
Abstract
The Eocene biota from the pre-rifting stage of the Upper Rhine Graben (southwest Germany) is, with exception of the world-famous fossil sites Messel and Bouxwiller, poorly known. While from these localities exclusively terrestrial and freshwater fossils were recovered, here we present floral and faunal assemblages from the middle Upper Rhine Graben which clearly indicate a temporarily brackish water environment and consist of a diverse palynomorph association, calcareous nannoplankton, foraminifers, ostracods and some fish and reptile remains. Based on the occurrence of Trochastrites hohnensis, a Lutetian age is assumed. Organic-walled dinoflagellate cysts, in particular Phthanoperidinium comatum, Phthanoperidinium echinatum, Apectodinium homomorphum and Apectodinium quinquelatum, suggest a late Ypresian to Lutetian age. The pollen and spores assemblage includes typical mid Eocene species, such as Tegumentisporis villosoides, Tricolporopollenites crassostriatus and representatives of Bombacaceae, but a late Ypresian to Priabonian age cannot be excluded. Foraminifers and ostracods do not further refine the biostratigraphical assignment. Hence, a Lutetian age is most probable. The mass occurrence of Neocyprideis, various foraminifer taxa and an organic-walled dinoflagellate cyst assemblage of very low diversity are indicative of a brackish water environment. Disarticulated vertebrate remains include fish teeth of Lepisosteidae, turtle plates and alligatoroid teeth of Diplocynodon and Hassiacosuchus. The present palaeogeographical scenarios do not consider a connection from the Upper Rhine Graben to the North Sea Basin, Alpine Sea/Paratethys or Paris Basin during the mid Eocene. Provided that the middle Upper Rhine Graben was land-locked and definitely not reached by a marine ingression during this time interval, we tentatively suggest that the brackish water taxa may have been accidentally introduced into a brackish inland sea by wind (anemochory), rain, highly mobile insects or vertebrates such as fish, birds and mammals (endozoochory/ectozoochory). The presumably freshwater calcareous nannoplankton species Nannoserratolithus minutus Martini is newly described.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
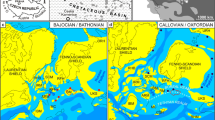
The almost north–south oriented Upper Rhine Graben is the central part of the intracontinental European Cenozoic Rift System which extends from the Western Mediterranean to the North Sea. The Upper Rhine Graben developed in the northwestern foreland of the Alps (Bourgeois et al. 2007). The continental rift zone is approximately 35–45 km across and ranges over 300 km from Basel in the south to Frankfurt in the north, i.e. from the Jura Mountains to the Rhenish Massif. Based on different syn-sedimentary graben subsidence rates and sediment fill various sub-grabens and -basins can be delimitated (Fig. 1). Precise levelling confirms that the fault system is still active to these days (e.g. Behrmann et al. 2003; Sissingh 2003).

Structural sketch map of the Upper Rhine Graben. URG Upper Rhine Graben; MB Mainz Basin; HB Hanau Basin; HeB Heidelberg Basin; KB Karlsruhe Basin; PB Pechelbronn Basin; SFS Saverne Fault System; MuB Mulhouse Basin. Modified after Sissingh (2006) and Grimm et al. (2011b). For detailed geological and palaeontological descriptions of important Eocene fossil sites in the middle and northern Upper Rhine Graben see e.g. Sprendlinger Horst: geological map of the area with fossil localities (Mutzl et al. 2022); geology and palaeontology (Smith et al. 2018); palynology (Thiele-Pfeiffer 1988; Lenz et al. 2007, 2011, 2015; Lenz and Wilde 2018; Moshayedi et al. 2018, 2020, 2021; Mutzl et al. 2022) – Eisenberg: description of boreholes, section in clay pit Erlenhof, lithostratigraphy (Schäfer 2000); palynology (Hottenrott 1998, 2000) – Bouxwiller and adjacent localities: lithology (Trauth et al. 1977); palaeontology (Lavoyer 2013); palynology (Schuler and Ollivier-Pierre 1981; Schuler 1990; Sittler and Ollivier-Pierre 1994)
At the northern end of the graben, between Mainz and Frankfurt, the Upper Rhine Graben bifurcates into a NNE–SSW and a NW–SE striking branch. This bifurcation is described as the Rhenish Triple Junction (Sissingh 2003). In the NNE–SSW branch the Upper Rhine Graben continues into several smaller Hessian grabens and basins (e.g. Wetterau Graben, Horloff-Graben, Hessen Depression with small grabens). In the Northwest German Basin, NNE–SSW striking faults are regarded as a prolongation of the graben. The NW–SE striking graben trend includes the Neuwied Basin and the Lower Rhine Embayment which structurally extends into the Roer Valley Graben. At least parts of the European Cenozoic Rift System are pre-Mesozoic structures. They developed in ancient crustal weakness zones which probably originated from late Variscan, Permo-Carboniferous fracture systems. The Paleozoic faults were repeatedly re-activated during the Mesozoic and Cenozoic due to roughly N–S oriented, syn-collisional compression (Sissingh 2003; Allenbach and Wetzel 2006) which probably has been induced by the evolving Alpine and Pyrenean orogens (e.g. Dèzes et al. 2004).
Due to intense hydrocarbon, potash/salt, and geothermal exploration in the Upper Rhine Graben, an extensive data set comprising numerous deep wells and seismic sections has accumulated and hence, the graben has become one of the best-studied rift zones in the world. The graben is the oldest known oil province in Western Europe. Oil shows and tar sands triggered exploitation dating back to 1498 (Sittler 1972; Böcker et al. 2017). The region is still in the focus of hydrocarbon exploration as in 2003 the Römerberg oil field, one of the most significant oil reservoirs of Germany, has been discovered near Speyer during drilling of a geothermal well. Further oil fields are located e.g. near Pechelbronn, Landau and Stockstadt (Perner et al. 2018). Potash/salt exploration concentrated on the southern depocentre of the Upper Rhine Graben where in the Mulhouse Basin upper Eocene (Priabonian) to Oligocene evaporites including potash and halite were precipitated. North of this basin, near Sélestat, Strasbourg and Pechelbronn, halite and sulphate bearing successions are known from the subsurface (e.g. Cendón et al. 2008). Numerous deep geothermal boreholes have been drilled in areas of positive temperature anomalies to exploit energy from deep-seated granite and Paleozoic and Mesozoic sedimentary successions, e.g. in Landau, Soultz-sous-Forêts and Rittershoffen in eastern France (see Baillieux et al. 2013; Duringer et al. 2019; Frey et al. 2022).
The sedimentary fill of the Upper Rhine Graben reaches a maximum thickness of as much as 4000 m (Behrmann et al. 2003). The tectono-sedimentary evolution of the graben is complex. The magnitude of subsidence was different in various parts of the graben and together with climatic effects and transgressions from the Paleogene North Sea and the Alpine Molasse Basin decisive for the depositional history of the graben fill.
In the Upper Rhine Graben, the early rifting stage started in the early Eocene (Ypresian) to mid Eocene (Lutetian/Bartonian). Numerous shallow and isolated depressions, partly fault-controlled, developed separated by highs. The palaeogeographically confined depressions were filled with predominantly fluvio-lacustrine sediments. The thickness of the lowermost Schliengen Formation (Fig. 2) (= Siderolith-Schichten; Schäfer 2013) in the southern and middle part of the graben varies between less than a metre and up to 108 m. The formation consists of residual soils and intercalated sand horizons. As the sediments are barren with regard to non-reworked fossils, age assignment is difficult. In places they are overlain by the middle Lutetian Bouxwiller Formation (see below) (Grimm et al. 2011b). The mostly pelitic Eozäner Basiston in the middle and northern part of the Upper Rhine Graben is similar to the Schliengen Formation. The Eozäner Basiston is characterised by strong thickness variation, deposition in local depressions and the lack of time-indicative fossils. The basal variegated Eocene clays are also barren with regard to palynomorphs (e.g. Sittler 1968a). A latest Ypresian to early Lutetian age is assumed (Grimm et al. 2011b). The upper Ypresian to lower Lutetian Messel Formation (Lenz et al. 2015), world-famous for a highly diverse and exquisitely preserved flora and fauna, is mostly restricted to ancient maar lakes on the Sprendlinger Horst (e.g. maar lakes Messel and Offenthal; Smith et al. 2018; Moshayedi et al. 2020). Another biostratigraphically important site is Bouxwiller, type locality for the middle Lutetian Bouxwiller Formation which is the basal unit of the Haguenau-Gruppe. The clayey marls and limestones, deposited in small freshwater lakes, are rich in e.g. charophytes, land and freshwater snails and vertebrate remains, in particular mammal marker species (Lavoyer 2013). An intercalated thin coal seam yielded a diverse and well preserved palynomorph association (Schuler and Ollivier-Pierre 1981; Schuler 1990; Sittler and Ollivier-Pierre 1994). The middle unit of the Haguenau-Gruppe are the Grüne Mergel which are at least partially time-equivalent to the Lymnäenmergel and Landauer Mergel sensu Breuer and Feist (1986) (for a detailed discussion on Eocene lithostratigraphical terms in the Upper Rhine Graben see Stratigraphical synthesis). Based on charophytes and mammal remains the Grüne Mergel are dated as mid Lutetian to mid Priabonian. Deposition took place in an inland sea subdivided by highs. In the middle Upper Rhine Graben, they consist mainly of grey to greenish clays and calcareous marls. Because of the occurrence of rare organic-walled dinoflagellate cysts (dinocysts) and a low-diversity foraminiferal fauna, brief marine ingressions from the south (Schäfer 2013) or west (Grimm et al. 2011b: 59) are assumed. In the southern part thick deposits of evaporites such as anhydrite/gypsum, halite and potash give evidence of hypersaline conditions. Highest thickness values exceeding several hundred metres have been recorded from the southern and middle part of the Upper Rhine Graben (Grimm et al. 2011b). Except for a few dinocysts, foraminifers and ostracods the invertebrate fauna is dominated by land and freshwater gastropods, questionable brackish water gastropods and bivalves (e.g. Sittler 1969). A well from the Landau oilfield (Landau 117) encountered 6 m of olive grey to green grey, fossiliferous claystones and clayey marlstones. Charophytes, foraminifers, ostracods and vertebrate remains have been recovered but only the charophytes are taxonomically treated indicating a position in the Lutetian Maedleriella embergeri charophyte zone (Breuer and Feist 1986). Some vertebrate remains from the same core of this well have been described and figured by Weiler (1963). The Grüne Mergel are overlain by the middle Priabonian Rote Leitschicht, a distinct marker horizon. In the southern Upper Rhine Graben, a bed containing foraminifers and calcareous nannoplankton is indicative of a marine ingression (Ohmert 1993; Martini and Reichenbacher 2007). Starting with the Pechelbronn-Gruppe, i.e. in the late Priabonian, the main rifting stage commences. At the base, terrestrial, limnic, fluviatile and marine deposits were laid down but during the early Rupelian (Mittlere Pechelbronn-Schichten) the marine influence increased due to the first marine transgression from the Paleogene North Sea into the Upper Rhine Graben via the Hessian Depression. It is assumed that this resulted in a connection between the North Sea Basin and the North Alpine Molasse Basin (Sissingh 2006).

Lithostratigraphy of the Paleocene to lowermost Oligocene in the middle Upper Rhine Graben. Based on Grimm et al. (2011b) and Schäfer (2013). Numerical ages (Ma) following International Chronostratigraphic Chart, 2022/10 (https://stratigraphy.org/ICSchart/ChronostratChart2022-10.pdf)
Apart from the fossil-rich Messel and Bouxwiller formations, exposed in few old pits, palaeontological data from Eocene sediments of the Upper Rhine Graben are sparse as they are mostly overlain by a thick sedimentary succession of Oligocene to Quaternary age (e.g. Straub 1955; Wirth 1969; Breuer and Feist 1986). Fossil assemblages from the early rifting stage are particularly interesting as they are useful in unravelling complicated facies relationships such as changes in salinity, water depth and oxygen supply. In addition, some fossil groups, e.g. palynomorphs, calcareous nannoplankton, charophytes, foraminifers, ostracods and mammals, have been successfully applied to develop a biostratigraphical scheme for marine to terrestrial sediments of the Upper Rhine Graben. However, very few Eocene fossil sites yielded more than one age-indicative fossil group (e.g. wells in the Landau oil field; see Wirth 1969; Breuer and Feist 1986) which complicates biostratigraphical correlations of the oldest graben-fill sediments deposited in small and isolated depressions.
This study focuses on Eocene fossil assemblages recovered from a borehole and two temporary outcrops on the western shoulder of the middle Upper Rhine Graben which were accessible during earthworks for the bypass highway B 10 between Albersweiler and Siebeldingen, west of Landau (Fig. 3). The assemblages comprise palynomorphs, calcareous nannoplankton, foraminifers, ostracods, molluscs (Kadolsky 2015; Kadolsky and Hartkopf-Fröder, in prep.) and vertebrate remains (teleosts, reptiles, but no mammals). Overall, fossils are abundant and well preserved but apart from palynomorphs and otoliths (Bettina Reichenbacher, written communication, February 01, 2023) of low diversity. The presence of various age diagnostic and palaeoecologically significant fossil groups is exceptional for Eocene sediments in the Upper Rhine Graben. Hence, the main objectives of this interdisciplinary study are to examine the fossiliferous sediments including organic geochemical analyses and to give a comprehensive taxonomic account of the palynomorphs, calcareous nannoplankton, foraminifers, ostracods and disarticulated vertebrate skeletal remains (fish teeth, reptiles) in order to present a precise age assignment based on pollen, spores, dinocysts and calcareous nannoplankton. In particular from the palaeontological data, a palaeoenvironmental reconstruction and palaeogeographical indications will be inferred. In addition, the hydrocarbon generation potential of the studied samples is compared with the age-equivalent succession from the nearby but more graben-wards Landau oilfield region.

Simplified map of the Birkweiler area showing outcrop localities and site of borehole B 804
In the Upper Rhine Graben many lithostratigraphical units are not yet formally defined. Stratigraphical correlations are frequently hampered by the presence of various sub-basins with different subsidence, thickness variations and separate geological history (e.g. Derer 2003; Duringer et al. 2019), small-scale facies heterogeneities and complex depositional patterns due to syn-sedimentary graben tectonics and relative sea-level fluctuations (e.g. marine–brackish–freshwater; Schuler 1983; Rauscher and Schuler 1988; Rauscher et al. 1988; see also Derer 2003), different stratigraphical schemes used in France, Germany and Switzerland in former times (e.g. Pirkenseer et al. 2018), scattered and scarce fossils, the lack of state-of-the-art taxonomic research, reworking of microfossils (e.g. Pirkenseer et al. 2011) and decalcification of the host sediment, to mention but a few. Hence, numerous local lithostratigraphical terms exist. We here follow Grimm et al. (2011b) and Schäfer (2013) who summarised Paleogene/Neogene lithostratigraphical units of the Upper Rhine Graben. We refrain from translating these units into English but keep applying the original German and French terms.
Geological setting of the Albersweiler–Siebeldingen area
The western shoulder of the Upper Rhine Graben in southern Palatinate and the northern Vosges (Fig. 1) is one of the key areas for the interpretation of the tectono-sedimentary evolution of the rift system (e.g. Illies 1963; Stapf 1988; Lopes Cardozo and Behrmann 2006). The area has been intensively studied with regard to stratigraphy, syn-rift sedimentation, structural configuration and petroleum systems because of relatively good exposures of upper Paleozoic and lower Mesozoic rocks, numerous borehole data obtained during hydrocarbon exploration of the economically important nearby Landau oilfield (e.g. Doebl and Bader 1970; Bruss 2000; Böcker and Littke 2014, 2016; Böcker et al. 2017), some isolated Upper Triassic and Lower Jurassic outcrops (Nottmeyer 1954), and the westernmost exposures of the Mid-German Crystalline High (Reischmann and Anthes 1996; Frey et al. 2021). Stratigraphical and tectonic research resulted in several large-scale geological maps (e.g. Nottmeyer 1954; Illies 1963; Doebl and Bader 1970; Stellrecht 1971; Hartkopf 1981) which provide a detailed survey on the regional geology between Eschbach and Burrweiler.
However, the existence of numerous deep-seated landslides widespread on hill slopes formed by Triassic, Paleogene and Neogene clays and marls has been neglected or underestimated (e.g. Nottmeyer 1954) although in some areas of the Mainz Basin, where similar geological conditions are found, extensive landslides are well known. Here, landslides with a slip plane in depths of up to 25 m can cover some square kilometres (Schäfer 2012). To the north and east of Birkweiler numerous landslides have been observed, e.g. by Nottmeyer (1954) and Hartkopf (1981), but their considerable dimensions have been fully captured not until earthworks started for the bypass highway B 10 by which an entire hill slope became displaced which subsequently required large-scale stabilisation measures (Wichter et al. 1988). The reactivated fossil slip planes are up to 26 m deep, indicating massive ground movements. Following Schröder (1984) and Wichter et al. (1988), in some of the numerous boreholes Paleogene/Neogene sediments are supposed to be overlain even by dislocated Permian (Rotliegend), Upper Triassic (Keuper) and Lower Jurassic rocks. The thickness of the displaced material is claimed to reach at least 13 m (e.g. in borehole B 704; Schröder 1984: 11; Wichter et al. 1988: fig. 2). In the nearby borehole B 703 Paleogene/Neogene strata are alleged to be overlain by ca. 29 m of Lower Jurassic rocks which may be due to a landslide with the slip plane near 17 m and additional compressional tectonics. However, a re-evaluation of the microfossil assemblage of boreholes B 703 and B 704 proved a Late Triassic (Keuper) age for those strata assigned to the Paleogene/Neogene by Wichter et al. (1988: fig. 2). Probably, in these two boreholes an undisturbed succession with Keuper sedimentary rocks at the base overlain by Lower Jurassic mudstones/marlstones has been encountered. Regardless of whether displaced Lower Jurassic mudstones overlay Paleogene/Neogene ones or the Upper Triassic/Lower Jurassic succession is undisturbed, in such an extensive landslide area in situ and displaced material are hardly to differentiate. Particularly in shallow outcrops and boreholes with a total depth above the slip planes, displaced material cannot be identified as a result of landslide mass movements. A reliable geological map depicting those strata which are still in their original position below dislocated rock masses can hardly be constructed, particularly as the area is also characterised by complex fault tectonics related to the western border fault of the Upper Rhine Graben. Consequently, Wichter et al. (1988) refrained from constructing a geological map in spite of having available data from 172 boreholes.
In the road cut of the bypass highway B 10 from west to east Permian (southwest of Albersweiler), Eocene (north of Birkweiler) and Lower Jurassic (west of Siebeldingen) sedimentary rocks were exposed.
To the east, the Upper Rotliegend (Permian) red beds of the Wadern Formation (Boy et al. 2012) are bordered by a major fault (“westliche Rheingrabenrandstörung” in the terminology of Stapf 1988), which roughly runs in north–south direction from west of Birkweiler through Albersweiler. The red beds are mainly composed of conglomerates and sedimentary breccias with some intercalated thin sandstone and mudstone beds. The coarse-grained facies is interpreted as alluvial fan deposits (fanglomerates) which unconformably overlay the Albersweiler gneiss complex (Reischmann and Anthes 1996).
Lower Jurassic mudstones were rather well accessible in the embankment of the ramp to the B 10 where still some small outcrops exist today. In the road cut, these mudstones were poorly exposed due to extensive landslides. The brickyard pit west of Siebeldingen was shut down years ago and when the roadworks started exposures were not accessible anymore due to recultivation. This locality was a well-known fossil site (e.g. Drexler 1958; Heidtke 2006) which yielded a diverse marine fauna including foraminifers, ostracods, molluscs and rare vertebrate remains. The Lower Jurassic sedimentary succession is composed of grey to black mudstones and marlstones. They are rich in organic matter reaching up to approx. 4.5 % TOC (total organic carbon; Böcker and Littke 2016). Thin horizons of marly limestones up to 20 cm thick and limestone concretions up to 10 cm in diameter are intercalated. The Lower Jurassic succession in the Siebeldingen area has been described in detail by Jüngst (1938) and Drexler (1958). Based on the succession in the brickyard pit west of Siebeldingen Jüngst (1938: 154) assigned the dark grey mudstones to the Lias α and lower Lias β (= Hettangian and Sinemurian) while the presence of Lias γ (Pliensbachian) was assumed to be doubtful. The occurrence of Passaloteuthis paxillosus (von Schlotheim, 1813) depicted in Heidtke (2006: fig. 6) clearly indicates that Pliensbachian sedimentary rocks were present in the brickyard section (Günter Schweigert, written communication, February 01, 2023).
Material
Samples were collected along the road cut of the new bypass highway B 10 north of Birkweiler (a municipality between Landau in der Pfalz and Annweiler am Trifels, southwestern Germany) in 1980–1984 and from boreholes drilled for highway construction and landslide stabilisation (Figs. 3, 4).

Section of borehole B 804 and Birkweiler road cut. Position of slip planes in borehole B 804 following Schröder (1984). Note that lithological description for borehole B 804 is based on data provided by the drilling company
Extensive earthworks were necessary as the bypass crosses an approx. 700 m long vineyard hill slope which was known to be prone to landslides since a long time (Schröder 1984; Wichter et al. 1988). During these earthworks fossiliferous Eocene sediments were exposed. Sampling concentrated on a stromatolite horizon and greenish marl horizons rich in hydrobioid gastropods as well as otoliths (Figs. 3, 4). This fossil site is hereafter named “Birkweiler road cut” (map TK 25 Sheet 6714 Edenkoben; UTM coordinates (WGS84) ca. 32U 429652 5451280; ca. R 34 29700 H 54 53022).
From this road cut Franz Doebl, then with the Wintershall AG, Betrieb Landau, sent six samples to the Geologischer Dienst Nordrhein-Westfalen, Krefeld for palynological analyses in June and July 1979. Of these a lignite sample (Fig. 3) yielded an Eocene miospore assemblage (sample no. 74933; UTM coordinates (WGS84) ca. 32U 429717 5451231; ca. R 34 29765 H 54 52973). The assemblage of a second sample (sample no. 74936, UTM coordinates (WGS84) ca. 32U 429735 5451220; ca. R 34 29783 H 54 52962) was badly preserved so that a biostratigraphical assignment was not possible. The four remaining samples proved to be barren with regard to palynomorphs.
After the beginning of the earthworks for the bypass highway B 10 north of Birkweiler new and newly reactivated landslides occurred which were tried to be stabilised by deep drainage galleries, concrete dowels, bore pile walls and rockfills. In addition, numerous exploration and inclinometer borings, some of them entirely cored, were drilled (Schröder 1984; Wichter et al. 1988). The samples were biostratigraphically investigated by Volker Sonne (then Geologisches Landesamt Rheinland-Pfalz) using foraminifers and ostracods but the results remained unpublished. This material, housed in the Landesamt für Geologie und Bergbau Rheinland-Pfalz, was available for examination. Especially important is borehole B 804 (map TK 25 Sheet 6714 Edenkoben; UTM coordinates (WGS84) 32U 429627 5451226; R 34 29675 H 54 52968; Figs. 3, 4) which yielded foraminifer, ostracod, hydrobioid and otolith assemblages. Core samples of some other boreholes (A 1–A 10) are housed in the Landesamt für Geologie und Bergbau Rheinland-Pfalz and were studied for palynology. Only samples from boreholes A 9 (map TK 25 Sheet 6714 Edenkoben; UTM coordinates (WGS84) 32U 429647 5451294; R 34 29695 H 54 53036) and A 10 (map TK 25 Sheet 6714 Edenkoben; UTM coordinates (WGS84) 32U 429704 5451191; R 34 29752 H 54 52933) yielded an Eocene palynomorph assemblage. All other samples proved either to be barren with regard to palynomorphs or to be Mesozoic in age and therefore are not considered in this study.
During the stabilisation measures, grey marls rich in hydrobioids, some other gastropod species and otoliths were collected from already excavated material of a three metres deep drainage pit right beside the highway and very close to the Birkweiler road cut locality (map TK 25 Sheet 6714 Edenkoben; UTM coordinates (WGS84) ca. 32U 429700 5451248; ca. R 34 29748 H 54 52990). As the pit was entirely boarded neither a profile could be measured nor any samples could be recovered directly from the section. Hence, samples are stratigraphically unconstrained spot samples not in stratigrapical order. Material from this site is referred to as “Birkweiler drainage pit” (Fig. 3).
Methods
Vitrinite reflectance and organic geochemistry
Vitrinite reflectance analysis was performed on six samples from the Birkweiler drainage pit and a well from the Landau oilfield following standard techniques as outlined in Böcker et al. (2017).
Three sediment samples (Albersweiler A 10, A 17 and A 19, all from the Birkweiler drainage pit) were analysed for saturated hydrocarbon distributions as described previously (Hartkopf-Fröder et al. 2007). Briefly, ground rock samples were extracted using dichloromethane containing 1 % methanol. Extract yields varied between 0.81 and 1.14 mg/g rock. The extracts were separated into compound class fractions by medium pressure liquid chromatography and the saturated hydrocarbon fractions subsequently analysed by gas chromatography and gas chromatography-mass spectrometry according to standard procedures.
Palynomorphs
Six palynological samples from the Birkweiler drainage pit were processed applying standard preparation methods, i.e. HCl-HF-HCl digestion and sieving with a 10 μm polyester fabric mesh (Ashraf and Hartkopf-Fröder 1996). Permanent strew mounts were produced with PVA as the mounting medium and Elvacite 2044TM epoxy resin as the embedding medium. Samples were analysed under a Zeiss Axioplan microscope (1000x magnification) and the software TILIA (Grimm 1993, 2004) was used for the calculation of the pollen histogram. Terrestrial pollen sums were > 250 for each sample. Dinocyst analysis was performed at 500x magnification using a Carl Zeiss Jena Jenaval microscope equipped with differential interference contrast optics. Photography was made in plain transmitted light at 1000x magnification in an Leitz Orthoplan microscope. After a first examination of an appropriate area of the slides and identification of taxa, a subsequent count of approx. 100 specimens was made, where possible. Except for sample 108581, a much higher number of specimens than counted has been observed in each sample.
Calcareous nannoplankton, foraminifers and ostracods
34 samples from the Birkweiler drainage pit were analysed for calcareous nannoplankton using smear slides, standard light microscopy and SEM techniques.
Foraminifer and ostracod samples were collected from the Birkweiler road cut, borehole B 804 and the Birkweiler drainage pit. The sediment was disaggregated by washing through a 63 μm sieve, followed by gravity separation with CCl4 in order to enrich the microfossils from the dried residue.
Vertebrate remains
To recover molluscs (Kadolsky and Hartkopf-Fröder, in prep.), otoliths and other vertebrate remains (e.g. scales, teeth, see below), sediment from the Birkweiler drainage pit was disaggregated using 5 % hydrogen peroxide solution and washed through a 200 μm sieve. From the dried residue the fossils were handpicked. To prepare cross-sections of fish teeth single specimens were embedded in epoxy resin, ground and polished by hand. Photographs were taken using a Zeiss Axio Imager.M2m incident light microscope equipped with EC Epiplan Neofluar objectives, a HXP 120C light source and a fluorescence filter set 09 (excitation BP 450–490 nm, beam splitter FT 510 nm, emission LP 515 nm) housed in the microscopy lab of the Lehrstuhl für Geologie, Geochemie und Lagerstätten des Erdöls und der Kohle, RWTH Aachen.
Storage of material
Material used in this study is accessioned into the following collections:
-
Sediment samples, thin sections, polished slabs of stromatolites: Geologischer Dienst Nordrhein-Westfalen, Krefeld; all material will be later transferred to the Landesamt für Geologie und Bergbau Rheinland-Pfalz, Mainz.
-
Polished sections for vitrinite reflectance: Lehrstuhl für Geologie, Geochemie und Lagerstätten des Erdöls und der Kohle, RWTH Aachen.
-
Palynological residues and slides: Geologischer Dienst Nordrhein-Westfalen, Krefeld.
-
Calcareous nannoplankton: at present E. Martini’s private collection; all specimens will be later transferred to the Senckenberg Forschungsinstitut und Naturmuseum, Frankfurt/Main where the holotype of Nannoserratolithus minutus Martini (see below) is already housed (catalogue number SM.B 17726).
-
Foraminifers, ostracods and vertebrate skeletal remains including polished sections of fish teeth: Landesamt für Geologie und Bergbau Rheinland-Pfalz, Mainz.
Results and discussion
Eocene sediments and lithostratigraphy
In the Birkweiler road cut mainly yellowish-greenish clays and marls were exposed. Marlstones, limestones and an organic matter-rich clay horizon with a thin lignite seam are intercalated (Fig. 4). West of the exposure of Lower Jurassic strata the succession starts with an approx. 3 m thick layer of light brown to white clay with numerous reddish brown to black concretions (Fig. 5 a). They consist of hematite, goethite, quartz and kaolinite (“Bohnerz”) while the white clay is composed of kaolinite, quartz and goethite as shown by XRD analyses. The concretions may become so abundant that the clay only fills the space between them. The “Bohnerz” concretions and the kaolinite clay represent a residual terrestrial sediment resulting from weathering under humid, tropical-subtropical conditions since the early Eocene or even earlier. Thus, the light-coloured clay with embedded “Bohnerz” concretions is a ferruginous palaeosol characterised by a ferric to pisoplinthic horizon which developed on Lower Jurassic mudstones and corresponds to a former land surface. This unit is attributed to the Schliengen Formation (= Siderolith-Schichten) which has been recorded in the middle and southern Upper Rhine Graben and adjacent areas where residual soils accumulated in depressions on karstified Jurassic limestones (Pirkenseer et al. 2018). Typically, the clay of the Schliengen Formation consists for the most part of kaolinite (Sittler 1965, 1968b) which is not a main constituent in other stratigraphical units (Grüne Mergel to Landau Formation) of the Upper Rhine Graben (Löschan et al. 2017). A stratigraphically younger unit similar to the Schliengen Formation and consisting of clay rich in limonite concretions is well known from the Mainz Basin where these “Bohnerztone” overlay e.g. limestones and marls of the upper Oligocene (Chattian) to lower Miocene Mainz-Gruppe (Schäfer 2012).

a Thin section of “Bohnerz” concretions of the Schliengen Formation; Birkweiler road cut. b Polished section of the stromatolite horizon; Birkweiler road cut. c Thin section of the stromatolite horizon; Birkweiler road cut. d Bedding plane of grey mudstone with numerous otoliths, fragments of fish bones and gastropods; Birkweiler drainage pit. e Enlargement of red square in 5 d showing ten otoliths; Birkweiler drainage pit. f Thin section of well-rounded stromatolite fragments and a single ooid from fossil rich grey marl; Birkweiler drainage pit (thin section AW4). g–i Thin sections of stromatolite fragments with well-preserved outward radiating tubes and growth zonation pattern; Birkweiler drainage pit (g: thin section AZ4a, h, i: thin section AZ1). j Thin section of sandstone with numerous angular stromatolite fragments (red arrows); Birkweiler drainage pit (thin section AZ4a). k, l Thin section of indurated clastic sedimentary rock mostly composed of angular stromatolite fragments; Birkweiler drainage pit (thin section AZ4b)
West of the layer with numerous “Bohnerz” concretions follows a poorly exposed succession of monotonous yellowish-greenish clays and marls. Unrecognised faulting and landslides may pretend a high thickness. Intercalated are a dark brown organic matter-rich clay horizon with a lignite seam, a very fossiliferous marl horizon (Fig. 4) and some limestone layers including a 50 cm thick stromatolite horizon (Figs. 4, 5 b, c). A sample from the lignite seam yielded an Eocene miospore assemblage (see above). Of special interest is an approx. 1.10 m thick marl layer (Fig. 4) which is extremely rich in hydrobioids (Kadolsky 2015; Kadolsky and Hartkopf-Fröder, in prep.), otoliths, vertebrate bones and teeth. The stromatolites from the Birkweiler road cut are very similar to a stromatolite horizon described by Heidtke (2006: 20, fig. 5) from the Lower Jurassic succession at the base of the brickyard pit west of Siebeldingen. In the fully marine, Lower Jurassic, organic-rich, dark grey mudstones the occurrence of stromatolites is at least unexpected. Schwarz and Griessemer (1998) reported green marls of mid Eocene age and landslides from the area of the former brickyard pit and hence, it is more likely to assume that Heidtke’s stromatolite horizon is in fact part of the middle Eocene succession and not of Early Jurassic age. This assumption is also supported by the fact that from the nearby village Birkweiler a 15 cm thick stromatolite horizon embedded in light greenish grey marls has been described by Bucher (1914: 69–70) and Reis (1923: 117–120) who postulated an Oligocene or Miocene age without further substantiation. For these marls an Early Jurassic age can be excluded as such sediments are unknown from the Siebeldingen brickyard pit and adjacent Lower Jurassic outcrops. In thin sections, the microstructure (e.g. outward radiating tubes, growth zonation; see Reis 1923: pl. 3, fig. 14, pl. 4, fig. 10) of the stromatolites from the Birkweiler village and the Birkweiler road cut are very similar suggesting a widespread occurrence of the distinct stromatolite horizon(s) in that area.
Due to extensive recent and reactivated fossil landslides and complex fault tectonics in this area, it is not reasonable to assume that the entire section is undisturbed and in its original position. In addition, it even cannot be ruled out that parts of the succession include strongly weathered Upper Triassic (Keuper) clays and marls. These sediments have been encountered in boreholes along the building site of the bypass highway B 10 (Schröder 1984; Wichter et al. 1988; re-evaluation of the sediments and microfossil assemblages in boreholes B 703 and B 704 by Peter Schäfer) and are difficult to distinguish from Paleogene/Neogene sediments based on petrographical criteria alone (e.g. Nottmeyer 1954). However, the lignite seam near the base and the fossil rich layer near the top of the succession are all Eocene in age (see below). In addition, four samples from the road cut investigated by Schuler (1990) have been assigned to the middle to upper Eocene Grüne Mergel (“Marnes vertes à limnées = Salifère inférieur”; see also Châteauneuf and Ménillet 2014) and two samples rich in dinocysts to the upper Eocene/lowermost Oligocene (Rupelian) Untere Pechelbronn-Schichten (“Couches de Pechelbronn inférieures = Sel III”; see also Duringer et al. 2019: fig. 7). From the area of the former brickyard pit near the Birkweiler road cut Schwarz and Griessemer (1998) reported Landauer Mergel sensu Breuer and Feist (1986) with charophytes indicative of the Maedleriella embergeri Zone. The recorded charophytes are clearly of Lutetian age. Additional samples from the Landauer Mergel in the Albersweiler area yielded late Eocene and perhaps even early Oligocene charophytes but no further information was provided (Schwarz and Griessemer 1998). Hence, in the Birkweiler road cut section the existence of a substantial amount of sediments other than of Eocene and early Oligocene age is rather unlikely.
The succession encountered in borehole B 804 (Fig. 4) is similar to the section in the Birkweiler road cut except that in the former limestone horizons are more abundant and dark grey sediments are more common indicating less weathered sediments. At least two slip planes have been identified in the borehole (at ca. 4 m and 26 m; Schröder 1984) and hence, the succession must be regarded as disturbed. However, the biostratigraphical data do not imply the presence of other than Eocene sediments below Quaternary overburden. Several samples of the cores from this borehole yielded foraminifer, ostracod, hydrobioid and otolith assemblages including a mass occurrence of hydrobioids and otoliths between 27.80 and 28.00 m. With regard to faunal composition this horizon is very similar to the one in the Birkweiler road cut. Due to landslides and fault tectonics these two horizons cannot be unequivocally correlated.
While in the Birkweiler road cut yellowish-greenish sediments prevail, probably indicating deep weathering enhanced by the mass movements, the marls from the Birkweiler drainage pit are predominantly light grey and medium light grey, in places medium grey and light olive grey in colour. Most of the marls lack macrofossils. However, many specimens of Anomalorbina?, a small-sized planorbid-like gastropod, were found on one bedding plane and specimens of the gastropod family Potamididae are concentrated in few thin layers (Kadolsky and Hartkopf-Fröder, in prep.). Some light grey, marly limestone horizons with disarticulated fish remains on the bedding planes are intercalated as well as up to few centimetres thick layers composed of marl, fine sand and masses of otoliths, hydrobioid fragments and complete hydrobioid shells (Kadolsky 2015; Kadolsky and Hartkopf-Fröder, in prep.) (Fig. 5 d, e). These layers have sharp lower and upper boundaries. After disaggregation of the sediment using 5 % hydrogen peroxide solution the resulting residue is composed of numerous hydrobioids, otoliths, stromatolite fragments, sandstone/clastic sedimentary rock fragments (Fig. 5 g–l) and very rare ooids (Fig. 5 f). In addition, rare hematite/goethite concretions (“Bohnerz”; see above) occur. By far most of the hydrobioids are broken but there are also many complete and well-preserved shells. The gastropod assemblage is of low diversity; each shell-bearing layer is almost monospecific, with the hydrobioid Lutetiella hartkopfi Kadolsky, 2015 being most abundant overall. The stromatolite fragments are mainly in the grain size of medium sand to medium gravel, well rounded and with smooth and even “polished” surface (Fig. 5 f). The stromatolites are well preserved showing outward radiating tubes and growth zonation patterns (Fig. 5 g–i). Dark brown to dark grey staining of the stromatolites is likely attributed to bitumen impregnation. The sandstone also contains numerous small, angular stromatolite fragments (Fig. 5 j). Some indurated clastic sedimentary rocks are mostly composed of stromatolite fragments (Fig. 5 k, l). The small angular stromatolite fragments are probably derived from stromatolites growing right next to the depositional site while the well-rounded fragments were abraded during transport from a more distant source. In either case a high-energy event such as a storm was necessary to break up stromatolite structures. In the Birkweiler drainage pit itself no stromatolites were exposed. However, in the nearby Birkweiler road cut a 50 cm thick stromatolite horizon was encountered (Fig. 4) indicating that proper environmental conditions for stromatolite growth existed during deposition of the Eocene succession.
Based on palynomorphs and calcareous nannoplankton (see below) the investigated sediments of the Birkweiler drainage pit are assigned to the mid Eocene. However, it may be that, due to landslides, the succession in borehole B 804 and the Birkweiler road cut includes middle and upper Eocene and perhaps even lower Oligocene sediments (Schwarz and Griessemer 1998). Lithostratigraphically the succession is assigned to the Landauer Mergel sensu Breuer and Feist (1986), lower part of the Lymnäenmergel, Haguenau-Gruppe, an informal unit which for the first time was used by Breuer and Feist (1986: tab. 2) for a 6 m thick succession of olive grey to green grey, fossiliferous claystones and clayey marlstones in well Landau 117. Validity and range of the Haguenau-Gruppe is still under discussion (Pirkenseer et al. 2018).
Thermal maturity and organic geochemistry
In samples from the Birkweiler drainage pit most laevigate trilete spores are lemon yellow to golden yellow in colour and hence, the SCS (Spore Color Standard of Pearson 1990) is between 2 and 3 on the 11-points gradation colour chart. These values indicate an immature rank with a vitrinite reflectance between 0.3 and 0.4 % VRr and are in good accordance with the measured vitrinite reflectance (see below; for an approximate correlation of the SCS scale and vitrinite reflectance see Marshall and Yule 1999; Hartkopf-Fröder et al. 2015). Some brown spores with SCS around 7 may be reworked.
Ichthyoliths such as the gar teeth may give some information on the thermal alteration of the sediment as they change colour from yellow to black and at very high maturity they become opaque white and finally clear and brittle (ichthyolith alteration index, IAI). Hence, with increasing maturation they follow the well-known colour path of conodonts (CAI; for more details see Hartkopf-Fröder et al. 2015). The gar teeth assemblage from Birkweiler is composed of two distinct colour populations, i.e. greyish orange to moderate yellowish brown and light grey to black. Few teeth show circular bands of both colours. Under fluorescence, most of the teeth cut and polished perpendicular to the longitudinal axis exhibit a yellow to orange fluorescence. Tooth enamel is distinguished from dentine by its stronger, bright yellow fluorescence (Fig. 6 a, b). In some teeth the inner parts of the dentine surrounding the pulp cavity exhibit a weaker fluorescence (dark brown) than in the outer parts. In one tooth no visible fluorescence of the dentine was observed but strong yellow fluorescence of the enamel. Provided that colour change of the ichthyoliths is solely controlled by maturation, the orange/brown specimens would imply a lower thermal maturity than the grey/black coloured ones. As both populations occur in the same sample reworking of those with darker colours might be assumed. While the samples from the Birkweiler drainage pit are thermally immature contemporaneous sediments from the Upper Rhine Graben fill show higher maturity (see below). The darker coloured teeth could therefore be reworked deriving from areas of higher subsidence and heat flow but this would imply, after an incipient maturation, the erosion of sediments bearing this distinct tooth type. However, this scenario is rather unlikely as it requires subsidence, thermal maturation and uplift within a short period. Irrespective of the colour all teeth are well preserved which also contradicts the assumption of reworking and transport. A more obvious explanation for the colour and fluorescence variation of the gar teeth and the occurrence of different colours and fluorescence in the very same tooth specimen takes into account that colonisation by microbes such as fungi and bacteria and subsequent degradation of organic matter causes colour change of the phosphatic fish remains (e.g. Tway et al. 1986; Koot et al. 2013). Hence, differences in colour and fluorescence of the Birkweiler ichthyoliths are here attributed rather to taphonomic processes (e.g. longer exposure times of the teeth before burial, more suitable microhabitat) than thermal alteration and reworking.

a Horizontal cross-sections close to the tip of a Lepisosteidae jaw tooth showing the strongly fluorescent enamel, the radially arranged dentine tubules and the narrowed pulp cavity; Birkweiler drainage pit. b Same specimen as a. Strongly fluorescent enamel (blue arrow), dentine tubules (white arrows), pulp cavity (red arrow). Both photographs incident light fluorescence microscopy using a Zeiss Axio Imager.M2m, a HXP 120C light source and a fluorescence filter set 09 (excitation BP 450–490 nm, beam splitter FT 510 nm, emission LP 515 nm)
Three samples from the Birkweiler drainage pit have been analysed for source rock potential and thermal maturity (Tab. 1).
Total organic carbon (TOC) contents between 0.73 and 1.05 % and hydrogen indices (HI) between 109 and 158 mg HC/g TOC indicate poor oil generation potential and only minor gas generation potential by type III kerogens (Peters and Cassa 1994). The samples from the Birkweiler drainage pit show vitrinite reflectance values of 0.37–0.39 % VRr indicating an immature rank. Tmax values between 427 and 435 °C correspond to an immature to early mature rank and are in good agreement with the VRr values (Peters et al. 2005).
From the Lymnäenmergel in wells of the Landau oil field, Teichmüller (1970, 1979), Doebl et al. (1974) and Teichmüller and Teichmüller (1981) published thermal maturity data ranging between 0.67 and 1.38 % VRr at 1585 to 2063 m depth. Three samples from one of these wells (1902–2036 m) of the Landau oil field have been re-analysed and show considerably lower values between 0.73 and 0.81 % VRr which is equivalent to a mid-oil window maturity (Fig. 7). However, the vitrinite reflectance data from the Lymnäenmergel in this well are still much higher than those from the Birkweiler drainage pit. A logarithmic trend line (Fig. 7) connecting outcrop and data from the wells of the Landau oil field indicates a major subsidence and heat flow related increase in VRr without major transient thermal effects. Maximum palaeotemperatures estimated from vitrinite reflectance data are in the range of ca. 120–130 °C (for 0.8 % VRr) at 2000 m depth in well Landau 2 (Deming 1994).

Vitrinite reflectance of the Landauer Mergel sensu Breuer and Feist (1986) (= lower part of the Lymnäenmergel) from Birkweiler drainage pit and the Lymnäenmergel encountered in wells of the Landau oil field at the western Upper Rhine Graben between Birkweiler and Landau. MD measured depth
Distribution patterns of saturated hydrocarbons in three analysed samples were similar and are exemplarily discussed for sample A 10 from the Birkweiler drainage pit (Fig. 8). In general, the distribution of hydrocarbons indicates mixed sources of the organic matter and is in agreement with low thermal maturity as assessed by Rock-Eval pyrolysis and vitrinite reflectance. n-Alkanes have been detected in the chain length range from C12 – C33 and showed a bimodal distribution with maxima at C17 and C27, respectively (Fig. 8 a). n-Alkanes in the longer chain length range (C23 – C33) displayed a significant odd-over-even predominance as expressed by the Carbon Preference Index CPI23–33 value which is a clear indication for immature terrestrial organic matter (e.g. Marynowski and Zatoń 2010). In contrast, no preference was observed for the alkane distribution around C17 which points to a contribution of marine organic matter. A generally similar n-alkane distribution has been reported for the Callovian (Middle Jurassic) Papartinė Formation of Lithuania (Marynowski and Zatoń 2010). In addition to n-alkanes, the isoprenoidal hydrocarbons pristane and phytane have been detected. Pristane dominated slightly over phytane as expressed by the Pr/Ph ratio which may indicate an oxic to suboxic depositional environment (Peters et al. 2005). Furthermore, the m/z 191 trace revealed the presence of a series of hopanoid hydrocarbons (Fig. 8 b) which can in principle be assigned to bacterial organic matter sources. The distribution pattern of these biomarkers was also in agreement with an immature signature of the organic matter, i.e. a significant value of the ββ/(ββ+βα+αβ) ratio indicates the presence of ββ-hopanes which disappear very early during maturation (Farrimond et al. 1998). Steranes, sterenes and steradienes were insignificant in the investigated samples.

a Total ion chromatogram displaying the distribution of n-alkanes in sample A 10, Birkweiler drainage pit; numbers on peaks indicate the number of carbon atoms in n-alkanes; IS, internal standard (5α-androstane); CPI23–33, 0.5 × (([C23]+[C25]+[C27]+[C29]+[C31]+[C33])/([C22]+[C24]+[C26]+[C28]+[C30]+[C32]) + 0.5 × (([C23]+[C25]+[C27]+[C29]+[C31]+[C33])/([C24]+[C26]+[C28]+[C30]+[C32]+[C34])); Pr/Ph, pristane/phytane ratio. b Selected ion chromatogram (m/z 191) displaying the distribution of hopanoid hydrocarbons in sample A 10; the numbers refer to the number of carbon atoms while Greek letters indicate the configurations at carbon atoms 17 and 21 of the hopane skeleton, respectively; ββ/(ββ+βα+αβ), 17β,21β-hopane/(17β,21β-hopane+17β,21α-hopane+17α,21β-hopane)
Pollen and spores
Six samples (108575, 108579, 108581, 108585, 108586, 108590) from the Birkweiler drainage pit were investigated for pollen, spores and algae (Figs. 9, 10, 11, Electronic Supplementary Material Fig. S1 and Tabs. S1, S2). Preservation of the predominantly yellow pollen and spores is reasonable. Most pollen are compressed, sometimes crumpled and corroded (thinning of exine) and many specimens contain black fragments, probably pyrite crystals.

Miospores from Birkweiler drainage pit. a Baculatisporites sp. (slide 108590 SG 063). b Retitriletes sp. (slide 108590 SG 061). c Cicatricosisporites paradorogensis (slide 108575 SG 003). d Polypodiaceoisporites sp. (slide 108581 SG 010). e Leiotriletes triangulus (slide 108590 SG 062). f Corrugatisporites multivallatus (slide 108581 SG 007). g Verrucatosporites sp. (slide 108575 SG 022). h Laevigatisporites haardti (slide 108596). i Inaperturopollenites dubius (slide 108585 SG 021). j Inaperturopollenites concedipites (slide 108590 SG 059). k Pinuspollenites labdacus (slide 108590 SG 058). l Ephedripites eocenipites (slide 108575 SG 005). m Restioniidites minimus (slide 108575 SG 016). n Graminidites sp. (slide 108585). o Platycaryapollenites lunatus (slide 108575 SG 009). p Plicapollis pseudoexcelsus (slide 108575 SG 017). q Myricipites sp. (slide 108586, with pyrite crystal embedded in the exine). r Polyatriopollenites stellatus (slide 108575 SG 010). s Compositoipollenites rhizophorus (slide 108585 SG 032, Nomarski differential interference contrast). t Malvacipollis sp. (slide 108575). u Arecipites convexus (slide 108575). v Oleoidearumpollenites microreticulatus (slide 108575 SG 020). SG = Single Grain Mount. All photographs were taken using a 100x oil immersion objective. Slides are stored in the palynological collection of the Geological Survey North Rhine-Westphalia, Krefeld

Miospores (a–s) and Botryococcus sp. (t) from Birkweiler drainage pit. a Retitricolpites gracilis (slide 108586). b Ilexpollenites margaritatus (slide 108575 SG 013). c Ericipites callidus (slide 108590 SG 065). d Tricolporopollenites crassostriatus (slide 108590 SG 036). e Tricolporopollenites parmularius (slide 108575 SG 001). f Tricolporopollenites microporitus (slide 108590 SG 034). g Tricolporopollenites pseudocingulum (slide 108575 SG 024). h Scabratricolporites sucii (slide 108585 SG 009). i Pentapollenites pentangulus (slide 108575 SG 008). j Margocolporites vanwijhei (slide 108590 SG 046). k Parthenopollenites marcodurensis (slide 108590 SG 030). l Eocaenipollis sp. 2 sensu Châteauneuf (1980) (slide 108575 SG 007). m Tetracolporopollenites sapotoides (slide 108590 SG 040). n Sapotaceoidaepollenites obscurus (slide 108590 SG 035). o Sapotaceoidaepollenites sculptatus (slide 108585). p Bombacacidites sp. (slide 108575). q Fususpollenites fusus (slide 108585). r Sparganiaceaepollenites sp. (slide 108585). s Nupharipollis sp. (slide 108581). t Botryococcus sp. (incident UVA–blue light fluorescence). SG = Single Grain Mount. All photographs were taken using a 100x oil immersion objective. Slides are stored in the palynological collection of the Geological Survey North Rhine-Westphalia, Krefeld

Stratigraphical ranges of selected pollen and spore taxa from Birkweiler drainage pit in the late Paleocene to early Oligocene of Central Europe (mainly after Lenz 2000). The assemblage suggests a mid Eocene, possibly a mid/late Eocene age. However, as the Mediocolpopollis Group is absent a late Eocene age is less likely. For discussion see text. Numerical ages (Ma) following International Chronostratigraphic Chart, 2022/10 (https://stratigraphy.org/ICSchart/ChronostratChart2022-10.pdf)
The Eocene palynoflora of Germany and France has been described in detail in several comprehensive monographs (e.g. Krutzsch 1959; Châteauneuf 1980; Thiele-Pfeiffer 1988; Schuler 1990; Lenz 2000, 2005). For detailed descriptions, nomenclatural discussions and illustrations we refer to these publications.
Pollen and spores stratigraphy
Identified palynomorph taxa are listed in Electronic Supplementary Material Tab. S1. The assemblage represents a typical Eocene palynoflora of probably mid Eocene age dominated by palaeotropical elements such as Pentapollenites spp., Monocolpopollenites tranquillus (Potonié, 1934) Thomson and Pflug, 1953 ex Jansonius and Hills, 1976, Bombacacidites spp., Compositoipollenites rhizophorus (Potonié, 1934) Potonié, 1960, Plicatopollis plicatus (Potonié, 1934) Krutzsch, 1962 (compare Krutzsch 1967) and characterised by the absence of typical early Eocene taxa, e.g. Thomsonipollis magnificus (Thomson and Pflug, 1953) Krutzsch, 1960 (compare Riegel et al. 2012). Distinctive late Eocene species such as those of the palynostratigraphically important Mediocolpopollis Group (Krutzsch 2008; Hottenrott in Grimm et al. 2011a) were not encountered in the Birkweiler drainage pit material and Pompeckjoidaepollenites subhercynicus (Krutzsch, 1954) Krutzsch in Góczán, Groot, Krutzsch and Pacltová, 1967, already rare in the late Eocene (Borkener Bild, Krutzsch 1966), is still rather common. Plicapollis pseudoexcelsus (Krutzsch, 1957) Krutzsch, 1961, another terminal member of the thermophile Normapolles Group, is typically abundant in the mid Eocene (Krutzsch 1966; Hottenrott 2002) and appears in moderate percentages (max. 5 %) at Birkweiler. Plicatopollis plicatus, a warmth loving species, restricted to the Eocene (Fig. 11) and especially abundant in the upper Ypresian to lower Lutetian Lower and Middle Messel Formation (Thiele-Pfeiffer 1988; Lenz et al. 2007, 2011; Lenz and Wilde 2018), appears regularly. Also typical for the mid Eocene are high percentages of Platycaryapollenites spp. (Juglandaceae) (Krutzsch 1992; Blumenstengel 1993; Blumenstengel et al. 1996). Especially specimens of the Bombacaceae (Bombacacidites spp.) and of Tegumentisporis villosoides (Krutzsch, 1959) Krutzsch, 1963 (e.g. sample 108579) could indicate a mid Eocene age (see Hochuli 1984, fig. 8). T. villosoides was known only from the Lower and Middle Messel Formation (upper Ypresian to lower Lutetian) of Messel (Thiele-Pfeiffer 1988; Lenz and Wilde 2018), the lower Eocene of Grube Prinz von Hessen (Moshayedi et al. 2018) and the middle Lutetian (Schilfkohle, late MP 13/early MP 14 following Haubold 1989) of Geiseltal (Krutzsch 1959). Some forms, e.g. Restioniidites minimus (Krutzsch, 1970) Kedves, 1974 and Ericipites callidus (Potonié, 1931) Krutzsch, 1970, appear regularly since the Eocene (Thomson and Pflug 1953; Krutzsch 1970; Thiele-Pfeiffer 1988). While most of the time-indicative taxa of the Birkweiler drainage pit assemblage point to a mid Eocene age, few species have been reported from the late Eocene only. Eocaenipollis sp. 2 sensu Châteauneuf (1980) seems to occur only in the late Eocene (Priabonian) (Fig. 11). Originally, the species has been described from the Paris Basin where it is rare and the actual stratigraphical range may be inadequately explored so far. The same may be true for Scabratricolporites sucii Schuler, 1990 which has been encountered by Schuler (1990: tab. 54) in the dinocyst-rich Birkweiler assemblage assigned to the Untere Pechelbronn-Schichten (see below). In general, this species is very scarce and may be of limited stratigraphical use. The presence of Margocolporites vanwijhei Germeraad, Hopping and Muller, 1968 in the Birkweiler drainage pit assemblage is noteworthy. The species is well known from other Paleogene basins, e.g. Paris Basin (Châteauneuf 1980). However, in Central Europe it is a rare component. Based on the overall composition of the assemblage and the lack of the Mediocolpopollis Group which is a very important marker for the late Eocene we assign the Birkweiler drainage pit association to the mid Eocene. A late Eocene age is less likely.
Schuler (1990: 210–211) studied eight samples from the road cut near Birkweiler but she neither published data on the exact locality nor petrography of the sediment. Detailed palynological data were provided for five samples and two rather similar palynomorph assemblages were distinguished. Four samples yielded a palynomorph assemblage composed of angiosperm pollen, few gymnosperm pollen, trilete spores and green algae. In one sample also rare dinocysts were encountered. The palynomorph assemblage of these four samples was assigned to the Grüne Mergel (“Marnes vertes à limnées = Salifère inférieur”) by Schuler (1990). Based on one sample and an additional one of which no species list is provided, a second rather similar assemblage was defined by her mostly differing in that it is rich in monospecific dinocysts (Spinidinium). Unfortunately, no specimens of Spinidinium were illustrated by Schuler (1990). Some species of Spinidinium resemble Apectodinium, but Spinidinium differs by having a hexa 2a plate, in contrast to the quadra 2a plate in Apectodinium. This distinctive feature may not always be easily observed. Most records and most species of Spinidinium are from Cretaceous–Paleocene, but a few species of Spinidinium range to late Eocene (Sluijs et al. 2009).
No specimens of Spinidinium were encountered in the present study (see below), but monotypic Apectodinium was recorded in one of our samples (see Dinoflagellate cysts (dinocysts) and green algae). We suspect that Schuler’s (1990) record may similarly be of monospecific Apectodinium, not of Spinidinium.
The presence of abundant dinocysts suggests a marine ingression and hence, Schuler (1990) ascribed this assemblage to the Untere Pechelbronn-Schichten (“Couches de Pechelbronn inférieures = Sel III”; see also Rauscher and Schuler 1988; Duringer et al. 2019: fig. 7). The Untere Pechelbronn-Schichten are of late Eocene/earliest Oligocene age (Grimm et al. 2011b). However, already Sittler (1969) reported dinocysts from the “Marnes calcaires grises à anhydrite et sel” which are equivalent to the Untere Grüne Mergel sensu Grimm et al. (2011b). With regard to the pollen and spores composition Schuler’s dinocyst-rich sample differs from those assigned to the “Marnes vertes à limnées” mainly by the occurrence of Scabratricolporites sucii and the lack of representatives of the Bombacaceae, both being very rare in the studied samples. The assignment to the Untere Pechelbronn-Schichten by Schuler (1990) which is solely based on abundant dinocysts of only one genus, therefore, seems not to be conclusive. Unfortunately, neither geographical coordinates nor a log including the sample position were published by Schuler (1990) so that it is unknown if the two dinocyst-rich samples were collected close to those assigned to the Grüne Mergel (“Marnes vertes à limnées = Salifère inférieur”) or derive from more distant localities within the course of the road cut near Birkweiler. These two samples, therefore, might come from a somewhat younger level of the Grüne Mergel (= Lymnäenmergel). Alternatively, Châteauneuf and Ménillet (2014) suggested that the Spinidinium dinocysts are reworked from Upper Cretaceous sediments.
Vegetation history
For a palaeoecological analysis the Restionaceae, represented by Restioniidites hungaricus (Kedves, 1965) Elsik, 1968, are included here to the land pollen sum (Electronic Supplementary Material Fig. S1) since modern members of the family, e.g. in southern Africa, occupy also terrestrial habitats (Moline and Linder 2005 and references therein). Krutzsch (1992) suggests that Restionaceae during the Paleogene were growing in coastal regions and even in dry habitats, e.g. on sandy substrates. However, they are often considered as representing marsh vegetation during the Eocene (see Lenz and Riegel 2001; Lenz et al. 2021). At Birkweiler Restioniidites spp., especially Restioniidites hungaricus, reaches > 25 % in three samples (108575, 108581, 108586). Sample 108586 is dominated by Restionaceae (> 65 %). Sparganiaceaepollenites spp. (Sparganiaceae, Typhaceae), moderately common at Birkweiler, are a typical swamp element which forms reed belts. Often Sparganiaceae appear together with Restionaceae in Eocene freshwater marshes (Lenz and Riegel 2001). Botryococcus, a colonial freshwater alga tolerating brackish water conditions (Guy-Ohlson 1992; Banerjee et al. 2002), is especially abundant when Restionaceae are most common, pointing to high water tables. Other algal remains, e.g. Ovoidites sp. (botanical affinity: Zygnemataceae; Rich et al. 1982) are rare but most of the samples contain chorate and also proximate dinocysts, which are generally more common when Restionaceae and Botryococcus are abundant. An exception is sample 108581, where dinocysts are extremely rare.
The most prominent arboreal elements are Inaperturopollenites concedipites (Wodehouse, 1933) Krutzsch, 1971 (botanical affinity: Taxodiaceae) and I. dubius (Potonié and Venitz, 1934) Thomsen and Pflug, 1953 (Cupressaceae) which point to locally wet conditions since many species of these families are nowadays dwellers of peat-forming swamp forests (see Schuler 1990; Lenz 2000, 2005). Plicapollis pseudoexcelsus and Pompeckjoidaepollenites subhercynicus probably belong to the Juglandaceae and are associated with both Restionaceae and Sparganiaceae. Therefore, they might be members of a marsh community, maybe growing on hammocks together with palms, as suggested at the middle Eocene site Helmstedt (Northern Germany, Lenz 2000; Lenz and Riegel 2001). On the other hand, an affinity to Rhoipteleaceae is suggested for Plicapollis (Krutzsch in Góczán et al. 1967; Krutzsch 1992). Plicapollis pseudoexcelsus and Pompeckjoidaepollenites subhercynicus rarely show percentages > 5 % at Birkweiler. Palm (Arecaceae) pollen, generally pointing to rather tropical, humid conditions, are regularly encountered at Birkweiler with taxa such as Arecipites convexus (Thiergart, 1938) Krutzsch, 1970 and Monocolpopollenites tranquillus, another typical Eocene element (Hottenrott 2002). Arecipites convexus reaches approx. 20 % in sample 108581 when also Myricipites pseudorurensis (Pflug in Thomson and Pflug, 1953) Grabowska and Ważyńska, 2009 (botanical affinity: Myricaceae), another element probably growing on hammocks in marsh land (Lenz and Riegel 2001), is common together with Restionaceae and Botryococcus. Triatriopollenites rurensis Pflug and Thomson in Thomson and Pflug, 1953 was also probably produced by Myricaceae and is here associated with Restionaceae. Bombacaceae (Bombacacidites spp.) and Icacinaceae (Compositoipollenites rhizophorus (Potonié, 1934) Potonié, 1960) are also considered marsh plants in the middle Eocene sediments of Helmstedt (Lenz 2000).
It is not clear, if the few pollen grains of Poaceae (Graminidites spp.) were produced by swamp elements of the grass family. Generally, grass pollen grains are rare until the Oligocene (Frederiksen 1985; Krutzsch 1992). Cyperaceae, also rare at Birkweiler, substitute Restionaceae as a humid element since the Oligocene (Krutzsch 1992). Direct evidence for mangrove vegetation, e.g. occurrence of Avicennia sp. or Rhizophora spp. (fossil pollen genus Zonocostites spp.), is missing at Birkweiler, but many Polypodiaceae, appearing at Birkweiler, might be associated with mangrove conditions. Although species of the genus Leiotriletes were not consistently differentiated in this study, some specimens of Leiotriletes adriennis (Potonié and Gelletich, 1933) Krutzsch, 1959 were identified. Some authors (e.g. Akkiraz et al. 2006) affiliate the taxon with the mangrove fern Acrostichum whereas others suggest a relationship to tropical Lygodium spp. (Schizaeaceae, see Dašková et al. 2011).
Drier, well-drained habitats are marked by the appearance of Juglandaceae (Plicapollis pseudoexcelsus, Platycaryapollenites spp.) as well as rare Pinuspollenites spp., e.g. Pinuspollenites labdacus (Potonié, 1931) Raatz, 1937 ex Potonié, 1958 and Pinuspollenites minutus (Zaklinskaya, 1957) Nagy, 1985 (see Châteauneuf 1980; Frederiksen 1985; Lenz et al. 2011). All those elements appear regularly, but never exceed 5 %, probably pointing to regional pollen transport from a less humid hinterland vegetation. Wilkinson et al. (1980) interpret pollen produced by Pinaceae (Pinuspollenites spp.) as an element of upland regions. Tricolporopollenites parmularius (Potonié, 1934) Krutzsch in Krutzsch, Pchalek and Spiegler, 1960, produced by Eucommiaceae, reaches up to 5 %. Today, Eucommia ulmoides Oliver, 1890 grows naturally in South China in hilly areas from 200 m to 1700 m above sea level where spring temperatures vary from 12.3 °C to 20.1 °C (Yu-Fei Wang et al. 2003). Pollen of a large number of paratropical trees, e.g. Sapotaceae (Sapotaceoidaepollenites spp., Tetracolporopollenites spp.), Myrtaceae (Cupanieidites spp., see Châteauneuf 1980), Caesalpiniaceae (Margocolporites vanwijhei Germeraad, Hopping and Müller, 1968, see Châteauneuf 1980), or Engelhardia (Momipites spp., see Lenz 2000) appear regularly. Whereas Sapotaceae, many Myrtaceae and Engelhardia might be indicative of wet, subtropical/tropical habitats, Caesalpiniaceae are adapted to hot climates with a distinctive season (see Brasero et al. 2009; Worobiec 2009). Engelhardia nowadays needs mean annual temperatures of minimally approx. 16 °C (Akkiraz et al. 2006).
Evergreen climbers were also present with taxa such as Parthenopollenites marcodurensis (Pflug and Thomson in Thomson and Pflug, 1953) Traverse, 1994 (botanical affinity: Parthenocissus, Worobiec 2009). Temperate or cosmopolitan families, e.g. Ulmaceae (Ulmipollenites undulosus Wolff, 1934), Ericaceae (Ericipites spp.) or Aquifoliaceae (Ilexpollenites spp.), are underrepresented at Birkweiler. Some taxa, e.g. Eocaenipollis sp. or Pentapollenites pentangulus (Pflug, 1953) Krutzsch, 1957, have a questionable botanical affinity but are characteristic for the Eocene and have a thermophilic character (Châteauneuf 1980; Schuler 1990; Lenz 2000). Gnetales are represented by few pollen grains of Ephedripites eocenipites (Wodehouse, 1933) Krutzsch, 1961, which is during the Eocene commonly associated with moist tropical and subtropical vegetation (Frederiksen 1985 and references therein).
Generally, a paratropical arboreal vegetation with a high quantity of wetland elements is suggested.
Schuler (1990: 210–211) identified similar pollen and spores spectra also pointing to a mid to late Eocene age. Samples were dominated by angiosperms (79–96 %) whereas pollen of Pinaceae (Pityosporites spp.) fluctuated between 2–11 % and Cupressaceae reached 3 % (Schuler 1990). This is in contrast to the current study where Cupressaceae and Taxodiaceae are dominant trees, clearly pointing to a massive influence of swamp forest elements (Page 1990). It is typical that Myricaceae, a family of approximately 50 shrub and tree species often today growing near streams and in swamps (Fitzgerald 1999), are common elements in our study, but much less abundant in Schuler (1990). Schuler (1990) reports higher percentages of Flagellariaceae (Milfordia spp. and Restioniidites hungaricus in this study) only for one sample, whereas percentages of marshy elements like Restionaceae and Sparganiaceae are much more abundant in samples from the Birkweiler drainage pit. We suggest that, although both studies deal with contemporaneous deposits, Schuler (1990) investigated a palynoflora pointing to a better drained, mesic habitat. In both studies Pinaceae, as an element of the hinterland, but also Gnetales are minor constituents whereas angiosperms typical for an Eocene rainforest, e.g. belonging to Sapotaceae, the Normapolles Group, Bombacaceae, Icacinaceae, are detected regularly by Schuler (1990) but are more diverse in our study. Presumably, cryptogams did not play a major role in both ecosystems.
Dinoflagellate cysts (dinocysts) and green algae
The same six samples as studied for pollen and spores were investigated for dinocysts. Table S3 (Electronic Supplementary Material) presents the dinocyst taxa recorded from the Birkweiler drainage pit and Figs. 12–14 show photographs of the dinocysts. Sample 108581 is essentially barren of dinocysts with only two specimens seen. The remaining five samples are relatively rich in dinocysts. Preservation is moderate to poor.

Dinocysts from Birkweiler drainage pit. a Apectodinium homomorphum (slide 108590/1). b Apectodinium homomorphum (slide 108579/1). c Apectodinium homomorphum (slide 108590/1). d Apectodinium homomorphum (slide 108590/1). e Apectodinium parvum – quinquelatum transitional (slide 108575/1). f Apectodinium parvum (slide 108590/1). g Apectodinium parvum (slide 108575/1). h Apectodinium parvum (slide 108575/1). i Apectodinium parvum – quinquelatum transitional (slide 108575/1). j Apectodinium quinquelatum (slide 108586/1). k Apectodinium quinquelatum (slide 108586/1). l Apectodinium quinquelatum (slide 108586/1). m Apectodinium quinquelatum (slide 108575/1). n Apectodinium quinquelatum (slide 108586/1). o Apectodinium quinquelatum (slide 108586/1). p Lejeunecysta hyalina (slide 108579/1). q Lejeunecysta hyalina (slide 108590/1). r Phthanoperidinium echinatum (slide 108579/1). s Phthanoperidinium echinatum (slide 108579/1). t Same specimen as s. All photographs were taken using a 100x oil immersion objective. Slides are stored in the palynological collection of the Geological Survey North Rhine-Westphalia, Krefeld

Dinocysts from Birkweiler drainage pit. a Polysphaeridium zoharyi (slide 108590/1). b Polysphaeridium zoharyi (slide 108590/1). c Polysphaeridium zoharyi (slide 108590/1). d Unknown dinocyst sp. 1 (slide 108590/1, low focus). e Same specimen as d (middle focus). f Same specimen as d (high focus). All photographs were taken using a 100x oil immersion objective. Slides are stored in the palynological collection of the Geological Survey North Rhine-Westphalia, Krefeld

Dinocysts from Birkweiler drainage pit. a Phthanoperidinium geminatum – P. regale group, uncertain identification (slide 108579/1). b Phthanoperidinium geminatum – P. regale group, uncertain identification (slide 108579/1). c Phthanoperidinium comatum (slide 108585/1). d Phthanoperidinium comatum (slide 108585/1). e Phthanoperidinium resistente (slide 108585/1). f Phthanoperidinium resistente (slide 108585/1). g Selenopemphix nephroides (slide 108579/1). All photographs were taken using a 100x oil immersion objective. Slides are stored in the palynological collection of the Geological Survey North Rhine-Westphalia, Krefeld
The dinocyst assemblages are similar in all samples. Diversity is very low, with only twelve species in total recognised in the samples (Electronic Supplementary Material Tab. S3). Except for a single specimen “Unknown dinocyst species 1” the dinocysts are well known from Eocene sections outside of the Upper Rhine Graben. No reworking is observed.
In addition to dinocysts the freshwater green algae Pediastrum and Botryococcus were also recorded. Pediastrum is included in Electronic Supplementary Material Tab. S3. Botryococcus is common in all samples, but is badly preserved and was not included in the counting and is omitted from Tab. S3.
Dinocyst taxonomy notes
Taxonomy follows Fensome et al. (2019).
The genus Apectodinium is represented by three species, A. homomorphum (Deflandre and Cookson, 1955) Lentin and Williams, 1977, A. parvum (Alberti, 1961) Lentin and Williams, 1977 and A. quinquelatum (Williams and Downie, 1966) Costa and Downie, 1979, which are all intergrading in morphology (Fig. 12 a–o). Cysts with incompletely developed processes are present. Such specimens probably represent failure to encyst completely (Denison 2021). Some other specimens of A. quinquelatum and A. parvum show a more or less clear expression of paratabulation. Such forms have previously been recorded in assemblages rich in Apectodinium (e.g. Châteauneuf and Gruas-Cavagnetto 1978; Heilmann-Clausen 1985). They conform with Axiodinium tesselatum (Châteauneuf and Gruas-Cavagnetto, 1978) Williams et al., 2015 (originally a subspecies of A. homomorphum) which is here considered as an ecophenotypic variation within Apectodinium.
Due to the rather poor preservation, it cannot be excluded that some specimens registered here as Apectodinium should instead be referred to Trivalvadinium formosum Islam, 1983a, an ill-known taxon resembling Apectodinium homomorphum except for apparently having a 3I archaeopyle. 3I archaeopyles could not be identified with certainty in any specimens in the present study. T. formosum has previously been recorded in marginal marine–lagoonal samples rich in Apectodinium from the late Ypresian–Lutetian of southern England (Islam 1983a, b). As noted by Islam (1983a), the two genera are probably related and favour a brackish water environment.
Phthanoperidinium comatum (Morgenroth, 1966) Eisenack and Kjellström, 1972 includes specimens with a reduced number of very thin spines which may also be rather short (possibly due to poor preservation). However, typical cysts with more numerous spines are also present in the material (Fig. 14 c, d).
Phthanoperidinium resistente (Morgenroth, 1966) Eisenack and Kjellström, 1972 and P. echinatum Eaton, 1976 are morphologically intergrading (Figs. 12 r–t, 14 e, f).
Two specimens in sample 108579 are assumed to either represent Phthanoperidinium geminatum Bujak in Bujak et al., 1980 or P. regale Bujak, 1994 which only differ in the type of archaeopyle (Fig. 14 a, b). As the specimens did not reveal the archaeopyle, they are grouped together in a Phthanoperidinium geminatum – P. regale group. Their shared diagnostic feature is the presence of distinctive simulate zones of ornament. However, less developed simulate zones may occur in other species (Bujak et al. 1980: 74). As only two specimens have been found, the identification is somewhat uncertain.
Polysphaeridium zoharyi (Rossignol, 1962) Bujak et al., 1980, in addition to typical specimens (Fig. 13 a–c), includes cysts with very thin, almost massive processes. They are considered to represent ecophenotypic varieties.
Palaeoecology
The most striking aspect of the dinocyst assemblage from Birkweiler is the very low diversity with only twelve species present (some of which are intergrading, cf. taxonomic notes). The gonyaulacoids, a major, mainly autotroph dinoflagellate group, is only represented by Polysphaeridium zoharyi. The numerous absent gonyaulacoid genera are associated with a wide range of neritic settings. Normally some of these genera would be represented in any neritic assemblages, at least in very small numbers, due to mixing of motile cells or cysts by currents, storms, reworking and time averaging. The missing gonyaulacoid genera include first and foremost the widespread, common Spiniferites group ‘generalists with open marine affinity’ (Frieling and Sluijs 2018) and the Areoligera – Glaphyrocysta group usually assumed to have lived in more proximal environments with normal salinity waters (e.g. Frieling and Sluijs 2018). Further notable absences include among others, Cleistosphaeridium, Operculodinium, Cordosphaeridium, Areosphaeridium and Thalassiphora.
The total absence of these genera suggests that the Birkweiler site was a completely landlocked water body, a scenario which would prevent the mixing of associations.
Except for Polysphaeridium zoharyi, the dinocysts present at Birkweiler are peridinoids, a group which today includes many heterotrophs. The proportion of heterotrophs is likely related to the production of prey in the surface waters and thus nutrient supply (Frieling and Sluijs 2018). The dominating cysts at Birkweiler, Apectodinium and Phthanoperidinium, are commonly considered to have been heterotrophs (cf. discussions in Sluijs and Brinkhuis 2009; Frieling and Sluijs 2018). The total dominance of peridinoids thus suggests a setting enriched in food/nutrients. Further ecological signals of individual taxa are given below.
Apectodinium (Costa and Downie, 1976) Lentin and Williams, 1977
Apectodinium is abundant in all samples (20–99 %, average 56 %) (Electronic Supplementary Material Tab. S3), except for sample 108581. This genus has been known since the 1970s for its abundance at the Paleocene–Eocene transition (e.g. Downie et al. 1971; Costa and Downie 1976; Harland 1979). It was later recognised that Apectodinium formed strong nearly global acmes on shelves during the hot initial Eocene PETM (Paleocene–Eocene Thermal Maximum; Bujak and Brinkhuis 1998; Crouch et al. 2001; Denison 2021). Palaeoecologically, Apectodinium was favoured by high temperatures and does not occur in great abundance below 20 °C (Frieling et al. 2014). It also seems to be favoured by coastal proximity, increased nutrient levels and freshwater input (Sluijs and Brinkhuis 2009; Sluijs et al. 2014; Frieling and Sluijs 2018). This preference seems to agree with the fact that most acmes of Apectodinium (except for the worldwide distribution during the PETM) are in lagoonal or estuarine environments (see below). The PETM distribution may be linked to increased nutrient input to shelves by rivers as predicted by circulation models (cf. discussion in Sluijs et al. 2007). Heilmann-Clausen (2020) discussed post-PETM examples from the Eocene of the North Sea Basin of strong (approx. 40–99 %) local Apectodinium acmes developed in brackish lagoons and some other very near-shore environments, while Apectodinium is rare or absent in coeval offshore sediments 600–700 km away. Apparently, these strong acmes were able to pop up very locally, whenever environmental conditions were optimal.
Phthanoperidinium Drugg and Loeblich Jr., 1967
Phthanoperidinium is abundant (16–66 %, average 34 %) except for the essentially dinocyst-free sample 108581. According to Sluijs et al. (2005) and Pross and Brinkhuis (2005) abundant Phthanoperidinium may in some cases be tied to high productivity in relatively shallow deltaic settings. In other cases, abundant Phthanoperidinium may indicate offshore upwelling areas. According to Frieling and Sluijs (2018) Phthanoperidinium is typically considered to be tolerant to low salinity.
Polysphaeridium zoharyi (Rossignol, 1962) Bujak et al., 1980
Polysphaeridium zoharyi is fairly common to abundant (1–19 %) in four of the five dinocyst-bearing samples. In contrast to all other species in the assemblages, P. zoharyi is the cyst of an extant dinoflagellate species. The motile stage of P. zoharyi, Pyrodinium bahamense Plate, 1906, mainly lives in equatorial lagoons and is tolerant to strong salinity fluctuations (e.g. Dale 1996; Pross and Brinkhuis 2005). P. zoharyi belongs to a group of dinocysts (epicystal Goniodomidae) which in the Paleogene have been particularly associated with seasonally elevated salinity levels (Frieling and Sluijs 2018).
Taken together, the Birkweiler dinocysts point to a completely landlocked water body with warm, nutrient-rich, brackish waters. Lagoons with fluctuating, seasonally high salinities may have been present.
The common occurrence of Pediastrum (14 %, cf. Electronic Supplementary Material Tab. S3) in sample 108575, and Botryococcus in all samples indicates significant influx of fresh water, at least temporarily.
Stratigraphical correlations and probable age
Due to the similarity of the dinocyst assemblages, the correlations and the age of the samples are discussed for all samples together.
Oldest possible age
Phthanoperidinium geminatum Bujak in Bujak et al., 1980 – P. regale Bujak, 1994 group
The oldest of these two species is P. regale with a lowest occurrence in the Norwegian-Greenland Sea near base of NP15b and near base of Chronozone C20r (Eldrett et al. 2004). The inception in Denmark is likewise in NP15 (Heilmann-Clausen and van Simaeys 2005: 195) and may be already in C21n, and thus slightly older than in the Norwegian-Greenland Sea, cf. discussion in King et al. (2016: 596). The influx of P. regale is a key-event defining Zone DE13 of King et al. (2016).
Relying on this group, the oldest possible age may therefore be mid Lutetian, early part of Chron C20r, or Chron C21n. It is important to note, however, that this group is only tentatively identified (cf. Dinocyst taxonomy notes).
Phthanoperidinium comatum (Morgenroth, 1966) Eisenack and Kjellström, 1972
Phthanoperidinium comatum is a well-known, distinctive dinocyst with a first occurrence in NW Europe in the late Ypresian (first occurrence in early Zone NP13, as discussed in King et al. 2016: 594). Numerous typical specimens were seen in the Birkweiler samples, and it is therefore rather certain that the samples cannot be older than early NP13.
Selenopemphix nephroides Benedek, 1972
The first occurrence of Selenopemphix nephroides is recorded in the North Sea Basin a little earlier than P. comatum, but still in the late Ypresian. It was recorded in Belgium in the Sables de Mons-en-Pévèle/Egem Member by Islam (1982), just below the lowest occurrence of the important marker Areosphaeridium diktyoplokum (Klumpp, 1953) Eaton, 1971, a level dated to uppermost NP12 (cf. King et al. 2016: 206). Heilmann-Clausen and Costa (1989) recorded S. nephroides from a similar level, just above the lowest occurrence of A. diktyoplokum in the Wursterheide borehole, northern Germany. However, Sluijs et al. (2009) recorded and illustrated S. nephroides from the PETM interval i.e. in the basal Ypresian, in the Bass River core, New Jersey. This species thus seems to have a very sporadic record, and is less reliable for dating.
Phthanoperidinium echinatum Eaton, 1976
The lowermost record of Phthanoperidinium echinatum is in early Ypresian in Belgium, at the base of the Dracodinium simile Zone (De Coninck 1991). Lowermost record in northern Germany seems to be in the mid Ypresian (Köthe 1990) and likewise in southern England (at base of the Bracklesham Beds, Eaton 1976).
Youngest possible age
Apectodinium spp.
The most dominating element in the Birkweiler assemblages is the genus Apectodinium with the closely related and intergrading species, A. homomorphum, A. quinquelatum and A. parvum. The genus is even monotypic in sample 108586. As mentioned above, this intergrading complex is well known especially from the initial Eocene PETM interval. However, Apectodinium also developed strong, even monotypic, associations very locally in brackish, paralic environments in the North Sea Basin at later times during the Ypresian and Lutetian, while it is absent, or nearly so, in coeval fully marine environments (Heilmann-Clausen 2020; see Palaeoecology). This raises the question: Can this genus have survived in very marginal settings, at later Eocene times than indicated from literature? The knowledge of dinocyst assemblages in well-dated brackish lagoons through geological time is limited. Bearing this uncertainty in mind, a survey of late Apectodinium records in NW Europe may still yield the best available information on the youngest possible age of the Birkweiler assemblages and will therefore be presented below.
Southern England
In southern England Apectodinium homomorphum ranges continuously up to the top of the Barton Clay according to Costa and Downie (1976). An isolated occurrence is indicated by these authors in the middle Priabonian Middle Headon Beds, but Apectodinium was not recorded in the 13 samples from the Middle Headon Beds studied by the same authors, in Liengjarern et al. (1980), so the last reliable record of A. homomorphum in southern England is in the late Bartonian. In a more detailed range chart of the Bartonian part of the same succession Bujak et al. (1980) indicated the top of A. homomorphum at a slightly lower level, in the middle of their Zone BAR-3 which is dated to the mid Bartonian (King et al. 2016: 421). No details on the abundance are given in these sources, and no other species of Apectodinium were recorded.
In the same area Eaton (1976) recorded sporadic A. homomorphum (which may include A. quinquelatum, by then ranked as a subspecies of A. homomorphum) in the Lutetian to earliest Bartonian.
Islam (1983a) recorded a strong acme of up to 40 % A. homomorphum (as the junior synonym A. folliculum Islam, 1983a) in a lower Lutetian lagoonal unit of the Earnley Formation at Bracklesham Bay. Similar observations of very abundant A. homomorphum were mentioned from some horizons of the upper Ypresian–Lutetian Bracklesham Beds at Alum Bay (Costa and Downie 1976). According to these authors A. homomorphum becomes dominant over A. quinquelatum in the late Ypresian and A. quinquelatum is only sporadically present in the Bracklesham Beds.
Northern France
In the Paris Basin Châteauneuf (1980) recorded common A. homomorphum in the ‘Lutétien supérieur–base de l'Auversien’, i.e. late Lutetian–early? Bartonian (King et al. 2016: 224). It is monotypic (90–98 %) in two samples from the basal ‘Auversien’ Horizon de Mont-Saint-Martin considered to be deposited in a lagoonal-continental or deltaic setting (Châteauneuf 1980: 149, 274). The Horizon de Mont-Saint-Martin is dated to the late Lutetian by King et al. (2016: 225). This is the only record of A. homomorphum in the mid Eocene–early Oligocene of the Paris Basin studied by Châteauneuf (1980).
No other species of Apectodinium were recorded by Châteauneuf (1980), except for rare A. parvum (as A. cf. parvum) in two sections of the lower Rupelian Marnes à Huîtres. It should be noted that, according to Châteauneuf (1980), there is no sign of reworking in these very young samples with A. parvum.
Northern Germany
In the Gartow borehole A. homomorphum is common to abundant in the middle of the mid Ypresian Subzone D8nb while it is absent above (Köthe 1990). In their integrated study of numerous other North German boreholes Köthe and Piesker (2007) indicated the tops of A. homomorphum and A. quinquelatum in Subzone D9na, which is near the Ypresian–Lutetian boundary according to King et al. (2016). No other species of Apectodinium ranged that high.
In the Wursterheide borehole the highest consistent occurrence of A. homomorphum is at 519 m in Zone D9 in which it accounts for 1–4 % of the dinocysts in several samples (Heilmann-Clausen and Costa 1989). This level is dated to late Ypresian by King et al. (2016). Sporadic and rare A. homomorphum continues up to 399.8 m, corresponding to early Lutetian. A. quinquelatum is very sporadic and rare up to earliest Lutetian.
To summarise, in northern Germany A. homomorphum is fairly common at a few late Ypresian levels and sporadic and rare in the early Lutetian. A. quinquelatum is very sporadic and rare in the earliest Lutetian.
Belgium
The Ypresian–Lutetian dinocyst succession is well known from Belgium due to several detailed studies by De Coninck (e.g. De Coninck 1976, 1977, 1985, 1991). Steurbaut et al. (2015) analysed a transitional Ypresian–Lutetian section in the Zemst Borehole for calcareous nannofossils, foraminifers and dinocysts. They discussed in particular the youngest occurrences of Apectodinium based on the Zemst section and previously studied sections. The following picture emerges from their discussion: Apectodinium is represented only by two species, A. homomorphum and A. quinquelatum, in the late Ypresian. It is frequent to abundant from the top of the Panisel Sand Member to the top of the Merelbeke Clay Member (i.e. from latest NP12 to mid NP13), with acmes of up to 14 % and 20 % at various levels at various localities. In all Belgian sections a major decrease of Apectodinium is observed from base of Pittem Clay (mid NP13) and upwards, as discussed in Steurbaut et al. (2015). This may be illustrated by the middle Lutetian Sands of Lede. Here De Coninck (1985) found < 1 % A. homomorphum in two of the four studied samples and < 1 % A. parvum in a third. These rare records may possibly be reworked. On the other hand, they are rather consistent with the data from northern Germany.
Western Alpine region, NW Tethyan Realm
In Haute-Savoie, France, the highest record of A. quinquelatum is in the Ypresian NP11 Zone. A. homomorphum and A. hyperacanthum (Cookson and Eisenack, 1965) Lentin and Williams, 1977 occur in the Lutetian and A. homomorphum also in the Priabonian (isolated record) (Jan du Chêne et al. 1975). In flysch from Switzerland A. homomorphum and A. quinquelatum occur in NP13, and A. homomorphum also in late NP14 and NP15 (Lutetian) (Jan du Chêne 1977).
Unspecified region
Costa and Downie (1979) stated that the last surviving species of Apectodinium are A. quinquelatum and A. homomorphum which survived to the late Eocene and early Oligocene, respectively. No details on these young records were given, however, and they are not taken into account in the present discussion.
To summarise the data on Apectodinium: For A. homomorphum the youngest reliable records are mid or late Bartonian. For A. quinquelatum the last reliable occurrences are somewhere in the Lutetian.
Phthanoperidinium echinatum Eaton, 1976
The youngest consistent records of Phthanoperidinium echinatum are mid or late Bartonian, as outlined below.
In a study of the lower and middle Eocene in southern England Bujak et al. (1980) recorded the last occurrence of P. echinatum within Zone BAR-3. The zone spans Beds E and F of the Naish Member and is dated to the mid Bartonian Chron C18n and NP17 (King et al. 2016: 421, fig. 152). P. echinatum was not recorded in the late Eocene of the same region studied by Liengjarern et al. (1980), thus supporting a Bartonian last occurrence.
In the north German Gartow borehole, the uppermost occurrence of P. echinatum is near top of Zone D10 (where it is common in one sample) (Köthe 1990). This level is dated to mid Bartonian (King et al. 2016: 25), and thus roughly coeval with the findings from England.
In Belgium De Coninck (1985) recorded P. echinatum in all four examined samples from the middle Lutetian Sands of Lede in the Mol borehole (common, with 4 % and 6 % in two of the samples). Rare P. echinatum (0.1–0.3 %) were recorded in six out of eight analysed samples from the Priabonian in the Dutch Woensdrecht borehole (De Coninck 1986).
In the Belgian Kallo borehole De Coninck (1995) found rare P. echinatum in the three Bartonian samples, but none in the five Priabonian samples (age of samples according to King et al. 2016).
In conclusion, in Belgium–the southern Netherlands P. echinatum is common in parts of the Lutetian, rare but consistent in the Bartonian and rare and inconsistent in the Priabonian.
P. echinatum was not recorded in the mid and late Eocene in Denmark (Heilmann-Clausen and van Simaeys 2005), nor in the late Lutetian–Oligocene of the Paris Basin studied by Châteauneuf (1980), although this author noted (p. 145) very rare Phthanoperidinium cf. echinatum in the ‘Auversien’ (i.e. late Lutetian/early Bartonian). This record is not included in his distribution chart, but a rare record in a single sample from the Rupelian is shown (p. 267).
Phthanoperidinium geminatum Bujak in Bujak et al., 1980 – P. regale Bujak, 1994 group
In the Phthanoperidinium geminatum – P. regale group P. geminatum ranges highest. The combined evidence shows that P. geminatum ranges to the mid Priabonian, as outlined below.
P. geminatum was established by Bujak in Bujak et al. (1980) and according to these authors the range in southern England is from Zone BAR-1 to the top of the studied interval (top of Zone BAR-5), approximately matching the Bartonian Stage (cf. King et al. 2016).
In the southern Netherlands P. geminatum is present in the Woensdrecht borehole in most samples up to the middle of the Priabonian Bassevelde Sands (De Coninck 1986, 1995). Similarly, in the Belgian Kallo borehole it is present in one Bartonian sample and in two out of three samples from the Bassevelde Sands, accounting for 1 % near base of this unit (De Coninck 1995).
In Denmark P. geminatum is very rare in two Lutetian samples from the Kysing-4 borehole, but consistent and more common in the late Bartonian and early Priabonian Chron C17n–C17r interval. P. geminatum is absent above, except for a single middle Priabonian sample from Chron C16n.1n and NP19/20 (at approx. 15 m). Here it accounts for 4 % and is presumably in situ (dinocyst data from Heilmann-Clausen and van Simaeys 2005; calcareous nannofossil- and magnetostratigraphy from Thomsen et al. 2012).
Stratigraphical conclusion
It seems certain, that the Birkweiler samples cannot be older than late Ypresian, Zone NP13 (based on Phthanoperidinium comatum). The Phthanoperidinium geminatum – P. regale group indicates an age no older than mid Lutetian. Unfortunately, this group is only tentatively identified.
Apectodinium quinquelatum suggests that the age can be no younger than Lutetian. Phthanoperidinium echinatum and Apectodinium homomorphum both suggest a youngest possible mid or late Bartonian age.
Calcareous nannoplankton
From the Birkweiler drainage pit 34 samples (M1 through M34) were investigated for calcareous nannoplankton. Since only excavated material could be sampled, the stratigraphical succession of the samples could not be established. Eleven of these samples contained autochthonous species, and in seventeen samples reworked Jurassic nannoplankton species such as Ellipsosphaera britannica (Stradner, 1963), Schizosphaerella punctulata Deflandre, 1939, Stephanolithion bigottii Deflandre, 1939 and some other species of Jurassic age were noted.
Two different autochthonous nannoplankton assemblages can be distinguished:
-
a monospecific Nannoserratolithus-Assemblage with the new species Nannoserratolithus minutus, which is very small and could only be identified under the SEM. It is rather common in samples M1, M7 (both lacking macrofossils), M22 (with mass occurrence of hydrobioids and otoliths), but rare to infrequent in samples M6, M10, M12 (all lacking macrofossils), M14 (with hydrobioids) and M23 (with mass occurrence of hydrobioids and otoliths).
-
a Nannoserratolithus/Trochoaster/Trochastrites-Assemblage which is present in samples M25, M26 and M31 (all with Anomalorbina? and otoliths).
Three additional samples from the Hydrobioid-, Anomalorbina?- and Potamididae-Association (see Kadolsky and Hartkopf-Fröder, in prep.) also contain the new nannoplankton species, which is rare in the Hydrobioid-Association and infrequent to common in the other two associations. In addition to the new nannoplankton species two Trochoaster species and one Trochastrites species were found in the Anomalorbina? sample, comparable with the above samples M25, M26 and M31. Rare reworked nannoplankton specimens from the Jurassic were noticed in all three associations.
In total, the following four calcareous nannoplankton species are present in the Birkweiler drainage pit samples.
Family Prinsiaceae Hay and Mohler, 1967
Genus Nannoserratolithus Martini and Schiller, 1998
Nannoserratolithus minutus Martini new species
(Fig. 15 a–d)

Calcareous nannoplankton from Birkweiler drainage pit. Nannoserratolithus minutus new species Martini. Different specimens (SEM). a Holotype SM.B 17726. a, c, d proximal sides, b distal side; sample M1
Holotype: Specimen shown in Fig. 15 a, stored in Senckenberg Forschungsinstitut und Naturmuseum, Frankfurt/Main, under catalogue number SM.B 17726.
Locus typicus: Birkweiler drainage pit, north of Birkweiler near Landau, (map TK 25 Sheet 6714 Edenkoben; UTM coordinates (WGS84) ca. 32U 429700 5451248; ca. R 34 29748 H 54 52990).
Stratum typicum: Landauer Mergel sensu Breuer and Feist (1986), lower part of the Lymnäenmergel, Haguenau-Gruppe, middle Eocene.
Derivatio nominis: minutus (lat.) = tiny.
Diagnosis: Very small, elliptical forms consisting of two shields, with the distal shield being slightly larger than the proximal one. Shields are built out of 10 to 17 segments, with short lateral overlaps like in tiles and tapering with an offset at their outer margins. The segments join in the central area in a slight bend. Long diameter: 2.5 to 3.5 μm. Small diameter: 2.0 to 3.0 μm.
Complete coccospheres, discovered in the Anomalorbina?-Association, have a diameter of 12 μm and consist of approximately 30 coccoliths.
Discussion: The new species seems to represent a freshwater form and resembles Nannoserratolithus dissolutus Martini and Schiller, 1998 from the lower Oligocene “G-Ton” (Maifeld Formation) of the Neuwied Basin (see Martini and Schiller 1998). However, this species is larger and has 20 to 26 segments, which do not join in the central area. From other freshwater species (see Kamptner 1930), especially Hymenomonas krejcigrafii Martini, 1988 from the “Untere Hydrobienschichten” (now Wiesbaden Formation; Reichenbacher and Keller 2002) of the Mainz Basin (Martini 1988), the new species can be easily differentiated by its two shields and shallower construction.
Family Lithostromationidae Deflandre, 1959
Genus Trochoaster Klumpp, 1953
Trochoaster deflandrei (Stradner, 1959) Martini and Stradner, 1960
(Fig. 16 a)

Calcareous nannoplankton from Birkweiler drainage pit. Sample M25 (a, c), and sample from the Anomalorbina?-Assemblage (b). a Trochoaster deflandrei. b Trochoaster simplex. c Trochastrites hohnensis
1959 Polycladolithus deflandrei nov. cent. – Stradner, fossile Discoasteriden Österreichs II, p. 18, fig. 76.
1960 Trochoaster deflandrei (Stradner) – Martini and Stradner, Nannotetraster, p. 3, tab. 1.
1991 Trochoaster deflandrei (Stradner) – Martini, Endemische Vertreter Trochoaster, p. 169, textfig. 2
Description: 6-rayed specimens, with 3 rays 120 degrees apart in an upper level and three rays in a lower level also 120 degrees apart but with a 60 degree offset. A groove is present on the central bridge which is built by ridges between the three main grooves. Towards the outer rim additional grooves and ridges are apparent. Size: 9.0–17.0 μm.
Remarks: Originally described from a marine succession of mid Eocene age from Aragon/Mexico, this species was found in lower Oligocene marine to brackish successions in the Upper Rhine Graben and Mainz Basin (Martini and Reichenbacher 2007), and also in the Chattian of the Lower Rhine region (unpublished).
Rare to few in the Nannoserratolithus/Trochoaster/Trochastrites-Assemblage.
Trochoaster simplex Klumpp, 1953
(Fig. 16 b)
1953 Trochoaster simplex n. sp. – Klumpp, Mikrofossilien Eozän, p. 385, textfig. 4/2, pl. 16 fig. 7.
1958 Trochoaster simplex Klumpp – Martini, Discoasteriden NW deutschen Eozän, p. 368, pl. 5 fig. 25 a, b.
1972 Trochoaster simplex Klumpp – Sherwood and Levin, Closer look at Trochoaster simplex, p. 593, textfig. 1.
Description: 6-rayed specimens with a central groove and three smaller grooves on the central bridge. Rays in one level and 60 degrees apart. Ridges radiate outward from the centre and extend downward into the arms. For further details see Sherwood and Levin (1972). Size: 7.5–17.0 μm.
Remarks: Trochoaster simplex and variants are found in the marine to brackish/limnic sediments of the Mittlere Pechelbronn-Schichten in the Upper Rhine Graben and the Mainz Basin, part of the Melania Clay in northern Hesse and in the Sieblos Formation in the Rhön Mountains (Martini 1991) as well as in the Maifeld Formation in the Neuwied Basin (Martini and Schiller 1998), all of early Oligocene age. A Trochoaster species also occurs in nannoplankton mass-layers in the “Untere Hydrobienschichten” (now Wiesbaden Formation; Reichenbacher and Keller 2002) of the Mainz and Hanau basins (Martini 1988), indicating a certain tolerance of decreasing salinity.
Rare to few in the Nannoserratolithus/Trochoaster/Trochastrites-Assemblage.
Trochoaster sp.
(not figured)
Remarks: Within the Nannoserratolithus/Trochoaster/Trochastrites-Assemblage a large number of aberrant or underdeveloped specimens of a Trochoaster species was found. Most seem to represent Trochoaster simplex, but some may also belong to Trochoaster deflandrei. General size and organisation of rays vary considerably and may result in 4 and up to 9 rays and irregular outline in a specimen.
Genus Trochastrites Stradner, 1961
Trochastrites hohnensis (Martini, 1958) Bouché, 1962
(Fig. 16 c)
1958 Discoaster hohnensis n. sp. – Martini, Discoasteriden NW-deutschen Eozän, p. 358, pl. 2 fig. 10 a, b.
1962 Trochastrites hohnensis (Martini) nov. comb. – Bouché, Nannofossiles Bassin de Paris, p. 91, pl. 4 figs. 13, 14.
1986 Trochastrites hohnensis (Martini) – Aubry, Paleogene nannoplankton stratigraphy, p. 294, pl. 4 fig. 24.
Description: Three-rayed form with a ridge along rays and a more or less prominent bifurcation at the ends of arms. Rays are normally 120 degrees apart, but slightly aberrant specimens are known. Some larger specimens have shallow grooves on the flat parts on both sides of the ridges. Size: 8.3–13.5 μm.
Remarks: Trochastrites hohnensis was first described from the marine “lower Upper Eocene” (“Unteres Ober-Eozän”) from well Hohne 1001 in northern Germany, and is associated with an assemblage belonging to nannoplankton zone NP15 (Martini 1958). Later it was found also in the marine Lutetian of the Paris Basin by Bouché (1962) and associated with Rhabdosphaera inflata Bramlette and Sullivan, 1961 by Aubry (1986). It seems to be restricted to nannoplankton zones NP14 and NP15 of mid Eocene age. The appearance together with a presumably freshwater species and the Trochoaster species, which arise sometimes also as mass occurrences in brackish waters (Martini 1991; Martini and Reichenbacher 2007), indicates a certain tolerance of reduced salinity.
Rare to few in the Nannoserratolithus/Trochoaster/Trochastrites-Assemblage.
The presence of only Jurassic reworked calcareous nannoplankton besides the autochthonous species indicates a closed system without significant long-distance influences. The Jurassic material was probably derived from nearby sources. The onset of numerous reworked species from the Alpine Cretaceous and lower Paleogene occurs in the Rote Leitschicht (Couche rouge), together with the first and sudden influx of calcareous nannoplankton of late Eocene age in the southern Upper Rhine Graben (Martini and Reichenbacher 2007). Also, in the post-Eocene successions in the Upper Rhine Graben, Mainz and Hanau basins reworked nannoplankton is not restricted to Jurassic species as in the Birkweiler samples, but contains always reworked Jurassic and Cretaceous as well as Paleogene species (Martini 1990) indicating long range transport from the Alps into the Upper Rhine Graben at those times.
To get more information on the stratigraphical position of the Birkweiler sediments within the Upper Rhine Graben succession, several samples from the Lymnäenmergel (wells Landau 10, Oberweiler 1973, Freiburg 1974) were investigated together with samples from younger deposits for comparison. The Lymnäenmergel samples also contain only reworked calcareous nannoplankton of Jurassic age, but lack the new species.
The two nannoplankton assemblages from Birkweiler seem to indicate slightly different environments with regard to salinity variation. The monospecific Nannoserratolithus-Assemblage, probably originally representing a freshwater environment, may be followed by the Nannoserratolithus/Trochoaster/Trochastrites-Assemblage which is tolerant of reduced salinity and may point to a brackish water influence. A similar succession from freshwater to brackish water conditions has been reported from the lower Oligocene deposits of Sieblos/Rhön Mountains (Martini and Rothe 1998).
Of stratigraphical importance is the presence of Trochastrites hohnensis in some of the Birkweiler samples. This species seems to be restricted to the mid Eocene as discussed above. Together with the allochthonous nannoplankton species of exclusively Jurassic age it indicates that the succession in the Birkweiler drainage pit belongs to the Landauer Mergel sensu Breuer and Feist (1986) (= lower part of the Lymnäenmergel) which are part of the Eocene Haguenau-Gruppe (Grimm et al. 2011b).
Foraminifera
Assemblages of calcareous foraminifers were found in some layers of borehole B 804 (Fig. 4), furthermore in one sample from the Birkweiler road cut and in one sample from the Birkweiler drainage pit (marls without gastropods, sample F1). The associations are of low diversity and indicate a brackish water environment of weakly increased salinity. Due to these palaeoecological conditions the specimens usually are growth-restricted. Especially the tests from the sample of the road cut often are somewhat corroded and/or recrystallised. The foraminiferal assemblages include “Rosalina” spp., Aubignyna? sp., “Nonion” sp., Anomalinoides sp., Caucasina? elongata (d’Orbigny, 1846), Caucasina? cf. coprolithoides Andreae, 1884, Turrilina sp. and some small Miliolacea referred to as Cycloforina? spp. and Quinqueloculina? sp.
Most of the foraminifers are not age-diagnostic and hence, a specified stratigraphical assignment of the Birkweiler assemblages is not possible. The occurrence of the genus Anomalinoides may indicate an Eocene age as it has been described by Gebhardt et al. (2013) from middle to upper Eocene deposits at the northern Tethyan margin near Adelholzen in southeastern Bavaria (Germany). For the Upper Rhine Graben region a species of this genus was only mentioned by Doebl (1958, 1962: 385) as Anomalina? sp. 300. The taxon is regarded as an index fossil of the Eocene Lymnäenmergel.
“Rosalina” sp. A
(Fig. 17 d–g)

Ostracods and foraminifers from Birkweiler road cut. a, b Cypridopsis? sp., carapaces from right lateral, in Fig. 17 a somewhat crumpled. c Neocyprideis sp., crumpled carapace (♀) from left lateral. d–g “Rosalina” sp. A, umbilical side (d, f); spiral side (e, g). Scale bar: 0.1 mm
Test pentalobulate in outline, often with distinctly projected final chamber. Chambers arranged in one and a half whorls. All chambers visible on the spiral side. Final whorl with 5 chambers, only these chambers visible on umbilical side. Periphery rounded. Sutures somewhat depressed, slightly curved back or nearly radial. Spiral side with densely packed large pores, diameter of pores larger than the distance between the pores. Abundant but not well preserved in the sample from the road cut. Diameter of tests: 0.25–0.35 mm.
“Rosalina” sp. B
(Fig. 18 a, b)

Foraminifers from borehole B 804. a, b “Rosalina” sp. B, spiral side, 7.8–8.0 m depth. d, e “Rosalina” sp. C, spiral side, 7.8–8.0 m depth. c, f Cycloforina? sp. A, lateral view, 11.8–12.0 m depth. g, h Anomalinoides sp., periphery of a broken test showing part of the aperture (g); umbilical side (h), 7.8–8.0 m depth. i Aubignyna? sp., umbilical side of a juvenile test, 23.8–24.0 m depth. Scale bar: 0.1 mm
Test quadrilobulate in outline. Chambers gradually increasing in size, arranged in one and a half whorls. All chambers visible on the spiral side. Final whorl with 4 chambers, only these chambers visible on umbilical side. Periphery rounded. Sutures distinctly depressed, nearly radial. Spiral side with moderately densely packed pores, diameter of pores smaller than the distance between the pores. The species occurs in some layers of borehole B 804, furthermore in the sample from the drainage pit and in the sample from the road cut. Diameter of tests: 0.20–0.23 mm.
“Rosalina” sp. C
(Fig. 18 d, e)
Test pentalobulate in outline. Chambers arranged in one whorl with 5 chambers. Periphery rounded. Sutures slightly depressed, nearly radial. Spiral side with wide-spaced large pores. The species occurs in some layers of borehole B 804 and in the sample from the drainage pit. Diameter of tests: 0.14–0.18 mm.
Aubignyna? sp.

Foraminifers from Birkweiler drainage pit. a–c Aubignyna? sp., spiral side (a), spiral side of a juvenile test (b), umbilical side of a juvenile test (c). d, f, g Caucasina? elongata, juvenile megalospherical test in lateral view (d), microspherical test in oblique lateral view (f), juvenile microspherical test in lateral view (g). e Caucasina? cf. coprolithoides, juvenile test in lateral view. h Cycloforina? sp. B, lateral view. Scale bar: 0.1 mm
Chambers arranged in a low trochospiral coil with 2–3 whorls in full-grown tests, but also small juvenile tests with only one and a half whorls are abundant. Final whorl with 7–8 chambers. Wall finely perforate. Sutures weakly depressed and curved back at the spiralside at least in the final whorl of full-grown tests, nearly radial in the initial whorl. Umbilical side with granules covering the umbilicus and the sutures. Last chamber somewhat inflated and strongly extended to the umbilical side. Aperture obscured by the granular deposit on the umbilical side. The species is abundant in the sample from the drainage pit, but also occurs in some samples of borehole B 804 and in the sample from the road cut. Diameter of tests: 0.11–0.34 mm.
Anomalinoides sp.
(Fig. 18 g, h)
1958 Anomalina? sp. 300 – Doebl, Stratigraphische und paläogeographische Ergebnisse
1962 Anomalina? sp. 300 – Doebl, Tertiär des Rheintalgrabens
Chambers arranged in a very low trochospiral coil with 2–3 whorls. Spiral side evolute. Final whorl with 6–9 coarsely perforated chambers. Umbilical side involute. Sutures incised, nearly radial or slightly curved back. Periphery outline lobulate or nearly rounded. Aperture a narrow slit extending from the base of the last chamber to the umbilical side. The species occurs in the sample from the road cut and in some layers of borehole B 804. Diameter of tests: 0.18–0.34 mm.
Caucasina? elongata (d’Orbigny, 1846)
(Fig. 19 d, f, g)
Test enlarging gradually from initial stage to final stage. Microspherical generation more or less elongated with a low trochospiral coil in initial stage. Later stage high spired with usually 3 chambers per whorl. Aperture a slit or broad oval opening at the face of the final chamber, bordered by a lip. The species occurs only in the sample of the drainage pit. Height of tests: 0.15–0.28 mm.
Caucasina? cf. coprolithoides (Andreae, 1884)
(Fig. 19 e)
Test more inflated. Chambers strongly increasing in size. Aperture a broad oval opening at the face of the final chamber, bordered by a lip. The species is proved only from the sample of the drainage pit. Height of figured test: 0.14 mm.
Cycloforina? sp. A
(Fig. 18 c, f)
Test quinqueloculine. Aperture circular, situated beside the penultimate chamber. No apertural tooth visible. The species is proved only from one sample of borehole B 804. Height of tests: 0.16–0.23 mm.
Cycloforina? sp. B
(Fig. 19 h)
Test quinqueloculine. Aperture circular with simple apertural tooth, situated at the distal end of the test covering parts of the penultimate chamber. The species is proved only from the sample of the drainage pit. Height of tests: 0.16–0.21 mm.
Quinqueloculina? sp.
Some small Miliolacea assigned to the genus Quinqueloculina? were found in some layers of borehole B 804 and in the sample from the drainage pit.
Turrilina sp.
The species only occurs in the sample from the drainage pit.
“Nonion” sp.
One specimen referred to as “Nonion” sp. was found in a sample from borehole B 804 between 7.8–8.0 m depth.
Ostracoda
Ostracods were recovered from two samples of the Birkweiler road cut of which one sample yielded a mass occurrence of the brackish water indicating ostracod genus Neocyprideis. Additional findings of ostracods in samples F1 and F2 from the drainage pit and in samples from borehole B 804 are rare. Some carapaces and often fragmented valves are assigned to the cypridid genus Cypridopsis?, which is well known from the Paleogene to the Recent. The genus usually indicates freshwater environments, but some species are tolerating also slightly increased salinities (Oertli 1963). Yet two other ostracod-species belonging to the genus Ilyocypris? and to the family Trachyleberididae? were found in the foraminifer yielding sample from the drainage pit (sample F1). Both species are documented only by a single carapace and no further detailed taxonomical classification is possible.
Cypridopsis? sp.
(Fig. 17 a, b)
Carapace elongated, highest in mid-length. Left valve larger than right valve. Posterior margin in lateral view somewhat narrower rounded than the anterior margin. Dorsal margin evenly curved or, especially in the right valve, very weakly angulated in mid-length. Vestibule clearly widened on the anterior margin. Fused zone on the ventral margin only widened in the anterior half of the valve. Central muscle scars could not be observed. The species occurs in one sample from the road cut and in sample F2 from the drainage pit. In the latter it is found together with the Hydrobioid-Association. Length of carapaces: 0.51–0.57 mm. Height of carapaces: 0.26–0.31 mm.
Neocyprideis sp.
(Fig. 17 c)
Unfortunately, the recovered material is not well preserved and does not allow a specific determination. Most of the specimens found are females, whose carapaces are inflated at its posterior end, which is caused by a breeding cavity developed in all species of Neocyprideis. Carapaces of the males are more evenly rounded in dorsal view. A comparison with Paleogene Neocyprideis species known from the southern North Sea Basin (see remarks) shows that Neocyprideis sp. resembles Neocyprideis apostolescui (Keij, 1957) in outline, but its dimensions (up to 0.95 mm length and 0.65 mm height measured for a female left valve) are larger. Better preserved material of Neocyprideis sp. is required to decide if this species belongs to the mid Eocene N. apostolescui – although there was no connection between the Upper Rhine Graben system and the North Sea Basin in Eocene times – or should be described as a new taxon. The species is very abundant in a sample of the road cut, rare in the foraminifer yielding sample from the drainage pit, but also occurs in one layer of borehole B 804 where one specimen could be found between 23.8 and 24.0 m depth. Length of carapaces: 0.85–0.95 mm.
Remarks: Keen (1990) studied the ecology and evolution of the ostracod genus Neocyprideis in Paleogene deposits of the southern North Sea Basin. He could recognise a phylogenetic lineage, which allows to separate four species. According to Keen (1990) the most ancestral species is the late Paleocene (Thanetian) Neocyprideis durocortoriensis Apostolescu, 1956. From it evolves N. apostolescui, known from middle Eocene sediments, subsequently the late Eocene Neocyprideis colwellensis (Jones, 1857) and finally Neocyprideis williamsoniana (Bosquet, 1852) in the early Oligocene. The latter species is also described from lower Oligocene sediments of Turkey (Gökçen 1973). The only mention so far of a Neocyprideis species from the Eocene of the entire Upper Rhine Graben area including the Mainz Basin can be found in Ohmert (2014), who specified N. aff. colwellensis from the Lymnäenmergel of a core drilling near Oberweiler in the southern Upper Rhine Graben. However, this is probably not the species described here as Neocyprideis sp. The genus Neocyprideis is most common in mesohaline to polyhaline and possibly in slightly hypersaline environments. Sometimes it is found together with euhaline marine ostracods, but it is always absent in oligohaline or freshwater habitats (Keen 1977, 1990).
Vertebrate remains (fish otoliths excluded)
In addition to the very abundant fish otoliths fish remains such as scales, vertebras, spines and teeth have been found in the Birkweiler samples and in a Hydrobioid-Association from borehole B 804 (27.80–28.00 m). Especially ganoid scales (Fig. 20 a) and jaw teeth (Figs. 6 a, b, 21 a–c) of gars (Lepisosteidae) are common. The gar teeth have a striated basal part and a smooth upper part (compare Grande 2010: 74, figs. 45, 240). The enamel is thin and strongly fluorescent. Dentine tubules which start immediately below the enamel are densely distributed and radiate inwards to the pulp cavity. The diameter of the tubules does not increase towards the pulp. Close to the tip the pulp cavity is very narrow but much wider at the tooth base (Fig. 21 a). Identical teeth and one scale are described by Weiler (1963: 8, figs. 2, 3) as Lepisosteus sp. from olive grey to green grey, fossiliferous claystones and clayey marlstones in well Landau 117 (825–830 m, Landauer Mergel sensu Breuer and Feist 1986, Lutetian) and, additionally, by Weiler (1961: pl. 8, figs. 6–8, 11–14) from the “Melanienton” of Borken (see also Grande 2010: 738). The same tooth type has been reported by Lavoyer (2013: pl. 12, fig. 4) from the Mittlere Pechelbronn-Schichten of the well Preuschdorf near Pechelbronn. In the uppermost Ypresian/lower Lutetian oil shales of the Middle Messel Formation (Lenz et al. 2015) the gars are represented by Atractosteus messelensis Grande, 2010. Today species of the genus Lepisosteus are common in North and Central American lakes and rivers as well as brackish waters of estuaries and marine coastal areas (e.g. Grande 2010; Echelle and Grande 2014). Under special weather conditions mass mortalities of gars may occur and were reported from Smithers Lake, Texas, by Weigelt (1927).

Vertebrate remains from Birkweiler road cut. a Lepisosteidae, scales. b Diplocynodon sp. tooth

Vertebrate remains from Birkweiler road cut. a, b Lepisosteidae, jaw teeth. c Lepisosteidae, jaw tooth, detail of striated basal part. d, e Hassiacosuchus sp., teeth, view at tip (e). f, g Diplocynodon sp., teeth
Spines and soft-rays of fins as well as vertebras of unidentifiable teleosts are rare to few in the collected material. Some of the spines may belong to the perches (Percidae). At present six species of five fish families are known from the classic oil shale of the Middle Messel Formation (Micklich 2018), which includes also three perches: Palaeoperca proxima Micklich, 1978, Rhenanoperca minuta Gaudant and Micklich, 1990 and Amphiperca multiformis Weitzel, 1933.
A sample rich in hydrobioid gastropods and otoliths from the Birkweiler road cut yielded disarticulated turtle plates, with the largest fragment measuring 17 x 25 mm. As the surface of the broken pieces are smooth and not sculptured they seem to belong to the swamp turtle Palaeoemys (syn. Palaeochelys), and not to Allaeochelys or Trionyx. Four species of turtles are known from the uppermost Ypresian/lower Lutetian Middle Messel Formation with Palaeoemys messeliana (Staesche, 1928) as the most common one, formerly placed in the genus Ocadia (see Cadena et al. 2018).
Ninety-five isolated teeth of alligatoroids were recovered from a Birkweiler road cut sample. They belong mainly to the genus Diplocynodon. These teeth are up to 17 mm long, are slightly bent and have weakly keeled sides (Figs. 20 b, 21 f, g). The larger ones are often split in two or more pieces. Very similar teeth were depicted by Weiler (1963: 10, fig. 10 a, b) from the Lutetian marls in borehole Landau 117 (825–830 m) and by Lavoyer (2013: pl. 13, fig. 1) from the middle Lutetian of Bouxwiller. A similar specimen (Heidtke 2006: fig. 14) is supposed to have been found in Lower Jurassic sedimentary rocks of the brickyard pit west of Siebeldingen but obviously originates from middle Eocene sediments which were exposed in the area of the former brickyard pit (Schwarz and Griessemer 1998). All material in our study was isolated from hydrobioid and otolith rich marl. The teeth are surprisingly abundant and rather well preserved. Hence, for our material reworking from Jurassic strata is excluded and the Diplocynodon teeth are definitely of mid Eocene age. Some sturdier and smaller teeth with more rounded tips seem to belong to the genus Hassiacosuchus (Fig. 21 d, e). This type was also reported from the middle Lutetian of Bouxwiller (Lavoyer 2013: pl. 13, fig. 4). Both alligatoroid genera were recorded from the uppermost Ypresian/lower Lutetian Middle Messel Formation as Diplocynodon darwini (Ludwig, 1877), Diplocynodon deponiae (Frey et al., 1987) and Hassiacosuchus haupti (Weitzel, 1935), formerly placed in the genus Allognathosuchus, together with representatives of the additional genera Asiatosuchus, Boverisuchus and Bergisuchus (Keller and Schaal 1988, Brochu and Miller-Camp 2018).
The above vertebrates of the Birkweiler locality are indicative of large water bodies with a well-established food chain and show some similarity regarding age and faunal composition to the latest Ypresian/early Lutetian Messel fauna (compare Smith et al. 2018).
Stratigraphical synthesis
The 3 m thick clay layer with numerous reddish brown to black concretions (“Bohnerz”) at the very base of the Birkweiler road cut section did not yield any macrofossils. Hence, the depositional environment and stratigraphy is not well known. The hematite/goethite concretions and the kaolinite-rich clay are interpreted as a residual terrestrial sediment, i.e. a ferruginous palaeosol which formed under tropical-subtropical conditions (Sittler 1970). In the middle and southern Upper Rhine Graben such sediments are widespread at the base of the Paleogene succession (e.g. Doebl 1970). Usually, these residual sediments whose thickness varies between less than a metre to more than 100 m overlay karstified Jurassic limestones (Pirkenseer et al. 2018). In the Mainz Basin similar formations are of late Miocene age (Schäfer 2012). In the Birkweiler road cut the lack of any fossils in the ferruginous palaeosol hampers a bio- and chronostratigraphical assignment. The designation of the lowermost part of the Birkweiler road cut section to the Schliengen Formation is therefore solely based on petrography and its position overlying the Lower Jurassic mudstones and underlying sediments dated as Eocene.
The sediments overlying the Schliengen Formation are exceptional in that they yielded pollen and spores, dinocysts, green algae, calcareous nannoplankton, foraminifers, gastropods, ostracods and vertebrate remains, including numerous otoliths, fish teeth and scales and reptile remains.
The presence of some typical mid Eocene pollen and spores such as Tegumentisporis villosoides, Tricolporopollenites crassostriatus and representatives of Bombacaceae is significant. However, T. villosoides and pollen of Bombacaceae were recently identified from the upper Ypresian to lower Lutetian of Messel and the nearby locality Grube Prinz von Hessen (Lenz and Wilde 2018; Moshayedi et al. 2018) and, consequently, a late Ypresian age cannot be ruled out. Likewise, a late Eocene age is less likely due to the absence of the Mediocolpopollis Group which is an important marker taxon for the late Eocene in Central Europe (Krutzsch 2008; Hottenrott in Grimm et al. 2011a) although it has been reported from the mid Eocene until the beginning of the late Eocene of the Paris Basin (Schuler 1990).
Although the dinocyst diversity is very low with only twelve species recorded the assemblage constrains the age of the sediments from the Birkweiler drainage pit to not older than late Ypresian and probably not younger than Lutetian. The presence of numerous specimens of Phthanoperidinium comatum indicates that the assemblage cannot be older than early NP13 (late Ypresian). The Phthanoperidinium geminatum – P. regale group would restrict the oldest possible age to mid Lutetian but only two specimens were encountered so that this age assignment must be taken with some caution. The occurrence of Apectodinium quinquelatum suggests that the samples cannot be younger than Lutetian while Phthanoperidinium echinatum and Apectodinium homomorphum imply an age not younger than mid or late Bartonian.
Trochastrites hohnensis seems to be restricted to nannoplankton zones NP14 and NP15 (Lutetian). Foraminifers, ostracods and vertebrate remains do not further refine this biostratigraphical dating.
Based on pollen and spores, dinocysts and calcareous nannoplankton a Lutetian age is most probable though a late Ypresian to Bartonian age cannot be completely ruled out at this point.
Because of the exceptional faunal composition the lithostratigraphical assignment of the Birkweiler fossiliferous marls, organic matter-rich clays and lignite is not obvious. In the Upper Rhine Graben the Schliengen Formation is overlain by the Haguenau-Gruppe which is subdivided by Grimm et al. (2011b) from base to top into the Bouxwiller Formation, Grüne Mergel and Rote Leitschicht. The Bouxwiller Formation, only present in some isolated areas of the middle and southern Upper Rhine Graben, is well dated by charophytes and mammals and assigned to the mid Lutetian MP13b zone (Berger et al. 2005a; Lavoyer 2013). In many boreholes a gradual transition from the Siderolith “Facies” into the Grüne Mergel “Facies” has been observed (Hinsken et al. 2007). The Grüne Mergel are still an informal formation but nevertheless were differentiated into three members, the basal Untere Grüne Mergel, the Brunstatt Subformation and the hombresi-Mergel at the top (Grimm et al. 2011b). The application of the Grüne Mergel and the Untere Grüne Mergel may be questionable as pointed out by Kadolsky and Hartkopf-Fröder (in prep.). A widely used term in hydrocarbon exploration and basic research for middle and upper Eocene sediments in the Upper Rhine Graben is the Lymnäenmergel (e.g. Doebl 1958, 1962; Wirth 1969; Schäfer 2000, 2013). The middle Eocene basal part of the Lymnäenmergel probably includes the Landauer Mergel sensu Breuer and Feist (1986: fig. 2) which is an informal and poorly defined lithostratigraphical six metre thick unit composed of olive grey to green grey, fossiliferous claystones and clayey marlstones in well Landau 117. In this well the Landauer Mergel overlies Triassic sedimentary rocks (middle Keuper) and is overlain by the middle Rupelian Alzey Formation. Hence, the lower and the upper boundary of the Landauer Mergel are hiatuses and the entire stratigraphical range of this unit is insufficiently known. The succession in well Landau 117 includes a horizon extremely rich in gyrogonites, foraminifers, ostracods, fish and crocodylian teeth, otoliths and fragments of vertebrates (Weiler 1963; Breuer and Feist 1986). According to the charophytes the Landauer Mergel is assigned to the Maedleriella embergeri Zone (Lutetian) (Breuer and Feist 1986). Gyrogonites are absent in our samples but have been recorded by Schwarz and Griessemer (1998) from green marls temporarily exposed in the area of the former brickyard pit near the Birkweiler road cut and assigned to the Landauer Mergel. Like in well Landau 117 this assemblage is indicative of the Maedleriella embergeri Zone. As in the faunal list of Breuer and Feist (1986) gastropods are not mentioned they are obviously lacking in this well. However, in several wells in the Landau oil field numerous mid Eocene gastropods, ostracods, gyrogonites and fish remains, including otoliths, have been encountered (Wirth 1969). Unfortunately, no data on calcareous nannoplankton and palynomorphs are available from this succession. Despite distinct differences between the Birkweiler and the Landauer Mergel assemblages in well Landau 117 they have characteristic elements in common, i.e. a fauna clearly indicating brackish water conditions. Hence, the sediments at least from the Birkweiler drainage pit and probably also from the basal part of borehole B 804 are provisionally assigned to the Lutetian Landauer Mergel sensu Breuer and Feist (1986) (= lower part of the Lymnäenmergel) until the lithostratigraphical subdivision of the Eocene succession in the Upper Rhine Graben is revised.
Palaeoecological synthesis
All fossil groups from the Birkweiler sites show very low diversity except of the pollen and spores and the moderately diverse otolith fauna (Bettina Reichenbacher, written communication, February 01, 2023) indicating an extreme environment probably due to reduced and fluctuating salinity as it is typical for marginal brackish water settings. Interestingly, all studied samples did not yield a single charophyte specimen though from the brickyard pit west of Siebeldingen Schwarz and Griessemer (1998) isolated numerous gyrogonites from green marls of Lutetian age.
The macropalaeontological data clearly indicate that at least three different gastropod associations are present in the sediments from the Birkweiler drainage pit (Kadolsky and Hartkopf-Fröder, in prep.):
-
the Hydrobioid-Association, also present in the Birkweiler road cut and in borehole B 804, is characterised by mass occurrences of Lutetiella hartkopfi Kadolsky, 2015 associated with few other rare gastropods, abundant otoliths and the presumably freshwater calcareous nannoplankton element Nannoserratolithus minutus n. sp.
-
the Potamididae-Association with Potamididae as the only gastropod taxon, abundant otoliths and Nannoserratolithus minutus n. sp.
-
the Anomalorbina?-Association distinguished by Anomalorbina?, some otoliths and fish bones and the calcareous nannoplankton Nannoserratolithus/Trochoaster/Trochastrites-Assemblage.
Neither the calcareous nannoplankton (except of Nannoserratolithus minutus n. sp.) nor the gastropod fauna from the Birkweiler drainage pit indicate a fully marine or a freshwater environment. Though some of the Trochoaster and Trochastrites species have been recorded in marine sediments they prevail in brackish water habitats in the Upper Rhine Graben, the Mainz Basin and some adjacent areas. All these species are tolerant of reduced salinity. The rare Viviparidae are the only unequivocal freshwater gastropod specimens. They occur as small fragments which are derived from a habitat different from that of the abundant Lutetiella. The hydrobioids and the Potamididae are all indicative of a brackish water environment (Kadolsky and Hartkopf-Fröder, in prep.). The same is true for the ostracod genus Neocyprideis which is extremely abundant at least in one horizon from the Birkweiler road cut. In almost all studied samples dinocysts are present but with very low diversity (only 12 species). It is noteworthy that gonyaulacoids are represented only by one species while genera typically abundant in marine environments are absent. The majority of dinocysts belong to heterotroph peridinoids such as Apectodinium and Phthanoperidinium indicating a nutrient-rich water body. Especially, the former is an opportunistic genus favoured by brackish lagoonal or estuarine to very near-shore environments. In addition, Phthanoperidinium and Polysphaeridium zoharyi are tolerant to salinity fluctuations (e.g. Sluijs and Brinkhuis 2009; Frieling and Sluijs 2018). The chlorococcalean green algae Pediastrum and Botryococcus point to freshwater input. Based on the dinocyst and green algae assemblage a completely landlocked, brackish water body with fluctuating salinity is assumed. The occurrence of foraminifers and a single microforaminiferal lining (Stancliffe 1996) suggest at least brackish conditions. A thick stromatolite horizon in the Birkweiler road cut section, similar to the Eocene/Oligocene microbial facies “biohermes” of Duringer and Gall (1994) in the southern Upper Rhine Graben, indicates a shallow water environment, possibly with temporarily dry periods.
A temporarily brackish water environment was also supposed for the Lymnäenmergel in the southwestern part of the Mainz Basin (in boreholes Ebertsheim and Lautersheim 1 near Eisenberg), where foraminifers together with gyrogonites and limnophilous ostracods have been encountered. As in borehole Lautersheim 1 the Lymnäenmergel reaches 28.4 m in thickness the northernmost Lymnäenmergel deposits with brackish influence can be expected in the Marnheim Bay sensu Spuhler (1937) few kilometres further north (Schäfer 2000). The moderate to high sulphur content and the maceral distribution in sapropelic coals from the Lymnäenmergel encountered in various oil wells in the central Upper Rhine Graben point to a paralic swamp, marsh or coastal plain environment (Böcker et al. 2017).
Though most recorded faunal taxa are indicative of a brackish water environment some as the alligatoroid genera Diplocynodon and Hassiacosuchus as well as the turtles have been documented in the Upper Rhine Graben exclusively from freshwater deposits.
Four species of turtles, all classified as freshwater turtles, have been reported from Messel (Cadena et al. 2018). Many modern freshwater turtles are vulnerable to increased salinity. In brackish and marine environments they become osmotically dehydrated, suffer from body mass loss and will not survive prolonged salinity exposure. However, numerous modern freshwater turtles inhabit estuarine and brackish water systems at least occasionally. In order to endure in such adverse environments freshwater turtles respond to elevated salinities by various behavioural or physiological strategies such as frequent retreats to freshwater habitats, reduction of feeding and drinking to avoid ingestion of salt water or drinking after heavy rainfalls when sea surface salinity becomes fresher. In addition, many modern freshwater turtle species are able to maintain osmoregulatory homeostasis in brackish water environments for a short time period and at least one species, the diamondback terrapin (Malaclemys terrapin), possesses a lachrymal gland to excrete excess salt (for detailed discussion on salinity tolerances of freshwater turtles see e.g. Bower et al. 2016; Agha et al. 2018, 2019). Reports from the fossil record of freshwater turtles found in indisputable brackish water or marine habitats seem to be rare (Agha et al. 2018). Nevertheless, it cannot be excluded that the turtles from Birkweiler were able to colonise transiently saline water habitats as long as they could temporarily move to freshwater areas.
The alligatoroid genera Diplocynodon and Hassiacosuchus are well known from Messel and Bouxwiller. Without doubt Lake Messel had no connection to brackish or marine areas. For Bouxwiller deposition in small lakes was ascertained (e.g. Grimm et al. 2011b). Recently, Lavoyer (2013) discussed the possibility that some taxa (Lamprothamnium, Hydrobiidae) from Bouxwiller may be tolerant of brackish water. As there are no additional indications for a brackish water influence, e.g. dinocysts and foraminifers, and the general palaeogeographical situation excludes a link to the open sea (see below) we prefer to retain the previous model of a small, fully freshwater lake for the Bouxwiller site. Basically, modern alligators are stenohaline and largely restricted to fresh water. Contrary to crocodiles, alligators do not possess salt glands so that salinity tolerance is low which prevents long-term survival in brackish or even fully marine water bodies (Mazzotti and Dunson 1989; Taplin and Grigg 1989). Adult alligators occasionally forage in brackish water environments such as highly-productive estuaries and near coastal areas (Mazzotti and Dunson 1989) while juvenile individuals exposed to brackish water (salinity concentration 12 ‰) suffer from loss of body mass, severe dehydration and adverse effects on various physiological functions (Faulkner et al. 2018). Based on modern and fossil evidence, a permanent colonisation of the Birkweiler brackish water habitat by the alligatoroid genera Diplocynodon and Hassiacosuchus is unlikely. At best, the brackish water body was visited for occasional forays to feed on various vertebrates, e.g. fish and turtles. If this conception is correct, the alligatoroid teeth were most presumably washed from the original freshwater environment to the final site of deposition. As the teeth are extremely compact and wear-resistant they can be transported over considerable distance without exhibiting abrasion features.
The pollen and spores suggest a palaeotropical, diverse vegetation with Cupressaceae/Taxodiaceae-dominated swamp forest, probably also associated with Myricaceae, Juglandaceae, Oleaceae and Aquifoliaceae. It is interesting, that Pentapollenites pentangulus, a species of uncertain botanical affinity, covaries with Cupressaceae/Taxodiaceae and other wet elements. We, hence, tentatively suggest that this also biostratigraphically important species – emerging in the Paris Basin since the mid Eocene according to Châteauneuf (1980) – might also favour locally wet conditions. A rainforest with palms, Sapotaceae, Bombacaceae, Juglandaceae, Malvaceae and many other families including evergreen climbers of the Vitaceae existed close to the site. The presence of mangroves, further pointing to a brackish water influence, cannot be excluded as deduced from the occurrence of Leiotriletes adriennis. The latter has been associated with the mangrove fern Acrostichum but Lygodium may be an additional/alternative producer of this spore.
Provided that temperature preference of modern Eucommia ulmoides can be extrapolated to Eocene Eucommia species, the spring temperatures during deposition of the Landauer Mergel was from 13 °C to 20 °C (Yu-Fei Wang et al. 2003). The dinocyst Apectodinium is very abundant and reaches 99 % in one sample. Such Apectodinium acmes point to a water temperature higher than 20 °C (Frieling et al. 2014). The presence of ectothermic crocodilians indicates a Mean Annual Temperature of > 14.2 °C and a Coldest Month Mean temperature of > 5.5 °C but alligatoroids, in particular larger individuals, seem to tolerate lower temperatures (Markwick 1998).
The high terrestrial input emphasised by type III kerogens and alkane distribution and the presence of a diverse pollen and spores assemblage including large and strongly ornamented species imply that the coast was not far from the depositional site. The lack of coarse-grained sediments in the Birkweiler sections located close to the evolving graben shoulder emphasises that subsidence was probably still negligible and that the sediments were deposited in regional shallow depressions. This is in accordance with e.g. Doebl and Bader (1970: 15, pl. A1), Sittler (1992: fig. 1), Duringer (1995: 167, 170, fig. 4) and Sissingh (1998: 261) who ascertained a negligible uplift of the graben shoulder during the mid Eocene.
Palaeogeographical implications
It is still a matter of debate if during the mid Eocene shallow marine connections existed between the Upper Rhine Graben and adjacent basins to enable migration of brackish water organisms. There is general consensus that a seaway from the northern Upper Rhine Graben to the North German-Polish Basin via the Hessen Depression or possibly also to the North Sea Basin via the Linz Passage was open not until the early Oligocene (e.g. Sissingh 2003, 2006; Ohmert 2017; Pirkenseer et al. 2018). Following the palaeogeographical reconstructions of Berger et al. (2005b) and Lavoyer (2013) the southern Upper Rhine Graben was land-locked during the mid Eocene and no marine communication established towards the North Alpine ‘Pre-Molasse’ Basin. This is in accordance with numerous other publications (e.g. Sissingh 1998, 2006; Dèzes et al. 2004; Pirkenseer et al. 2018). However, it must be noted that the mid Eocene palaeogeography as proposed by Sissingh (1998, 2006) is based on the assumption of a late Lutetian to Bartonian hiatus in the Upper Rhine Graben due to non-deposition and erosion as well as an age of the “Limnaea Marls” not older than Priabonian. More recent studies do not support such a hiatus (Hinsken et al. 2007; Lavoyer 2013) and the age of the “Limnaea Marls” ranges from the mid Lutetian to mid Priabonian (Grimm et al. 2011b). Following Sissingh (1998, 2003, 2006) the first marine ingression into the Upper Rhine Graben occurred during the Priabonian via the Rhône and Bresse graben systems, the Burgundy Passage and the Délemont and Laufen basins but palaeoecological, sedimentological and geochemical data may preclude such a seaway (Fontes et al. 1991). A third option, a short-term mid Eocene marine incursion from the Paris Basin, has been suggested by Grimm et al. (2011b: 59) but a connection between the Paris Basin and the Upper Rhine Graben is not considered by e.g. Gély (2008) and Huyghe et al. (2015).
Provided that the mid Eocene Upper Rhine Graben was palaeogeographically isolated from the North German-Polish Basin, the North Sea Basin, Paris Basin and North Alpine ‘Pre-Molasse’ Basin colonisation of the brackish water inland sea in the Upper Rhine Graben by aquatic invertebrates was possible through passive dispersal vectors such as wind (anemochory, e.g. Rivas et al. 2019) and rain. Additionally, numerous invertebrates (e.g. flying insects) and vertebrates (e.g. aquatic birds, mud-wallowing mammals) have been empirically identified as crucial in the dispersal of viable propagules and living individuals since Darwin (1859: 385; 1882) reported on freshwater bivalves sticking to aquatic insects and birds. Thus, long-distance transport of fish eggs by water fowl or land animals has been taken into consideration by Reichenbacher et al. (2007) for the distribution of gobiid fishes from the Paratethys to the Upper Rhine Graben during the late Burdigalian. Transport can be achieved both internally (endozoochory) and externally (ectozoochory) (Bilton et al. 2001; Green and Figuerola 2005; Vanschoenwinkel et al. 2011; Coughlan et al. 2017, with further references therein). Diapausing, desiccation-resistant resting stages that adhere to plumage, bills and legs of waterbirds or the moist fur of aquatic mammals can easily be transported from one water body to another (Coughlan et al. 2015). Various studies proved that some larvae can survive out of water long enough for travel distances of 10 km and more if they are attached to waterfowl, ducks or mallards. Aquatic and semi-aquatic birds were already present during the Eocene (Mayr 2017). Various (resting) eggs can even hatch after passing the digestive tract of insects, fish (ichthyochory, Guy-Haim et al. 2017) and birds. Recently, Silva et al. (2019) and Lovas-Kiss et al. (2020) demonstrated that not only highly resistant, diapaused eggs of killifish but even the soft-membraned eggs of cyprinids survive ingestion, the acidic and anoxic environment in the digestive tract of waterbirds and egestion. Although only 0.2–1 % of the ingested eggs were found viable in the fresh faeces it is assumed that bird-mediated, internal transport of fish eggs enables the establishment of new populations in isolated, remote water bodies.
Deduced from numerous zoological studies on the dispersal of invertebrate eggs, larvae and even adult individuals via mechanisms such as anemochory, endozoochory and ectozoochory, it seems to be conclusive that a connection from the Upper Rhine Graben to adjacent marine basins was not a prerequisite to explain the presence of a brackish water fossil assemblage of low diversity. Rather, wind, rain or highly mobile animals were vectors for the dispersal of dinocysts and invertebrates which resulted in the successful establishment of a brackish water biota in a remote water body near the western shoulder of the evolving middle Upper Rhine Graben far away from the open sea.
Conclusion
Various thermal maturity parameters, in particular Spore Color Standard, Tmax, VRr and biomarkers, indicate that the organic matter of the middle Eocene Landauer Mergel in the Birkweiler drainage pit site is immature and thermal maturity is distinctly lower compared to approximately time equivalent sediments in the Landau oilfield. While the Birkweiler sites are located next to the graben shoulder, this oilfield is situated more graben-wards where the overburden thickness for the Landauer Mergel is much higher (Schad 1962) and in the case of the studied samples more than 1900 m. The hydrocarbon generation potential of the Birkweiler marl samples was analysed by TOC and Rock-Eval measurements and show low values indicative of a poor generative potential. However, liptinite-rich (sapropelic) coals and organic-rich mudstones as were encountered at the base of the Birkweiler road cut section (Fig. 4) are considered as source rocks of ‘high wax oils’, which also contributed to the main oil fields (e.g. Landau and Römerberg). These sapropelic coals are characterised by high amounts of terrestrial organic matter. In various wells, e.g. near Speyer, about 30 km northeast of Birkweiler, they show extraordinarily high hydrocarbon potential which may be even higher than that of the Lower Jurassic Posidonia Shale (Bruss 2000; Böcker et al. 2017).
Following age indicative pollen and spores, dinocysts and calcareous nannoplankton, a Lutetian age is considered to be most probable while a late Ypresian to Bartonian age is considerably less likely.
Most of the dinocysts, calcareous nannoplankton, foraminifers, ostracods and gastropods (Kadolsky and Hartkopf-Fröder, in prep.) point to a brackish water environment. Only the Viviparidae are restricted to fresh water and were likely washed into the brackish water environment (Kadolsky and Hartkopf-Fröder, in prep.). The alligatoroids Diplocynodon and Hassiacosuchus are well known from freshwater lakes such as Messel (Brochu and Miller-Camp 2018). As for the Viviparidae remains of alligatoroids and turtles may have been displaced from their freshwater habitats into a brackish water environment. However, it cannot be excluded that alligatoroids and turtles were also able to occasionally inhabit brackish water settings. Trochoaster and Trochastrites are well known to occur in brackish water habitats in the Upper Rhine Graben. The occurrence of the ostracod Neocyprideis and dinocyst and foraminifer assemblages of very low diversity are all typical for brackish water habitats while unequivocal marine indications such as diverse dinocyst or foraminifer associations are lacking. Also, the n-alkane distribution does not support a marine environment. A thick stromatolite horizon indicates a shallow water environment.
Fossil-rich middle Eocene sediments deposited in such an environment have rarely been reported from the Upper Rhine Graben (for examples see Kadolsky and Hartkopf-Fröder, in prep.) and the concurrent occurrence of pollen and spores, dinocysts, green algae, calcareous nannoplankton, foraminifers, gastropods, ostracods and vertebrate remains (particularly otoliths, reptiles) has been unknown so far.
The Birkweiler assemblages are similar but not identical to the one reported from the Landauer Mergel in well Landau 117 by Breuer and Feist (1986). The main differences are the lack of charophytes in the Birkweiler assemblages which, however, have been reported by Schwarz and Griessemer (1998) from a nearby site. In addition, in well Landau 117 gastropods are lacking suggesting some environmental differences between both sites. Nevertheless, the occurrence of foraminifers indicates a brackish water environment similar to Birkweiler.
The Birkweiler assemblages are the most diverse brackish water assemblages reported so far from the Eocene of the Upper Rhine Graben. The fauna flourished in a shallow brackish water inland sea with fluctuating salinity located near the western shoulder of the evolving Upper Rhine Graben. Along water bodies wetlands developed where sapropelic peat was deposited.
Regardless of whether a short-lived communication between the Upper Rhine Graben and adjoining marine basins was maintained or the Upper Rhine Graben was land-locked with no connection to any marine basin during deposition of the Birkweiler succession, brackish water organisms may have been dispersed over long distances by different vectors such as wind, rain, highly mobile insects or vertebrates. Obviously, dispersal of aquatic invertebrates by endo- and ectozoochory has so far been underestimated to conclusively explain the occurrence of Paleogene brackish water faunas in the land-locked Upper Rhine Graben.
Data availability
Fossil material, sediment samples, thin sections and polished epoxy-mounted samples are stored in various collections open to the public on reasonable request (for details see Material). All data generated or analysed during this study are included in this published article and its supplementary information files.
References
Agha, M., Ennen, J. R., Bower, D. S., Nowakowski, A. J., Sweat, S. C., & Todd, B. D. (2018). Salinity tolerances and use of saline environments by freshwater turtles: implications of sea level rise. Biological Reviews, 93(3), 1634–1648.
Agha, M., Yanagitsuru, Y. R., Fangue, N. A., Nowakowski, A. J., Kojima, L. V., Cech Jr., J. J., Riley, M. K., Freeman, J., Cocherell, D. E., & Todd, B. D. (2019). Physiological consequences of rising water salinity for a declining freshwater turtle. Conservation Physiology, 7(1). https://doi.org/10.1093/conphys/coz054.
Akkiraz, M. S., Akgün, F., Orcen, S., Bruch, A. A., & Mosbrugger, V. (2006). Stratigraphic and palaeoenvironmental significance of Bartonian, Priabonian (Middle, Late Eocene) microfossils from the Bascesme Formation, Denizli Province, western Anatolia. Turkish Journal of Earth Sciences, 15(2), 155–180.
Allenbach, R. P., & Wetzel, A. (2006). Spatial patterns of Mesozoic facies relationships and the age of the Rhenish Lineament: a compilation. International Journal of Earth Sciences, 95(5), 803–813.
Ashraf, A. R., & Hartkopf-Fröder, C. (1996). Die Siebverfahren bei der Aufbereitung palynologischer Proben. Neues Jahrbuch für Geologie und Paläontologie Abhandlungen, 200(1–2), 221–235.
Aubry, M.-P. (1986). Paleogene calcareous nannoplankton biostratigraphy of northwestern Europe. Palaeogeography, Palaeoclimatology, Palaeoecology, 55(2–4), 267–334.
Baillieux, P., Schill, E., Edel, J.-B., & Mauri, G. (2013). Localization of temperature anomalies in the Upper Rhine Graben: insights from geophysics and neotectonic activity. International Geology Review, 55(14), 1744–1762.
Banerjee, A., Sharma, R., Chisti, Y., & Banerjee, U. C. (2002). Botryococcus braunii: a renewable source of hydrocarbons and other chemicals. Critical Reviews in Biotechnology, 22(3), 245–279.
Behrmann, J. H., Hermann, O., Horstmann, M., Tanner, D. C., & Bertrand, G. (2003). Anatomy and kinematics of oblique continental rifting revealed: A three-dimensional case study of the southeast Upper Rhine graben (Germany). AAPG Bulletin, 87(7), 1105–1121.
Berger, J.-P., Reichenbacher, B., Becker, D., Grimm, M., Grimm, K., Picot, L., Storni, A., Pirkenseer, C., & Schäfer, A. (2005a). Eocene–Pliocene time scale and stratigraphy of the Upper Rhine Graben (URG) and the Swiss Molasse Basin (SMB). International Journal of Earth Sciences, 94(4), 711–731.
Berger, J.-P., Reichenbacher, B., Becker, D., Grimm, M., Grimm, K., Picot, L., Storni, A., Pirkenseer, C., Derer, C., & Schäfer, A. (2005b). Paleogeography of the Upper Rhine Graben (URG) and the Swiss Molasse Basin (SMB) from Eocene to Pliocene. International Journal of Earth Sciences, 94(4), 697–710.
Bilton, D. T., Freeland, J. R., & Okamura, B. (2001). Dispersal in freshwater invertebrates. Annual Review of Ecology and Systematics, 32, 159–181.
Blumenstengel, H. (1993). Die Pollenzone 16 des Paläogens Mitteldeutschlands – Probleme der Definition und Abgrenzung. In Pflanzen der geologischen Vergangenheit. Festschrift Prof. W. Krutzsch (pp. 105–111). Berlin: Museum für Naturkunde der Humboldt-Universität, Paläontologisches Museum.
Blumenstengel, H., Krutzsch, W., & Volland, L., with contributions by Knoth, W., & Knuth, G. (1996). Revidierte Stratigraphie tertiärer Ablagerungen im südlichen Sachsen-Anhalt. Teil 1: Raum Halle-Merseburg. Hallesches Jahrbuch für Geowissenschaften, Reihe B, Beiheft, 1, 1–101.
Böcker, J., & Littke, R. (2014). Source rock characterisation and thermal maturity of the Rupelian Fish Shale (Bodenheim Fm./Hochberg Subfm.) in the central Upper Rhine Graben. Zeitschrift der Deutschen Gesellschaft für Geowissenschaften, 165(2), 247–274.
Böcker, J., & Littke, R. (2016). Thermal maturity and petroleum kitchen areas of Liassic Black Shales (Lower Jurassic) in the central Upper Rhine Graben, Germany. International Journal of Earth Sciences, 105(2), 611–636.
Böcker, J., Littke, R., & Forster, A. (2017). An overview on source rocks and the petroleum system of the central Upper Rhine Graben. International Journal of Earth Sciences, 106(2), 707–742.
Bouché, P. M. (1962). Nannofossiles calcaires du Lutétien du Bassin de Paris. Revue de Micropaléontologie, 5(2), 75–103.
Bourgeois, O., Ford, M., Diraison, M., Le Carlier de Veslud, C., Gerbault, M., Pik, R., Ruby, N., & Bonnet, S. (2007). Separation of rifting and lithospheric folding signatures in the NW-Alpine foreland. International Journal of Earth Sciences, 96(6), 1003–1031.
Bower, D. S., Scheltinga, D. M., Clulow, S., Clulow, J., Franklin, C. E., & Georges, A. (2016). Salinity tolerances of two Australian freshwater turtles, Chelodina expansa and Emydura macquarii (Testudinata: Chelidae). Conservation Physiology, 4(1). https://doi.org/10.1093/conphys/cow042.
Boy, J. A., Haneke, J., Kowalczyk, G., Lorenz, V., Schindler, T., Stollhofen, H., & Thum, H. (2012). Rotliegend im Saar-Nahe-Becken, am Taunus-Südrand und im nördlichen Oberrheingraben. Schriftenreihe der Deutschen Gesellschaft für Geowissenschaften, 61, 254–377.
Brasero, N., Nel, A., & Michez, D. (2009). Insects from the Early Eocene amber of Oise (France): diversity and palaeontological significance. Denisia, 26, 41–52.
Breuer, R., & Feist, M. (1986). Biostratigraphisch bedeutsame Charophyten aus dem Alttertiär von Erdölfeldern des Oberrheingrabens. Newsletters on Stratigraphy, 16(3), 139–147.
Brochu, C., & Miller-Camp, J. (2018). Crocodyliforms – large-bodied carnivores. In K. T. Smith, S. F. K. Schaal, & J. Habersetzer (Eds.), Messel – An ancient greenhouse ecosystem (pp. 158–167). Stuttgart: Schweizerbart.
Bruss, D. (2000). Zur Herkunft der Erdöle im mittleren Oberrheingraben und ihre Bedeutung für die Rekonstruktion der Migrationsgeschichte und der Speichergesteinsdiagenese. Berichte des Forschungszentrums Jülich, 3831, I–XII + 1–222.
Bucher, W. (1914). Beitrag zur geologischen und paläontologischen Kenntnis des jüngeren Tertiärs der Rheinpfalz. Geognostische Jahreshefte, 26, 1–103.
Bujak, J. P., & Brinkhuis, H. (1998). Global warming and dinocyst changes across the Paleocene/Eocene Epoch boundary. In M.-P. Aubry, S. G. Lucas, & W. A. Berggren (Eds.), Late Paleocene – early Eocene climatic and biotic events in the marine and terrestrial records (pp. 277–295). New York: Columbia University Press.
Bujak, J. P., Downie, C., Eaton, G. L., & Williams, G. L. (1980). Dinoflagellate cysts and acritarchs from the Eocene of southern England. Special Papers in Palaeontology, 24, 1–100.
Cadena, E., Joyce, W. G., & Smith, K. T. (2018). Turtles – armored survivalists. In K. T. Smith, S. F. K. Schaal, & J. Habersetzer (Eds.), Messel – An ancient greenhouse ecosystem (pp. 148–157). Stuttgart: Schweizerbart.
Cendón, D. I., Ayora, C., Pueyo, J. J., Taberner, C., & Blanc-Valleron, M.-M. (2008). The chemical and hydrological evolution of the Mulhouse potash basin (France): Are “marine” ancient evaporites always representative of synchronous seawater chemistry? Chemical Geology, 252(3–4), 109–124.
Châteauneuf, J.-J. (1980). Palynostratigraphie et paléoclimatologie de l’Éocène supérieur et de l’Oligocène du Bassin de Paris. Mémoire du Bureau de Recherches Géologiques et Minières, 116, 1–424.
Châteauneuf, J.-J., & Gruas-Cavagnetto, C. (1978). Les zones de Wetzeliellaceae (Dinophyceae) du bassin de Paris. Comparaison et corrélations avec les zones du Paléogène des bassins du nord-ouest de l'Europe. Bulletin du Bureau de Recherches Géologiques et Minières (2. Série), Section IV, 2, 59–93.
Châteauneuf, J.-J., & Ménillet, F. (2014). Découverte d’une microflore bartonienne dans le Fossé rhénan supérieur: la formation de Mietesheim (Bas-Rhin, Alsace, France). Géologie de la Fance, 2014(1), 3–20.
Costa, L. I., & Downie, C. (1976). The distribution of the dinoflagellate Wetzeliella in the Palaeogene of north-western Europe. Palaeontology, 19(4), 591–614.
Costa, L. I., & Downie, C. (1979). The Wetzeliellaceae; Palaeogene dinoflagellates. In D. C. Bharadwaj, H. P. Singh, & R. S. Tiwari (Eds.), Proceedings of the 4th International Palynological Conference, Lucknow (1976–77) Vol. 2 (pp. 34–46). Lucknow: Birbal Sahni Institute of Palaeobotany.
Coughlan, N. E., Kelly, T. C., Davenport, J., & Jansen, M. A. K. (2015). Humid microclimates within the plumage of mallard ducks (Anas platyrhynchos) can potentially facilitate long distance dispersal of propagules. Acta Oecologica, 65–66, 17–23.
Coughlan, N. E., Stevens, A. L., Kelly, T. C., Dick, J. T. A., & Jansen, M. A. K. (2017). Zoochorous dispersal of freshwater bivalves: an overlooked vector in biological invasions? Knowledge and Management of Aquatic Ecosystems, 418(42). https://doi.org/10.1051/kmae/2017037.
Crouch, E. M., Heilmann-Clausen, C., Brinkhuis, H., Morgans, H. E. G., Rogers, K. M., Egger, H., & Schmitz, B. (2001). Global dinoflagellate event associated with the late Paleocene Thermal Maximum. Geology, 29(4), 315–318.
Dale, B. (1996). Dinoflagellate cyst ecology: Modeling and geological applications. In J. Jansonius, & D. C. McGregor (Eds.), Palynology: Principles and applications Vol. 3 (pp. 1249–1275). Dallas: AASP Foundation.
Darwin, C. (1859). On the origin of species by means of natural selection, or the preservation of favoured races in the struggle for life (pp. X + 502). London: John Murray.
Darwin, C. (1882). On the dispersal of freshwater bivalves. Nature, 25(649), 529–530.
Dašková, J., Konzalová, M., & Cílek, V. (2011). Tracing of palynomorphs in the Eastern Slovakian Karst. Acta Musei Nationalis Pragae Series B — Historia Naturalis, 67(1–2), 51–62.
De Coninck, J. (1976). Microfossiles à paroi organique de l'Yprésien du Bassin Belge. Service Géologique de Belgique, Professional Paper, 1975(12), 1–151.
De Coninck, J. (1977). Organic walled microfossils from the Eocene of the Woensdrecht borehole, southern Netherlands. Mededelingen Rijks Geologische Dienst, Nieuwe Serie, 28(3), 33–64.
De Coninck, J. (1985). Microfossiles à paroi organique dans les Sables de Lede (Éocène moyen) du sondage de Mol (Belgique). Bulletin de la Société Belge de Géologie, 94(1), 65–78.
De Coninck, J. (1986). Organic walled phytoplankton from the Bartonian and Eo-Oligocene transitional deposits of the Woensdrecht Borehole, southern Netherlands. Mededelingen Rijks Geologische Dienst, 40(2), 1–49.
De Coninck, J. (1991). Ypresian organic-walled phytoplankton in the Belgian Basin and adjacent areas. Bulletin de la Société Belge de Géologie, 97(3–4), 287–319.
De Coninck, J. (1995). Microfossiles à paroi organique du Bartonien, Priabonien et Rupélien inférieur dans le sondage de Kallo; espèces significatives dans les sondages de Woensdrecht, Kallo et Mol. Mededelingen Rijks Geologische Dienst, 53, 65–105.
Deming, D. (1994). Overburden rock, temperature, and heat flow. In L. B. Magoon, & W. G. Dow (Eds.), The petroleum system – from source to trap (pp. 165–186). Tulsa, OK: AAPG.
Denison, C. N. (2021). Stratigraphic and sedimentological aspects of the worldwide distribution of Apectodinium in Paleocene/Eocene Thermal Maximum deposits. In F. Marret, J. O’Keefe, P. Osterloff, M. Pound, & L. Shumilovskikh (Eds.), Applications of non-pollen palynomorphs: from palaeoenvironmental reconstructions to biostratigraphy (pp. 269–308). London: Geological Society.
Derer, C. E. (2003). Tectono-sedimentary evolution of the northern Upper Rhine Graben (Germany), with special regard to the early syn-rift stage (pp. II + 103). Dissertation Universität Bonn.
Dèzes, P., Schmid, S. M., & Ziegler, P. A. (2004). Evolution of the European Cenozoic Rift System: interaction of the Alpine and Pyrenean orogens with their foreland lithosphere. Tectonophysics, 389(1–2), 1–33.
Doebl, F. (1958). Stratigraphische und paläogeographische Ergebnisse neuerer mikropaläontologischer Untersuchungen im Tertiär des Rheintal-Grabens. Erdöl und Kohle, 11(6), 373–376.
Doebl, F. (1962). Foraminifera. In Arbeitskreis deutscher Mikropaläontologen (Ed.), Leitfossilien der Mikropaläontologie (pp. 385–391). Berlin: Borntraeger.
Doebl, F. (1970). Die tertiären und quartären Sedimente des südlichen Rheingrabens. In J. H. Illies, & S. Mueller (Eds.), Graben problems (pp. 56–66). Stuttgart: Schweizerbart.
Doebl, F., & Bader, M. (1970). Die Geologie des Gebietes der Kleinen Kalmit (westlich Landau/Pfalz) zur Zeit des Tertiärs. Mitteilungen der Pollichia, III. Reihe, 17, 14–23.
Doebl, F., Heling, D., Homann, W., Karweil, J., Teichmüller, M., & Welte, D. (1974). Diagenesis of Tertiary clayey sediments and included dispersed organic matter in relationship to geothermics in the Upper Rhine Graben. In J. H. Illies, & K. Fuchs (Eds.), Approaches to taphrogenesis (pp. 192–207). Stuttgart: Schweizerbart.
Downie, C., Hussain, M. A., & Williams, G. L. (1971). Dinoflagellate cyst and acritarch associations in the Paleogene of Southeast England. Geoscience and Man, 3, 29–35.
Drexler, E. (1958). Foraminiferen und Ostracoden aus dem Lias α von Siebeldingen/Pfalz. Geologisches Jahrbuch, 75, 475–554.
Duringer, P. (1995). Dynamik der detritischen Ablagerungen am Rande des Oberrheingrabens (Obereozän–Unteroligozän) (Exkursion G am 21. April 1995). Jahresberichte und Mitteilungen des Oberrheinischen Geologischen Vereins, Neue Folge, 77, 167–200.
Duringer, P., Aichholzer, C., Orciani, S., & Genter, A. (2019). The complete lithostratigraphic section of the geothermal wells in Rittershoffen (Upper Rhine Graben, eastern France): a key for future geothermal wells. BSGF – Earth Sciences Bulletin, 190(1). https://doi.org/10.1051/bsgf/2019012.
Duringer, P., & Gall, J.-C. (1994). Morphologie des constructions microbiennes en contexte de fan-delta Oligocène. Exemple du rift rhénan (Europe occidentale). Palaeogeography, Palaeoclimatology, Palaeoecology, 107(1–2), 35–47.
Eaton, G. L. (1976). Dinoflagellate cysts from the Bracklesham Beds (Eocene) of the Isle of Wight, southern England. Bulletin of the British Museum (Natural History) Geology, 26(6), 227–332.
Echelle, A. A., & Grande, L. (2014). Lepisosteidae: Gars. In M. L. Warren, Jr., & B. M. Burr (Eds.), Freshwater fishes of North America, Vol. 1, Petromyzontidae to Catostomidae (pp. 243–278). Baltimore: Johns Hopkins University Press.
Eldrett, J. S., Harding, I. C., Firth, J. V., & Roberts, A. P. (2004). Magnetostratigraphic calibration of Eocene–Oligocene dinoflagellate cyst biostratigraphy from the Norwegian–Greenland Sea. Marine Geology, 204(1–2), 91–127.
Farrimond, P., Taylor, A., & Telnæs, N. (1998). Biomarker maturity parameters: the role of generation and thermal degradation. Organic Geochemistry, 29(5–7), 1181–1197.
Faulkner, P. C., Burleson, M. L., Simonitis, L., Marshall, C. D., Hala, D., & Petersen, L. H. (2018). Effects of chronic exposure to 12‰ saltwater on the endocrine physiology of juvenile American alligator (Alligator mississippiensis). Journal of Experimental Biology, 221(14). https://doi.org/10.1242/jeb.181172.
Fensome, R. A., Williams, G. L., & MacRae, R. A. (2019). The Lentin and Williams index of fossil dinoflagellates 2019 edition. AASP Contributions Series, 50, 1–1173.
Fitzgerald, J. A. (1999). Pollen and spore assemblages from the Oligocene Lough Neagh Group and Dunaghy Formation, Northern Ireland (pp. VIII + 380). Unpublished PhD thesis University of Sheffield.
Fontes, J.-C., Filly, A., Gaudant, J., & Duringer, P. (1991). Origine continentale des évaporites paléogènes de Haute Alsace: arguments paléoécologiques, sédimentologiques et isotopiques. Bulletin de la Société Géologique de France, 162(4), 725–737.
Frederiksen, N. O. (1985). Review of Early Tertiary sporomorph palaeoecology. AASP Contributions Series, 15, 1–91.
Frey, M., Weinert, S., Bär, K., Vaart, J. van der, Dezayes, C., Calcagno, P., & Sass, I. (2021). Integrated 3D geological modelling of the northern Upper Rhine Graben by joint inversion of gravimetry and magnetic data. Tectonophysics, 813. https://doi.org/10.1016/j.tecto.2021.228927.
Frey, M., Bär, K., Stober, I., Reinecker, J., Vaart, J. van der, & Sass, I. (2022). Assessment of deep geothermal research and development in the Upper Rhine Graben. Geothermal Energy, 10(18). https://doi.org/10.1186/s40517-022-00226-2.
Frieling, J., Iakovleva, A. I., Reichart, G.-J., Aleksandrova, G. N., Gnibidenko, Z. N., Schouten, S., & Sluijs, A. (2014). Paleocene–Eocene warming and biotic response in the epicontinental West Siberian Sea. Geology, 42(9), 767–770.
Frieling, J., & Sluijs, A. (2018). Towards quantitative environmental reconstructions from ancient non-analogue microfossil assemblages: Ecological preferences of Paleocene–Eocene dinoflagellates. Earth-Science Reviews, 185, 956–973.
Gebhardt, H., Ćorić, S., Darga, R., Briguglio, A., Schenk, B., Werner, W., Andersen, N., & Sames, B. (2013). Middle to Late Eocene paleoenvironmental changes in a marine transgressive sequence from the northern Tethyan margin (Adelholzen, Germany). Austrian Journal of Earth Sciences, 106(2), 45–72.
Gély, J.-P. (2008). La stratigraphie et la paléogéographie du Lutétien en France. In D. Merle (Coord.), Stratotype Lutétien (pp. 182–227). Paris: Muséum national d’Histoire naturelle, BRGM.
Góczán, F., Groot, J. J., Krutzsch, W., & Pacltová, B. (1967). Die Gattungen des “Stemma Normapolles Pflug 1953b” (Angiospermae). Neubeschreibung und Revision europäischer Formen (Oberkreide bis Eozän). Paläontologische Abhandlungen B, 2(3), 429–539.
Gökçen, N. (1973). Age and lateral variations in environmental conditions of the Pinarhisan Formation north-northeastern Turkish Thrace. In Congress of Earth Sciences on the Occasion of the 50th Anniversary of the Turkish Republic (pp. 131–145). Ankara: Mineral Research and Exploration Institute.
Grande, L. (2010). An empirical synthetic pattern study of gars (Lepisosteiformes) and closely related species, based mostly on skeletal anatomy. The resurrection of holostei (pp. X + 871). American Society of Ichthyologists and Herpetologists Special Publication, 6.
Green, A. J., & Figuerola, J. (2005). Recent advances in the study of long-distance dispersal of aquatic invertebrates via birds. Diversity and Distributions, 11(2), 149–156.
Grimm, E. C. (1993). TILIA: A Pollen Program for Analysis and Display. Springfield: Illinois State Museum.
Grimm, E. C. (2004). TGView. Version 2.0.2. Springfield: Illinois State Museum Research Collection Center.
Grimm, K. I., Radtke, G., Köthe, A., Reichenbacher, B., Schwarz, J., Martini, E., Kadolsky, D., Hottenrott, M., & Franzen, J. L. (2011a). Regionale Biostratigraphie. Schriftenreihe der Deutschen Gesellschaft für Geowissenschaften, 75, 43–56.
Grimm, M. C., Wielandt-Schuster, U., Hottenrott, M., Grimm, K. I., & Radke, G., with contributions by Berger, J.-P., Ellwanger, D., Harms, F.-J., Hoselmann, C., Picot, L., & Weidenfeller, M. (2011b). Oberrheingraben. Schriftenreihe der Deutschen Gesellschaft für Geowissenschaften, 75, 57–132.
Guy-Haim, T., Hyams-Kaphzan, O., Yeruham, E., Almogi-Labin, A., & Carlton, J. T. (2017). A novel marine bioinvasion vector: Ichthyochory, live passage through fish. Limnology and Oceanography Letters, 2(3), 80–89.
Guy-Ohlson, D. (1992). Botryococcus as an aid in the interpretation of palaeoenvironment and depositional processes. Review of Palaeobotany and Palynology, 71(1–4), 1–15.
Harland, R. (1979). The Wetzeliella (Apectodinium) homomorpha plexus from the Palaeogene/earliest Eocene of North-west Europe. In D. C. Bharadwaj, H. P. Singh, & R. S. Tiwari (Eds.), Proceedings of the 4th International Palynological Conference, Lucknow (1976–77) Vol. 2 (pp. 59–70). Lucknow: Birbal Sahni Institute of Palaeobotany.
Hartkopf, C. (1981). Ein Beitrag zur Geologie und Tektonik in der Umgebung von Albersweiler/Pfalz (pp. IV + 82). Unpublished Diplomkartierung Johannes Gutenberg-Universität Mainz.
Hartkopf-Fröder, C., Kloppisch, M., Mann, U., Neumann-Mahlkau, P., Schaefer, R. G., & Wilkes, H. (2007). The end-Frasnian mass extinction in the Eifelian Mountains, Germany: new insights from organic matter composition and preservation. In R. T. Becker, & W. T. Kirchgasser (Eds.), Devonian events and correlation (pp. 173–196). London: Geological Society.
Hartkopf-Fröder, C., Königshof, P., Littke, R., & Schwarzbauer, J. (2015). Optical thermal maturity parameters and organic geochemical alteration at low grade diagenesis to anchimetamorphism: A review. International Journal of Coal Geology, 150–151, 74–119.
Haubold, H. (1989). Die Referenzfauna des Geiseltalium, MP levels 11 bis 13 (Mitteleozän, Lutetium). Palaeovertebrata, 19(3), 81–93.
Heidtke, U. H. J. (2006). Fossilien aus dem Lias von Siebeldingen (Pfalz). Pollichia-Kurier, 22(2), 19–22.
Heilmann-Clausen, C. (1985). Dinoflagellate stratigraphy of the uppermost Danian to Ypresian in the Viborg 1 borehole, central Jylland, Denmark. Danmarks Geologiske Undersøgelse, Serie A, 7, 1–69.
Heilmann-Clausen, C. (2020). Observations of the dinoflagellate Wetzeliella in Sparnacian facies (Eocene) near Epernay, France, and a note on tricky acmes of Apectodinium. Proceedings of the Geologists’ Association, 131(5), 450–457.
Heilmann-Clausen, C., & Costa, L. I. (1989). Dinoflagellate zonation of the uppermost Paleocene? to Lower Miocene in the Wursterheide Research Well, NW Germany. Geologisches Jahrbuch A, 111, 431–521.
Heilmann-Clausen, C., & Simaeys, S. van (2005). Dinoflagellate cysts from the Middle Eocene to ?lowermost Oligocene succession in the Kysing Research Borehole, central Danish Basin. Palynology, 29, 143–204.
Hinsken, S., Ustaszewski, K., & Wetzel, A. (2007). Graben width controlling syn-rift sedimentation: the Palaeogene southern Upper Rhine Graben as an example. International Journal of Earth Sciences, 96(6), 979–1002.
Hochuli, P. A. (1984). Correlation of Middle and Late Tertiary sporomorph assemblages. Paléobiologie Continentale, 14(2), 301–314.
Hottenrott, M. (1998). Eine eozäne Mikroflora aus dem Eisenberger Becken (Nordpfalz, Deutschland). Mainzer Naturwissenschaftliches Archiv, Beiheft, 21, 65–69.
Hottenrott, M. (2000). Palynologische Altersdatierungen im Tertiär des Eisenberger Beckens (Nordpfalz). Jahresberichte und Mitteilungen des Oberrheinischen Geologischen Vereins, Neue Folge, 82, 261–277.
Hottenrott, M. (2002). Neue palynologische Daten zur stratigraphischen Einstufung der älteren Tonserie (Unteres Tonlager, Mittel-Eozän) im Westerwald. Courier Forschungsinstitut Senckenberg, 237, 69–75.
Huyghe, D., Lartaud, F., Emmanuel, L., Merle, D., & Renard, M. (2015). Palaeogene climate evolution in the Paris Basin from oxygen stable isotope (δ18O) compositions of marine molluscs. Journal of the Geological Society, 172(5), 576–587.
Illies, H. (1963). Der Westrand des Rheingrabens zwischen Edenkoben (Pfalz) und Niederbronn (Elsaß). Oberrheinische Geologische Abhandlungen, 12, 1–23.
Islam, M. A. (1982). Dinoflagellate age of the boundary between Ieper and Panisel Formations (Early Eocene) at Egem, Belgium, and its significance. Neues Jahrbuch für Geologie und Paläontologie Monatshefte, 1982(8), 485–490.
Islam, M. A. (1983a). Dinoflagellate cyst taxonomy and biostratigraphy of the Eocene Bracklesham Group in southern England. Micropaleontology, 29(3), 328–353.
Islam, M. A. (1983b). Dinoflagellate cysts from the Eocene of the London and the Hampshire basins, southern England. Palynology, 7, 71–92.
Jan du Chêne, R. (1977). Nouvelles données sur la palynostratigraphie des Flyschs des Préalpes externes (Suisse). Archives des Sciences Genève, 30(1), 53–63.
Jan du Chêne, R., Gorin, G., & van Stuijvenberg, J. (1975). Étude géologique et stratigraphique (palynologie et nannoflore calcaire) des Grès des Voirons (Paléogène de Haute-Savoie, France). Géologie Alpine, 51, 51–78.
Jüngst, H. (1938). Der Pfälzer Lias und seine paläogeographische Bedeutung. Mitteilung des Saarpfälzischen Vereins für Naturkunde und Naturschutz, Pollichia, Neue Folge, 7, 145–168.
Kadolsky, D. (2015). Lutetiella, a new genus of hydrobioids from the Middle Eocene (Lutetian) of the Upper Rhine Graben and Paris Basin (Mollusca: Gastropoda: Rissooidea s. lat.). Geologica Saxonica, 61(1), 35–51.
Kamptner, E. (1930). Die Kalkflagellaten des Süßwassers und ihre Beziehungen zu jenen des Brackwassers und des Meeres. Internationale Revue der Gesamten Hydrobiologie und Hydrographie, 24(1–2), 147–163.
Keen, M. (1977). Ostracod assemblages and the depositional environments of the Headen, Osborne and Bembridge Beds (Upper Eocene) of the Hampshire Basin. Palaeontology, 20(2), 405–445.
Keen, M. (1990). The ecology and evolution of the Palaeogene ostracod Neocyprideis. Courier Forschungsinstitut Senckenberg, 123, 217–228.
Keller, T., & Schaal, S. (1988). Krokodile – urtümliche Großechsen. In S. Schaal, & W. Ziegler (Eds.), Messel – Ein Schaufenster in die Geschichte der Erde und des Lebens (pp. 109–118). Frankfurt a. M.: Kramer.
King, C., Gale, A. S., & Barry, T. L. (2016). A revised correlation of Tertiary rocks in the British Isles and adjacent areas of NW Europe. Geological Society, London, Special Reports, 27, 1–724.
Klumpp, B. (1953). Beiträge zur Kenntnis der Mikrofossilien des Mittleren und Oberen Eozän. Palaeontographica A, 103(5–6), 377–406.
Köthe, A. (1990). Paleogene dinoflagellates from Northwest Germany – biostratigraphy and paleoenvironment. Geologisches Jahrbuch A, 118, 3–111.
Köthe, A., & Piesker, B. (2007). Stratigraphic distribution of Paleogene and Miocene dinocysts in Germany. Revue de Paléobiologie, 26(1), 1–39.
Koot, M. B., Cuny, G., Tintori, A., & Twitchett, R. J. (2013). A new diverse shark fauna from the Wordian (middle Permian) Khuff Formation in the interior Haushi-Huqf area, Sultanate of Oman. Palaeontology, 56(2), 303–343.
Krutzsch, W. (1959). Mikropaläontologische (sporenpaläontologische) Untersuchungen in der Braunkohle des Geiseltales. I. Die Sporen und die sporenartigen sowie ehemals im Geiseltal zu Sporites gestellten Formeinheiten der Sporae dispersae der mitteleozänen Braunkohle des mittleren Geiseltales (Tagebau Neumark-West i. w. S.), unter Berücksichtigung und Revision weiterer Sporenformen aus der bisherigen Literatur. Geologie, 8, Beiheft, 21/22, 1–425.
Krutzsch, W. (1966). Die sporenstratigraphische Gliederung des älteren Tertiär im nördlichen Mitteleuropa (Paleozän-Mitteloligozän). Methodische Grundlagen und gegenwärtiger Stand der Untersuchungen. Abhandlungen des Zentralen Geologischen Instituts, 8, 112–149.
Krutzsch, W. (1967). Der Florenwechsel im Alttertiär Mitteleuropas auf Grund von sporenpaläontologischen Untersuchungen. Abhandlungen des Zentralen Geologischen Instituts, 10, 17–37.
Krutzsch, W. (1970). Atlas der mittel- und jungtertiären dispersen Sporen- und Pollen- sowie der Mikroplanktonformen des nördlichen Mitteleuropas. Lieferung VII: Monoporate, monocolpate, longicolpate, dicolpate und ephedroide (polyplicate) Pollenformen (pp. 1-175). Berlin: VEB Gustav Fischer Verlag.
Krutzsch, W. (1992). Paläobotanische Klimagliederung des Alttertiärs (Mitteleozän bis Oberoligozän) in Mitteldeutschland und das Problem der Verknüpfung mariner und kontinentaler Gliederungen (klassische Biostratigraphien – paläobotanisch-ökologische Klimastratigraphie – Evolutions-Stratigraphie der Vertebraten). Neues Jahrbuch für Geologie und Paläontologie Abhandlungen, 186(1–2), 137–253.
Krutzsch, W. (2008). Die Bedeutung der fossilen Pollengattung Mediocolpopollis Krutzsch 1959 (fam. Santalaceae) für die Gliederung des Obereozän im mitteldeutschen Ästuar. Hallesches Jahrbuch für Geowissenschaften, Beiheft, 25, 1–103.
Lavoyer, T. (2013). Paléontologie et stratigraphie de la partie nord du fossé rhénan supérieur moyen au cours du Paléogène: relations entre le système du rift, les transgressions marines et le paléoclimat. GeoFocus, 35, 1–210.
Lenz, O. K. (2000). Paläoökologie eines Küstenmoores aus dem Eozän Mitteleuropas am Beispiel der Wulfersdorfer Flöze und deren Begleitschichten (Helmstedter Oberflözgruppe, Tagebau Helmstedt) (pp. II + 230). Dissertation Universität Göttingen.
Lenz, O. K. (2005). Palynologie und Paläoökologie eines Küstenmoores aus dem Mittleren Eozän Mitteleuropas – Die Wulfersdorfer Flözgruppe aus dem Tagebau Helmstedt, Niedersachsen. Palaeontographica B, 271(1–6), 1–157.
Lenz, O. K., & Riegel, W. (2001). Isopollen maps as a tool for the reconstruction of a coastal swamp from the Middle Eocene at Helmstedt (Northern Germany). Facies, 45(1), 177–194.
Lenz, O. K., Riegel, W., & Wilde, V. (2021). Greenhous conditions in lower Eocene coastal wetlands? – Lessons from Schöningen, Northern Germany. PloS One, 16(1). https://doi.org/10.1371/journal.pone.0232861.
Lenz, O. K., & Wilde, V. (2018). Changes in Eocene plant diversity and composition of vegetation: the lacustrine archive of Messel (Germany). Paleobiology, 44(4), 709–735.
Lenz, O. K., Wilde, V., Mertz, D. F., & Riegel, W. (2015). New palynology-based astronomical and revised 40Ar/39Ar ages for the Eocene maar lake of Messel (Germany). International Journal of Earth Sciences, 104(3), 873–889.
Lenz, O. K., Wilde, V., & Riegel, W. (2007). Recolonization of a Middle Eocene volcanic site: quantitative palynology of the initial phase of the maar lake of Messel (Germany). Review of Palaeobotany and Palynology, 145(3–4), 217–242.
Lenz, O. K., Wilde, V., & Riegel, W. (2011). Short-term fluctuations in vegetation and phytoplankton during the Middle Eocene greenhouse climate: a 640-kyr record from the Messel oil shale (Germany). International Journal of Earth Sciences, 100(8), 1851–1874.
Liengjarern, M., Costa, L., & Downie, C. (1980). Dinoflagellate cysts from the Upper Eocene–Lower Oligocene of the Isle of Wight. Palaeontology, 23(3), 475–499.
Löschan, G., Emmerich, K., Reinhold, C., & Reinecker, J. (2017). Clay mineralogy of Tertiary formations in the northern Upper Rhine Graben – New insights from geothermal and hydrocarbon exploration. Zeitschrift der Deutschen Gesellschaft für Geowissenschaften, 168(2), 233–244.
Lopes Cardozo, G. G. O., & Behrmann, J. H. (2006). Kinematic analysis of the Upper Rhine Graben boundary fault system. Journal of Structural Geology, 28(6), 1028–1039.
Lovas-Kiss, Á., Vincze, O., Löki, V., Pallér-Kapusi, F., Halasi-Kovács, B., Kovács, G., Green, A. J., & Lukács, B. A. (2020). Experimental evidence of dispersal of invasive cyprinid eggs inside migratory waterfowl. Proceedings of the National Academy of Sciences of the United States of America, 117(27), 15397–15399.
Markwick, P. J. (1998). Fossil crocodilians as indicators of Late Cretaceous and Cenozoic climates: implications for using palaeontological data in reconstructing palaeoclimate. Palaeogeography, Palaeoclimatology, Palaeoecology, 137(3–4), 205–271.
Marshall, J. E. A., & Yule, B. L. (1999). Spore colour measurement. In T. P. Jones, & N. P. Rowe (Eds.), Fossil plants and spores: modern techniques (pp. 165–168). London: Geological Society London.
Martini, E. (1958). Discoasteriden und verwandte Formen im NW-deutschen Eozän (Coccolithophorida). 1. Taxionomische Untersuchungen. Senckenbergiana lethaea, 39(5–6), 353–388.
Martini, E. (1988). Nannoplankton-Massenvorkommen in den Corbicula- (Schichten mit Hydrobia inflata) und Hydrobienschichten des Oberrheingrabens, des Mainzer und des Hanauer Beckens (Miozän). Geologisches Jahrbuch A, 110, 205–227.
Martini, E. (1990). The Rhinegraben system, a connection between northern and southern seas in the European Tertiary. Veröffentlichungen Übersee-Museum A, 10, 83–98, 208–211.
Martini, E. (1991). Endemische Vertreter der Gattung Trochoaster (kalkiges Nannoplankton) im westdeutschen Unter-Oligozän. Geologisches Jahrbuch A, 128, 167–177.
Martini, E., & Reichenbacher, B. (2007). Nannoplankton und Fisch-Otolithen in den Mittleren Pechelbronn-Schichten (Unter-Oligozän, Oberrheingraben/Mainzer Becken). Geologische Abhandlungen Hessen, 116, 235–273.
Martini, E., & Rothe, P. (Eds.) (1998). Die alttertiäre Fossillagerstätte Sieblos an der Wasserkuppe/Rhön. Geologische Abhandlungen Hessen, 104, 1–274.
Martini, E., & Schiller, W. (1998). Kalkiges Nannoplankton von Sieblos/Rhön und dem Neuwieder Becken (Unter-Oligozän). Geologische Abhandlungen Hessen, 104, 165–172.
Martini, E., & Stradner, H. (1960). Nannotetraster, eine stratigraphisch bedeutsame neue Discoasteridengattung. Erdoel-Zeitschrift, 76(8), 266–270.
Marynowski, L., & Zatoń, M. (2010). Organic matter from the Callovian (Middle Jurassic) deposits of Lithuania: Compositions, sources and depositional environments. Applied Geochemistry, 25(7), 933–946.
Mayr, G. (2017). Avian evolution. The fossil record of birds and its paleobiological significance (pp. XIII + 309). Chichester: Wiley.
Mazzotti, F. J., & Dunson, W. A. (1989). Osmoregulation in crocodilians. American Zoologist, 29(3), 903–920.
Micklich, N. (2018). Actinopterygians – the fishes of the Messel Lake. In K. T. Smith, S. F. K. Schaal, & J. Habersetzer (Eds.), Messel – An ancient greenhouse ecosystem (pp. 104–111). Stuttgart: Schweizerbart.
Moline, P. M., & Linder, H. P. (2005). Molecular phylogeny and generic delimitation in the Elegia group (Restionaceae, South Africa) based on a complete taxon sampling and four chloroplast DNA regions. Systematic Botany, 30(4), 759–772.
Moshayedi, M., Lenz, O. K., Wilde, V., & Hinderer, M. (2018). Controls on sedimentation and vegetation in an Eocene pull-apart basin (Prinz von Hessen, Germany): evidence from palynology. Journal of the Geological Society, 175(5), 757–773.
Moshayedi, M., Lenz, O. K., Wilde, V., & Hinderer, M. (2020). The recolonisation of volcanically disturbed Eocene habitats of Central Europe: the maar lakes of Messel and Offenthal (SW Germany) compared. Palaeobiodiversity and Palaeoenvironments, 100(4), 951–973.
Moshayedi, M., Lenz, O. K., Wilde, V., & Hinderer, M. (2021). Lake-level fluctuations and allochthonous lignite deposition in the Eocene pull-apart basin “Prinz von Hessen” (Hesse, Germany) – a palynological study. In M. R. Rosen, D. B. Finkelstein, L. Park Boush, & S. Pla-Pueyo (Eds.), Limnogeology: Progress, challenges and opportunities (pp. 69–106). Cham: Springer.
Mutzl, J., Lenz, O. K., Wilde, V., Krahn, K. J., Moshayedi, M., & Hinderer, M. (2022). Vegetation dynamics in a disturbed lacustrine record: The Eocene maar lake of Groß-Zimmern (Hesse, SW Germany). Palaeobiodiversity and Palaeoenvironments. https://doi.org/10.1007/s12549-022-00543-1.
Nottmeyer, D. (1954). Stratigraphische und tektonische Untersuchungen in der rheinischen Vorbergzone bei Siebeldingen–Frankweiler. Mitteilungen der Pollichia, 43, 36–93.
Oertli, H. J. (1963). Fossile Ostracoden als Milieuindikatoren. Fortschritte in der Geologie von Rheinland und Westfalen, 10, 53–66.
Ohmert, W. (1993). Eine obereozäne Foraminiferenfauna aus dem südlichen Oberrhein-Graben. Zitteliana, 20, 323–329.
Ohmert, W. (2014). Ostracoden aus dem eozänen Lymnäenmergel (Haguenau-Formation) der Bohrung Oberweiler (Gemeinde Badenweiler, südlicher Oberrhein). Berichte der Naturforschenden Gesellschaft zu Freiburg im Breisgau, 104, 131–168.
Ohmert, W. (2017). Mikrofaunen der “Versteinerungsreichen Zone” (Mittlere Pechelbronn-Formation, Unter-Oligozän), ihre stratigraphische und regionale Verteilung am östlichen Oberrheingraben-Rand (Südwestdeutschland). Zitteliana, 89, 39–111.
Page, C. N. (1990). Key to families of Coniferophytina. In K. U. Kramer, & P. S. Green (Eds.), The families and genera of vascular plants, Vol. 1, Pteridophytes and Gymnosperms (p. 283). Berlin: Springer.
Pearson, D. L. (1990). Pollen/spore color “standard“. 2nd Printing of Version #2. Phillips Petroleum Company.
Perner, M., Jäger, H., Reinhold, C., Bechstädt, T., & Stinnesbeck, W. (2018). Impact of rift dynamics on palaeoenvironmental conditions and hydrocarbon system development (northern Upper Rhine Graben, SW-Germany). Petroleum Geoscience, 24(4), 425–439.
Peters, K. E., & Cassa, M. R. (1994). Applied source rock geochemistry. In L. B. Magoon, & W. G. Dow (Eds.), The petroleum system – from source to trap (pp. 93–120). Tulsa, OK: AAPG.
Peters, K. E., Walters, C. C., & Moldowan, J. M. (2005). The biomarker guide, 2nd ed. (pp. XXXVI + 1155). Cambridge: Cambridge University Press.
Pirkenseer, C., Rauber, G., & Roussé, S. (2018). A revised Palaeogene lithostratigraphic framework for the northern Swiss Jura and the southern Upper Rhine Graben and its relationship to the North Alpine Foreland Basin. Rivista Italiana di Paleontologia e Stratigrafia, 124(1), 163–246.
Pirkenseer, C., Spezzaferri, S., & Berger, J.-P. (2011). Reworked microfossils as a paleogeographic tool. Geology, 39(9), 843–846.
Pross, J., & Brinkhuis, H. (2005). Organic-walled dinoflagellate cysts as paleoenvironmental indicators in the Paleogene; a synopsis of concepts. Paläontologische Zeitschrift, 79(1), 53–59.
Rauscher, R., & Schuler, M. (1988). Les dinokystes, des témoins d’influences marines dans le Paléogène d’Alsace. Bulletin des Centres de Recherches Exploration-Production Elf-Aquitaine, 12(1), 405–425.
Rauscher, R., Schuler, M., & Sittler, C. (1988). Les dinokystes dans le Paléogène d’Alsace: le problème de l’origine du sel. Comptes Rendus de l’Académie des Sciences, Série II, 307, 175–178.
Reichenbacher, B., & Keller, T. (2002). Neudefinition von stratigraphischen Einheiten im Tertiär des Mainzer und Hanauer Beckens (Deutschland, Oligozän–Miozän), Teil 2: Wiesbaden-Formation (= Untere Hydrobien-Schichten). Mainzer Geowissenschaftliche Mitteilungen, 31, 99–122.
Reichenbacher, B., Gaudant, J., & Griessemer, T. W. (2007). A late Burdigalian gobiid fish, Gobius brevis (Agassiz, 1839), in the Upper Hydrobia Beds in the middle Upper Rhine Graben (W-Germany). Paläontologische Zeitschrift, 81(4), 365–375.
Reis, O. M. (1923). Kalkalgen und Seesinterkalke aus dem rheinpfälzischen Tertiär. Geognostische Jahreshefte, 36, 103–130.
Reischmann, T., & Anthes, G. (1996). Geochronology of the mid-German crystalline rise west of the river Rhine. Geologische Rundschau, 85(4), 761–774.
Rich, F. J., Kuehn, D., & Davies, T. D. (1982). The paleoecological significance of Ovoidites. Palynology, 6, 19–28.
Riegel, W., Wilde, V., & Lenz, O. K. (2012). The Early Eocene of Schöningen (N-Germany) – an interim report. Austrian Journal of Earth Sciences, 105(1), 88–109.
Rivas Jr., J. A., Schröder, T., Gill, T. E., Wallace, R. L., & Walsh, E. J. (2019). Anemochory of diapausing stages of microinvertebrates in North American drylands. Freshwater Biology, 64(7), 1303–1314.
Schad, A. (1962). Das Erdölfeld Landau. Abhandlungen des Geologischen Landesamtes in Baden-Württemberg, 4, 81–101.
Schäfer, P. (2000). Zur Stratigraphie und Genese der tertiären Sedimente zwischen Eisenberg und Lautersheim im südwestlichsten Teil des Mainzer Beckens. Jahresberichte und Mitteilungen des Oberrheinischen Geologischen Vereins, Neue Folge, 82, 175–222.
Schäfer, P. (2012). Mainzer Becken. Stratigraphie – Paläontologie – Exkursionen (pp. VIII + 333). Sammlung geologischer Führer, 79.
Schäfer, P. (2013). Grabenbildung – Tertiäre Tektonik in der Pfalz. In J. Haneke, & K. Kremb (Eds.), Beiträge zur Geologie der Pfalz (pp. 35–48). Speyer: Verlag der Pfälzischen Gesellschaft zur Förderung der Wissenschaften.
Schröder, G. (1984). Ingenieurgeologische Untersuchungen an rezenten und fossilen Rutschungen im Raum Albersweiler/Birkweiler (Ostpfalz) (pp. IV + 41). Unpublished Diplomarbeit Johannes Gutenberg-Universität Mainz.
Schuler, M. (1983). Pollens et spores des Séries salifères (Éocène/Oligocène) du bassin potassique de Mulhouse (France). Relations entre la microflore et les dépôts évaporitiques. Sciences Géologiques Bulletin, 36(4), 255–265.
Schuler, M. (1990). Environnements et paléoclimats paléogènes. Palynologie et biostratigraphie de l’Éocène et de l’Oligocène inférieur dans les fossés rhénan, rhodanien et de Hesse. Document du BRGM, 190, 1–503.
Schuler, M., & Ollivier-Pierre, M.-F. (1981). Étude palynologique (pollen et spores) du Lutétien continental de Bouxwiller (Bas-Rhin, France). Sciences Géologiques Bulletin, 34(4), 219–238.
Schwarz, J., & Griessemer, T. W. (1998). Eine neue Charophyten-Art aus dem Lutetium (Mitteleozän) des Oberrheingrabens bei Albersweiler (Rheinland-Pfalz, SW-Deutschland): Chara rothauseni sp. n. Mainzer Naturwissenschaftliches Archiv, Beiheft, 21, 57–63.
Sherwood, R. W., & Levin, H. L. (1972). A closer look at Trochoaster simplex Klumpp. Journal of Paleontology, 46(4), 591–594.
Silva, G. G., Weber, V., Green, A. J., Hoffmann, P., Silva, V. S., Volcan, M. V., Lanés, L. E. K., Stenert, C., Reichard, M., & Maltchik, L. (2019). Killifish eggs can disperse via gut passage through waterfowl. Ecology, 100(11). https://doi.org/10.1002/ecy.2774.
Sissingh, W. (1998). Comparative Tertiary stratigraphy of the Rhine Graben, Bresse Graben and Molasse Basin: correlation of Alpine foreland events. Tectonophysics, 300(1–4), 249–284.
Sissingh, W. (2003). Tertiary paleogeographic and tectonostratigraphic evolution of the Rhenish Triple Junction. Palaeogeography, Palaeoclimatology, Palaeoecology, 196(1–2), 229–263.
Sissingh, W. (2006). Syn-kinematic palaeogeographic evolution of the West European Platform: correlation with Alpine plate collision and foreland deformation. Netherlands Journal of Geosciences, 85(2), 131–180.
Sittler, C. (1965). Le Paléogène des fossés rhénan et rhodanien. Études sédimentologiques et paléoclimatiques. Mémoires du Service de la Carte Géologique d’Alsace et de Lorraine, 24, 1–392.
Sittler, C. (1968a). L‘analyse pollinique dans l’est de la France. Étude des formations éocènes ou rapportées à l’Éocène et des stratotypes palynologiques de Borken et de Messel. Mémoires du Bureau de Recherches Géologiques et Minières, 58, 165–171.
Sittler, C. (1968b). Les indications stratigraphiques fournies par les minéraux argileux des différentes formations éocènes du fossé rhénan. Comparaison avec les régions voisines. Mémoires du Bureau de Recherches Géologiques et Minières, 58, 495–503.
Sittler, C. (1969). L’Éocène dans le fossé rhénan. Mémoires du Bureau de Recherches Géologiques et Minières, 69, 371–383.
Sittler, C. (1970). Die Korrelationsmöglichkeiten der ältesten tertiären Ablagerungen des Rheingrabensystems unter Berücksichtigung ihres Tonmineralbestandes. In J. H. Illies, & S. Mueller (Eds.), Graben problems (pp. 69–75). Stuttgart: Schweizerbart.
Sittler, C. (1972). Le pétrole dans le département du Haut-Rhin. Bilan d’un siècle et demi de recherches et d’exploitations. Sciences Géologiques Bulletin, 25(2–3), 151–161, 189–200.
Sittler, C. (1992). Illustration de l’histoire géologique du Fossé rhénan et de l’Alsace. Neues Jahrbuch für Geologie und Paläontologie Abhandlungen, 186(3), 255–282.
Sittler, C., & Ollivier-Pierre, M.-F. (1994). Palynology and palynofacies analyses: some essential clues to assess and identify West-European Tertiary depositional environments in terms of relative high or lowstands. Application to the case of three Eocene and Oligocene sections in France. Bulletin des Centres de Recherches Exploration-Production Elf-Aquitaine, 18(2), 475–488.
Sluijs, A., & Brinkhuis, H. (2009). A dynamic climate and ecosystem state during the Paleocene–Eocene Thermal Maximum: inferences from dinoflagellate cyst assemblages on the New Jersey Shelf. Biogeosciences, 6(8), 1755–1781.
Sluijs, A., Pross, J., & Brinkhuis, H. (2005). From greenhouse to icehouse; organic-walled dinoflagellate cysts as paleoenvironmental indicators in the Paleogene. Earth-Science Reviews, 68(3–4), 281–315.
Sluijs, A., Bowen, G. J., Brinkhuis, H., Lourens, L. J., & Thomas, E. (2007). The Palaeocene – Eocene Thermal Maximum super greenhouse: biotic and geochemical signatures, age models and mechanisms of global change. In M. Williams, A. M. Haywood, F. J. Gregory, & D. N. Schmidt (Eds.), Deep-time perspectives on climate change: Marrying the signal from computer models and biological proxies (pp. 323–349). London: The Micropalaeontological Society.
Sluijs, A., Brinkhuis, H., Williams, G. L., & Fensome, R. A. (2009): Taxonomic revision of some Cretaceous–Cenozoic spiny organic-walled peridiniacean dinoflagellate cysts. Review of Palaeobotany and Palynology, 154(1–4), 34–53.
Sluijs, A., Roij, L. van, Harrington, G. J., Schouten, S., Sessa, J. A., LeVay, L. J., Reichart, G.-J., & Slomp, C. P. (2014). Warming, euxinia and sea level rise during the Paleocene–Eocene Thermal Maximum on the Gulf Coastal Plain: implications for ocean oxygenation and nutrient cycling. Climate of the Past, 10(4), 1421–1439.
Smith, K. T., Schaal, S. F. K., & Habersetzer, J. (Eds.) (2018). Messel – An ancient greenhouse ecosystem (pp. XVI + 355). Stuttgart: Schweizerbart.
Spuhler, L. (1937). Die Marnheimer Bucht. Mitteilungen des Saarpfälzischen Vereins für Naturkunde und Naturschutz Pollichia, Neue Folge, 6, 3–59.
Stancliffe, R. P. W. (1996). Microforaminiferal linings. In J. Jansonius, & D. C. McGregor (Eds.), Palynology: Principles and applications Vol. 1 (pp. 373–379). Dallas: AASP Foundation.
Stapf, K. R. G. (1988). Zur Tektonik des westlichen Rheingrabenrandes zwischen Nierstein am Rhein und Wissembourg (Elsaß). Jahresberichte und Mitteilungen des Oberrheinischen Geologischen Vereins, Neue Folge, 70, 399–410.
Stellrecht, R. (1971). Geologisch-tektonische Entwicklung im Raum Albersweiler/Pfalz. Jahresberichte und Mitteilungen des Oberrheinischen Geologischen Vereins, Neue Folge, 53, 239–262.
Steurbaut, E., King, C., Matthijs, J., Noiret, C., Yans, J., & van Simaeys, S. (2015). The Zemst borehole, first record of the EECO in the North Sea Basin and implications for Belgian Ypresian–Lutetian stratigraphy. Geologica Belgica, 18(2–4), 147–159.
Stradner, H. (1959). Die fossilen Discoasteriden Österreichs. 2. Teil. Erdoel-Zeitschrift, 75(12), 472–488.
Straub, E. W. (1955). Über einen Fund von fraglichem Mitteleozän im Erdölfeld Stockstadt bei Darmstadt. Notizblatt des Hessischen Landesamtes für Bodenforschung zu Wiesbaden, 83, 220–227.
Taplin, L. E., & Grigg, G. C. (1989). Historical zoogeography of the eusuchian crocodilians: a physiological perspective. American Zoologist, 29(3), 885–901.
Teichmüller, M. (1970). Bestimmung des Inkohlungsgrades von kohligen Einschlüssen in Sedimenten des Oberrheingrabens – ein Hilfsmittel bei der Klärung geothermischer Fragen. In J. H. Illies, & S. Mueller (Eds.), Graben problems (pp. 124–142). Stuttgart: Schweizerbart.
Teichmüller, M. (1979). Die Diagenese der kohligen Substanzen in den Gesteinen des Tertiärs und Mesozoikums des mittleren Oberrheingrabens. Fortschritte in der Geologie von Rheinland und Westfalen, 27, 19–49.
Teichmüller, M., & Teichmüller, R. (1981). The significance of coalification studies to geology – a review. Bulletin des Centres de Recherches Exploration-Production Elf-Aquitaine, 5(2), 491–534.
Thiele-Pfeiffer, H. (1988). Die Mikroflora aus dem mitteleozänen Ölschiefer von Messel bei Darmstadt. Palaeontographica B, 211(1–3), 1–86.
Thomsen, E., Abrahamsen, N., Heilmann-Clausen, C., King, C., & Nielsen, O. B. (2012). Middle Eocene to earliest Oligocene development in the eastern North Sea Basin: Biostratigraphy, magnetostratigraphy and palaeoenvironment of the Kysing-4 borehole, Denmark. Palaeogeography, Palaeoclimatology, Palaeoecology, 350–352, 212–235.
Thomson, P. W., & Pflug, H. (1953). Pollen und Sporen des mitteleuropäischen Tertiärs. Gesamtübersicht über die stratigraphisch und paläontologisch wichtigen Formen. Palaeontographica B, 94(1–4), 1–138.
Trauth, N., Cavelier, C., Sommer, F., Tourencq, J., Pomerol, C., & Thiry, M. (1977). Aperçu sur la sédimentation paléogène du synclinal de Bouxwiller, comprise entre les marnes à Rhynchonelles (Bathonien) et le conglomérat du Bastberg (Oligocène). Sciences Géologiques Bulletin, 30(2), 91–100.
Tway, L. E., Harrison, W. E., & Zidek, J. (1986). Thermal alteration of microscopic fish remains – an initial study. Palaios, 1(1), 75–79.
Vanschoenwinkel, B., Waterkeyn, A., Nhiwatiwa, T., Pinceel, T., Spooren, E., Geerts, A., Clegg, B., & Brendonck, L. (2011). Passive external transport of freshwater invertebrates by elephant and other mud-wallowing mammals in an African savannah habitat. Freshwater Biology, 56(8), 1606–1619.
Weigelt, J. (1927). Rezente Wirbeltierleichen und ihre paläobiologische Bedeutung (pp. XVI + 227). Leipzig: Verlag Max Weg.
Weiler, W. (1961). Die Fischfauna des unteroligozänen Melanientons und des Rupeltons in der Hessischen Senke. Notizblatt des Hessischen Landesamtes für Bodenforschung zu Wiesbaden, 89, 44–65.
Weiler, W. (1963). Die Fischfauna des Tertiärs im oberrheinischen Graben, des Mainzer Beckens, des unteren Maintals und der Wetterau, unter besonderer Berücksichtigung des Untermiozäns. Abhandlungen der Senckenbergischen Naturforschenden Gesellschaft, 504, 1–75.
Wichter, L., Krauter, E., & Meiniger, W. (1988). Landslide stabilization using drainage wells, concrete dowels and anchored bore pile walls. In C. Bonnard (Ed.), Landslides. Proceedings of the Fifth International Symposium on Landslides (pp. 1023–1028). Rotterdam: Balkema.
Wilkinson, G. C., Bazley, R. A. B., & Boulter, M. C. (1980). The geology and palynology of the Oligocene Lough Neagh Clays, Northern Ireland. Journal of the Geological Society, 137(1), 65–75.
Wirth, E. (1969). Die Probleme des Eozäns im deutschen Anteil der Oberrheinebene und ihrer Randzonen. Mémoires du Bureau de Recherches Géologiques et Minières, 69, 287–306.
Worobiec, E. (2009). Middle Miocene palynoflora of the Legnica lignite deposit complex, Lower Silesia, Poland. Acta Palaeobotanica, 49(1), 5–133.
Yu-Fei Wang, Cheng-Sen Li, Collinson, M. E., Jian Lin, & Qi-Gao Sun (2003). Eucommia (Eucommiaceae), a potential biothermometer for the reconstruction of paleoenvironments. American Journal of Botany, 90(1), 1–7.
Acknowledgements
This study was initiated by the late Karl R. G. Stapf who introduced C. Hartkopf-Fröder to the geology of the Haardt during a university course. Günter Schweigert is thanked for discussions on Lower Jurassic biostratigraphy. Dietrich Kadolsky kindly commented on an earlier version of the manuscript. Oliver Hampe and Bettina Reichenbacher gave advice on the fish teeth structure and otolith fauna, respectively. Ralf Littke provided access to a Zeiss Axio Imager.M2m incident light microscope and Donka Macherey assisted with the photographs of the fish teeth cross-sections. The help of Ursula Amend, Ulrike Lux and Jörg Schardinel in producing the figures is gratefully acknowledged. The manuscript considerably benefitted from perceptive advice and constructive comments by Kirsten I. Grimm and Olaf K. Lenz. We thank Anna-Kristin Stössel and Sinje Weber for technical editing of the manuscript. We dedicate this publication to our friend and colleague Martin Hottenrott who was involved in the initial stage of this project but passed away on September 29, 2013. German palynology has suffered a sad loss by his untimely death. He is greatly missed by all of us.
Funding
No funding was received to assist with the preparation of this manuscript.
Author information
Authors and Affiliations
Author notes
Martin Hottenrott is deceased. This paper is dedicated to his memory.
- Martin Hottenrott
Corresponding author
Ethics declarations
Conflict of Interest
The authors have no relevant financial or non-financial interests to disclose.
Additional information
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.

Rights and permissions
Springer Nature or its licensor (e.g. a society or other partner) holds exclusive rights to this article under a publishing agreement with the author(s) or other rightsholder(s); author self-archiving of the accepted manuscript version of this article is solely governed by the terms of such publishing agreement and applicable law.
About this article

{kind=link}
Cite this article
Hartkopf-Fröder, C., Martini, E., Heilmann-Clausen, C. et al. Eocene sediments and a fresh to brackish water biota from the early rifting stage of the Upper Rhine Graben (west of oil field Landau, southwest Germany): implications for biostratigraphy, palaeoecology and source rock potential. Palaeobio Palaeoenv 104, 53–102 (2024). https://doi.org/10.1007/s12549-023-00577-z
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s12549-023-00577-z