
Abstract
Several fossil beetles from the Late Pliocene (Piacenzian) of Willershausen (Lower Saxony, Germany) previously identified as Tenebrionidae and Staphylinidae are reviewed. One species, formerly determined as “Staphylinoidea” is identified as Euboeus mimonti Boieldieu, 1865 (Tenebrionidae: Helopini). This species is the oldest record of an extant species of darkling beetles, which occurs now in the Balkans and Anatolia. The discovery of Euboeus mimonti in the Late Pliocene of Germany indicates that the range of this species was much wider, and the climate in the Late Pliocene in the modern territory of Lower Saxony was much milder. One species formerly determined as “Alleculidae gen. sp.” belongs to the subfamily Alleculinae (Tenebrionidae) and it is identified here as Pseudocistela aff. ceramboides. Two species, misidentified by a previous author as Tenebrionidae belong to Elateridae and Cerambycidae respectively. Generic and species composition, as well as the ratio of extinct and extant beetle taxa in the Willershausen Fossil Lagerstätte are discussed.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Darkling beetles are poorly presented in fossil deposits from the Late Jurassic to the Pliocene but they are well represented in Cenozoic ambers (Nabozhenko 2019). The fossil record contains 133 fossil species from 86 genera (Nabozhenko 2019; Nabozhenko and Tanaka 2022, 2023; Nabozhenko and Perkovsky 2023). One species was recently transferred to Zopheridae (Alekseev and Nabozhenko 2023) and another one to the family Tetratomidae (Aalbu et al. 2023).
Only two recently determined species of Lagria Fabricius, 1775 (Tenebrionidae: Lagriinae) are known from Pliocene deposits: Lagria koshimizui Nabozhenko et Tanaka, 2023 and Lagria cf. formosensis Borchmann, 1912 from the Kabutoiwa Member of the Motojuku Formation (lower Piacenzian) in Japan (Nabozhenko and Tanaka 2023). While Paleogene and older records of darkling beetles are primarily important for understanding the evolutionary history of the family, evolutionary scenarios, and phylogenetic models, Neogene (especially Pliocene) records provide valuable data for palaeoclimatic and faunistic reconstructions. For example, the record of thermophilic fossil Lagria spp. and other subtropical insects on the main islands of Japan made it possible to refute the opinion of a cold-temperate climate in this region in the Late Pliocene based on the descriptions of the frog genus Rana Linnaeus, 1758 (Nabozhenko and Tanaka 2023).
Below we present an interesting compression print of darkling beetle of the tribe Helopini (subfamily Tenebrioninae) from the Willershausen Fossil Lagerstätte, Germany (Pliocene, Piacenzian), which was originally assigned to Staphylinoidea (Gersdorf 1969). Three specimens from this deposit that were previously determined as tenebrionids (Gersdorf 1969, 1971, 1976) are also discussed.
Material and methods
The studied specimens are deposited in the Geoscience Museum at the Georg-August University in Göttingen and bear collection numbers prefixed by GZG.W. (Geowissenschaftliches Zentrum Göttingen, Fossillagerstätte Willershausen), and associated original numbers that were assigned for the publication of specimens in the 1970s and 1980s.
The extant specimens of Euboeus mimonti are deposited in the Zoological Institute of the Russian Academy of Sciences (ZIN, St Petersburg, Russia).
Fossil specimens were photographed with a Sony Alpha 99 II camera equipped with a Tamron SP 90mm macro lens. Line drawings were hand-drawn and hand stroked through transmitted light and enhanced with Photoshop (Adobe Systems software) from the photographs and checked against the specimens. Extant specimens were studied using a binocular microscope Micromed MC-4 Zoom Led and their photographs were taken with a Canon EOS 5D Mark IV Body, Canon MP-E65MM F2.8 Macro lens and Canon Macro Twin Lite MT-26X-RT flash bulb, and stacking was done using Stack-shot 3X with enlarged macro rails s/n 3734; the photosystem is installed on a Kaiser Copy Stand RS 1 reproduction machine. Images were stacked in Helicon Focus 7.7.4 Pro.
Geological setting and locality
The former clay-pit is located in the township Willershausen (51.783724 N, 10.113421 E), ca. 30 km north of Göttingen, Lower Saxony, Germany. From the 16th century to its closure in 1975, the pit exploited clay for nearby brickworks in the last decades of its existence (Ziegelei Schlange). Since 1977 the abandoned pit is a protected Natural Monument (Meischner, 2000), and it was included in the Geopark “Harz, Braunschweiger Land, Ostfalen” in 2012. The Lagerstätte formed in a sinkhole in tectonically disturbed Lower (Buntsandstein) and Middle (Muschelkalk) Triassic sediments above leached late Permian evaporites (Zechstein) (Meischner 2000). A lake infilling in the sinkhole was 150–200 m in diameter, steep-sided and several tens of meters deep, with a well oxygenated upper water body (epilimnion) and anoxic conditions in lower water column (hypolimnion). Principle lacustrine lithologies comprise a lake margin facies of sand and light-coloured clay with few and poorly preserved fossils (mainly plant detritus, gastropods, bivalves, crayfish exuviae) and a central lake facies of dark clay and carbonates with common and well-preserved fossil content. Almost all of the ~50.000 fossils recovered from Willershausen come from a 30 cm thick, laminated bed of lime marl and dolomitic marlstone of the central lake facies (Meischner 2000). Based on the find of Anancus arvernernsis, Wegele (1914) proposed a Late Pliocene age for the Willershausen biota. Biostratigraphic data from palynomorphs, macroflora and vertebrates (Mohr 1986; Mai 1995; Knobloch 1998) support a Late Pliocene (Piacenzian, 3.6 – 2.588 Mya) age of the Fossil Lagerstätte Willershausen.
Brief information about fossil Coleoptera of Willershausen
The most recent informative bibliographic reviews of the fossil fauna and flora of this locality have been presented by Ferguson and Knobloch (1998) and Kolibáč et al. (2016). Here we focus on the coleopterofauna of the Fossil Lagerstätte Willershausen. Korge (1967) described the first clear genus and a species of Staphylinidae (Table 1) from Willershausen, whose status as a palaeoendemic genus is incorrect and will be dealt with separately (Jenkins Shaw et al. in prep). Schmidt (1967) reviewed the fossil Cerambycidae from the Willershausen deposits, describing two new genera and three new species, one of those genera was later assigned to an extant Eastern Asian species (Vitali 2011). An additional specimen of Acanthocinus schmidti Schmidt, 1967 that was misidentified and described by Jordan (1967) as Pseudonepa bitarsalis Jordan, 1967 (Heteroptera) was identified by Gersdorf (1976), who also gave the most comprehensive information about fossil beetles from Willershausen (Gersdorf 1969, 1971, 1976). In the latter three articles, he described the prints of 120 beetles belonging to 16 families. Only 21 taxa of these beetles were identified to genus and/or species, including three new extinct genera and seven new species. Much later, Schweigert (2003) determined one species of longhorn beetle (Cerambycidae) and one stag beetle (Lucanidae) from the Pliocene of Willershausen. Brauckmann et al. (2013) revised the identification of Oryctes nasicornis (Linnaeus, 1758) of Gersdorf (1971) as Oryctes? sp. (Scarabaeidae: Dynastinae). Finally, a new extinct species of Trogossitidae was described by Kolibáč et al. (2016).
Systematic palaeontology
Family Tenebrionidae Latreille, 1802
Subfamily Tenebrioninae Latreille, 1802
Tribe Helopini Lacordaire, 1859
Subtribe Helopina Lacordaire, 1859
Genus Euboeus Boieldieu, 1865
Euboeus mimonti Boieldieu, 1865

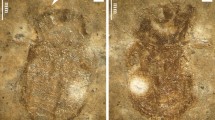
Euboeus mimonti, fossil (GZG) and extant (ZIN) specimens: a print GZG.W.16854. Original number: 599–60, side lighting; b the same, direct lighting and scale; c reconstruction; d extant male (Greece, Peloponnese); e the same, female

Euboeus mimonti, fossil (GZG) and extant (ZIN) specimens, head and pronotum: a print GZG.W.16854. Original number: 599–60; b extant specimen, male
Gersdorf, 1969: 303: Staphylinoidea
Material: One print, sex unknown: GZG.W.16854. Original number: 599–60.
Description of compression print: The body has been strongly deformed during the fossilization, black with reddish inlays at base of head, across the pronotum and partly elytra. Only several right antennomeres and right leg without tarsus are partly preserved. Coarse puncturation of round, dense punctures is well visible on the head and the pronotum. The pronotum is partly deformed: the right margin with posterior angle is normal, while the left margin was tucked in when fossilised (Figs 1c, 2a). The scutellar shield is absent. Left elytron is damaged at the basal third and the apical quarter, whereas the right elytron is partly damaged only at apical portion, but its apex is visible. Widely rounded humeri are well visible. The long striolas are well presented, as well as seven visible elytral striae.
Comments: This specimen was originally assigned to “Staphylinoidea” (Gersdorf 1969) on the base of supposedly shortened elytra. Gersdorf (1969) assumed that this species may belong to any staphylinid genus with shortened, but still relatively long elytra, for example, Ptomascopus Kraatz, 1876 (Staphylinidae: Silphinae), Velleius Leach, 1819 (Staphylinidae: Staphylininae) or Megarthrus Curtis, 1829 (Staphylinidae: Proteininae). Gersdorf (1969) misinterpreted some structures of this print, especially the elytra, which are not shortened or reduced.
Despite the absence of some structures necessary for diagnostics, we can with some confidence attribute this specimen to Euboeus mimonti based on the combination of the following features, which are not characteristic of beetles from other families and other groups of Tenebrionidae: eyes reniform, with ocular canthus; anterior margin of epistoma widely emarginated (Fig. 2, compare with Fig. 2b); lobe-shaped genae, covering the base of the antennae dorsally; moniliform antenna; pronotum and head coarsely punctured by round punctures; pronotum bell-shaped, widest at base (Figs 1c, 2); elytral base much wider than the pronotal one; humeral angles widely rounded, not projected; clear long scutellary striole; at least seven elytral striae well visible as in extant specimens of Euboeus mimonti; walking legs; print length 18 mm (result of fossilization; body length was shorter but not much).
At first glance, this beetle resembles the large European comb-clawed beetle Upinella aterrima (Rosenhauer, 1987) (see images and description in Novák (2015), which has a similar habitus. This impression is especially reinforced by the right side of the pronotum, which is very similar to Upinella: lateral margin evenly rounded, pronotum looks like widest at middle, posterior angles obtuse, not projected. However, we observe a strong deformation of the prothorax during the process of fossilization, because the right side has a different structure: pronotum looks widest at base, posterior angles are projected and acute. The structure of the head refutes the belonging of this fossil species to Upinella Mulsant, 1856 (species of this genus have very large eyes, small genal lobes, short and straight anterior margin of epistoma and very long antennomeres) and fully corresponds to the subfamily Tenebrioninae, especially the small reniform eyes, well-expressed genae and widely emarginated anterior margin of epistoma. In Europe, only Euboeus mimonti has a combination of characters mentioned above, body shape and size (Figs 1d, e).
Euboeus mimonti is externally similar to some flying Helopini from the genera Adelphinus Fairmaire et Coquerel, 1866 and Apterotarpela Kaszab, 1954. The mentioned taxa combine sets of characters, which cannot be strongly transformed even after deformation during fossilization: the anterior margin of epistoma is widely emarginated; the base of the elytra is much wider than the base of the pronotum; humeral angles are widely rounded (acute, pointed, narrowly rounded or absent in other Helopini). The second character might potentially look like a result of deformation in the fossil specimen of Euboeus mimonti. However, in the case of strong deformation (divergence of the elytra along the suture and eversion of the lateral sides), we would observe all nine striae, whereas on the elytra of the fossil and extant specimens only 7–8 striae are visible dorsally. Thus, we do not interpret here the feature of the elytral base as a result of deformation. Species of the genus Adelphinus differ from E. mimonti by the rectangular pronotum widest at middle and much smaller body (Nabozhenko 2015; Nabozhenko and Grimm 2019). One species, Apterotarpela clypealis (Kaszab, 1954), having an emarginated epistoma, differs from E. mimonti in the shape of the pronotum, widest at middle and smaller body (Nabozhenko and Ando 2018).
Family Tenebrionidae Latreille, 1802
Subfamily Alleculinae Laporte, 1840
Tribe Alleculini Laporte, 1840
Subtribe Gonoderina Seidlitz, 1896
Genus Pseudocistela Crotch, 1874
Pseudocistela aff. ceramboides (Linnaeus, 1758)
(Fig. 3)

Pseudocistela aff. cerambioides (GZG.W.35146a+b. Original number: 599–22): a part; b counterpart; c apex of abdomen, counterpart (arrows show intersegmental membranes between 3–5 abdominal ventrites) (scale bar = 10 mm for a and b; 5 mm for c)
Gersdorf, 1969: 323–324, taf. 16, fig. 9: Alleculidae gen. sp.
Material: 1♂: GZG.W.35146a+b. Original number: 599–22.
Comments: Part and counterpart of this print were well described by Gersdorf (1969), who suggested that the habitus and venation of hindwing of this species are similar to Prionichus Solier, 1835, Hymenalia Mulsant, 1856 and Podonta Solier, 1835. The hind wings are partially folded and cannot be used for diagnostics. Gersdorf (1969, p. 324) indicated that representatives of Podonta have most similar ratios of prothoracic length/prothoracic width, prothoracic length/elytral length and elytral length/width. He placed this species to Alleculinae (originally Alleculidae) as unclear genus. The first co-author supports the opinion of Gersdorf. The specimen has a combination of features usual for Alleculinae and not characteristic of other beetle families in the extant fauna of Europe: size moderate, length 15.8 mm (the beetle was obviously little shorter before fossilization) which makes it possible to exclude small beetles of the superfamily Tenebrionoidea with a similar habitus; large convex eyes; trapezoidal pronotum with weakly rounded lateral margins, strongly narrowed from base to apical margin; basal width of pronotum subequal to elytral base, only slightly narrower; elytra with clear striae consisting of coarse, dense round punctures; well preserved intersegmental membranes between abdominal ventrites 3–5 (Fig. 3c). These membranes are presented only in the tenebrionoid branch of Tenebrionidae (subfamilies Tenebrioninae, Blaptinae, Diaperinae, Stenochiinae and Alleculinae) and indicate the presence of defensive glands, which are widely used in the classification and phylogeny of darkling beetles (e.g. Doyen 1972; Watt 1974; Doyen and Lawrence 1979; Doyen and Tschinkel 1982; Matthews and Bouchard 2008; Matthews et al. 2010) as well as in determination of fossil tenebrionids (Nabozhenko 2019).
We can exclude from the analysis the genus Podonta (tribe Cteniopodini), which has six visible abdominal ventrites and unclear puncturation in striae (small punctures in striae are not different from those in interstriae), because the fossil species has five ventrites and clear large strial punctures. The genera Prionichus, Gonodera Mulsant, 1856 and Pseudocistela are the most similar to the fossil specimen given the size of the body. However, species of Prionichus have a more robust body and a much wider and shorter pronotum, and representatives of the genus Gonodera have the pronotum with obtuse or right posterior angles and unevenly rounded lateral margins. Species of the genus Pseudocistela are the most similar in the combination of the following characters: shape of body, large convex eyes, shape of the pronotum with acute posterior angles and punctures in elytral striae. Species of Pseudocistela have serrate antennomeres, whereas the (partly preserved) antennae of the fossil specimen look filiform. In fact, these filiform remnants are only narrow fragments of antennomeres and cannot be used for generic diagnosis.
The shape of the pronotum (evenly rounded lateral margins of the pronotum in contrast to other Western Palaearctic species having the bell-shaped pronotum with slightly emarginated or straight lateral margins in the anterior quarter (Novák 2013)) of the studied fossil corresponds to that of extant Pseudocistela ceramboides, widely distributed in Europe north to the Baltic Sea.
Family Elateridae Leach, 1815
Subfamily Denticollinae Gistel, 1856
Tribe Selatosomini Schimmel, Tarnawski, Han et Platia, 2015
Genus Selatosomus Stephens, 1830
Selatosomus sp.
(Fig. 4a)

Fossil beetles from different families, interpreted as Tenebrionidae (GZG): a Selatosomus sp. (Elateridae), GZG.W.19266, original number 52 30143; b Xylotrechus sp. (Cerambycidae), GZG.W.2901, original number 599-38, part; c the same, counterpart (scale bar = 10 mm for all images)
Gersdorf, 1976: 116, taf. 2, fig. 18a: Tenebrionidae, subfamily Akidinae Solier, 1836
Material: One print, sex unknown: GZG.W.19266. Original number: 52.30143.
Comments: This specimen was determined by Gersdorf (1976) as belonging to the tenebrionid genera Akis Herbst, 1799 or Cyphogenia Solier, 1837 (erroneously written as “Cephogonia” by Gersdorf) (Tenebrionidae: Pimeliinae: Akidini) and accompanied by a photo of Akis bacarozzo (Schrank, 1786) under the name “Cephogonia aurita”. Gersdorf based his identification solely on the shape of the pronotum with depressed lateral sides. In fact, this beetle belongs to the genus Selatosomus (Elateridae), and the edges of the prothorax were depressed during the process of fossilization. The following characters allow to exclude this species from the tenebrionid tribe Akidini: first antennomere is not concealed by gena (almost completely concealed by large genal lobe in Akidini); anterior margin of epistoma short, not emarginated (apical margin wide, widely emarginated in Akidini); eyes round, comparatively large (reniform and dorsally narrow in Akidini); antennomeres short, antennomere 3 short (at least antennomeres 3 and 4 strongly elongate, antennomere 3 longer than antennomeres 4 and 5 together in Akis and Cyphogenia); elytra with clear striae consisting of dense round punctures (without striae and strial puncturation, often without visible elytral puncturation in Akidini).
Diagnoses of the genus Selatosomus and other genera of the tribe Selatosomini are published by Schimmel et al. (2015).
Family Cerambycidae Latreille, 1802
Subfamily Cerambycinae Latreille, 1802
Tribe Clytini Mulsant, 1839
Genus Xylotrechus Chevrolat, 1860
Xylotrechus sp.
(Figs 4b, c)
Gersdorf, 1971: 652–653, taf. 62, fig. 5: Tenebrionidae.
Material: 1♀, part and counterpart: GZG.W.2901a+b. Original number: 599-38.
Comments: This species was attributed to the family Tenebrionidae based only on oral opinions of G. Schmidt (expert on Cerambycidae) and H. Korge (expert on Staphylinidae) without any argumentation. The phrase “... but the antennae expressly speak against this, and to a lesser extent the shape of the pronotum” (Gersdorf 1971, p. 653) served as an exception to this species from Cerambycidae. The following characters allow us to exclude this species from Tenebrionidae and assign it to Clytini: antennae are attached to the anterior portion of frons; eyes large and convex, emarginated around the first antennomere attachment point; elytra shorter than abdomen, with rounded apices; intersegmental membranes are absent between 3–5 abdominal ventrites. The pronotum, although somewhat deformed, has the shape and puncturation typical for Clytini. The fossil specimen has the following combination of features characteristic the genus Xylotrechus (Plavilstshikov 1940): short antennae reaching only the base of the prothorax, thickened hind femora not reaching the apex of the elytra, and an extremely elongated apex of the abdomen protruding beyond the apex of the elytra by several segments (taking into account possible deformation, three segments protrude, and in females of extant Xylotrechus spp. last two segments are often protruded). Dr Denis Kasatkin (Rostov-on-Don, Russia), an expert on Cerambycidae, confirmed our opinion and noted that the species similar to the genus Xylotrechus judging by the shape and length of femora, antennae and the last segment of the female abdomen.
Discussion
To date, 30 Coleoptera taxa determined to genus or species are known from the Willershausen Lagerstätte (Table 1). Some identifications of Erasmus Gersdorf require careful rechecking and revision as our results show. The ratio of extinct and extant taxa in this fauna deserves attention. In total, 28 genera have been identified, from which 24 (86%) are extant and four (14%) are extinct; half of the 20 identified species are extant and half are extinct. Only one genus, Cupes Fabricius, 1801 (Cupedidae), is not represented in the extant fauna of Europe and the genus Euboeus (Tenebrionidae) is absent in the extant fauna of Germany and Central Europe in general. It is noteworthy that all identified species are either extant taxa widespread in Central Europe or extinct, the latter probably representing more thermophilic forms. This material might evoke the false impression that extant species, which reduced their range as a result of the Pleistocene glaciations, did not occur in the Late Pliocene of Willershausen. For beetles, the record of a fossil specimen of the Mediterranean Euboeus mimonti fills this gap. The existence of extant beetles with strongly reduced ranges in the Late Pliocene of Willershausen may be confirmed by the probable presence of an extant Chinese species of Saperda Fabricius, 1775, which was described under the name Saperdopsis robusta G. Schmidt, 1967 (Vitali 2011, 2015).
Among other insect groups we also find extant taxa that once lived at Willershausen and have now more restricted distributions in warmer regions. Examples are the praying mantis (Mantis religiosa (Linnaeus, 1758)), in Europe restricted to the Mediterranean region and warmer parts of western Europe, although now re-colonizing northern parts of Germany (Beier 1967; Berg and Keller 2004), or the termite genus Reticulitermes Holmgren, 1913, now restricted to south-western Europe (Weidner 1971).
As a result of our study, the number of tenebrionids known from the fossil record has increased to 135 species from 88 genera. The most interesting result of our study is the record of the Mediterranean extant species Euboeus mimonti in the Late Pliocene of Willershausen.
The genus Euboeus Boieldieu, 1865 contains 78 species distributed from the Atlantic coast of Europe to Kopet Dag Mts. in western Turkmenistan and the Caspian depression in western Kazakhstan (Nabozhenko 2020, 2022). All species of this genus are flightless, the majority of species are wingless and some of them have reduced wings. The only exception is Euboeus (s. str.) mimonti, a well flying species (also fly to the light trap; personal communication and collection of beetles in Bornova – Izmir, Turkey by Bekir Keskin). This species is distributed in North Macedonia, Bulgaria, Greece and Western Turkey east to Isparta Province (Nabozhenko et al. 2017a). One adventive population is known from Erzurum in Eastern Anatolia (Nabozhenko et al. 2022). The majority of Euboeus species feeds on lichens (Nabozhenko et al. 2017b, 2021), whereas four species of the nominotypical subgenus are saprophagous and frugivorous beetles (Liberto and Leo 2006) and occur in Mediterranean landscapes with sparse forests. The record of Euboeus mimonti at Willershausen indicates that the range of this species was much wider, and the climate in the Late Pliocene in the modern territory of Lower Saxony was much milder.
Based on the climatic requirements of living counterparts, presence of growth rings (Gottwald 1981) and the high percentage of deciduousness, Ferguson and Knobloch (1998) inferred the climate at Willershausen as seasonal with mean temperatures slightly higher than at present. The palaeoforest at Willershausen was dominated by taxa such as Acer, Aesculus, Carpinus, Fagus, Quercus, Sassafras, Tilia (Mai 1995; Knobloch 1998) that are typical of hilly mesophytic woodland (Ferguson and Knobloch 1998; Knobloch 1998). These taxa also indicate relatively warmer conditions in Europe during the Late Pliocene than the present day (Adroit et al. 2018). Uhl et al. (2007) inferred an MAT at Willershausen of between 10.6 and 15.6 °C based on leaf morphology and of diversity of plant species niches. The mean temperature of the coldest month (CMMT) is estimated between 0.6 and 3.2 °C and the MAP between 897 and 1151 mm per year (Uhl et al. 2007; Thiel et al. 2012; Adroit et al. 2018).
Decreasing temperatures throughout the Pliocene caused vegetation in Europe to change from highly diverse subtropical and warm-temperate forests to temperate deciduous forests (Mai 1995). These changes, in combination with the onset of significant glaciations in the Northern Hemisphere (ca. 2.75 mya) caused widespread range retractions and distributional changes, with the presence of Euboeus mimonti at Willershausen another piece of evidence in this respect.
Kolibáč et al. (2016) presented useful information and a comprehensive bibliography about the palaeolandscapes and palaeoflora of the Willershausen Fossil Lagerstätte. They noted that insect taxa typical for mesophytic woodland dominated in this locality. Extant populations of Euboeus mimonti inhabit mainly xerophytic woodlands, which were probably present in the Pliocene of Willershausen along with more mesophytic habitats. Findings of plants common in this Lagerstätte, such as Liquidambar (Mai 1995) and Magnolia (Mai 1995; Wolkenstein and Arp 2021) may testify to this.
The genus Pseudocistela includes more than 120 species widely distributed in the world except for Australia and South America (Novák 2017); eight species are known in the Palaearctic (Novák 2020). Only one extant species Pseudocistela ceramboides occurs in Central Europe, whereas other western Palaearctic species are distributed in Mediterranean region and relic subtropical forests of Iran and Azerbaijan (Novák 2013). We can not identify this fossil specimen to any species with confidence and we admit the existence of other (extinct or extant) species of this genus in the Pliocene of Willershausen. It can only be noted that the western Palaearctic representatives of Pseudocistela inhabit old deciduous forests (P. ceramboides also in coniferous ones), and their larvae develop in rotten wood (Dubrovina et al. 1979), which corresponds to the reconstructed landscapes of this Late Pliocene locality.
Data availability
The original museum information about the material on fossil Coleoptera (Tenebrionidae and identified as Tenebrionidae) from Fossil Lagerstätte Willershausen (deposited in the Geoscience Museum at the Georg-August University in Göttingen) is available at ZENODO repository in open access: https://doi.org/10.5281/zenodo.8318703.
References
Aalbu, R. L., Kanda, K., Merkl, O., Ivie, M. A., & Johnston, A. (2023). Reconstitution of some tribes and genera of Lagriinae (Coleoptera, Tenebrionidae). ZooKeys, 1172, 155–202. https://doi.org/10.3897/zookeys.1172.103149
Adroit, B., Girard, V., Kunzmann, L., Terral, J.F., & Wappler, T. (2018). Plant-insect interactions patterns in three European paleoforests of the late-Neogene –early-Quaternary. PeerJ, 6, e5075. https://doi.org/10.7717/peerj.5075
Alekseev, V. I. , & Nabozhenko, M. V. (2023). News from Baltic amber: a new tribe of extinct Paleogene ironclad beetles (Coleoptera: Zopheridae: Zopherinae). Zootaxa, 5297(3), 435–445. https://doi.org/10.11646/zootaxa.5297.3.8
Beier, M. (1967). Mantis religiosa L. im Pliozän des Harzvorlandes. Berichte der Naturhistorischen Gesellschaft Hannover, 111, 63–64.
Berg, M., & Keller, M. (2004). Die Gottesanbeterin, Mantis religiosa Linnaeus, 1758 (Mantodea: Mantidae), im Stadtgebiet von Berlin-Schöneberg – Ihre Lebensweise und faunistische Beobachtungen in den Jahren 1998 bis 2003. Märkische Entomologische Nachrichten, 6(1), 55–84.
Brauckmann, C., Gröning, E., & Ilger, J.-M. (2013). The Late Pliocene Clausthal rhinoceros beetle from Willershausen (Scarabaeidae: Dynastinae: Oryctes? sp.; Lower Saxony, Germany). Documenta naturae, 192(1), 39–49.
Doyen, J. T. (1972). Familial and subfamilial classification of the Tenebrionoidea (Coleoptera) and a revised generic classification of the Coniontini (Tentyriidae). Quaestiones entomologicae, 8, 357–376.
Doyen, J. T., & Lawrence, J. F. (1979). Relationships and higher classification of some Tenebrionidae and Zopheridae (Coleoptera). Systematic Entomology, 4, 333–337. https://doi.org/10.1111/j.1365-3113.1979.tb00619.
Doyen, J. T., & Tschinkel, W. R. (1982). Phenetic and cladistic relationships among tenebrionid beetles (Coleoptera). Systematic Entomology, 7, 127–183. https://doi.org/10.1111/j.1365-3113.1982.tb00129.x.
Dubrovina, M. I., Gusakova, T. V. & Mamaev, B. M. (1979). Larvae of comb-clawed beetles of the subfamily Alleculinae of the fauna of the USSR. In F.N. Pravdin (Ed.), Insects – wood destroyers and their entomophages (pp. 98–129). Moscow: Nauka. [in Russian]
Ferguson, D.K., & Knobloch, E. (1998). A fresh look at the rich assemblage from the Pliocene sink-hole of Willershausen, Germany. Review of Palaeobotany and Palynology, 101, 271–286.
Gersdorf, E. (1969). Käfer (Coleoptera) aus dem Jungtertiär Norddeutschlands. Geologisches Jahrbuch, 87, 295–332.
Gersdorf, E. (1971). Weitere Käfer (Coleoptera) aus dem Jungtertiär Norddeutschlands. Geologisches Jahrbuch, 88, 629–670.
Gersdorf, E. (1976). Dritter Beitrag über Käfer (Coleoptera) aus dem Jungtertiär von Willershausen, Bl. Northeim 4226. Geologisches Jahrbuch, Reihe A, 36, 103–145.
Gottwald, H. (1981). Anatomische Untersuchungen an pliozänen Hölzern aus Willershausen bei Göttingen. Palaeontographica Abt. B, 179, 138–151.
Jordan, K.H.C. (1967). Wanzen aus dem Pliozän von Willershausen. Berichte der Naturhistorischen Gesellschaft Hannover, 115, 77–90.
Knobloch, E. (1998). Der Pliozäne Laubwald von Willershausen. Documenta naturae, 120, 1–302.
Kolibáč, J., Adroit, B., Gröning, E., Brauckmann, C., & Wappler, T. (2016). First record of the family Trogossitidae (Insecta, Coleoptera) in the Late Pliocene deposits of Willershausen (Germany). Paläontologische Zeitschrift, 90, 681–689. https://doi.org/10.1007/s12542-016-0316-6.
Korge, H. (1967). Ein fossiler Staphylinide aus dem pliozänen Ton von Willershausen im westlichen Harzvorland. Berichte der Naturhistorischen Gesellschaft Hannover, 111, 109–111.
Liberto, A., & Leo, P. (2006). Una nuova Halammobia del Peloponneso e nuovi dati faunistici sui tenebrionidi della Grecia (Coleoptera, Tenebrionidae). Fragmenta entomologica, 38(2), 251–277. https://doi.org/10.4081/fe.2006.9.
Mai, D.H. (1995). Tertiäre Vegetationsgeschichte Europas (pp. 1-691). Jena, Stuttgart, New York: G. Fischer.
Matthews, E. G., & Bouchard, P. (2008). Tenebrionid beetles of Australia: descriptions of tribes, keys to genera, catalogue of species. Canberra: Australian Biological Resources Study.
Matthews, E. G., Lawrence, J. F., Bouchard, P., Steiner, W. E. Jr., & Ślipiński, S. A. (2010) 11.14. Tenebrionidae Latreille, (1802). In R. A. B. Leschen, R. G. Beutel & J. F. Lawrence (Eds.), Handbook of zoology. Arthropoda: Insecta. Part 38. Coleoptera, beetles. Vol. 2. Morphology and systematics (Elateroidea, Bostrichiformia, Cucujiformia partim) (pp. 574–659). Berlin: Walter de Gruyter.
Meischner, D. (2000). Der Pliozäne Teich von Willershausen am Harz. In D. Meischner (Ed.), Europäische Fossillagerstätten (pp. 223–228). Berlin, Heidelberg: Springer.
Mohr, B. A. R. (1986). Die Mikroflora der oberpliozänen Tone von Willershausen (Kreis Northeim, Niedersachsen). Palaeontographica (B), 198, 133–156.
Nabozhenko, M. V. (2015). New species of the genus Adelphinus Fairmaire et Coquerel, 1866 (Coleoptera: Tenebrionidae) from Iran. Caucasian Entomological Bulletin, 11(2), 363–365. https://doi.org/10.23885/1814-3326-2015-11-2-363-365.
Nabozhenko, M. V. (2019). The fossil record of darkling beetles (Insecta: Coleoptera: Tenebrionidae). Geosciences, 9(12), 514. https://doi.org/10.3390/geosciences9120514.
Nabozhenko, M. V. (2020). Tribe Helopini Latreille, 1802. In D. Iwan & I. Löbl (Eds.), Catalogue of Palaearctic Coleoptera, Volume 5. Revised and updated second edition. Tenebrionoidea (pp. 314–339). Leiden: Brill. https://doi.org/10.1163/9789004434998_004.
Nabozhenko, M. V. (2022). A taxonomic review of the genus Euboeus Boieldieu, 1865 (Coleoptera: Tenebrionidae: Helopini) of the Caucasus, Iran and Turkmenistan. Zootaxa, 5159(4), 451–486. https://doi.org/10.11646/zootaxa.5159.4.1.
Nabozhenko, M. V., & Ando, K. (2018). Subtribal, generic and subgeneric composition of darkling beetles of the tribe Helopini (Coleoptera: Tenebrionidae) in the Eastern Palaearctic. Acta zoologica Academiae Scientiarum Hungaricae, 64(4), 277–327. https://doi.org/10.17109/AZH.64.4.277.2018.
Nabozhenko, M. V., & Grimm, R. (2019). New species and new records of darkling beetles of the tribe Helopini (Coleoptera: Tenebrionidae) from the Western Palaearctic. Caucasian Entomological Bulletin, 15(1), 107–116. https://doi.org/10.23885/181433262019151-107116.
Nabozhenko, M. V., & Perkovsky, E. E. (2023). Oracula campbelli sp. n. – the first fossil darkling beetle (Coleoptera: Tenebrionidae: Alleculinae) from the late Eocene Rovno amber (Ukraine). Zootaxa, 5230(2), 245–250. https://doi.org/10.11646/zootaxa.5230.2.8.
Nabozhenko, M. V., & Tanaka, T. (2022). Promethis undulatus sp. n. – the first fossil darkling beetle (Coleoptera: Tenebrionidae: Stenochiinae: Cnodalonini) from the late Miocene of Japan. Caucasian Entomological Bulletin, 18(2), 273–278. https://doi.org/10.23885/181433262022182-273278.
Nabozhenko, M. V., & Tanaka, T. (2023). Species of the genus Lagria Fabricius, 1775 from Kabutoiwa Member in Central Japan – the first fossil darkling beetles (Coleoptera: Tenebrionidae: Lagriinae) from the Pliocene. Paleontological Research, 28(2), 1–8. https://doi.org/10.2517/PR220040.
Nabozhenko, M. V., Nikitsky, N. B., & Keskin, B. (2017a). Taxonomic review of the genus Euboeus s. str. Boieldieu, 1865 (= Probaticus s. str. Seidlitz, 1896, syn. n.) (Coleoptera, Tenebrionidae). Zootaxa, 4358(3), 494–506. https://doi.org/10.11646/zootaxa.4358.3.6.
Nabozhenko, M. V., Keskin, B., & Nabozhenko, S. V. (2017b). Life forms and strategies of lichen-feeding darkling beetles (Coleoptera, Tenebrionidae: Helopini). Entomological Review, 97(6), 735–746. https://doi.org/10.1134/S0013873817060045.
Nabozhenko, M. V., Keskin, B., Alpagut Keskin, N., Gagarina, L. V., & Nabozhenko, S. V. (2021). Two new species and new records of lichen-feeding darkling beetles (Coleoptera: Tenebrionidae: Helopini) from Turkey with notes on bionomics and trophic relations. Zootaxa, 5057(1), 69–86. https://doi.org/10.11646/zootaxa.5057.1.4.
Nabozhenko, M. V., Doğan, D., & Yıldırım, E. (2022). Additions to the knowledge of the diversity of darkling beetles (Coleoptera: Tenebrionidae) from Turkey with new records and taxonomic notes. Journal of insect biodiversity, 32(1), 5–25. https://doi.org/10.12976/jib/2022.32.1.2.
Novák, V. (2013). Review of the West Palaearctic Pseudocistela with description of P. hajeki sp. nov. from Iran (Coleoptera: Tenebrionidae: Alleculinae). Acta Entomologica Musei Nationalis Pragae, 53(1), 293–301.
Novák, V. (2015). Review of the genus Upinella Mulsant, 1856 stat. nov. (Coleoptera: Tenebrionidae: Alleculinae). Folia Heyrovskyana, series A, 23(2), 89–111.
Novák, V. (2017). Pseudocistela ornata sp. nov. from Laos and new nomenclatory acts in Gonoderini (Coleoptera: Tenebrionidae: Alleculinae). Studies and Reports. Taxonomical Series, 13(2), 447–453.
Novák, V. (2020). Subfamily Alleculinae Laporte, 1840. In D. Iwan & I. Löbl (Eds.), Catalogue of Palaearctic Coleoptera, Volume 5. Revised and updated second edition. Tenebrionoidea (pp. 417–453). Leiden: Brill. https://doi.org/10.1163/9789004434998_004.
Plavilstshikov, N.N. (1940). Fauna of the USSR. Insects Coleoptera. Vol. 22. Longhorn beetles (Part 2). Moscow, Leningrad: Academy of sciences of the USSR publ.
Schimmel, R., Tarnawski, D., Han T., & Platia, G. (2015). Monograph of the new tribe Selatosomini from China (Elateridae: Denticollinae). Part I. Polish Entomological Monographs, Vol. 11. Polish Entomological Society, Poznan.
Schmidt, G. (1967). Die Bockkäfer (Cerambycidae) von Willershausen. Berichte der Naturhistorischen Gesellschaft Hannover, 111, 113–120.
Schweigert, G. (2003). Alpenbock und Hirschkäfer im Pliozän von Willershausen. Fossilien, 20(3), 178–182.
Thiel, C., Klotz, S., & Uhl, D., 2012. Palaeoclimate estimates for selected leaf floras from the late Pliocene (Reuverian) of Central Europe based on different palaeobotanical techniques. Turkish Journal of Earth Sciences, 21(2), 263–287. https://doi.org/10.3906/yer-1007-41.
Uhl, D., Klotz, S., Traiser, C., Thiel, C., Utescher, T., Kowalski, E., & Dilcher, D.L. (2007). Cenozoic paleotemperatures and leaf physiognomy – a European perspective. Palaeogeography, Palaeoclimatology, Palaeoecology, 248(1–2), 24–31. https://doi.org/10.1016/j.palaeo.2006.11.005.
Vitali, F. (2011). Short Notes IV. Saperdopsis robusta Schmidt, 1967: an extant Chinese species? (Coleoptera Cerambycidae). Entomapeiron (P. S.), 4(1), 35–36.
Vitali, F. (2015). Saperda caroli n. sp., a new fossil species from Early Eocene of Colorado, with taxonomic remarks on its extinct congeners (Coleoptera, Cerambycidae). Les Cahiers Magellanes, NS, 17, 20–28.
Watt, J. C. (1974). A revised subfamily classification of Tenebrionidae (Coleoptera). New Zealand Journal of Zoology, 1, 381– 452. https://doi.org/10.1080/03014223.1974.9517846.
Wegele, H. (1914). Stratigraphie und Tektonik der tertiären Ablagerungen von Oldenrode-Düderode-Willershausen (pp.1-38). Inaugural Dissertation, Georg August Universität Göttingen.
Weidner, H. (1971). Zwei neue Termitenarten aus dem Pliozän von Willershausen. Berichte der Naturhistorischen Gesellschaft Hannover, 115, 41–46.
Wolkenstein, K., & Arp, G. (2021). Taxon- and senescence-specific fluorescence of colored leaves from the Pliocene Willershausen Lagerstätte, Germany. Paläontologische Zeitschrift, 95, 747–756. https://doi.org/10.1007/s12542-020-00538-3.
Acknowledgements
The authors are sincerely grateful to Dr Denis Kasatkin (Rostov Branch of the All-Russian Plant Qurantine Center, Rostov-on-Don, Russia) for confirming the identification of the fossil Cerambycidae and help with the diagnosis of the genus, Dr Francesco Vitali (Musée national d’histoire naturelle de Luxembourg) for sending important bibliographic sources and Ivan Chigray (ZIN) for the loan specimens of Euboeus mimonti. Two reviewers, André Nel (Muséum National d’Histoire Naturelle, Paris, France) and Wolfgang Schawaller (Staatliches Staatliches Museum für Naturkunde Stuttgart, Germany) added useful comments and corrections, for which the authors are very grateful. This work was supported by the state project AAAA-A17-117081640018-5 (Ministry of Higher Education and Science of the Russian Federation) for M.V. Nabozhenko.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
The authors declare that they have no conflict of interest.
Additional information
Publisher's note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Springer Nature or its licensor (e.g. a society or other partner) holds exclusive rights to this article under a publishing agreement with the author(s) or other rightsholder(s); author self-archiving of the accepted manuscript version of this article is solely governed by the terms of such publishing agreement and applicable law.
About this article

Cite this article
Nabozhenko, M.V., Jenkins Shaw, J., Gehler, A. et al. Euboeus mimonti Boieldieu, 1865, the oldest record of an extant species of Tenebrionidae (Coleoptera) and notes on other species identified as darkling beetles from the Late Pliocene of Willershausen (Germany). Palaeobio Palaeoenv 104, 141–151 (2024). https://doi.org/10.1007/s12549-023-00592-0
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s12549-023-00592-0