that cause shell thickening in oysters [3] and imposex (penis induction) in some female snails [4–6]. While OTCs have been banned from use in anti-fouling paints, some sites remain contaminated with relatively high concentrations [6–10].
There are two routes by which aquatic organisms accumulate chemicals from the environment. The first, bioconcentration, involves uptake from water through the gills and the second, biomagnification, involves uptake through the food chain. Research suggests that OTCs are transferred through the food chain [11–14]. In our previous study, the results suggested that the rock shell, Thais clavigera, assimilates OTCs from the sediment and suspended particles as most of their prey are filter or suspension feeders in Kagoshima Bay, Japan. Currently the OTCs levels in bivalves, snails and sediment are relatively high.
To evaluate OTCs pollution and the potential for imposex induction in T. clavigera, we translocated some of them from a non-polluted area to a polluted area and held them with prey (P) or without prey (NP) in Kagoshima Bay. We also evaluated the contributions of OTCs in the water and their prey organisms to those of rock shells.
Materials and methods
Sample collection and experimental design
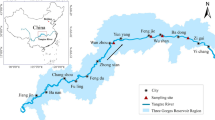
Rock shells were collected at Iso, a non-polluted site, and translocated to Akobaru where almost all female rock shells are still showing imposex from our annual observation of a polluted site in Kagoshima Bay (Figs. 1, 2) The rock shells were divided into two groups. One group (P) of 20 rock shells was placed in a mesh cage (33 × 24 × 7 cm; mesh size 2 mm) with prey organisms such as mussels, Septifer virgatus, and oyster, Saccostrea mordax, which we collected at Akobaru. The second group (NP) of 20 rock shells was placed inside a net (mesh size 1 mm) to isolate them from their prey and this net was placed in the same cage as the first group (Fig. 3). Five cages (40 individuals/cage) were placed in Akobaru in July 2006. Rock shells from both groups (P and NP) were collected from each cage in weeks 1, 2, 4 and 6. Thereafter, individuals from the NP group were sampled in week 8 and individuals from the P group in week 13, as shown in Table 1. We removed all individuals in the NP group at week 8 because of high mortality due to starvation. First, we measured the shell size and total weight of each individual, and then dissected them to examine them for imposex (induction of penis in females). After measuring the weight of the rock shells’ flesh, their condition factor, weight of flesh/(shell height)3 × 1000, was calculated. The flesh was removed and stored at −10 °C in the freezer until analysis. To determine the extent of imposex induction, we calculated a relative penis length (RPL) index as follows [5, 15]:

Study area and sampling sites

Induction of imposex in rock shells in Kagoshima Bay

Schematic of experimental design of the translocation experiment. Illustration of the cage containing rock shells that had access to prey (P) or did not have access to prey (NP)
Analysis of OTCs
We measured the concentrations of the following OTCs in water samples and in rock shell samples collected in Akobaru: monobutyltin (MBT), dibutyltin (DBT), tributyltin (TBT), monophenyltin (MPT), diphenyltin (DPT), and triphenyltin (TPT). The samples were analyzed following the method described in Uno et al. [16]. Briefly, we added surrogate OTCs (MBT-d 9, DBT-d 18, TBT-d 27, MPT-d 5, DPT-d 10, and TPT-d 15), acetic acid-sodium acetate trihydrate buffer (pH 5), and sodium tetraethylborate (NaBEt4) to the water samples then extracted OTCs using n-hexane. The soft tissue samples were thawed then spiked with OTC surrogates in a similar manner. The spiked samples were homogenized then extracted with methanol-ethyl acetate (1:1, volume/volume) containing 1 M hydrobromic acid for 30 min by ultrasonication followed by centrifugation. After dehydrating the extract with anhydrous sodium sulfate, the OTCs were ethylated using a NaBEt4 solution. The lipids were saponified with an ethanol solution containing 1 M potassium hydroxide. OTCs were extracted twice with n-hexane. After concentrating the hexane solutions derived from the water and tissue samples, they were purified using a hexane-washed florisil column. The OTCs eluted by 5 % diethyl ether-hexane were analyzed by gas chromatography–mass spectroscopy (Agilent Technologies 6890 Series gas chromatograph equipped with a 5973 MSD mass selective detector). The detection limits of OTCs were 0.1 ng/l and 0.1 ng/g w.w., respectively. The recovery rates were 83.3 % for TBT, 71.2 % for DBT, 66.5 % for MBT, 88.4 % for TPT and 102.6 % for DPT in aquatic organisms and we did not correct our analysis results by these recovery rates.
Statistical analysis
The data were analyzed using a one-way ANOVA followed by Dunnett’s test and a t-test. All statistical analyses were performed using SPSS 10.0J (SPSS, Tokyo). The level of significance was P < 0.05.
Results
Condition of translocated rock shell
The water temperature in Kagoshima Bay during our study period ranged from 23.5 to 30.2 C°. The shell height and total body weight were 22.8 ± 1.4 mm and 2.19 ± 0.3 g at the beginning (n = 30, mean ± SD), 23.3 ± 1.0 mm and 2.23 ± 0.29 g at week 8 in the NP group (n = 9) and 22.7 ± 1.1 mm and 2.58 ± 0.32 g at week 13 in the P group (n = 13), respectively. There were significant differences in the condition factor after week 2, but not in shell height and total body weight of rock shell individuals between the P and NP groups (Fig. 4). Survival was 50 and 45 % in the cages collected at week 6 and 8, respectively in the NP group. Conversely, survival was 95 % and 65 % at weeks 6 and 13 in the P group (Fig. 5).

Changes in the condition factor of translocated rock shells (mean ± SD). For abbreviations, see Fig. 3. ** Significantly different from P group (<0.01)

Survival of translocated rock shells. For abbreviations, see Fig. 3
Induction of imposex
Induction of imposex was observed at week 13 in the P group. A high proportion (71 %) of female rock shells in the P group (five of seven individuals) exhibited imposex (Fig. 6) and the RPL index of all female individuals was 15.1 % (26.3 % in imposex females).

Proportion of individuals exhibiting imposex induction, and relative penis length (RPL) index of translocated rock shells. For other abbreviations, see Fig. 3
OTC concentrations in translocated rock shells
The OTC concentrations are given in Table 1 for translocated rock shells and Table 2 for water, sediment, prey and wild rock shells. The ΣOTCs concentrations were higher in the P group than the NP group or the prey (Table 1). While the ratios of TBT in butyltins (sum of MBT, DBT and TBT) were around 50 % in the P group in week 13 and NP group in week 8, those of TPT in phenyltins (sum of MPT, DPT and TPT) were 56 % for the P group at week 13 and 22 % for the NP group at week 8, respectively. Similarly, the TBT concentrations were higher in individuals from the P group than the NP group. The TPT concentrations were not much different between the P group and NP group (Fig. 7). The tissue concentration of butyltins was higher than that of phenyltins.

Tributyltin (TBT) and triphenyltin (TPT) concentrations in translocated rock shells (mean ± SD). SD bars of TBT and TPT concentrations in the NP group were too small to show. For other abbreviations, see Fig. 3
The TBT concentration was 1.6 ng/g w.w. [3.9 ng/g dry weight (d.w.)] at the beginning and had increased by 483 and 424 ng/g w.w. (1210 and 1311 ng/g d.w.) by weeks 6 and 13 in the P group and 68.3 and 178 ng/g w.w. (206 and 548 ng/g d.w.) at weeks 6 and 8 in the NP group, respectively. The TPT concentrations was 3.9 ng/g w.w. (9.8 ng/g d.w.) at the beginning and had increased by 61.4 and 110 ng/g w.w. (154 and 339 ng/g d.w.) by weeks 6 and 13 in the P group and 21.3 and 47.0 ng/g w.w. (64.6 and 144 ng/g d.w.) by weeks 6 and 8 in the NP group, respectively. The TBT concentration almost reached steady state in the P group until week 6.
Discussion
The OTC concentrations found in the sediments, wild rock shells and prey organisms that were analyzed in 2005 but seemed not to have changed much by 2006, were higher in Akobaru than in Iso (Table 2). Conversely, the OTC concentrations in the water in 2006 had decreased relative to those in 2005 reported by Koyama et al. [6]. However, almost all female rock shells still exhibited imposex, as found in our annual survey at Akobaru (Fig. 2). The OTCs seem to have been released from a small shipyard and fishing boats in the past. Taken together, these observations suggest that Akobaru has been contaminated by OTCs and that the residue levels are high enough to induce imposex in female rock shell even presently. Interestingly, the initial tissue TBT and TPT concentrations were 1.6 and 3.9 ng/g w.w. (3.9 and 9.8 ng/g d.w.), thus, lower than levels reported from other sites: 4.3–16.7 ng Sn/g d.w. (10.5–40.8 ng TBT/g d.w., [17]), 18.0–400 ng TBT/g w.w., and 36.9–782 ng TPT/g w.w. [18], suggesting that the rock shells collected from Iso were less exposed to OTCs.
According to Shim et al. [19], TBT and TPT concentrations of translocated rock shells without prey in south Korea, whose RPL index increased from 36 to 74 %, reached a steady state on days 17 and 32 and increased from 179 to 436 ng TBT/g d.w. (66–160 ng TBT/g w.w., estimation by our w.w./d.w. ratio) and 47–218 ng TPT/g d.w. (17–80 ng TPT/g w.w.), respectively. While the TBT concentration in the waters of Akobaru was almost 1/10 of that reported by Shim et al. [19], TBT and TPT concentrations of the translocated rock shells of the NP group were 68.3 and 21.3 ng/g w.w. at week 6 and 178 and 47.0 ng/g w.w. at week 8, respectively, in the present study. These OTCs levels are similar to those reported by Shim et al. [19]. The estimated bioaccumulation factor (OTCshell/OTCwater) of TBT in rock shells (w.w. basis) in the present study, 1.48 × 105 (178/0.0012), was higher than 1.29 × 104 (168/0.013) estimated from Shim et al. [19]. The bioconcentration factor of TBT can increase depending on the reduction of the TBT concentration in marine gastropods [20] and fishes [21]. The efficiency of bioconcentration by the translocated rock shells also seemed to be relatively effective in lowering the TBT concentration in the water of Akobaru in the present study.
The TBT concentrations of the P group at weeks 6 and 13 (Table 1) were significantly higher than those of wild rock shells in Akobaru (Table 2). Rock shells prey on a variety of organisms such as barnacles, limpets and bivalves [22]. As reported by Tang et al. [23], TBT was also bioaccumulated by oysters and mussels. The OTCs accumulated in these prey organisms appear to be transferred to rock shells as well as the OTCs present in the water. Therefore, the translocated rock shells were caged with mussels and oysters in the present study. On the other hand, most wild rock shells distributed near a mussel community seemed to prey on mussels in our annual observation at Akobaru. As shown in Table 2, the OTC concentrations in oyster were much higher than those of mussels. These results suggest that the translocated rock shells preyed not only on mussels but also on oysters and, therefore, accumulated higher OTCs than wild ones.
Based on Horiguchi et al. [24] and Shim et al. [19], a RPL index >80 % was observed in wild rock shells which accumulated approximately 200 ng TBT/g w.w. and 200 ng TPT/g w.w. (estimation by our w.w./d.w. ratio). As shown in TBT and TPT accumulation profiles in Fig. 7, the TBT concentration in the translocated rock shells almost reached a steady state within 6 weeks. Their mean concentrations of TBT and TPT, 424 and 110 ng/g w.w. in the P group, were sufficient to induce imposex at week 13, suggesting that OTCs in water and prey at Akobaru were still at sufficiently toxic levels to induce imposex in rock shells in 2006 and until the present.
Although OTC concentrations in translocated rock shells of the P group were sufficient to induce imposex in the current study, their RPL index was 15.1 % (26.3 % in imposex females). According to the results of Shim et al. [19], the RPL index increased from 36 to 74 % in translocated rock shells without prey for 55 days. While the TBT and TPT concentrations of translocated rock shells of P group in the present study, i.e., 424 and 110 ng/g w.w., were much higher than those reported by Shim et al. [19], their RPL index, i.e., 15.1 % (26.3 % in imposex females), was smaller than those reported by Shim et al. [19].; however, the increase in the RPL index, 38 % (36–74 %) in their study was not so very different from our results. It took 13 weeks to induce imposex in the present study; however, it took only 32 days to induce a significantly longer mean female penis in translocated rock shells [19]. It possibly takes longer to induce imposex in non-imposex rock shells.
While TBT and TPT concentrations of translocated rock shells in the NP group in the present study were similar to those reported by Shim et al. [19], they did not induce imposex. This suggests that longer starvation suppressed the induction of imposex in non-imposex rock shells in NP group.
The concentration of TBT was higher in the P group than in the NP group suggesting that the potential of imposex induction in the sites of Kagoshima Bay is still high due to higher OTC concentrations in prey organisms. TBT can be accumulated from both water and prey bivalves [25] and marine snails [14] through the food web. Based on the lower OTC concentrations in the water, but higher concentrations in prey and sediment in Akobaru, OTCs in prey seemed to be one of the important sources of OTCs for rock shells in Akobaru. After the banning of OTCs, the sediment TBT concentration did not change significantly in south Korea [10]. The sediment TBT concentration of 123 ng/g d.w. in Akobaru in 2005 was medium level when compared with that of South Korea. This suggests that recovery from sediment pollution by TBT is difficult and that sediment can be one of the sources of TBT for coastal biota by desorption or resuspension of sediment-bound TBT.
In conclusion, OTCs levels in wild and translocated rock shells have been sufficient to induce imposex, suggesting that OTCs levels in the environment of Kagoshima Bay are also sufficient to induce imposex in rock shells there.
References
Yamada H, Takayanagi K, Tateishi M, Tagata H, Ikeda K (1997) Organotin compounds and polychlorinated biphenyls of livers in squid collected from coastal waters and open oceans. Environ Pollut 96:217–226
Ueno D, Inoue S, Takahashi S, Ikeda K, Tanaka H, Subramanian AN, Fillmann G, Lam PKS, Zheng J, Muchtar M, Prudente M, Chung K, Tanabe S (2004) Global pollution monitoring of butyltin compounds using skipjack tuna as a bioindicator. Environ Pollut 127:1–12
Alzieu CI, Sanjuan J, Michel P, Borel M, Dreno JP (1989) Monitoring and assessment of butyltins in Atlantic coastal waters. Mar Pollut Bull 20:22–26
Smith BS (1981) Tributyltin compounds induce male characteristics on female mud snails Nassarius obsoletus = Ilyanassa obsolete. J Appl Toxicol 1:141–144
Horiguchi T, Shiraishi H, Shimizu M, Morita M (1994) Imposex and organotin compounds in Thais clavigera and T. bronni in Japan. J Mar Biol Assoc UK 74:651–669
Koyama J, Takenouchi A, Nigaya S, Kokushi E, Uno S (2012) Organotin residues in rock shell, Thais clavigera, and their sources in Kagoshima Bay, Japan. Aquat Ecosyst Health 15:260–266
Inoue S, Abe S, Oshima Y, Kai N, Honjo T (2006) Tributyltin contamination of bivalves in coastal areas around northern Kyushu, Japan. Environ Toxicol 21:244–249
Berto D, Giani M, Boscolo R, Covelli S, Giovanardi O, Massironi M, Grassia L (2007) Organotins (TBT and DBT) in water, sediments and gastropods of southern Venice lagoon (Italy). Mar Pollut Bull 55:425–435
Lopes-dos-Santos RMA, Galante-Oliveira S, Lopes E, Almedia C, Barroso C (2014) Assessment of imposex and butyltin concentration in Gemophos viverratus (Kiener, 1834), from São Vicente, Republic of Cabo Verde (Africa). Environ Sci Pollut Res 21:10671–10677
Kim NS, Hong SH, Yim UH, Shin KH, Shim WJ (2014) Temporal changes in TBT pollution in water, sediment and oyster from Jinhae Bay after the total ban in South Korea. Mar Pollut Bull 86:547–554
Coelho MR, Bebianno MJ, Langston WJ (2002) Routes of TBT uptake in the clam Ruditapes decussatus. II. Food as vector of TBT uptake. Mar Environ Res 54:193–207
Kono K, Minami T, Yamada H, Tanaka H, Koyama J (2008) Bioaccumulation of tributyltin and triphenyltin compounds through the food web in deep offshore water. Coast Mater Sci 32:102–107
Murai R, Sugimoto A, Tanabe S, Takenouchi I (2008) Biomagnification profiles of tributyltin (TBT) and triphenyltin (TPT) in Japanese coastal food webs elucidated by stable nitrogen isotope ratios. Chemosphere 73:1749–1756
Bryan GW, Bright DA, Hummerstone LG, Burt GR (1993) Uptake, tissue distribution and metabolism of C14-labelled tributyltin (TBT) in the dog-whelk, Nucella lapillus. J Mar Biol Assoc UK 73:889–912
Gibbs PE, Bryan GW, Pascoe PL, Burt GR (1987) The use of the dog-whelk, Nucella lapillus, caused by imposex induced by tributyltin from antifoulding paints. J Mar Biol Assoc UK 66:767–777
Uno S, Murakami M, Kokushi E, Koyama J (2011) Interspecies differences in the accumulation of tributyltin and its metabolites under dietary exposure in sea perch, Lateolabrax japonicas and red sea bream, Pagrus major. Environ Toxicol 26(1):29–36
Leung KMY, Kwong RPY, Ng WC, Horiguchi T, Qiu JW, Yang R, Song M, Jiang G, Zheng GJ, Lam PKS (2006) Ecological risk assessments of endocrine disrupting organotins compounds using marine neogastropods in Hong Kong. Chemosphere 65:922–938
Horiguchi T, Shiraishi H, Shimizu M, Morita M (1997) Imposex in sea snails, caused by organotin (tributyltin and triphenyltin) pollution in Japan: a survey. Appl Organomet Chem 11:451–455
Shim WJ, Kahng SH, Hong SH, Kim NS, Kim SK, Shim JH (2000) Imposex in the rock shell, Thais clavigera, as evidence of organotins contamination in the marine environment of Korea. Mar Environ Res 49:435–451
Oehlmann J, Stoben E, Schulte-Oehlmann U, Bauer B (1998) Imposex development in response to TBT pollution in Hinia incrassate (Ström, 1768) (Prosobranchia, Stenoglossa). Aquat Toxicol 43:239–260
Yamada H, Takayanagi K (1992) Bioconcentration and elimination of bis (tributyltin) oxide (TBTO) and triphenyltin chloride (TPTC) in several marine fish species. Water Res 26:1589–1595
Blackmore G (2000) Field evidence of metal transfer from invertebrate prey to an intertidal predator, Thais clavigera (Gastropoda: Muricidae). Estuar Coast Shelf Sci 51:127–139
Tang C-H, Hsu C-H, Wang W-H (2010) Butyltin accumulation in marine bivalves under field conditions in Taiwan. Mar Environ Res 70:125–132
Horiguchi H, Shiraishi H, Shimizu M, Yamazaki S, Morita M (1995) Imposex in Japanese gastropods (Neogastropoda and Mesogastropoda): effects of tributyltin and triphenyltin from antifouling paints. Mar Pollut Bull 31:402–405
Shim WJ, Yim UK, Kim NS, Hong SH, Oh JR, Jeon JK, Okamura H (2005) Accumulation of butyl -and phenyltin compounds in starfish and bivalves from the coastal environment of Korea. Environ Pollut 133:489–499
Acknowledgments
The authors would like to acknowledge Kagoshima University for financial support for this study. We also thank Prof. Miguel Vazquez Archdale (Kagoshima University) for his improvements to and comments on this manuscript.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article

Cite this article
Koyama, J., Nigaya, S., Takenouchi, A. et al. Bioaccumulation of environmental organotin compounds in translocated rock shell Thais clavigera in Kagoshima Bay. Fish Sci 82, 975–982 (2016). https://doi.org/10.1007/s12562-016-1031-7
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s12562-016-1031-7