
Abstract
This article compiles data on the Devonian-Carboniferous Boundary successions from countries and regions, which have not been dealt with separately in this special issue. Data derive from different palaeocontinents on a large palaeoclimatic gradient from the southern high latitudes of western Gondwana through the palaeotropics into the temperate/boreal northern latitudes of Siberia. The quality of the data is variable, but often surprisingly good, although major uncertainties can persist and traditional positions of the boundary do not withhold modern stratigraphic results and concepts. Not all these regions have been on the forefront of the DCB discussions in recent years, but they provide invaluable insights into the regional and global dimensions of uppermost Devonian and lowermost Carboniferous changes in the biosphere and geosphere. Almost globally documented are faunal turnovers and collapse of ecosystems in the Hangenberg extinctions. Those allow the placement of the DCB even if index fossils are absent or scarce. However, it also shows the regional differences, which advocate for a holistic approach to the boundary by ensuring global correlation without relying on the conceptual dominance a single taxon, which might be less powerful outside its usual facies.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
In order to fulfil the ambition of this special issue of presenting a global perspective on the Devonian-Carboniferous Boundary (DCB) succession, this contribution presents brief overviews and data for regions and countries (Fig. 1), which have not been treated in the other contributions. The reader is referred to those contributions for the Montagne Noire (Feist et al. 2021, this issue), Belgium (Denayer et al. 2021, this issue), the Carnic Alps (Spalletta et al. 2021, this issue), China (Qie et al. 2021, this issue), Czech Republic (Kumpan et al. 2021, this issue), Germany (Becker et al. 2021, this issue), Greenland (Marshall 2021, this issue), Iran (Königshof et al. 2021, this issue), Ireland (Graham and Sevastopulo 2021, this issue), Poland (Matyja et al. 2021, this issue), some parts of Russia and Kazakhstan (Kulagina et al. 2021, this issue), Sardinia (Corradini et al. 2021, this issue) and the USA (Over 2021, this issue).

Geographic overview on a modern world geography: the countries dealt with in this contribution are indicated in orange. The black dots indicate approximate positions of DCB sections in those countries. Note that a dot may represent several geographical rather close sections
These certainly short and partly incomplete overviews presented in this paper are also thought to give the interested reader a starting point for a more in-depth research on the DCB interval in those regions.
This does do not mean that these regions and sections can be neglected for our understanding of the DCB interval. In fact, they are important cornerstones and test cases that a global picture for the understanding of the geological and biological history of the DCB interval can be developed, including our search for dividing it and defining the base of the Carboniferous. This overview also shows that it seems to be likely that important discoveries are still to be made in those regions, which sometimes have only been superficially studied, and/or modern data are not available or only limited to single sections. Although presented according to modern geography, these regional overviews encompass various palaeocontinents and palaeoenvironments, from the high latitudes of western Gondwana to the equatorial and temperate seas in the northern palaeohemisphere (Fig. 2).

Palaeogeographic map of C. R. Scotese (Palaeomap Project map 65) for the time interval of the Devonian-Carboniferous Boundary with approximate position of the main sections mentioned herein. Note that the depositional settings of several sections do not correspond to the land-sea distribution of the map
The disparity in the abundance and precision of stratigraphic data has been a challenging problem for the authors of this contribution. Where possible, older data were re-interpreted and translated into modern taxonomic and stratigraphic nomenclature. The following abbreviations of genera are used in this paper. Conodonts: Bi.: Bispathodus; Br.: Branmehla; Gn.: Gnathodus; M.: Mehlina; Pa.: Palmatolepis; Po.: Polygnathus; Pr.: Protognathodus; Ps.: Pseudopolygnathus; Rh.: Rhodalepis; Si.: Siphonodella; Spores: K.: Knoxisporites; R.: Retispora; Ru.: Rugospora; V.: Verrucosisporites; W.: Waltzispora, Miospore zones: LL (lepidophyta–literatus), LE (lepidophyta–explanatus), LN (lepidophyta–nitidus), Rle (lepidophyta), LVa (lepidophyta–vallatus).
South America
Due to the palaeogeographic position of South America in high southern latitudes during the DCB interval (Fig. 2), the correlation to the biostratigraphic schemes developed in the low palaeolatidudes is often difficult. Correlation between these two realms is mainly achieved thanks to the apparent cosmopolitan nature of some miospores (e.g. Melo and Loboziak 2003; Playford 1990). Upper Devonian–lower Carboniferous strata is exposed in the intercratonic basins covering parts of western Gondwana, and arc-related and retroarc basins related to the active western Gondwana margin, nowadays mainly situated east of the Andes (Precordillera). Uppermost Devonian glacial records (e.g. diamictites) are found all over the region (e.g. Caputo et al. 2008; Isaacson et al. 2008; Lakin et al. 2016; Streel et al. 2013). Dating them has been a crucial step as they are the first markers of the climate detoriation leading to the Late Palaeozoic Ice Age. In the context of the DCB, these glacial deposits have been considered to represent time equivalents of the Rhenish Hangenberg Sandstone (e.g. Kaiser et al. 2015). However, this correlation is too simple and may only hold up for the top the glacial deposits, since theirs bases are often dated as LE miospore zone, which clearly predates the Hangenberg Sandstone.
Brazil
The Upper Devonian-Carboniferous successions have been described from several intercratonic basins. The successions of the Amazon Basin have played a key role not only in the establishment of a miospore biostratigraphic scheme for South America (e.g. Loboziak et al. 1995, 2000; Loboziak and Melo 2002; Melo and Loboziak 2003; Melo and Playford 2012), but also for the correlation into other parts of Gondwana (e.g. Loboziak et al. 1999) and most important with the palaeotropical miospore biostratigraphy of Western Europe (e.g. Clayton et al. 1977; Streel et al. 1987).
In the Amazon Basin, the Upper Cuirri Formation (diamictites, siltstones and sandstone lenses) has a Rle-LVa age (Melo and Loboziak 2003), which is the equivalent of the LE-LN interval in the zonal scheme for Western Europe. The glacial pulse ends below the base of the Carboniferous. The DCB is situated in the sandy non-glacial deposits of the basal Oriximiná Formation. In other boreholes, the boundary is placed between the two formations (Melo and Loboziak 2003). It is important to note that in the Amazon Basin, the boundary between the youngest Famennian and oldest Tournaisian miospore biozones (LVa–AL) corresponds to an erosional surface (Melo and Playford 2012). Hence, the record of the DCB interval is incomplete, and differences in the ages of the basal Oriximiná Formation (Melo and Loboziak 2003) may be rooted in this problem. From the subsurface of the neighbouring Solimões Basin, Caputo et al. (2008) correlated glaciomarine diamictites (Jaraqui Member of the Jandiatuba Formation) with the upper Curiri Formation of the Amazon Basin. However, as outlined by Lakin et al. (2016), the primary sedimentological and biostratigraphic data are difficult to access.
In the Parnaíba Basin, the uppermost Devonian–lowermost Carboniferous succession (Loboziak et al. 1992) has some interesting similarities to those of the Amazon Basin. Indeed, the upper part of the Cabeças Formation attributed to the LVa mio-spore Zone is composed of glacial and periglacial deposits (Streel et al. 2000). An unconformable contact is found when the following Longá Formation is entirely Tournaisian in age (Streel et al. 2013). However, a conformable contact is developed when the base of the Longa Formation corresponds to ‘end-Devonian transgressive marine shales’ (Streel et al. 2013). The boundary between both formations is also indicative for the retreat of the glacial facies, which took place immediately below the DCB (Caputo et al. 2008). In southeastern Brazil, uppermost Devonian diacmitites dated as LN Zone (Loboziak et al. 1995) are found in the subsurface of the Paraná Basin (Lakin et al. 2016). These so-called Ortigueira Diamictite form the top of the Ponta Grossa Formation, which is unconformably overlain by the Moscovian or younger rocks of the Itararé Group (Milani et al. 2007).
Bolivia and Peru
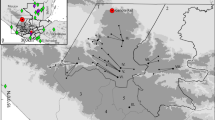
In the Bolivian Altiplano, around the Lake Titikaka, the Upper Devonian–lower Carboniferous strata are regrouped in ascending order into Colpacucho, Cumaná and Kasa formations (Fig. 3). The Upper Devonian Calpacucho Formation is composed of marine mudstones and sandstones and, in its upper part, deeper-water offshore facies dominates. The following Cumana Formation is composed of up to 100-m-thick glacial marine diamictites and shales with dropstones of latest Fammenian palynology-based age (Díaz-Martínez and Isaacson 1994; Lakin et al. 2016). The Kasa Formation represents a complex deltaic progradational sequence of Tournaisian-Visean age (Díaz-Martínez et al. 1999). As previously described from Brazil, the glacial pulse occurred just below the DCB. Palynological data (Díaz-Martínez et al. 1999; Vavrdová et al. 1993) show that the Cumaná Formation belongs to the LE Zone. The absence of Verrucosisporites nitidus does not allow to identify the LN Zone. However, Lakin et al. (2016) concluded that the extinction of Retispora lepidophyta and marine phytoplankton occurred within or immediately above the diamictites, and hence allows to place the DCB at this extinction level (= base of VI Zone). The position of the DCB above the glacial levels can be seen on the northeastern side of the Lake Titikaka (Marshall et al. 2020; Vavrdová et al. 1993). In the Chaguaya section, Marshall et al. (2020) used the last appearances of R. lepidophyta and Umbellasphaeridium saharicum to place the DCB 12 m above a 60–140-m-thick interval of glaciogenic sediments (coarse-grained sandstones, striated/grooved hardgrounds and diamictites). Within the glaciogenic sediments, they found a single specimen of V. nitidus, which may indicate the LN Zone. Older data from Vavrdová et al. (1993) and Díaz-Martínez et al. (1999) for the Villa Molina section show the presence of the LN Zone few metres (sample MM 9a) above the level interpreted as glaciogenic sediments by Marshall et al. (2020).

DCB sections in the vicinity of Lake Titikaka, Bolivia. The sections are famous for the record of glacial deposits, which are all Famennian in age (Marshall et al. 2020). Figure slightly modified from Lakin et al. (2016). Biostratigraphic data according to Vavrdová et al. (1993), Díaz-Martínez and Isaacson (1994), Díaz-Martínez et al. (1999), Lakin et al. (2016)
The most controversial Upper Devonian glacial records in Bolivia are from the Bermejo West section in the Sub-Andean Zone (see summary in Lakin et al. 2016). The section starts with black shales attributed to the late Famennian (?VCo Zone). This succession is topped by 18 m of diamictites and sandstone lenses of the Itacua Formation. The data of Wicander et al. (2011) showed that the base of the Itacua Formation in this section is within the LL Zone, and the top in the LN Zone. Hence, they concluded that the latest Devonian glaciation was a 1-3-My-long multiphase event. This interpretation was challenged by Streel et al. (2013) who proposed an alternative interpretation of the ages and spore assembalges, resulting in a shorter and simplier glacial event.
Isaacson et al. (1995) report a very unusual record of the DCB in 2 boreholes from the Bolivian part of the Madre de Dios Basin (Subandean Zone). In these boreholes, diamictites regrouped in the Toregua Formation are thought to be of Late Devonian and early Carboniferous age. Vavrdová et al. (1996) indicated the presence of the biozones LL, LN and VI in the Toregua Formation, shortly above the (?) lower Famennian (TG Zone). Lakin et al. (2016) indicate that the lower formation boundary is close to the base of the late Famennian VH/Hys Zone. Therefore, they attributed a VI age for the unconformable base of the overlying Yaurichambi Formation. Melo (2005) studied sections in the western part of the Subandean Zone. There, neither the Toregua Formation nor the undifferentitated Retama Group provided in situ latest Famennian and early Tournaisian palynofloras. Miospores of these ages are found as reworked elements in younger strata.
In SE Peru, the Ccatcca Formation shares many lithological similarities with the Bolivian Cumaná Formation and is considered to be age-equivalent (Díaz-Martínez 2004; Isaacson et al. 2008). If this correlation is correct, the DCB may be found in the lower part of the Ambo Group just above the Ccatcca Formation.
Chile
In northern Chile, Upper Devonian to lower Carboniferous siliciclastic rocks are found in two major facies belts, which Bahlburg and Breitkreuz (1993) termed western and eastern basins. The eastern basin (Sierra Almeida) contains a 2700-m-thick succession of shallow-marine platform deposits ranging from Middle Devonian to Visean (Zorritas Formation) (e.g. Boucot et al. 2008; Isaacson et al. 1985; Niemeyer et al. 1997).
A first hint for the position of the DCB in the Middle Member of the Zorritas Formation was the record of Wocklumeria (Breitkreuz 1986). Rubinstein et al. (1996) recognised the VI Zone (oldest Carboniferous miospore biozone), but the study was based on a single sample. According to a detailed palynological study, palynological assemblages were assigned to the latest Famennian (Strunian)?, Tournaisian and Tournaisian-Visean (Rubinstein et al. 2017). The position of the DCB falls into a 15-m-thick mudstone interval between their samples 7051 and 7050. However, characteristic for the assemblages is the high number of reworked spores, which make precise attributions to palynozones impossible; e.g. the marker of the latest Devonian R. lepidophyta is only found in the middle of the youngest assemblages.
In the western basin (Costal Cordillera), several kilometres-thick turbiditic facies developed (e.g. Hervé et al. 1981). Biostratigraphic data are rare, and the different formations are attributed to the Devonian and Carboniferous based on plants, brachiopods and conodonts (see data and references in Hervé et al. 2007).
Argentina
The DCB in Argentina is in general characterised by an unconformity, showing Devonian folded sediments overlain by less disturbed Carboniferous beds (López Gamundi and Rossello 1993). However, a biostratigraphic control of the succession is scarce in many areas, and data are available only in the central and northern Precordillera.
In the Rio Blanco Basin (San Juan province), a hiatus, possibly spanning from Frasnian to the early Tournaisian, is documented (Amenábar et al. 2009). The green-brown shales, with limestone nodules and lenses and subordinate sandstones of the Chigua Formation are followed by thick-bedded paraconglomerates with a few intercalated fine-grained sandstones and shales of the Malimán Formation. The units are separated by an angular unconformity. Palynomorphs indicate a Givetian age for the Chigua Formation, and a late Tournaisian-Visean for the lower part of the Malimán Formation (Amenábar et al. 2009). A similar situation is documented slightly to the south in the Uspallata-Callingasta Basin, where the green greyish sandstones and pelites of the Codo Formation, dated to the Frasnian, are followed by the sandstones, conglomerates and pelites of the El Ratón Formation of ?late Tournaisian-Visean age (Amenábar and di Pasquo 2008; Milana and Di Pasquo 2019).
A possibly different context occurs in the Sierra de Las Minitas (northern Precordillera, La Rioja province). There, a wide range of sedimentary siliciclastic lithotypes is assigned to a poorly defined Jagüel Formation (González and Bossi 1986), which age encompasses most of the Devonian and Mississippian. These sediments are strongly folded and faulted and include a number of glacial diamictites (Prestianni et al. 2015, and referenced therein). In the lower part of the unit, Lower Devonian trilobites are documented (e.g. Holloway and Rustán 2012), but no stratigraphic data are available above until the diamictite levels, interpreted as Late Devonian Glacial Event (see discussion in Prestianni et al. 2015). In a section located in the southern part of the Sierra de Las Minitas, Prestianni et al. (2015) documented the miospore Waltzispora lanzonii from horizons located about 35 m above the diamictites in a palynological association mainly composed of Devonian taxa. However, W. lanzonii is documented only from the Tournaisian of north-northwestern Gondwana, and therefore the Devonian taxa should be considered reworked by glacial erosion (Prestianni et al. 2015).
Africa
South Africa
The correlation of strata of the large intracontinental Cape-Karoo Basin to global stratigraphic units is often complex. The Witteberg Group consisting of marginal marine deposits of mudstones, siltstones and sandstones encompasses the Givetian to Tournaisian interval. Nowadays (Gess 2016), the DCB can be precisely placed in this succession based on a rich and diversified fish fauna. The stratigraphic marker is well documented by the extinctions of placoderms and of most of acanthodians and sarcopterygians at the top of the sandstone-dominated Witpoort Formation. The succeeding mudstone- and siltstone-dominated Kweekvlei Formation is characterised by the radiation of the actinopterygians (Gess 2016). This faunal signal confirms the earlier idea of a regressive sandy Famennian and transgressive muddy to silty Tournaisian strata (Cooper 1986) and the position of the DCB close or at the contact between the Witpoort and Kweekvlei formations (Streel and Theron 1999; Theron 1993). An interesting element for the correlation with South America is the glacial signal in the Peerdepoort Member (upper part of the Witpoort Formation) (Almond et al. 2002).
Ghana
In equatorial Africa, Ghana is the only country with a precise record of the DCB. The palynological records place the DCB within the Takoradi Shale Formation at Essipon Beach near Sekondi (Atta-Peters 1996; Atta-Peters and Anan-Yorke 2003). The formation is part of the Ordovician to Lower Cretaceous Sekondi Group.
The c. 200-m-thick Takoradi Shale Formation is composed of black or dark grey compact mudstone and shales with intercalations of sandstones. Atta-Peters and Anan-Yorke (2003) illustrate an about 20-m-thick section exposing the uppermost Devonian and lowermost Carboniferous. The palynological content of 23 samples have been correlated to Western European standard palynozones by Atta-Peters (1996). Samples 1–6 are correlated to the LE Zone, sample 7 to the LN Zone and samples 8 to 23 to the VI Zone. Sample 7 was taken from the top of a 40-cm-thick sandstone bed, and sample 6 1 m up-section in a 2.5-m-thick unit of shales and mudstones. Atta-Peters and Anan-Yorke (2003) added more samples to the critical interval around the DCB, and they could place the boundary about 45 cm above sample 7. Besides the very precise palynology-based position for the DCB in the Takoradi Shale Formation, it is interesting to note that as in South Africa the DCB is at or near the lithological changes from coarse to fine sediments.
Northern Africa
In northern Africa, the South Atlas Fault and its eastward prolongation separate the mobile Variscan belt in the north from a tectonically less deformed domain in the south, the so-called Northern African or Sahara Platform, which formed on the cratonic basement of Gondwana. Uppermost Devonian and basal Carboniferous strata crop out in both domains, but most information are from the remnants of the epicontinental sea, often called basins, which covered the Sahara Platform in southern Morocco, Algeria and Libya. North of the fault, almost all Variscan structural units contain strata attributed to the Upper Devonian and/or lower Carboniferous, but a lot of those units are affected by early Variscan deformation, called Eovariscan (370–360 Ma after Hoepffner et al. 2005), in the Late Devonian and early Tournaisian. Overall, unfavourable facies (e.g. unfossiliferous shales), reworked and re-sedimented material, important stratigraphic gaps and synsedimentary tectonics hamper the precise determination of the DCB. Continuous stratigraphic sections are often missing and data remain often scattered over the entire succession. Hence, the DCB is often approximately placed by dates coming from much further down and/or up in these successions. The currently most-studied outcrops are situated in the Western Moroccan Meseta.
Morocco
Morocco is the African country, where the most modern biostratigraphic data for the DCB interval are available. Cephalopod faunas of the Upper Devonian successions of the eastern Anti-Atlas (e.g. Tafilalt, Maïder, Dra Valley) have been intensively investigated since the pioneer studies of Termier and Termier (1950) and Petter (1959, 1960). With additional data for conodonts, a rather robust stratigraphic framework for the sections straddling the DCB has been established (e.g. Becker et al. 2013; Kaiser et al. 2007, 2011; Korn et al. 2004) in the different facies realms defined in the Tafilalt and Maïder (Wendt 1988; Wendt et al. 1984). In this region, the black shales or their weathered equivalents at the base of the Fezzou and Aoufital formations (e.g. Becker et al. 2013; Kaiser et al. 2011, 2013; Korn 1999) are considered to be equivalent to the Rhenish Hangenberg Black Shale.
In the northern Maïder, the fossiliferous upper Devonian-lower Carboniferous succession around the Lalla Mimouna hill has gained a lot of interest in recent years (Becker et al. 2012, 2013; Fröhlich 2004; Hartenfels and Becker 2016; Korn et al. 2004). Works concentrated on two sections called Lalla Mimouna A and B by Korn et al. (2004) and respectively Lalla Mimouna South and North by Becker et al. (2012, 2013). The sections are important since they are the only ones with abundant conodont assemblages across the DCB interval in this area.
The research interest in this area was triggered by the findings of Korn et al. (2004), on the northern slope of Lalla Mimouna, of an ammonoid-bearing horizon at the top of a 1.1-m-thick unit of sandy and marly limestones. These contain Acutimitoceras hilarum and Postclymenia evoluta, and conodonts indicative for the Upper praesulcata Conodont Zone (= Pr. kockeli Zone). Hence, in this section, the Acutimitoceras-Postclymenia Ammonoid Assemblage has a latest Devonian age.
Later, Becker et al. (2013) coined for the latest Devonian and earliest Carboniferous strata the name Lalla Mimouna Formation. This formation is 9-m-thick and consists of a silty shales/marls package sandwiched between two crinoidal limestone units (Fig. 4). The lower member (1.4-m-thick) is composed of coarse-grained crinoidal limestone (beds 1–4b), a poorly exposed silty/marly interval (bed 5), topped by a sandy limestone bed (bed 6). This last bed contains the conodonts Palmatolepis gracilis gracilis, Bispathodus stabilis vulgaris, Bi. aculeatus aculeatus and unnamed siphonodellids. According to Becker et al. (2013), the entire lower member belongs to the Si. praesulcata Zone although Si. praesulcata is absent from that member. This lower member is considered to represent the pre-Hangenberg sedimentation. The middle member (bed 7, 7.5-m-thick) consists of silty shales/marls with several intercalations of thin-bedded, calcareous siltstones, which have been individualized as sub-beds. One of those, Bed 7b, contains a rich brachiopod fauna typical for the latest Famennian. Becker et al. (2013) correlate the base of this unit with the onset of the Hangenberg crisis, but could not find the anoxic level as equivalents of the Hangenberg Black Shale.

DCB in Lailla Mimouna North section, Morocco. The DCB is placed at the base of bed 12 (12a) by Becker et al. (2013), although Si. sulcata appeared slightly higher. The slightly modified log from Becker et al. (2013) shows the main lithological and stratigraphic information available for this section. The lateral section inserted shows the highly variable thicknesses of the beds around the DCB. This is most likely a depositional signal, and it certainly influences fossil content and hence the boundary position
The upper member (0.57 thick) is three-folded. The lower sub-unit (beds 8–10) comprises rather sandy, crinoidal pack-rudstones with intercalated siltstone layers. It contains a rich conodont fauna including Protognathodus meischneri, Pr. collinsoni, Polygnathus cf. purus purus and Pseudopolygnathus aff. primus. The middle sub-unit (bed 11) consists of deeply weathered calcareous siltstones, but laterally sandy or silty crinoidal pack-rudstones are intercalated in this subunit. Relatively pure crinoidal pack-rudstones separated by discrete siltstone layers form the upper sub-unit (beds 12–13). From thin limestones (beds 12b–c), Becker et al. (2013) mentioned a single specimen that falls into the variability of Si. sulcata (s.l.) sensu Flajs and Feist (1988). It resembles specimens regarded as transitional morphotypes between Si. praesulcata and Si. sulcata or as Si. sulcata (Ziegler and Sandberg 1996). Bed 12e contains a single specimen of Si. sulcata (s.l.) falling into group 5 of Kaiser and Corradini (2011). It suggests that bed 12e belongs to the sulcata Zone and hence it is undoubtfully of Carboniferous age. However, Becker et al. (2013) place the base of the Si. sulcata Zone, and in consequence the DCB at the base of bed 12. Bed 12g is attributed to the Upper duplicata Zone (= Si. jii Zone), which clearly hints to a, important stratigraphic gaps. A marked change in the conodont biofacies is observed in bed 13a, where siphonodelloids disappear and protognathodids (Pr. meischneri, Pr. collinsoni, Pr. kockeli) re-appear. The Lalla Mimuona Formation is topped by deeply weathered, greenish silty shales of the Fezzou Formation.
The major problem of the Lailla Mimouna sections are the important lateral variable thickness of the beds, several beds wedging out laterally. In fact, some of the bed contacts may correspond to hummocky cross-stratifications, pointing to very dynamic depositional systems for which the reworked conodonts mentioned by Becker et al. (2013) are a further evidence. Overall, the sections at Lalla Mimouna share many characters with the succession exposed in the GSSP at La Serre (Feist et al. 2021, this issue), including its shortcomings.
In the southeastern Amessoui Syncline, two sections—El Atrous and Jebel Ouaoufilal—expose a several 100-m-thick DCB interval within the Aoufital Formation. The most recent detailed data are available for the El Atrous section (Kaiser et al. 2013). The missing Wocklumeria Zone (UD VI-D) indicates that the top of the Erfoud Formation corresponds to an unconformity. Above this unconformity, a 2.5-m-thick unit of reddish-white unfossiliferous shales (bed lower K3), which can contain sulphates from pyrite weathering, is considered to be the Hangenberg Black Shale equivalent. Up-section, the lower part of the Aoufital Formation (beds upper K3-K28) consists of a more than 260-m-thick succession of alternating unfossiliferous, greenish shales and thin-bedded siltstones and fine-grained sandstones. At the Jebel Ouoaoufilal, this stratigraphic level contained a thin sideritic layer with the oldest Acutimitoceras (Stockumites), which indicates the lower UD VI-F Ammonoid Zone. The sandstones of bed K 28 are interpreted as laterally wedging mass-flow deposits, which are an equivalent of the Hangenberg Sandstone. The DCB is placed at the base of bed K 31 into a coarsening-upward sequence (beds K29-36; 60-m-thick) from shales and intercalated thin sandstone beds to proximal sandstone deposits (K33-36). The position of the DCB is based by default on the presence of the rhynchonellid brachiopod Hemiplethorhynchus (?) sp. cf. allani and the absence of ?Centrorhynchus, which possibly indicates a Tournaisian age. The top of the Aoufital Formation contains about 30 m of olive-green shales, topped by black shales of the basal Oued Znaigui Formation correlated with the Lower Alum Shale Event (upper part of the lower Tournaisian or ‘middle’ Tournaisian in the older literature).
North of the South Atlas Fault, the allochthonous nappes at Mrirt at the eastern margin of the Western Moroccan Meseta are an example for several problems to locate the DCB (Becker et al. 2020; Walliser et al. 2000). Although several sections contain stratigraphic markers typical for the DCB interval (Becker et al. 2020), the precise position of the DCB remains unknown. More detailed data are available from the western part of the Western Moroccan Meseta. Kaiser et al. (2007) studied in the Oulmes region the Ain Jemaa sections. In the Ain Jemaa 1 section, basal clymeniid-rich nodular limestones (beds 1–2, middle member of the Bou Gzem Formation) contained conodonts indicating the Bi. ac. aculeatus Zone. Above these limestones follows the upper member of the Bou Gzem Formation composed in ascending order of a 3-m-thick unit of unfossiliferous, silty shales (bed 2b), a 7-m-thick unit of unfossiliferous pyritic black shales (bed 2c) and 15 m of shales with haematite-rich nodules and concretions (bed 2d). Up-section follow about 20 m of grey quartzitic sandstones (bed 2e, Ta’arraft Formation). Despite the absence of biostratigraphic evidence above the nodular limestones, bed 2c is interpreted as equivalent of the Hangenberg Black Shale, bed 2d as Hangenberg Shale equivalents and bed 2e as Hangenberg Sandstone equivalent. This interpretation was based on the application of a model for a supposed global sequence stratigraphic framework for the DCB interval (Becker 1996; Bless et al. 1993). The validity of this assumption cannot be verified without re-consideration of all data.
Algeria
In Algeria, uppermost Devonian and lowermost Carboniferous strata crop out in several basins south of the South Atlas Fault (Conrad et al. 1986). A general problem is that many studies rely on the data calibrated to the then ground-breaking cephalopod stratigraphic schemes of Petter (1959, 1960) and Pareyn (1961), and not to modern stratigraphic zonations as, e.g. Korn et al. (2004, 2007).
The successions of the Béchar Basin sensu lato are often considered to be a reference. Those sections are fully marine and carbonate-dominated in its northern part (Djebel Antar and Horreit, Ben Zireg area), whereas in the southern part, siliciclastic deposits define the boundary interval in the Saoura Valley and in the transition to the Ougarta (Marhouma).
In the NE of Djebel Antar (Soltane et Betoum Anticline), a condensed Upper Devonian-Tournaisian succession is only complete in the East. A 2–3-m-thick unit of shales is attributed to the latest Devonian (Strunian) based on the presence of Bi. costatus (Weyant 1976). This is followed by a 30-m-thick unit divided into a lower shaly sub-unit topped by a limestone sub-unit dated as late Tournaisian (Si. anchoralis Zone). The shaly unit contains at its base undeterminable goniatites and the conodonts Pr. meischneri and Pr. kuehni. Lemosquet et al. (1980) correlate this level with the Stockum level of the Rhenish succession. In the Ben Zirg area, the Viséan wildflysch contains blocks from which Alberti (1972) described latest Devonian trilobites (Phacops (Ph.) accipitrinus) and conodonts, which may occur from the Bi. costatus to the Si. quadruplicata zones. Weyant (1985) mentioned for the north of the Bechar Basin sensu lato ‘a Protognathodus fauna at the boundary between the upper Famennian and Tournaisian …’, which most likely corresponds to the Pr. kockeli Zone placed at the base of the Carboniferous (Lemosquet et al. 1980), but also includes the basal Carboniferous conodonts mentioned by Alberti (1972).
At the southern end of the Bechar Basin sensu lato, a facies change from the sandstone-dominated Ouarourout Formation to the mudstone-dominated Olad-Bou Hadid Formation materialises the boundary interval. The DCB is placed at the base of the Olad-Bou-Hadid Formation after Conrad et al. (1970). In a section north of Ouarourout, they found a thin limestone bed near the formation base containing the conodonts Po. inornatus, Po. communis, Ps. dentilineatus, Siphonodella sp. and ‘Ozarkodina’ sp. The mudstones below this limestone bed yielded a cephalopod assemblage containing small specimens of Gattendorfia, Imitoceras and an undetermined clymenid. In consequence, Weyant (1985) placed the sulcata Zone at the base of the Olad-Bou Hadid Formation. Korn et al. (2007) correlated the Ouarourout ammonoid fauna with their Gattendorfia-Eocanites Assemblage. They indicate that this assemblage does not represent the earliest Carboniferous, but a higher stratigraphic level equivalent to the grey shales above the Hangenberg Limestone of the Rhenish succession. Hence, the actual DCB in the Algerian sections has to be positioned lower probably near the top of the Ouarourout Formation.
An important palynostratigraphic record for the DCB interval is known from the so-called argillaceous series of the basal Ioucha 1 Formation in borehole ut2 in the Western Grand Erg at the eastern side of the Bechar Basin sensu lato. In their update of the classical work of Lanzonih and Magloire (1969), Coquel and Abdesselam-Rouighi (2000) identified the oldest association (Biozone L7 of Lanzonih and Magloire, 1969) in −590 to −687.50-m depth. This rich assemblage contains R. lepidophyta and hence is of latest Famennian age. Coquel and Abdesselam-Rouighi (2000) pointed to the absence of Lophozonotriletes cristifer and the rareness of Rugospora flexuosa compared to contemporaneous assemblages in the Sahara. The second assemblage (Biozone M2 of Lanzonih and Magloire 1969) in −540 to −590-m depth is less diversified. The new data show the presence of Vallatisporites vallatus, which combined to the absence/rareness of Aratrisporites saharaeasis indicates the lower to middle Tournaisian. Coquel and Abdesselam-Rouighi (2000) used the sudden disappearance of R. lepidophyta in ut2 as an argument to support a stratigraphic gap at the top of the Devonian (latest Strunian), and possibly the basal Carboniferous, but the later assumption lacks palynological arguments.
In the Timimoun region east of the Ougarta, the DCB interval is found in the argillaceous Kahla Sandstones, which are upper Devonian to lower Tournaisian (Conrad et al. 1986; Legrand-Blain 1979). The Kahla Sandstones are divided into the lower and upper Kahla Sandstones. The basal part of the lower Kahla Sandstones contains coquina-like limestone intercalations yielding a late Famennian brachiopod assemblage (Legrand-Blain 1979). This is topped by 180 m of fluvio-deltaic deposits of Strunian (age based on the report of R. lepidophyta by Lanzonih and Magloire (1969)). The upper Khala Sandstones contains two major shaly intervals. The lower interval contains a Gattendorfia-Kahlacanites Assemblage (Ebbighausen et al. 2004; Korn et al. 2007), which is placed near the top of the Gattendorfia-Eocanites Ammonoid Genus Zone (Korn et al. 2004, 2007). In comparison to Central Europe (Ebbighausen et al. 2004), this assemblage points most likely to a stratigraphic level above the Gattendorfia Limestone. The upper shale interval contains a Kazakhstania-Acrocanites Assemblage and a Pericyclus-Progoniatites Assemblage, which indicate a late early Tournaisian to early late Tournaisian age for this shaly unit (Korn et al. 2010).
In other Algerian basins, the situation is comparable to the above-described successions. The uppermost Famennian is dominated by coarser-grained silicilastic deposits, which on a N-S gradient show a trend from marine, deltaic to continental facies. Those deposits are often described as being regressive (see, e.g. Conrad et al. 1970, 1986; Legrand-Blain 1979). The following Carboniferous deposits are fine-grained and argillaceous intercalations become more common or the lithology is more variegated. They often contain a marine fauna of Tournaisian age, which leads to their description as a transgressive phase. Commonly, the base of the Carboniferous is equaled with the lithological change, e.g. lower and upper Khenig Sandstone, north of the Hoggar (Reggan-Ahnet-Mouydir area), but equivalent to the Timimoum region, the first Carboniferous fauna seems to occur stratigraphically a little bit higher than the earliest Carboniferous. Hence, it seems to be likely to assume that the DCB should be placed slightly lower as commonly done.
Libya, Mali, Niger
In Lybia, Devonian and Carboniferous strata have been of high interest for the hydrocarbon exploitation (e.g. Belhaj 2000). In eastern and western Libya, sedimentary rocks of latest Famennian age have been identified by palynomorphs. In the subsurface of eastern Libya (Cirenaica Platform), Streel et al. (1988) attributed an assemblage of R. lepidophyta, Ru. flexuosa, V. pusillites and K. literatus to the LL Zone of the latest Famennian (praesulcata Conodont Zone). In western Libya (Rhadamès and Mourzouk-Djado basins), the palynozone 11 of Massa et al. (1979) seems to cover a similar stratigraphic level. In the traditional view (e.g. Belhaj 2000; Conrad et al. 1986), the latest Famennian rocks are unconformably overlain by the Marar Formation. Its base contains a diverse marine fauna including brachiopods (e.g. Fustella), ammonoids (e.g. Muensteroceras) and conodonts (e.g. Gn. texanus) of late Tournaisian age (Conrad et al. 1986; Mergl and Massa 2000).
However, a completely different scenario is reported by Mergl and Massa (2000). They suggest for a rich brachiopod-bivalve assemblage from the middle Tahara Formation in the Awaynat Wanin Area (Ghadames Basin) a Famennian age. In the upper part of the same formation, sandstones yield a brachiopod association attributed to the Hastarian (early Tournaisian). The lower part of the overlying Marar Formation also contains brachiopods attributed to the Hastarian. Mergl and Massa (2000) dated the so-called ‘brachiopod sandstone marker’ of Seidl and Röhlich (1984) in the upper Ashkidah Formation of the northern Murzuq Baisn (Wadi ash Shati area) as definitely early Tournaisian.
In Niger, Devonian-Carboniferous succession are known from the Iullemedden Basin and Djado Sub-basin. The Djado Sub-Basin shares many similarities with the northward situated Murzuq Basin in Libya, but the upper Devonian strata is poorly studied and may even be largely absent. In the Carboniferous, the transgressive marine deposits of the Marar Formation are well-developed (Mergl and Massa 2000). The views on the upper Devonian and lower Carboniferous succession of the Iullemedden Basin (Tim Mersoi Sub-basin) are contradicting (see Conrad et al. 1986; Legrand-Blain 1985a). However, the DCB has so far not been precisely located and it may even be absent in this basin, which would agree with the strong regressive character of the uppermost Devonian, and the gradual flooding of the margin of the African craton during the Tournaisian. In Mali, the Devonian-Carboniferous transition is highly condensed and incomplete in the Taoudenni Basin (Legrand-Blain 1985b). The basal part of the Bir en Naharat Formation is not dated, but may be Devonian in age. Higher up in the formation, a level with the conodont Ps. primus is succeeded by a ferruginous conglomerate and fine-grained sandstones, both containing upper Tournaisian brachiopods. The rest of the Bir en Naharat Formation has been attributed to the Visean.
Australia
Strata straddling the DCB can be found in western and eastern Australia. One recurrent problem is the absence of precise biostratigraphic data and zonations, which often renders the placement of the boundary difficult or impossible. This problem is further enhanced, especially in the Carboniferous parts of the eastern Australian successions by a marked faunal provincialism (e.g. Jones 1995).
Around the DCB, this handicap can be overcome when palynomorphs are present. In many of the Australian basins, the recognition of the widespread, and almost cosmopolitan, Retispora lepidophyta Assemblage can be used to identify the latest Famennian (e.g. Jones 1995; Kemp et al. 1977).
In Queensland, the DCB can be situated in shallow-water carbonates (oolithic limestones), which are intercalated within thick volcanoclastic and siliciclastic complex (Bryan et al. 2003; Roberts et al. 1993). On example is the Gudman Formation (see Webb 2005 for details), west-southwest of Rockhampton in eastern-central Queensland. It is mostly early Tournaisian in age based on conodonts (Si. sulcata to Si. crenulata zones) (Mory and Crane 1982) and brachiopods (Tulcumbella tenuistriata Zone) (Roberts et al. 1993). However, the lower part of the formation contained conodonts of latest Devonian age. Mory and Crane (1982) reported an undifferentiated latest Famennian Si. praesulcata Zone. The data of Webb (2005), a posterior tip of a Palmatolepis and one element tentatively assigned to (?)Bi. bispathodus from the lower part of the formation, may extend the biostratigraphic range of the formation even deeper into the late Famennian. But up to now, the position of the DCB is not located precisely in the Gudman Formation. By default, it is located below the base of the first reefal unit, which plays an important role for the understanding of the recovery phase after the Hangenberg extinction (Denayer and Webb 2017; Webb 1998; Webb 2005).
Further north, in the hinterland of Townsville, the DCB is situated in several formations defined in different basins/regions (see details in Talent et al. 2002). Those formations are all dominated by mainly non-marine siliciclastic sedimentary rocks, but marine carbonated levels productive for conodonts, though rare and unevenly distributed, are intercalated into the succession. Mawson and Talent (1997) identified major marine pulses in the succession, one ‘early’ in the Si. sulcata Zone. This gives a relatively precise position for the boundary, especially if the underlying strata contains late Devonian conodonts (e.g. Si. praesulcata Zone in the Hardwick Formation of the Burdekin Basin).
In western Australia, in the Canning Basin, Talent et al. (1993) correlated the change from the mixed siliciclastic-carbonate succession of the Gumhole Formation to the overlying sandstones of the Yellow Drum Formation with the major global regressive phase of the Hangenberg Sandstone Event. Hence, these sandstones would (at least partly) be an equivalent to the Hangenberg sandstones of the Rhenish succession. Mory and Hocking (2011) summarised the available biostratigraphic data for the two formations. The best age for the upper part of the Gumhole Formation derives from the already abovementioned Retispora lepidophyta Assemblage. The local Po. communis collinsoni and Icriodus platys zones for shallow-water conodonts are recognised in the Gumpole Formation (Nicoll and Druce 1979), but their precise correlation to the international zonal schemes remains questionable.
Mory and Hocking (2011) place the DCB within the lower part of the Yellow Drum Formation. This position is based on the observations of a nowadays-filled trench next to Linesman Creek. There, the income of Si. sulcata marks the base of the Carboniferous, at a level also marked by a shift in δ13C (Andrew et al. 1994). Since the Gumhole Formation was not exposed in this trench, the precise position of the DCB within the Yellow Drum Formation remains unknown.
Asia
Vietnam
The Cat Co section, located in the Cat Ba Island (northwestern Vietnam) and exposing rocks of the Pho Han Formation, has been studied in detail in the recent period with multidisciplinary studies including conodont biostratigraphy, facies analysis, trace elements, isotopes and organic geochemistry (Komatsu et al. 2012, 2014; Paschall et al. 2019; Shizuya et al. 2020). The section (Fig. 5) is very condensed around the D/C boundary and consists of micritic light grey to grey limestones, marls and interbedded thin dark grey limestones and organic carbon-rich, black, friable limestones (Komatsu et al. 2014). The Hangenberg Event is expressed by a c. 1 m (beds 116–118) of laminated black shales alternated with dark grey limestone (Komatsu et al. 2014; Shizuya et al. 2020).

DCB in the Cat Co section, Vietnam. Overview on stratigraphy, lithology, δ13Corg curve and occurrences of selected conodonts important for placing the DCB. The DCB is placed at the base of bed 122. HBS = Hangenberg Black Shale. (modified from Shizuya et al. 2020)
Conodonts are well preserved, but the association lack of some typical elements of the DCB interval, bispathodids and protognathodids is not reported. The base of the Carboniferous is detected by the first occurrence of Si. sulcata in bed 119. Beside conodonts, calcareous algae, foraminifers, brachiopods and crinoids are reported. Geochemical studies (trace element proxies, total organic carbon, framboidal pyrite distributions, etc.) and a significant mercury enrichment suggest dysoxic/anoxic conditions from the latest Famennian through the early Tournaisian with severe anoxia (approaching euxinia) throughout the Hangenberg Event interval (Paschall et al. 2019; Shizuya et al. 2020). The authors interpret the mercury anomaly as the result of large-scale volcanic activity.
Malaysia and Thailand
Devonian and Carboniferous sedimentary successions are known in SE Asia but the precise stratigraphy is still poorly known (Hamada 1967; Oliver et al. 1995). On both sides of the Malaysia-Thailand border near the western coast, the Sanai Limestone Member of the Jentick yielded diagnostic Famennian conodonts (Meor and Lee 2003) whereas the overlying Telegu Jatoh and Wang Kehan silicoclastic member yielded the trilobite Macrobole kedahensis, which indicates an early Tournaisian age (Cocks et al. 2005). From the Kathan Limestone Formation cropping out in Peninsular Malaysia, Metcalfe (2002) described conodonts from the upper Tournaisian Sc. anchoralis Zone, together with reworked late Devonian and early Tournaisian species, and evidenced a disconformity between Eifelian and upper Tournaisian strata. From Western Peninsular Malaysia, Tsegab et al. (2017) described a diverse conodont fauna ranging from the Famennian to the Tournaisian, including abundant polygnatids and siphonodellids, from the Kintab Limestone Formation. However, the DCB markers are not described.
In the Chiang Dao area (northern Thailand), Randon et al. (2006) described conodonts from a thick-bedded chert sequence of the Fang Chert Formation. The authors documented late Frasnian, early Famennian (Pa. gl. prima Zone) and late Tournaisian (Sc. anchoralis Zone) conodonts from the ‘Chiang Dao Cherts’ section, without possibility to better constrain the stratigraphic context. This is a rather common situation in Thailand, where Late Devonian-early Carboniferous ages are attributed to black lydite/bedded chert sequences, based on very poor detailed stratigraphic control (see also discussions in Fontaine et al. 2005, 2009).
NW India
The DCB is documented in the Pin Valley of Spiti (NW India) by Draganits et al. (2002). The authors studied several sections within the Middle Devonian-lower Carboniferous Lipak Formation, mainly represented by mixed siliciclastic and calcareous sediments in the lower part, followed by fossiliferous limestones and sandstones and, at place, rare dark mudstones and thinly bedded siltstones. The authors provided a reconstruction of the depositional environment in terms of sequence stratigraphy, supported by biostratigraphic data based on a few conodont samples.
C. 75 m above the base of the Lipak Formation, a 58-cm-thick unit of dark grey carbonaceous shales sandwiched between crinoidal grainstone is interpreted as a Hangenberg Shale equivalent. Conodonts from the samples just below and above this level yielded a fauna documented from both the latest Famennian and basal Tournaisian (e.g. Bi. ac. aculeatus, Bi. spinulicostatus, Clydagnathus cavusformis and Ps. primus).
Pakistan and Afghanistan
Late Famennian and Tournaisian conodonts were illustrated by Molloy et al. (1997) from the Khyber region, north-west Pakistan. A few sections were measured in the area, and two of them (sections SM and SB) span the DCB. The sections are mainly constituted by limestone with interbedded shales and siltstones, and look continuous across the boundary, but a detailed log or a description is currently not available.
In the SM section, the boundary can be constrained in a few metres’ interval around a ‘shale and siltstone’ unit between the last occurrence of Bi. costatus and the first occurrence of Po. rostratus. In the SB section, the same interval looks thicker (more than 10 m) and includes several shaly beds. However, it should be noted that the conodont abundance is scarce in that part of the sections, protognathodids are not reported and only one specimen of Si. cooperi is documented from the upper part of the SM section.
In its synopsis of the stratigraphy of the Carboniferous stratigraphy for the western Karakorum, Gaetani et al. (2004) defined for the Karambar Unit the Margach Formation. Although the fossil content of the formation is very poor and patchy, and stratigraphic diagnostic fossils apparently lack, the stratigraphic range has been indicated as mid Famennian to ?early middle Tournaisian.
Sections spanning the DCB in Afghanistan are mentioned by Mistiaen et al. (2015). However, detailed data are not available.
Iraq and Saudi Arabia
In the northern part of Iraq, the shallow-water uppermost Famennian-Tournaisian rocks belong to the Ora Formation. A rich spore association from the Kaista section was illustrated by Naquishbandi et al. (2010), but precise data on the DCB are not available, nor a detailed description of the section and on other fossil groups. The Ora Formation is also known in the subsurface of western and southern-eastern Iraq (Al-Juboury and Al-Hadidy 2008; Aqrawi 1998), but again, detailed data are not available.
In Saudi Arabia, the uppermost Devonian and lowermost Carboniferous strata are known from the subsurface in the NE and E of the country (Bassis et al. 2016). Conformable contacts between the two systems seem to be rare, and in many regions the strata are eroded and truncated by the pre-Khuff or pre-Unayzah unconformities. In this context, the presence of reworked latest Devonian miospores in the younger Tournaisian and Visean strata (e.g. Clayton 1995; Clayton et al. 2000) clearly indicates the erosion of the strata of the DCB interval.
The DCB lies within the upper part of the Jubah Formation. This formation consists of continental and fluvial depositional environments. Al-Hajri et al. (1999) and Clayton et al. (2000) provided slightly different lists of the miospore taxa from the uppermost Jubah Formation in the Harmaliyah field (eastern Saudi Arabia). However, both report the abundance of R. lepidophyta, which results in the identification of the regional Saudi Aramco palynozone D0 (Al-Hajri et al. 1999). Clayton et al. (2000) concluded that the upper part of this Retispora lepidophyta Assemblage corresponds to the LE and possible LN zones of Western Europe. The lower part could be older than those two zones. In the Abu Safah-29 borehole, Clayton et al. (2000) identified a Indotriradites explanatus Assemblage (also called Saudi Aramco palynozone C6, Hooker et al. 2011). They attributed a Tournaisian (‘probably early Tournaisian’, op. cit.), and indicate the similarities in the floral composition to Palynozone 12 of Grignani et al. (1992) in Libya.
Hence, the precise location of the DCB has so far not been identified, and Clayton et al. (2000) speculated about erosion or non-depositional as possible causes in their studied boreholes.
Turkey
Upper Devonian and lower Carboniferous rocks are widely exposed in various regions of Turkey, but the DCB is rarely precisely placed.
In the Istanbul-Zonguldak area (NW Turkey), the DCB is exposed in the shallow-water facies in the eastern part and deeper-water flysch facies in the western part (Okay et al. 2006). In the western part, conodont data from various localities (Çapkinoğlu 2000, 2005) suggest that the boundary should be placed in the upper part of the Ayineburnu Formation, but precise studies are missing. However, a lenticular massive, poorly sorted sandstone-greywacke body with intraformational intraclasts (Küçükyalı Member) is locally equivalent to the upper part of the Ayineburnu Formation. This unit possibly represents an equivalent of the Hangenberg Sandstone event, but further evidence is needed.
In the eastern part, in the Topluca section (Bartın area), where the upper Strunian succession includes stromatoporoid biostromes, Denayer (2016) placed the DCB in a 1-m-thick poorly exposed interval between bed 33 that yielded the last Quasiendothyra kobeitusana (DFZ7 Foraminifera Zone) and bed 35 that yielded a depauperate fauna of unilocular foraminifers (MFZ1).
In the central and eastern parts of Turkey, Upper Devonian and lower Carboniferous successions are exposed in few localities of the Taurides. In the Geyık Dağı tectonostratigraphic unit, the DCB was investigated in detail in the Natlaş section. Here, the boundary lies in the Natlaş Member of the Ziyarettepe Formation. It starts with a 20-m-thick unit of yellowish sandy limestone with brachiopods, followed by an about 55-m-thick unit of dark grey, locally bituminous argillaceous limestone passing upwards to shales. Plodowski and Salanci (1990) placed the DCB about 30 m below the top of the Natlaş Member on the basis of the last occurrence of trilobites of the genus Phacops, whereas the Tournaisian genera Cummungsella and Piltonia are reported less than 10 m above the boundary. Typical Strunian brachiopods occur in the same beds than the phacopid trilobites, while only Aulacella and Ovatia species occur in the upper part of the Natlaş Member, above the DCB (Plodowski and Salanci 1990). The overlying shales yielded the Tournaisian brachiopods Rugosochonetes sp. and Schizophoria sp., as well as the spores Umbonatisporites disctinctus and Raistrickia clavata (among others) that indicate the upper lower Tournaisian PC Palynozone. Recently, Atakul-Özdemir (2017) described the conodont fauna of the member in its type section, precisely documenting a continuous sequence from the Bi. ultimus to the Si. bransoni zones, and placed the DCB at the first occurrence of Si. sulcata in sample SN18, 25 m above the base of the Ziyarettepe Formation. The foraminifers were preliminary reported by Altiner et al. (2012) but no diagnostic taxon was identified near the DCB, with the notable exception of quasiendothyrids in the upper part of the Natlaş Mbr and tubular unilocular foraminifers 3 m above the boundary, based on conodonts.
In the Silifke area (Antalya tectonostratigraphic unit), the Korucuk Formation is the equivalent of the Ziyarettepe Formation. It starts with 15 m of greenish-greyish siltstone with plant fragments yielding the Strunian guide miospores R. lepidophyta and V. pussilites passing upwards to sandstone-shale-sandy limestone alternations rich in trilobites (Demirtaşlı et al. 1978). The upper part of the Korucuk Formation is a 22-m-thick unit made of dark grey platy limestones with dark shaly intercalations yielding Tournaisian brachiopods and foraminifers (Earlandia spp. and Septabrunsiina spp.) indicating an early Tournaisian age. The brachiopods are abundant in the uppermost Devonian part of the formation, where Plodowski and Salanci (1990) reported the same taxa as in the Natlaş Member, but without clear evidence of Tournaisian material.
In the Hakkari province (SE Turkey), close to the border with Iran and Iraq, the Devonian-Carboniferous succession is exposed in the Zap river valley. The Köprülü Formation starts with a c. 30-m-thick unit of dark sandy limestones interbedded with shales that include two horizons yielding late Devonian fish remains (Janvier et al. 1984). Above the last fish horizon, a 200-m-thick shale interval with small intercalated limestone beds yielded a diverse ostracod fauna indicating the change from restricted marine to intertidal or brackish conditions at the base of the formation to more open-marine conditions in the middle part. Janvier et al. (1984) placed the DCB about 80 m above the base of the Köprülü Formation based on a change in ostracod assemblages. A shaly horizon 50 m above this boundary yielded spores indicating the lower Tournaisian PC Palynozone (Higgs et al. 2002).
Azerbaijan and Armenia
The Geran-Kalasi section (Grechichnikova and Levitskii, 2011) is one of the most complete sections of the Upper Devonian–lower Carboniferous in Azerbaijan. It is located in the Nakhichevan Autonomous Region (west Azerbaijan) and mainly represent a marine continuous sequence with rich fossil associations: brachiopods, foraminifera, bryozoans, tabulate and rugose corals, brachiopods, ostracodes, crinoids, conodonts, algae and ammonoids are present. The section was described in detail by Grechichnikova and Levitskii (2011), who listed the fossil occurrences throughout the section and introduced brachiopod-based biozones. The same levels crop out in the Armenian flank of the Daralagez mountain range, notably in the Arshaki-Arhbyur Gorge (Tolokonnikova 2016).
The DCB interval is represented by about 8 m of thinly bedded sandy limestones with bryozoans, brachiopods, ostracodes and conodonts (Bi. costatus, among others), followed by 5 m of ‘shaly silty, poorly exposed’ sediments and 7 m of sandy, thinly to medium-bedded sandy limestones. Grechichnikova and Levitskii (2011) attributed those beds to the upper part of the latest Famennian Sphenospira julii–Spinocarinifera nigra Brachiopod Zone. The lowermost Tournaisian is identified by the Unispirifer praeulbanensis–Rhytiophora curtirostris Brachiopod Zone, and is represented by 6 m of dark grey shales, with thin interbeds of siltstones and sandy limestones with brachiopods, tabulate and rugose corals, crinoids, conodonts and bryozoans (Abrahamyan et al. 1975; Alekseeva et al. 2018).
Uzbekistan
Preliminary data at the DCB in Uzbekistan are available from the Kule (Erina 2008; Narkiewicz et al. 2017) and Novchomok (Narkiewicz et al. 2017) sections. Both are located in the eastern part of the Kitab Geological Reserve area.
In the Kule section, two lithostratigraphic units are discriminated in the Famennian and Tournaisian: the Akbasay Formation composed of reddish grey cherty limestones and siliciclastics, followed by the Novchomok Formation, represented by ‘grey and light grey, fine-grained and micritic, sometimes brecciated crinoidal limestones’ (Yolkin et al. 2008). No shaly interval is reported near the boundary. A detailed sampling was carried by Erina (2008), who positioned the basal Carboniferous in the lower part of the Novchomok Formation based on the occurrence of questionable element of Si. sulcata. However, the presence in the same bed of an element of Si. lobata (figured as ‘Si. angulata n.sp.’ Erina) questions the position of the boundary. Narkiewicz et al. (2017) did not recover siphonodellids from the boundary beds, but placed the boundary in the uppermost part of the Akbasay Formation on the basis of the entry of Po. p. subplanus. Further studies confirmed the datum, thanks to the recovery of one element of Si. sulcata (Narkiewicz et al. 2021). Only one juvenile element of Protognathodus sp. is documented so far from the Si. jii Zone of the Kule section (Narkiewicz et al. 2021).
In the Novchomok section, the lower part is represented by marly shales and marls interbedded with pelitic and crinoidal limestones of the Yatavluk Formation, overlaid by the limestones of the Novchomok Formation. On the basis of a poorly preserved conodont fauna, including a few protognathodids, the boundary is tentatively placed in the uppermost part of the Yatavluk Formation (Narkiewicz et al. 2017). High mercury concentration in the latest Devonian were documanted by Rakociński et al. (2021).
Tajikistan
Successions spanning the DCB crop out in southern Tien-Shan (western Tajikistan), not far from the border with Uzbekistan (Bardashev et al. 2005, 2006; Bardasheva et al. 2004). Three parallel sections, named Shishkat 3, Shishkat 3a and Shishkat 3b, were measured and sampled in detail by Bardasheva et al. (2004). The DCB occurs within the Surkhobi Formation, a grey to light grey-cream limestone mostly made up by ‘clastic detrital and coarse-detrital rocks, as well as limestone breccia’ (Bardashev et al. 2006). Oolites were reported from the boundary beds, whereas mudstones or shales are not present. Foraminifers and conodonts are abundant; crinoids, brachiopods and ostracods are also present.
Despite the abundance of conodonts, the early siphonodellids are rare in the boundary beds, and only one element of Si. sulcata is reported both from sections Shishkat 3 and Shishkat 3a, and rare protognathodids are reported only from the upper part of the Si. sulcata Zone and above. Also, only an undifferentiated ‘praesulcata Zone’ is discriminated, and the scarce fauna makes further subdivisions difficult, even if the last occurrence of Pa. gr. gonioclymeniae about 11 m below the occurrence of Si. sulcata may indicate the beginning of the Hangenberg Event.
Detailed distribution data of foraminiferas are not provided, but faunal lists are provided for each lithological unit (Bardasheva et al. 2004). Several taxa of genera Cribosphaeroides, Endoglomospiranella, Parathuramminites, Quasiendothyra, Septabrunsiina and Uralinella are listed from a 10-m-thick interval approximately including the upper part of the praesulcata Zone, but their precise occurrence is not available. A less differentiated association, including Parathurammina radiosphaerica, Parathuramminites mikhonoae, Bisphaera malevkensis, B. irregularis and Earlandia minima is listed from a 5-m interval more or less coincident with the Si. Sulcata Zone.
Kazakhstan
Sections exposing the DCB in Western Kazakhstan are treated in a dedicated chapter of the present special issue by Kulagina et al. (2021, this issue). However, a recent re-excavation and description of the Berchogur sections has been published by Nikolaeva et al. (2021). The first occurrence of Si. sulcata in Member 3 allows the localisation of the DCB. However, in their revision, Nikolaeva et al. (2021) identified a regressive event that most probably records the ‘Hangenberg Sandstone event’ concomitantly with a clear separation of Famennian and Tournaisian faunal associations (foraminifers, conodonts, ammonoids, etc.) within the uppermost part of Member 2b of the Dzhangana Formation. The position of the DCB should therefore be somewhat lowered in the succession compared to previously published data.
In southern Kazhakstan, the DCB is exposed in the Bolshoi Karatau Mountains, where upper Frasnian to lower Bashkirian carbonates are exposed along plateform-bassin transects. Detailed biostratigraphic data for calcareous foraminifers (Zhaimina et al. 2017; Zhaimina and Kulagina 2019) allow to place the DCB, e.g. bed 15A in the Karamurum Section. As elsewhere in the former USSR, the base of the Carboniferous corresponds to the base of the Tournayellina pseudobeata Zone. Another interesting stratigraphic signal are breccias in the Balaturlan Member (Shukurganat Formation), which have been correlated with a global regressive trend (Zhaimina et al. 2017), supposedly correlatable to the Hangenberg Sandstone event.
Mongolia
Rocks of Late Devonian and lower Carboniferous age crops out in several areas of Mongolia (Ariunchimeg et al. 2014; Ariunchimeg and Nyamsuren 2001), but only very few data are available. The DCB is not documented so far, as it corresponds to a formational boundary (e.g. Wang et al. 2005; Wang and Minjin 2004) or studies are limited to parts of the successions (e.g. Upper Devonian, Munkhjargal et al. 2021a, b, Suttner et al. 2020).
Russia
This short overview on the DCB in Russia is necessarily incomplete. Information on the boundary are available from various regions in Russia (East-European Platform, Urals, Siberia, North-east Russia, Russian Arctic) and from a large suite of facies ranging from continental and coastal to basinal environments. The contribution of Kulagina et al. (2021, this issue) in this special issue provides a lot of details about DCB boundary sections with a special focus on the foraminifera biostratigraphy.
In the Moscow Basin, in the central part of the East-European Platform, the DCB corresponds to an unconformity between the Khovanshchinian and Malevkian Regional substages, and the correlation to global chronostratigraphy is complicated by abundant shallow-water conodont assemblages (e.g. Alekseev et al. 1996; Gatovsky et al. 2011). Although time is clearly missing, the region is important for latest Famennian vertebrate assemblages (e.g. Lebedev et al. 2018).
The Gumerovian Regional Substage, in which the DCB is situated, fills the stratigraphic gap in the Moscow Basin. It originates from the Zigan section on the western slope of the Urals (e.g. Pazukhin et al. 2009, Kulagina et al. 2021, this issue), and is today widely recognised in the stratigraphic schemes of the East European Platform and Urals. In its type section, the Gummerovian Regional Substage consists of c. 2 m of strata (bed 20 of the local nomenclature); its base corresponds to a sequence boundary. The base of the Carboniferous is placed at the base of bed 20c. However, Gatovsky (2015) reports an almost synchronous first occurrence of the conodont Si. sulcata and the miospore V. pusillites in the upper part of bed 20c. In the sequence stratigraphic interpretation of Gatovsky (2015), bed 20a corresponds to a lowstand systems tract, bed 20b represents a transgressive systems tract and beds 20c to 20f to a highstand systems tract.
In the northern Urals, a lot of new data have been published from the Timan-Pechora region in recent years, especially from the Kamenka River and Vangyr River sections (e.g. Zhuravlev 2020; Zhuravlev and Sobolev 2019). The DCB is placed in the lower part of the Edzhid Formation. The latest Famennian is characterised by lagoonal, clayey carbonates, which contained conodonts as Si. praesulcata, Si. bella, Si. quasinuda, Po. parapetus, Po. communis communis, Hindeodus penescitulus, Ps. graulichi and Ligonodina discreta. The basal Carboniferous is represented by shoal and open marine carbonates containing Si. sulcata, Si. bella, Si. quasinuda, Si. semichatovae and Patrognathus crassus. The foraminifer Tournayellina (Eotournayellina) pseudobeata is found 0.5 m above the boundary (Vevel et al. 2012). The topmost Famennian carbonates are thought to have been deposited under regressive conditions, whereas the basal Carboniferous is transgressive. Tentative correlations to the Rhenish sequence stratigraphic model are discussed, especially to the relative sea-level curve. Vevel et al. (2012) indicated that the maximum regression occurs 0.3 m below the biostratigraphic DCB in the Kamenka River section.
In the northern Urals, important for the ongoing discussions for the revision of the DCB are records of Pr. kockeli in the Izyael River and Kozhim River sections (Nemirovskaya et al. 1993; Plotitsyn 2016).
The Omolon Massif in Far East Russia is often overlooked when discussing the DCB. The Upper Devonian-lower Carboniferous succession of the area had gathered a lot of attention in the 1980s (e.g. Bogush et al. 1987; Conil et al. 1982; Gagiev 1979; Shilo et al. 1984; Simakov et al. 1983; Swennen et al. 1986). Interestingly, the position of the DCB remains disputed. Depending on the interpretation of the fossil data and their correlation to other regions, the DCB of Gagiev (1979) is correctly placed after Kulagina et al. (2021, this issue) but has to be considerably moved up-section after Poty (1999) based on the distribution of both macro- and microfossils. This problem is important for our understanding of the DCB, the associated biological crises and recovery, and the Omolon Massif could play a central role in it. If the DCB remains in the 1980s position, the Omolon Massif can be considered a Carboniferous refuge for Devonian survivors of the biocrises. In this case, the Omolon Massif would be a starting point of the Carboniferous repopulation. However, this also means that the stratigraphic ranges for some fossil groups have to be extended at least locally, and that traditional biostratigraphic signals will fail in those refuges. If the proposed boundary of Poty (1999) is correct, then the faunal extinctions in the Omolon Massif follow the global scheme, but some organisms would have a (significant) earlier first occurrence than elsewhere if the original taxonomical attribution is correct. Doubts can be raised concerning the original identification of the boundary solely based on a fragment of siphonodellid. Hence, these ‘Carboniferous’ newcomers would survive the end-Devonian extinction and provide the stock for the Carboniferous repopulation and diversification.
In Arctic Russia, Upper Devonian and lower Carboniferous rocks are exposed north of the Lena river. They belong to the Stolb, the Ebelakh and Bastakh formations (Mezhvilk 1958). The DCB should occur within the Ebelakh Formation, constituted by fine- to medium-grained feldspar-quartz sandstone with carbonate cement and small amount of dolomite and felsic volcanic matrix (Saraev et al. 2013). However, it is not possible precisely placing the boundary, as fossils are very scarce (Mezhvilk 1958). From the Lena River estuary area, Ivanovski (1967) noted that the oldest corals found in the Carboniferous strata are Caninia cornucopiae and Uralinia multiplex, which indicate probably an upper Tournaisian age, whereas older macrofauna seems to be lacking. Conodonts collected in the Stolb Formation (Izokh 2017) indicate an age not younger than the Pa. rhomboidea Zone, while the older assemblage from the Bastakh Formations is referred to the isosticha-Upper crenulata Zone (Izokh 2017; Izokh and Yazikov 2017).
The situation in Novaya Zemlya is also poorly known at a detailed scale. In the southern part of Novaya Zemlya, the uppermost Devonian-Tournaisian Lazarevskaya Formation includes neritic carbonate facies. The Famennian part of the Lazarevskaya Formation is composed of bioclastic grainstone and packstone and stromatoporoid boundstone (Schecoldin et al. 1994) that yield a rich and diverse assemblage of benthic organisms, including rugose corals (Gorsky 1938), brachiopods and foraminifers (Rzhonsnitskaya 1967). Shallow-marine carbonates extend at least up to the Viséan, with some intercalations of shale and bedded chert in the Tournaisian (Schecoldin et al. 1994). On the western coast of the southern area, Schecoldin et al. (1994) report in the Lazarevskaya Formation the occurrence of black shale and radiolarite that yielded lower Tournaisian conodonts allowing to place the DCB. However, only the Tournaisian species Si. lobata, Si. crenulata and Si. obsoleta were reported. In the Kara area (central-eastern part of the archipelago), the succession across the DCB consists of black bedded cherts, radiolarites, shales and rare carbonate intercalations. The conodont fauna consists of Siphonodella sp., Patrognathus crassus and Clydognathus cf. cavusiformis.
Europe
Ukraine
In the Donets Basin, the DCB is located at the interface of the Devonian Porfiritovaya and Bazalievskaya formations. The uppermost Famennian is characterised by shallow-water carbonate facies, often dolomitized, with an abundant foraminiferal fauna including Quasiendothyra communis, Q. kobeitusana, Q. konensis, Cribrosphaeroides sp., Paracaligelloides florennensis, Tournayella sp., Septatournayella sp., Septaglomospiranella sp. and Septabrunsiina sp., as well as the index conodont Si. praesulcata (Poletaev et al. 1990, 2011). Brachiopods and stromatoporoids are abundant (Berchenko and Poletaev 2007). An unconformity is developed at the base of the overlying Bazalievskaya Formation, but after Poletaev (1981) it might reflect local condition of sedimentation. However, this unconformity should probably be reconsidered under the perspective of the Hangenberg Sandstone event. This formation passes southwardly into the Porfiritiv Formation, which is composed of fine-grained, often dolomitic limestone with the previously reported foraminiferal assemblage, overlain by black shale with brachiopods and a more diverse conodont fauna composed of Antognathus volnovachaensis, Bi. costatus, Bi. aculeatus, Si. praesulcata, Po. parapetus and Pseudopolygnathus spp. (Poletayev et al. 2013), which likely indicate some intermixing of Devonian and Carboniferous species.
The lower Tournaisian is recognised regionally as the Buzinivian Horizon and corresponds to the Bazalievskaya Formation. The basal beds of this unit contain the Tournaisian conodonts Siphonodella aff. sulcata, S. semichatovae and Patrognathus andersoni (Poletayev et al. 2013). The foraminifers, though less diverse than in the underlying strata, are well characterised by Bisphaera malevkensis, B. irregularis and Earlandia minima and form the local ‘bispheric foraminifers zone’ (Poletaev et al. 2011) that possibly corresponds to the ‘unilocular zone’ of Poty et al. (2006).
Belarus
The DCB is documented in two boreholes from the Pripyat Depression, in the southern part of Belarus (Avchimovitch et al. 1988, 1993). The authors studied a rich miospore association and noted a strong floral turnover across the boundary. Unfortunately, detailed lithological and stratigraphic data are not available. However, malformed spores of R. lepidophyta described by Avkhimovich (1993) allow the recognition of the tener event (sensu Prestianni et al. 2016) in this succession, i.e. the recognition of the HBS event equivalent.
Serbia
The DCB is exposed in NW Serbia at the Milivojevica Kamenjar section, a large overturned olistolith spanning from latest Famennian to Serpukhovian (Korn and Sudar 2016; Sudar et al. 2018). Ammonoid and conodont fauna and biostratigraphy were recently documented by Korn and Sudar (2016) and Sudar et al. (2018), respectively.
The DCB interval ‘occurs in a continuous limestone succession, not interrupted by strata indicating the Hangenberg Event’ (Sudar et al. 2018, p. 172). However, a minor lithological change between dark-grey nodular and flaser limestone with high shale content (Famennian) to grey bedded and partly nodular limestone with minor shale content (Tournaisian) is documented. Ammonoids do not allow a precise biostratigraphy of this part of the section, as they are concentrated in a few levels dated to an early ‘Wocklumeria Stufe’ and to a generic ‘middle Tournaisian’ (Korn and Sudar 2016), i.e. the upper lower Tournaisian. Conodont data are more precise and a clear difference in the association is documented in connection with the lithological change: typical latest Famennian species occur in the topmost shaly limestone, whereas Carboniferous taxa as Si. sulcata and Po. purus subplanus are documented just above. A recent attempt to recover palynomorphs from the DCB interval shales failed as the material has a high thermal maturity (Prestianni, pers. com., 2020).
Great Britain
In many parts of Great Britain, the DCB beds are found in the stratigraphic interval often called or associated with the ‘upper’ Old Red Sandstone (ORS). During the Upper Devonian to lower Carboniferous the palaeo-coastline moved from the south of the British Isles towards the north, and marine conditions gradually installed during this time slice. Hence, depending on the geographical position of the studied section, the DCB can be found in neritic marine, marginal marine or continental facies.
Marine conditions dominate the Devonian successions of Devon and Cornwall (SW Britain). There, a classical example for the DCB documented in neritic facies is the Pilton Mudstone Formation of the Culm Basin. In this formation, trilobites and cephalopods are documented ranging from the late Famennian Wocklumeria to the early Tournaisian Gattendorfia ammonoid zones (e.g. Goldring 1955). Further north, where the continental character of the deposits increases, especially palynomorphs have been demonstrated to be of critical stratigraphic importance (e.g. Clayton et al. 1986; McNestry 1988). In this special issue, the contribution of Graham and Sevastopulo (2021, this issue) illustrates this facies realm widely developed in the Upper Devonian and lower Carboniferous in southern Ireland, southern Wales and southern England (e.g. Mendips).
Further north, in the entirely continental facies realm, a recent biostratigraphic study in Scotland (Marshall et al. 2019) demonstrates the importance for the correct placement of the DCB. These new data for the stratigraphic distribution of palynomorphs show that the Upper ORS Kinnesswood Formation is Late Devonian in age, and not early Carboniferous as previously postulated. The DCB approximates with the base of the following Ballagan Formation and its coastal wetland environments (Millward et al. 2018), in which the oldest Tournaisian miospore zone was found at its base at Burnmouth.
These new dates change the perception of the vertebrate evolution in the late Devonian and early Tournaisian. The previously postulated range extension of typical Devonian fish into the Carboniferous can be refuted with the new age of the Kinnesswood Formation. This increases the amplitude of the magnitude of the Hangenberg event for the terrestrial realm, and the DCB becomes a clear barrier in the evolution of terrestrial vertebrates. This study also shows that the Ballagan Formation comprises the entire Tournaisian, and hence the tetrapods and arthropods of this formation fill important parts of the Roemer’s Gap (Marshall et al. 2019).
France
In France, Devonian-Carboniferous successions have been documented in different tecto-sedimentary contexts in various parts of the country, north and south of the future Variscan Chain. The best-known successions in the Montagne Noire and Avesnois are treated in this special issue respectively by Feist et al. (2021, this issue) and Denayer et al. (2021, this issue).
The Boulonnais area in northern France is the equivalent of the Belgian Brabant Parautochton as its western continuation emerging below the post-Palaeozoic cover near the Boulogne-sur-Mer area. The Devonian and Carboniferous successions of the Massif de Ferques are comparable to the one, e.g. in the Tournai drillings and Feluy area in Belgium (Denayer et al., 2021, this issue) but the transition is far less well dated. The Sainte-Godeleine Formation is a greenish to reddish sandy and shaly unit c. 50 m thick displaying shallow-water bioturbated facies almost devoid of fossil (Ager and Wallace 1967). Its age is extrapolated from the lower Famennian acritarch assemblage known at the top of the underlying Hydrequent Formation (Loboziak and Streel 1981). Rare limestone intercalation near the top of the Sainte-Godeleine Formation yielded ostracods that Lethiers (1982) attributed to the Tournaisian. A hiatus of the Strunian and possibly lower Tournaisian is possible after Hance et al. (2001) and Mansy et al. (2007).
South of the Boulonnais, the Palaeozoic strata equivalent to the Dinant Synclinorium are known through drillings below the Mesozoic-Cenozoic strata of the Paris Bassin. The Brevilliers drillings reached the Devonian and the DCB was documented by Combaz and Streel (1971). The Strunian part is typically an alternation of crinoidal grainstone and dark bioclastic shale and yielded the diagnostic miospores Retispora lepidophyta, Raistrickia spp. and Vallatisporites spp., including abnormal forms pointing to the tener event (Prestianni et al. 2016). The top of the Famennian (−195-m depth) is marked by a sandy limestone bed (probably the Hangenberg Sandstone equivalent basal bed of the Hastière Formation) followed by more massive crinoidal limestone devoid of miospores, i.e. the lower part of the lower Tournaisian Hastière Formation.
In Brittany, the Vosges and the Morvan, the Devonian successions are usually well known, but in all these areas, the DCB has not been placed with certainty as the transition towards the Carboniferous is overwhelmed by coarse-grained siliciclastic deposits traditionally regarded as the record of the Bretonic phase of the Variscan orogeny.
In the Armorican Massif of Brittany, the Devonian and Carboniferous strata occur in the Chateaulin and Morlaix Synclines. A minimum latest Famennian age (late Strunian) for the youngest Devonian deposits has been documented (Zorn Formation, Morzadec and Streel 1980; Kermerrien Formation, Rolet et al. 1986; Dourduff Breccia, Coquel and Deunff 1977). The overlying mixed silicoclastic and volcano-clastic Kertangy Formation has yielded only badly preserved late Tournaisian miospores (Morzadec et al. 1975); the DCB and lower Tournaisian are either recorded in unfossiliferous volcanic tuff or in a hiatus.
In the northern Vosges Massif, the situation is similar to the Armorican one, with a thick sequence of Upper Devonian and lower Carboniferous flyschoid mixed siliciclastic and volcano-clastic sequence (Bruche Series, Blanalt and Lillié 1973) where the DCB cannot be placed with certainty (Fluck et al. 1987). In the southern Vosges, the Chagey and Stalbert formations represent the fossiliferous Strunian (Asselberghs 1926) and passed upwards to the flyschoid Bois de la Gigoutte Formation and its lateral equivalent Markstein Series, where Maass and Stoppel (1982) documented a conodont assemblage indicating the DCB. In the Morvan Massif (north-eastern tip of the French Central Massif), the facies are similar with the southern Vosges. The thick silicoclastic sequence exposed in Savigny-Poil-Fol starts with greenish siltstone that yield a (late) Strunian assemblage of brachiopods and trilobites (Weyer 1965), overlain by sandstone and conglomerate in which intercalated siltstone beds yielded the brachiopod Ziganella attributed to the basal Hastarian (basal Carboniferous) by Legrand-Blain (1997).
In the Pyrenees, the DCB is found in the upper part of the Barousse Formation (sensu Perret 1993), which crops out on both sides of the French-Spanish border. The currently best and most detailed studied sections (Milles, Saubette, Garcet) are found on the French side (e.g. Cygan et al. 2002; Perret 1988, 1993). These sections represent pelagic carbonate facies (grey micritic limestones), which near the boundary is interrupted by the intercalation of an argillaceous level up to several decimetres-thick, which can be correlated with the Hangenberg (Black) shale and maybe also the Hangenberg Sandstone. The general facies evolution, but also the faunal content, show striking similarities with many sections in the Montagne Noire (see Feist et al. 2021, this issue). In the current context of the position of a revised DCB, it is interesting to note that Perret (1988) highlighted the stratigraphic importance of protognathodids (Pr. meischneri, Pr. collinsoni, Pr. kockeli) in the interval between the top of the siliciclastic level and the first occurrence of Si. sulcata several decimetres higher in the nodular limestone facies.
In the Milles section (Fig. 6), based on rich conodont assemblages, the DCB was placed between samples 11 and 12 by Cygan et al. (2002) using the first occurrence of Si. sulcata. Later, Kaiser et al. (2009) lowered the DCB within bed 9 due to the occurrence of Pr. kuehni in the upper part of the bed. S. sulcata was surely reported from bed 12, and as Si. cf. sulcata from bed 10. Hence, the DCB is now located only one decimetre above the level of the equivalent of the Hangenberg Sandstone. In the Saubette section, the situation is almost identical. The base of the Carboniferous is found 50 cm above the HBS equivalent (sample 14 in Perret 1988, 1993). The Garcet section is important for the absence of a lithological change in the DCB Interval. It remains completely carbonate. The DCB is placed between samples F and G (Cygan et al. 2002), based on the FO of Si. sulcata. In contrast to the other Pyrenean sections, protognathodids have not been reported so far from this section.

In northern Corsica, an upper Famennian neritic succession contains a rich benthic fauna (corals, foraminifers, stromato-poroids) (Krylatov and Mamet 1966). The Capitello Limestone yielded a diverse assemblage of Quasiendothyra spp. indicating the Strunian. Baudelot et al. (1981) indicate the occurrence of conodonts pointing also to a Strunian age for the unit. The occurrence of Po. ex. gr. marburgensis, known to disappear during the Hangenberg Black Shale event, might tie up the age to the upper part of the Strunian. Up-section, the Capitello Limestone is silicified and passes to calcareous shales then to a Culm-like succession with sandstone and conglomerate intercalations. The DCB, though not placed with precision, should fall into the silicified limestone unit (Baudelot et al. 1976).
Spain and Portugal
In the Cantabrian Zone in Northwestern Spain, the uppermost Devonian and lowermost Carboniferous strata are represented by the Ermita Formation and lateral equivalents. On the Cantabrian block, the Ermita Formation is missing or highly condensed. Towards the Asturian Arc, the sections become more complete. The uppermost Ermita Formation consists of channelled quartz arenites and conglomerates that cut into the underlying quartz sandstones (Keller et al. 2008). They called this the Llombera beds. Keller et al. (2008) pointed that crinoidal shoal deposits called Baleas Limestone are at least locally sandwiched into the Llombera beds. However, in a more distal setting, the Llonbera beds are replaced by muddy limestone and dark shales, which can be topped by crinoidal limestones of the Bales Limestone (van Loevezijn and Raven 2017). The DCB is often placed in the top of the Ermita Formation and the carbonated lateral equivalents. García-López and Sanz-López (2002) summarised available biostratigraphic data. In the classical Olleros de Alba section, the top of the Ermita Formation contains a conodont assemblage, which has been attributed to the Si. Sulcata Zone, but together with Si. praesulcata and Si. sulcata appears a single element, which may already indicate the Si. duplicata Zone (sample OLL-3 of García-López and Sanz-López 2002). A similar earliest Carboniferous age was reported for the top of the Ermita Formation at Santiago de las Villas. Here a conodont assemblage contains Ps. primus, Bi. stabilis, Pr. kockeli, Pr. kuehni, Pr. meischneri and Siphonodella sp. In the Baleas Quarry, García-Alcalde and Menéndez-Alvarez (1988) placed the DCB in the middle of the Baleas Limestone, using the first occurrence of Si. sulcata in their sample J-263. The bed J-259, 75 cm below, yielded a rich conodont assemblage including Si. praesulcata and Pr. meischneri. In the Palentine Domain, Sanz-López et al. (1999) tentatively placed the DCB in the upper part of the Vidrieros Formation using conodont assemblages containing protognathodids and siphonodellids.
In the Spanish Pyrenees, the Barousse Formation (Perret 1993) ranges from the middle Famennian Pa. marginifera Zone to the Tournaisian Si. sandbergi Zone (Sanz-Lopéz 2002a, b). The unit consists of nodular limestones with an increase in the abundance of crinoids and cephalopods in the upper part. A shale bed, 0.2 to 2-m-thick (‘level B’ of Boyer et al. 1974), occurs within the unit and is considered an equivalent of the Hangenberg Black Shales (Sanz-Lopéz 2002a). Conodonts collected from the limestones just above yielded conodonts from the Si. sulcata to possibly the Si. crenulata Zones (Boersma 1973; Perret 1993).
In the southern Central Iberian Zone, Garcia-Alcalde et al. (2000) correlated in the lower part of the Casa de La Vega Formation unfossiliferous dark claystones with the Hangenberg (Black) Shale. These claystones are intercalated into limestones, which contained late Famennian to late Tournaisian conodonts.
In SW Spain and Portugal, the succession of the Iberian Pyrite Belt (IPB) and neighbouring structural units in the South Portuguese Zone exposes uppermost Devonian to lowermost Carboniferous strata (e.g. Moreno et al. 1996; Pereira et al. 2008). Within the stratigraphic section of the IPB, the Phyllite Quartzite Group (PQG) forms the sedimentary basement of the Volcanic-Siliceous Complex (VSC), which is composed of a complex patchwork of intrusive and extrusive volcanic rocks, siliciclastic sedimentary rocks and siliceous rocks. Strata of the PQG often contains late Famennian palynozones (VCo, VH), but in some cases, the latest Famennian palynozones (LL and LN) are found in shales attributed to the PQG (e.g. Nasceidos Member, Pomarão anticline, Oliveira et al. 2006; Mina de São Domingos section, Pereira et al. 2007). However, the earliest Carboniferous palynozones have so far not been found in the VSC, but the boundary is often placed within that lithostratigraphic unit.
In Southwest Portugal, the DCB is situated within the Tercenas Formation. Oliveira (1990) reported clymenids and brachiopods indicating a latest Famennian to early Carboniferous age. Pereira et al. (2007) showed the presence of the LN and VI palynozones in the Pedra Ruiva section. Hence, the DCB has to be placed in the c. 2.5-m-thick interval between the samples 279 and 280 of Pereira et al. (2007).
Conclusions
This compilation shows that a lot of data on the knowledge of the DCB exist even in regions and countries (Fig. 7), which have not been on the forefront of discussions on the boundary in recent years. Detailed modern data are surprisingly available more often than expected, but there are also many regions, which requires new detailed looks to avoid any miscorrelations. Around the globe, when facies changes to dark shales or sandstones occur, those units have often been correlated with the well-known name Hangenberg of the Rhenish succession. Although this approach might be useful, without biostratigraphic control this seems to be an overoptimistic approach for boundary identification. However, many sections recorded faunal turnover and collapse of ecosystems in the Hangenberg extinctions. This allows the placement of the boundary even if index fossils are absent or scarce.

This compilation clearly shows the need to correlate the boundary through different facies and on the different palaeocontinents. This will not be achieved with a single criterion, but requires multiple lines of evidences. If the boundary definition should work globally, we have to step back from the focus on conodont-bearing carbonate pelagic facies, and better integrate the calendar of the often, dramatic changes in the latest Devonian–basal Carboniferous biosphere and geosphere.
References
Abrahamyan, M. S., Arakelyan, R. A., & Papoyan, A. S. (1975). Granitsa Devona i Nizhnego Karbona [The Devonian and Lower Carbon-iferous boundary]. In Stratigrafiya Karbona i Geologiya Uglenosnykh Formatsii SSSR [Stratigraphy of the Carboniferous and Geology of Coal–bearing Formations of the USSR] (pp. 49–55). Moscow: Nedra [in Russian].
Ager, D. Y., & Wallace, P. (1967). The environmental history of the Boulonnais, France. Proceedings of the Geological Association, London, 77, 385–417.
Alberti, H. (1972). Zur geologischen Entwicklung der Sahara Occidental an der Wende Devon–Karbon. Newsletters on Stratigraphy, 2, 149–152.
Alekseev, A. S., Kononova, L. I., & Nikishin, A. M. (1996). The Devonian and Carboniferous of the Moscow Syneclise (Russian Platform): stratigraphy and sea-level changes. Tectonophysics, 268, 149–168.
Alekseeva, R. E., Afanasjeva, G. A., Grechishnikova, I. A., Oleneva, N. V., & Pakhnevich, A. V. (2018). Devonian and Carboniferous brachiopods and biostratigraphy of Transcaucasia. Paleontological Journal, 52(8), 829–967.
Al-Hajri, S. A., Filatoff, J., Wender, L. E., & Norton, A. K. (1999). Stratigraphy and operational palynology of the Devonian system in Saudi Arabia. GeoArabia, 4, 53–68.
Al-Juboury, A., & Al-Hadidy, A. (2008). Facies and depositional environments of the Devonian–Carboniferous succession of Iraq. Geological Journal, 43, 383–396.
Almond, J., Marshall, J., & Evans, F. (2002). Latest Devonian and earliest Carboniferous glacial events in South Africa. In 16th International Sedimentological Congress (Abstract Volume, pp. 11–12). Johannesburg: Rands Afrikaans University.
Altiner, D., Şahin, N., & Sancay, H. (2012). Carboniferous. In Paleozoic of Northern Gondwana and Its Petroleum Potential A Field Workshop (Vol. 7, pp. 100–114). Turkish Association of Petroleum Geologists, special publication. https://doi.org/10.3997/2214-4609-pdb.367.7.
Amenábar, C. R., & di Pasquo, M. M. (2008). Nuevos aportes a la palinología, cronología y paleoambiente de la Precordillera Occidental de Argentina: formaciones El Planchón, Codo (Devónico) y Del Ratón (Mississippiaño). Acta Geológica Lilloana, 21, 3–20.
Amenábar, C. R., di Pasquo, M., & Azcuy, C. L. (2009). Palynofloras of the Chigua (Devonian) and Malimán (Mississippian) formations from the Precordillera Argentina: age, correlation and discussion of the D/C boundary. Revista Española de Micropaleontología, 41, 217–239.
Andrew, A. S., Hamilton, P. J., Mawson, R., & Talent, J. A. (1994). Isotopic correlation tools in the mid-Palaeozoic and their relation to extinction events. Australian Petroleum Exploration Association Journal, 34, 268–277.
Aqrawi, A. A. M. (1998). Paleozoic stratigraphy and petroleum systems of the Western and Southwestern Deserts of Iraq. Geoarabia, 3, 229–248.
Ariunchimeg, Y., & Nyamsuren, G. (2001). Upper Famennian and Lower Carboniferous stratigraphy in Mandalovoo and Shin Jinst Areas, Mongolia. Transaction Mongolian Science and Technology Geology, 2-3, 24–34.
Ariunchimeg, Y., Bayasgalan, A., Waters, J. A., Kido, E., Suttner, T. J., Sersmaa, G., Undariya, J., & Otgonbaatar, D. (2014). IGCP 596 IGCP 580 Field Workshop Guidebook. 8-18th August, 2014, Ulaanbaatar, Mongolia (pp. 1–55). Paleontological Center, Mongolian Academy of Sciences.
Asselberghs, E. (1926). Sur l’existence du Famennien (Néodévonien) à Chagey (Belfort). Bulletin de la Société Géologique de France, 26, 67–74.
Atakul-Özdemir, A. (2017). Conodont zonation and the Devonian/Carboniferous (Famennian/Tournaisian) boundary in the Naltaş section, eastern Taurides, Turkey. Journal of Asian Earth Sciences, 134, 94–102.
Atta-Peters, D. (1996). Late Devonian and Early Carboniferous miospores from the Takoradi Shale Formation of the Sekondian Group, Western Ghana. Africa Geoscience Review, 3, 413–427.
Atta-Peters, D., & Anan-Yorke, R. (2003). Latest Devonian and Early Carboniferous pteridophytic spores from the Sekondi Group of Ghana. Revista Española de Micropaleontología, 35, 9–27.
Avchimovitch, V. I., Turnau, E., & Clayton, G. (1993). Correlation of uppermost Devonian and Lower Carboniferous miospore zonations in Byelorussia, Poland and western Europe. Annales de la Société géologique de Belgique, 115, 453–458.
Avchimovitch, V. I., Byvcheva, T. V., Higgs, K., Streel, M., & Umnova, V. T. (1988). Miospore systematics and stratigraphic correlation of Devonian–Carboniferous Boundary deposits in the European part of the USSR and western Europe. In G. Flajs, R. Feist, & W. Ziegler (Eds.), Devonian-Carboniferous Boundary. Results of recent studies, Courier Forschungsinstitut Senckenberg 100, 169–191.
Avkhimovich, V. I. (1993). Zonation and spore complexes of the Devonian and Carboniferous boundary deposits of the Pripyat Depression (Byelorussia). Annales de la Société géologique de Belgique, 115, 425–451.
Bahlburg, H., & Breitkreuz, C. (1993). Differential response of a Devonian-Carboniferous platform-deeper basin system to sea-level change and tectonics, N Chilean Andes. Basin Research, 5, 21–40.
Bardashev, I. A., Bardasheva, N. P., Weddige, K., & Ziegler, W. (2005). Stratigraphy and facies of the Middle Paleozoic parts of southern Tien-Shan in Tajikistan and Uzbekistan. Senckenbergiana lethaea, 85, 319–364.
Bardashev, I. A., Bardasheva, N. P., Weddige, K., & Ziegler, W. (2006). Stratigraphy of the Shiskat Paleozoic reference section (southern Tien-Shan, Central Tajikistan). Senckenbergiana lethaea, 86, 289–319.
Bardasheva, N. P., Bardashev, I. A., Weddige, K., & Ziegler, W. (2004). Stratigraphy and conodonts of the Lower Carboniferous of the Shishkat section (southern Tien-Shan, Tajikistan). Senckenbergiana lethaea, 84, 225–301.
Bassis, A., Hinderer, M., & Meinhold, G. (2016). Petrography and geochemistry of Palaeozoic quartz-rich sandstones from Saudi Arabia: implications for provenance and chemostratigraphy. Arabian Journal of Geosciences, 9, 400. https://doi.org/10.1007/s12517-016-2412-z.
Baudelot, S., Doubinger, J., Durand-Delga, M., & Vellutini, P. (1976). Caractéristiques et âges des cinq cycles paléozoiques du Nord-Ouest de la Corse. Bulletin de la Société Géologique de France, 7(5), 1221–1228.
Baudelot, S., Durand-Delga, M., Mirouse, R., Perret, M.-F., & Taugourdeau-Lantz, J. (1981). Le Dévonien de Galeria en Corse septentrionale, sa datation et sa place dans le cadre de la Méditerranée occidentale. Comptes Rendus de l’Académie des Sciences, Paris, Série II, 292(3), 347–355.
Becker, R. T. (1996). New faunal records and holostratigraphic correlation of the Hasselbachtal D/C-boundary auxiliary stratotype (Germany). Annales de la Société géologique de Belgique, 117, 19–45.
Becker, R. T., Aboussalam, Z. S., & Hartenfels, S. (2012). The Devonian–Carboniferous boundary at Lalla Mimouna (Northern Maider, Anti-Atlas, SE Morocco)– preliminary new data. SDS Newsletter, 27, 31–37.
Becker, R. T., Hartenfels, S., Aboussalam, Z. S., Tragelehn, H., Brice, D., & El Hassani, A. (2013). The Devonian–Carboniferous boundary at Lalla Mimouna (northern Maider) – a progress report. Documents de l’Institut Scientifique, Rabat, 27, 109–120.
Becker, R. T., Aboussalam, Z. S., El Hassani, A., Hartenfels, S., & Hünecke, H. (2020). Devonian and basal Carboniferous of the allochthonous nappes at Mrirt (eastern part of Western Meseta) - review and new data. Hassan II Academy of Science and Technology, Frontiers in Science and Engineering, 10(1), 87–126.
Becker, R.T., Hartenfels, S., & Kaiser, S.I. (2021). Review of Devonian-Carboniferous Boundary sections in the Rhenish Slate Mountains (Germany). In M. Aretz & C. Corradini (Eds.) Global review of the Devonian-Carboniferous Boundary. Palaeobiodiversity and Palaeoenvironments 101(2). https://doi.org/10.1007/s12549-020-00469-6. [this issue]
Belhaj, F. (2000). Chapter 6 - Carboniferous and Devonian Stratigraphy—the M’rar and Tadrart Reservoirs, Ghadames Basin, Libya. In M. A. Sola & D. Worsley (Eds.), Geological Exploration in Murzuq Basin (pp. 117–142). Elsevier Science B.V.
Berchenko, O. I., & Poletaev, V. I. (2007). To refine. Stratigraphic Scheme of Devonian Deposits of Donbas. Geologiski zhurnal, 4, 65–68. [in Ukrainian]
Blanalt, J.-G., & Lillié, F. (1973). Données nouvelles sur la stratigraphie des terrains sédimentaires dévono-dinantiens de la vallée de la Bruche (Vosges septentrionales). Sciences Géologiques, bulletins et mémoires, 26(1), 69–74.
Bless, M. J. M., Becker, R. T., Higgs, K., Paproth, E., & Streel, M. (1993). Eustatic cycles around the Devonian–Carboniferous boundary and the sedimentary and fossil record in Sauerland (Federal Republic of Germany). Annales de la Société géologique de Belgique, 115, 689–702.
Boersma, K. T. (1973). Devonian and Lower Carboniferous conodont biostratigraphy, Spanish Central Pyrenees. Leidse Geologische Mededelingen, 49, 303–377.
Bogush, O. I., Lipina, O. A., & Reitlinger, E. A. (1987). Foraminifera of the boundary sediments of the Devonian and Carboniferous of the Omolon Massif. In A. V. Kanygin (Ed.), Mikrofauna i biostratigrafiya fanerozoya Sibiri I smezhnykh regionov [Microfauna and the Biostratigraphy of the Phanerozoic of Siberia and Adjacent Regions] (pp. 4–16). Novosibirsk: Nauka, Sib. Otdel. (Trudy Instituta geologii i Geofiziki, Sibirskogo Otdeleniya Akademii Nauk SSSR, 651).
Boucot, A. J., Racheboeuf, P., & Niemeyer, H. (2008). Sierra de Almeida Zorritas Formation, northern Chile, Devonian brachiopods: more is better. Fossil and Strata, 54, 69–73.
Boyer, F., Krylatov, S., & Stoppel, D. (1974). Sur le probléme de l'existence d'une lacune sous le lydiennes à nodules phosphatés du Dinantien des Pyrénées et de la Montagne Noire (France, Espagne). Geologisches Jahrbuch B, 9, 1–60.
Breitkreuz, C. (1986). Das Paläozoikum in den Kordilleren Nordchiles (21°-25°). Geotektonische Forschungen, 70, 1–88.
Bryan, S. E., Fielding, C. R., Holcombe, R. J., Cook, A., & Moffitt, C. A. (2003). Stratigraphy, facies architecture and tectonic implications of the Upper Devonian to Lower Carboniferous Campwyn Volcanics of the northern New England Fold Belt. Australian Journal of Earth Sciences, 50, 377–401.
Çapkinoğlu, S. (2000). Late Devonian (Famennian) Conodonts from Denizliköyü, Gebze, Kocaeli, Northwestern Turkey. Turkish Journal of Earth Sciences, 9, 91–112.
Çapkinoğlu, S. (2005). Famennian conodonts from the Ayineburnu Formation of the Istanbul Zone (NW Turkey). Geologica Carpathica, 56(2), 113–122.
Caputo, M. V., Melo, J. H. G., Streel, M., & Isbell, J. L. (2008). Late Devonian and Early Carboniferous glacial records of South America. In C. R. Fielding, T. D. Frank, & J. L. Isbell (Eds.), Resolving the Late Paleozoic Ice Age in Time and Space, 441 (pp. 161–173). Geological Society of America, Special Papers.
Clayton, G. (1995). Carboniferous miospore and pollen assemblages from the Kingdom of Saudi Arabia. Review of Palaeobotany and Palynology, 89, 115–123.
Clayton, G., Coquel, R., Doubinger, J., Gueinn, K. J., Loboziak, S., Owens, B., & Streel, M. (1977). Carboniferous miospores from Western Europe: illustrations and zonation. Mededelingen Rijks Geologische Dienst, 29, 1–71.
Clayton, G., Graham, J. R., Higgs, K., Sevastopulo, G. D., & Welsh, A. (1986). Late Devonian and Early Carboniferous palaeogeography of Southern Ireland and Southwest Britain. Annales de la Société géologique de Belgique, 109, 103–111.
Clayton, G., Owens, B., Al-Hajri, S. A., & Filatoff, J. (2000). Latest Devonian and early Carboniferous miospore assemblages from Saudi Arabia. In S. A. Al-Hajri & B. Owens (Eds.), Stratigraphic Palynology of the Paleozoic of Saudi Arabia (Vol. 1, pp. 146–153). GeoArabia Special Publication.
Cocks, L. R. M., Fortey, R. A., & Lee, C. P. (2005). A review of Lower and Middle Palaeozoic biostratigraphy in west peninsular Malaysia and southern Thailand in its context within the Sibumasu Terrane. Journal of Asian Earth Sciences, 24(6), 703–717.
Combaz, A., & Streel, M. (1971). Microfossiles végétaux du Tournaisien inférieur dans le core-drill de Brévillers (Pas-de-Calais, France). In Streel, M., & Wagner, R. (Eds). Compte-rendus de la 8e réunion de la Commission internationale de Microflore du Paléozoïque et de l'Assemblée générale de l'IUGS, Liège, 1970. Subcommission on Carboniferous Stratigraphy. Les Congrès et Colloques de l'Université de Liège, 55, 227–240.
Conil, R., Poty, E., Simakov, K. V., & Streel, M. (1982). Foraminifères, spores et coraux du Famennien supérieur et du Dinantien du Massif d’Omolon (Extrême-Orient soviétique). Annales de la Société géologique de Belgique, 105, 145–160.
Conrad, J., Pareyn, C., & Weyant, M. (1970). Mise en évidence du Tournaisien inférieur dans la Vallée de la Saoura (Sahara nord-occidental) et conséquences paléogéographiques. Comptes Rendus Académie Sciences Paris, 271, 900–903.
Conrad, J., Massa, D., & Weyant, M. (1986). Late Devonian regression and early Carboniferous transgression on the Northern African platform. Annales de la Société géologique de Belgique, 109, 113–122.
Cooper, M. R. (1986). Facies shifts, sea-level changes and event stratigraphy in the Devonian of South Africa. South African Journal of Science, 82, 255–258.
Coquel, R., & Abdesselam-Rouighi, F. (2000). Révision palynostratigraphique du Dévonien terminal-Carbonifère inférieur dans le Grand Erg occidental (bassin de Béchar) Sahara Algérien. Revue de Micropaléontologie, 43, 353–364.
Coquel, R., & Deunff, J. (1977). Sur la découverte de spores du passage Dévonien-Carbonifère (Strunien) dans le complexe schisteux de la “Brèche du Dourduff” (région de Morlaix, Finistère) et sa signification. Compte Rendu de l’Académie des Sciences, Paris, Série D, 285(1), 15–18.
Corradini, C., Mossoni, A., Corriga, M.G., & Spalletta, C. (2021). The Devonian/Carboniferous Boundary in Sardinia (Italy). In M. Aretz & C. Corradini (Eds.) Global review of the Devonian-Carboniferous Boundary. Palaeobiodiversity and Palaeoenvironments, 101(2). https://doi.org/10.1007/s12549-020-00440-5. [this issue]
Cygan, C., Deramond, J., Majesté-Menjoulas, C., Joseph, J., & Perret, M.-F. (2002). Pyrenees Field Trip Guide Book (pp.1-128). Toulouse: Eighth European Conodont Symposium (ECOS 8).
Demirtaşlı, E., Çatal, E., Dîl, N., Kirağlı, C., & Salanci, A. (1978). Carboniferous of the Silifke area. In E. Demirtaşlı (Ed.), Field Excursion on the Carboniferous of Turkey Guidebook, IUGS Subcommission on Carboniferous Stratigraphy (pp. 31–37). Ankara: Maden Tetkik ve Arama Enstuts.
Denayer, J. (2016). Rugose corals across the Devonian–Carboniferous boundary in NW Turkey. Acta Palaeontologica Polonica, 61, 51–70.
Denayer, J., & Webb, G.E. (2017). Earliest Mississippian rugose corals of eastern Australia: post-disaster fauna across the Devonian-Carboniferous Boundary? In B. Mottequin, L. Slavík, & P. Königshof (Eds.) Climate change and biodiversity patterns in the mid-Palaeozoic. Palaeobiodiversity and Palaeoenvironments, 97(3), 655–667.
Denayer, J., Prestianni, C., Mottequin, B., Hance, L., & Poty, E. (2021). The Devonian–Carboniferous boundary in Belgium and surrounding areas. In M. Aretz & C. Corradini (Eds.) Global review of the Devonian-Carboniferous Boundary. Palaeobiodiversity and Palaeoenvironments, 101(2). https://doi.org/10.1007/s12549-020-00440-5. [this issue]
Díaz-Martínez, E. (2004). La glaciación del Devónico superior en Sudamérica: estado del conocimiento y perspectivas. In XII Congreso Peruano de Geología, Resúmenes Extendidos (Vol. 6, pp. 440–443). Sociedad Geológica del Perú, Publicación Especial.
Díaz-Martínez, E., & Isaacson, P. E. (1994). Late Devonian glacially-influenced marine sedimentation in western Gondwana: the Cumana Formation, Altiplano, Bolivia. In A. F. Embry, B. Beauchamp, & D. J. Glass (Eds.), Pangea: Global Environments and Resources, 17 (pp. 511–522). Calgary: Canadian Society of Petroleum Geology, Memoir.
Díaz-Martínez, E., Vavrdová, M., Bek, J., & Isaacson, P. (1999). Late Devonian (Famennian) Glaciation in Western Gondwana: evidence from the Central Andes. Abhandlungen der Geologischen Bundesanstalt, 54, 213–237.
Draganits, E., Mawson, R., Talent, J. A., & Krystyn, L. (2002). Lithostratigraphy, conodont biostratigraphy and depositional environment of the Middle Devonian (Givetian) to early Carboniferous (Tournaisian) Lipak Formation in the Pin Valley of Spiti (NW India). Rivista Italiana di Paleontologia e Stratigrafia, 108, 7–35.
Ebbighausen, V., Bockwinkel, J., Korn, D., & Weyer, D. (2004). Early Tournaisian ammonoids from Timimoun (Gourara, Algeria). Mitteilungen des Museums der Naturkunde Berlin, Geowissenschaftliche Reihe, 7, 133–152.
Erina, M. V. (2008). Conodonts. In E. A. Yolkin, A. I. Kim, & J. A. Talent (Eds.), Devonian Sequences of the Kitab Reserve Area. Field Excursion Guidebook. International Conference “Global Alignments of Lower Devonian Carbonate and Clastic Sequences” (SDS/IGCP 499 Project joint field meeting), (pp. 1–45). Kitab State Geological Reserve, Uzbekistan, August 25 – September 3, 2008.
Feist, R., Cornée, J.-J., Corradini, C., Hartenfels, S., Aretz, M., & Girard, C. (2021). The Devonian/Carboniferous boundary in the stratotype area (SE Montagne Noire, France). In M. Aretz & C. Corradini (Eds.) Global review of the Devonian-Carboniferous Boundary. Palaeobiodiversity and Palaeoenvironments 101(2). https://doi.org/10.1007/s12549-019-00402-6. [this issue]
Flajs, G., & Feist, R. (1988). Index conodonts, trilobites and environment of the Devonian-Carboniferous boundary beds at La Serre (Montagne Noire, France). In G. Flajs, R. Feist, & W. Ziegler (Eds.) Devonian-Carboniferous Boundary. Results of recent studies. Courier Forschungsinstitut Senckenberg, 100, 53–107.
Fluck, P., Edel, J.-B., Gagny, C., Montigny, R., Piqué, A., Schneider, J.-L., & Whitechurch, H. (1987). Géologie du socle vosgien. Documents BRGM., 146, 1–97.
Fontaine, H., Salyapongse, S., Suteethron, V., Tian, P., & Vachard, D. (2005). Sedimentary Rocks of the Loei Region, Northeast Thailand: Stratigraphy, Paleontology, Sedimentology. Bangkok, Thailand: Department of Mineral Resources 165 pp.
Fontaine, H., Hoang, T. T., Juanngam, S., Kavinate, S., Salyapongsse, S., Suteethorn, V., & Vachard, D. (2009). Paleontology and stratigraphy of the Northwest Thailand: paleogeographical implications. Bangkok, Thailand: Department of Mineral Resources 207 pp.
Fröhlich, S. (2004). Evolution of a Devonian carbonate shelf at the northern margin of Gondwana (Jebel Rheris, eastern Anti-Atlas, Morocco). Ph.D. Dissertation, Eberhard-Karls-Universität Tübingen, 71 pp.
Gaetani, M., Zanchi, A., Angiolini, L., Olivini, G., Sciunnach, D., Brunton, H., Nicora, A., & Mawson, R. (2004). The Carboniferous of the Western Karakoram (Pakistan). Journal of Asian Earth Sciences, 23(2), 275–305.
Gagiev, M. H. (1979). Conodonts from the Devonian–Carboniferous boundary deposits of the Omolon Massif. Field Excursion Guidebook Tour IX, XIV Pacific Science Congress, Khabarovsk, 1979(Suppl. 2), 3–104.
García-Alcalde, J. L., & Menéndez-Alvarez, J. R. (1988). Devonian-Carboniferous boundary in the Asturo-Leonese Domain (Cantabrian Mountains, NW Spain). In G. Flajs, R. Feist, & W. Ziegler (Eds.), Devonian-Carboniferous Boundary, Results of recent studies. Courier Forschungsinstitut Senckenberg 100, 21–37.
Garcia-Alcalde, J. L., Truyols-Massoni, M., Pardo-Alonso, M., Bultynck, P., & Carls, P. (2000). Devonian chronostratigraphy of Spain. In P. Bultynck (Ed), Recognition of Devonian series and stage boundaries in geological areas. Courier Forschungsinstitut Senckenberg, 225, 131–144.
García-López, S., & Sanz-López, J. (2002). Devonian to Lower Carboniferous conodont biostratigraphy of the Bernesga Valley section (Cantabrian Zone, NW Spain). In S. García-López & F. Bastida (Eds.), Palaeozoic conodonts from northern Spain. Eight International Conodont Symposium held in Europe (pp. 163–205). Madrid: Publicaciones del Instituto Geológico y Minero de España (Serie: Cuadernos del Museo Geominero, 1).
Gatovsky, Y. A. (2015). Famennian–Tournaisian boundary on the western slope of the South Urals, Russia: new look. In B. Mottequin, J. Denayer, P. Königshof, C. Prestianni, & S. Olive (Eds.), IGCP 596 - SDS SYMPOSIUM Climate change and Biodiversity patterns in the Mid-Palaeozoic. Abstracts, STRATA Série 1 16, 51–52.
Gatovsky, Y. A., Polyansky, B. V., & Pekin, A. A. (2011). Upper Devonian and Lower Carboniferous deposits of the Southwestern Moscow Syneclise based on drilling data from Aleksandrovskaya Stratigraphic Test Well (Kaluga District). Moscow University Geology Bulletin, 66, 45–51.
Gess, R. W. (2016). Vertebrate biostratigraphy of the Witteberg Group and the Devonian–Carboniferous boundary in South Africa. In B. Linol & M. J. de Wit (Eds.), Origin and evolution of the Cape Mountains and Karoo Basin. Regional Geology Reviews (pp. 131–140). Cham: Spinger. https://doi.org/10.1007/978-3-319-40859-0_13.
Goldring, R. (1955). The Upper Devonian and Lower Carboniferous trilobites of the Pilton Beds in North Devon. Senckenbergiana lethaea, 36, 27–48.
González, C. R., & Bossi, G. E. (1986). Los depósitos carbónicos al oeste de Jagüel, La Rioja. 4° Congreso Argentino de Paleontología y Bioestratigrafía (Mendoza), Actas. 1, 231–236.
Gorsky, J. J. (1938). Carboniferous corals of Novaya Zemlya. Paleontologiya Sovetskoi Arktiki, Trudy Vsesoyuznego Instituta Arktiki, 93, 1–221.
Graham, J. R., & Sevastopulo, G. D. (2021). The stratigraphy of latest Devonian and earliest Carboniferous rocks in Ireland. In M. Aretz & C. Corradini (Eds.) Global review of the Devonian-Carboniferous Boundary. Palaeobiodiversity and Palaeoenvironments, 101(2). https://doi.org/10.1007/s12549-020-00455-y. [this issue]
Grechichnikova, I. A., & Levitskii, E. S. (2011). The Famennian-Lower Carboniferous reference section Geran-Kalasi (Nakhichevan Autonomous Region, Azerbaijan). Stratigraphy and Geological Correlation, 19, 21–43.
Grignani, D., Lanzoni, E., & Elatrash, H. (1992). Palaeozoic and Mesozoic subsurface palynostratigraphy in the Al Kufrah Basin, Libya. In Proceedings of the 3rd Symposium on the Geology of Libya (Tripoli 1987) (pp. 1159–1227). Amsterdam: Elsevier.
Hamada, T. (1967). Devonian of East Asia. In D. H. Oswald (Ed.), International Symposium on the Devonian, Volume I (pp. 583–596). Calgary: Alberta Society of Petroleum Geologists.
Hance, L., Poty, E., & Devuyst, F.-X. (2001). Stratigraphie séquentielle du Dinantien type (Belgique) et corrélation avec le Nord de la France (Boulonnais, Avesnois). Bulletin de la Société géologique de France, 172(4), 411–426.
Hartenfels, S., & Becker, R. T. (2016). Age and correlation of the transgressive Gonioclymenia Limestone (Famennian, Tafilalt, eastern Anti-Atlas, Morocco). Geological Magazine, 155(3), 586–629.
Hervé, F., Davidson, J., Godoy, E., Mpodozis, C., & Covacevich, V. (1981). The Late Paleozoic in Chile: stratigraphy, structure and possible tectonic framework. Revista da Academia do Ciencias Brasil, 53, 361–373.
Hervé, F., Faundez, V., Calderón, M., Massonne, H.-J., & Willner, A. (2007). Metamorphic and plutonic basement complexes. In T. Moreno & W. Gibbons (Eds.), The Geology of Chile (pp. 5–19). Geological Society of London.
Higgs, K., Finucane, D., & Tunbridge, I. P. (2002). Late Devonian and early Carboniferous microfloras from the Hakkarı Province of southeastern Turkey. Review of Palaeobotany and Palynology, 118, 141–156.
Hoepffner, C., Soulaimani, A., & Piqué, A. (2005). The Moroccan Hercynides. Journal of African Earth Sciences, 43, 144–165.
Holloway, D. J., & Rustán, J. J. (2012). The trilobite Reedops (Phacopidae) in the Lower Devonian of Argentina (Malvinokaffric Realm). Journal of Paleontology, 86, 253–257.
Hooker, N. P., Breuer, P., & Tourqui, H. (2011). Palynostratigraphy and palaeoenvironments of the Mississippian to Pennsylvanian succession in the subsurface, northern Saudi Arabia. In E. Hakansson & J. A. Trotter (Eds.), Programme & Abstracts: The XVII International Congress on the Carboniferous and Permian, 2011/20 (pp. 1–70). Geological Survey of Western Australia, Record.
Isaacson, P., Fisher, L., & Davidson, J. (1985). Devonian and Carboniferous stratigraphy of Sierra de Almeida, Northern Chile, preliminary results. Revista Geológica de Chile, 25-26, 113–124.
Isaacson, P. E., Palmer, B. A., Mamet, B. L., Cooke, J. C., & Sanders, D. E. (1995). Devonian–Carboniferous stratigraphy in the Madre de Dios Basin, Bolivia: Pando X-1 and Manuripi X-1 wells. In A. J. Tankard, R. Suarez, & H. J. Welsink (Eds.), Petroleum Basins of South America. American Association of Petroleum Geologists Memoirs. 62, 501–511.
Isaacson, P., Díaz-Martínez, E., Grader, G., Kalvoda, J., Babek, O., & Devuyst, F. X. (2008). Late Devonian–earliest Mississippian glaciation in Gondwanaland and its biogeographic consequences. Palaeogeography, Palaeoclimatology, Palaeoecology, 268, 126–142.
Ivanovski, A. B. (1967). Etyudy o rannekamennougolinikh rougozakh [Study on Lower Carboniferous rugose corals] (pp. 1–92). Moscow: Nauka. [In Russian]
Izokh, N. G. (2017). Late Devonian – Early Carboniferous conodonts from the Northeastern Siberia, Arctic Russia. In J.-C. Liao & J. I. Valenzuela-Rios (Eds.), Fourth International Conodont Symposium. ICOS IV. “Progress on Conodont Investigation”, Cuadernos del Museo Geominero (Vol. 22, pp. 201–204). Madrid: Instituto Geologico y Minero de Espana.
Izokh, N. G., & Yazikov, A. Y. (2017). Discovery of Early Carboniferous conodonts in Northern Kharaulakh Ranges (lower reaches of the Lena River, northeastern Siberia, Arctic Russia). Revue de micropaléontologie, 60, 213–232.
Janvier, P., Lethiers, F., Monod, O., & Balkaş, Ö. (1984). Discovery of a vertebrate fauna at the Devonian-Carboniferous Boundary in SE Turkey (Hakkari Province). Journal of Petroleum Geology, 7, 147–168.
Jones, P. J. (1995). Australian Phanerozoic Time-scales 5. Carboniferous. Biostratigraphic charts and explanatory notes. Bureau of Mineral Resources. Geology & Geophysics Record, 1995(34), 1–45.
Kaiser, S. I., & Corradini, C. (2011). The early siphonodellids (Conodonta, Late Devonian-Early Carboniferous): overview and taxonomic state. Neues Jahrbuch für Geologie und Paläontologie, Abhandlungen, 261, 19–35.
Kaiser, S. I., Becker, R. T., & El Hassani, A. (2007). Middle to Late Famennian successions at Ain Jemaa (Moroccan Meseta) implications for regional correlation, event stratigraphy and synsedimentary tectonics of NW Gondwana. In R. T. Becker & W. T. Kirchgasser (Eds.), Devonian events and correlations (Vol. 278, pp. 237–260). Special Publications Geological Society of London.
Kaiser, S. I., Becker, R. T., Spalletta, C., & Steuber, T. (2009). High-resolution conodont stratigraphy, biofacies, and extinctions around the Hangenberg Event in pelagic successions from Austria, Italy, and France. In D.J. Over (Ed.) Studies in Devonian Stratigraphy: Proceedings of the 2007 International Meeting of the Subcommission on Devonian Stratigraphy and IGCP 499. Palaeontographica Americana, 63, 99–143.
Kaiser, S. I., Becker, R. T., Steuber, T., & Aboussalam, Z. S. (2011). Climate-controlled mass extinctions, facies, and sea-level changes around the Devonian-Carboniferous boundary in the eastern Anti-Atlas (SE Morocco). Palaeogeography, Palaeoclimatology, Palaeoecology, 310, 340–364.
Kaiser, S. I., Becker, R. T., Hartenfels, S., & Aboussalam, Z. S. (2013). Middle Famennian to middle Tournaisian stratigraphy at El Atrous (Amessoui syncline, southern Tafilalt). Documents de l'Institut Scientifique, Rabat, 27, 77–87.
Kaiser, S. I., Aretz, M., & Becker, R. T. (2015). The global Hangenberg Crisis (Devonian–Carboniferous transition): review of a first-order mass extinction. In R. T. Becker, P. Königshof, & C. E. Brett (Eds.), Devonian climate, sea level and evolutionary events (Vol. 423, pp. 355–386). Special Publications Geological Society of London.
Keller, M., Bahlburg, H., & Reuther, C. D. (2008). The transition from passive to active margin sedimentation in the Cantabrian Mountains, Northern Spain: Devonian or Carboniferous? Tectonophysics, 461, 414–427.
Kemp, E. M., Balme, B. E., Helby, R. J., Kyle, R. A., Playford, G., & Price, P. L. (1977). Carboniferous and Permian palynostratigraphy in Australia and Antarctica: a review. BMR Journal of Australian Geology & Geophysics, 2, 177–208.
Komatsu, T., Takashima, R., Naruse, H., Ta, P. H., Dang, T. H., Nguyen, D. P., Dinh, C. T., Ho, T. C., Tran, T. V., Satoru, K., Maekawa, T., & Gatovsky, Y. (2012). Devonian–Carboniferous transition in the Pho Han Formation on Cat Ba Island, northeastern Vietnam. The Journal of the Geological Society of Japan, 118, V–VI.
Komatsu, T., Kato, S., Hirata, K., Takashima, R., Ogata, Y., Oba, M., Naruse, H., Phuong, H., Phong Nguyen, D., Dang, H. T., Doan, T. N., Nguyen, H. H., Sakata, S., Kaiho, K., & Königshof, P. (2014). Devonian–Carboniferous transition containing a Hangenberg Black Shale equivalent in the Pho Han Formation on Cat Ba Island, northeastern Vietnam. Palaeogeography, Palaeoclimatology, Palaeoecology, 404, 30–43. https://doi.org/10.1016/j.palaeo.2014.03.021.
Königshof, P., Bahrami, A., & Kaiser, S.I. (2021). Devonian-Carboniferous boundary sections in Iran. In M. Aretz & C. Corradini (Eds.) Global review of the Devonian-Carboniferous Boundary. Palaeobiodiversity and Palaeoenvironments, 101(2). https://doi.org/10.1007/s12549-020-00438-z. [this issue]
Korn, D. (1999). Famennian ammonoid stratigraphy of the Ma´der and Tafilalt (eastern Anti-Atlas, Morocco). Abhandlungen der Geologischen Bundesanstalt, 54, 147–179.
Korn, D., & Sudar, M. N. (2016). The Late Devonian and Early Carboniferous ammonoids (Cephalopoda) from Milivojevića Kamenjar, Družetić (NW Serbia) and their stratigraphy. Bulletin of Geosciences, 91(1), 187–220. https://doi.org/10.3140/bull.geosci.1578.
Korn, D., Belka, Z., Fröhlich, S., Rücklin, M., & Wendt, J. (2004). The youngest African clymeniids (Ammonoidea, Late Devonian) – failed survivors of the Hangenberg Event. Lethaia, 37, 307–315.
Korn, D., Bockwinkel, J., & Ebbighausen, V. (2007). The Tournaisian and Viséan ammonoid stratigraphy in North Africa. Neues Jahrbuch für Geologie und Paläontologie, 243, 127–148.
Korn, D., Ebbighausen, V., & Bockwinkel, J. (2010). The ammonoids from the Grès du Kahla supérieur of Timimoun (Middle-early Late Tournaisian; Gourara, Algeria). Fossil Record, 13, 13–34.
Krylatov, S. M., & Mamet, B. (1966). Données nouvelles sur les terrains paléozoiques de l'Argentella-Tour Margine (Corse); attribution à la limite dévono-carbonifère du calcaire de Capitello. Bulletin de la Société géologique de France, 7(1), 73–79.
Kulagina, E.I., Zaytseva, E.L., Vevel Y.A., Stepanova, T.I., Gibshman, N.B., Nikolaeva, S.V., Kononova, L.I., & Plotitsyn, A.N. (2021). The foraminiferal zonal scale of the Devonian–Carboniferous boundary beds in Russia and Western Kazakhstan and its correlation with ammonoid and conodont scales. In M. Aretz & C. Corradini (Eds.) Global review of the Devonian-Carboniferous Boundary. Palaeobiodiversity and Palaeoenvironments, 101(2). https://doi.org/10.1007/s12549-020-00439-y. [this issue]
Kumpan, T., Kalvoda, J., Bábek, O., Grygar, T.M., & Fryda, J. (2021). The Devonian-Carboniferous boundary in the Moravian Karst (Czech Republic). In M. Aretz & C. Corradini (Eds.) Global review of the Devonian-Carboniferous Boundary. Palaeobiodiversity and Palaeoenvironments, 101(2) https://doi.org/10.1007/s12549-019-00409-z. [this issue]
Lakin, J. A., Marshall, J. E. A., Troth, I., & Harding, I. C. (2016). Greenhouse to icehouse: a biostratigraphic review of latest Devonian–Mississippian glaciations and their global effects. In R. T. Becker, P. Königshof, & C. E. Brett (Eds.), Devonian climate, sea level and evolutionary events (Vol. 423, pp. 439–464). Special Publications Geological Society of London.
Lanzonih, E., & Magloire, L. (1969). Associations palynologiques et leurs applications stratigraphiques dans le Dévonien supérieur et le Carbonifère inférieur du Grand Erg Occidental (Sahara algérien). Revue de l'Institut français du pétrole et Annales des combustibles liquides, 24(4), 441–468.
Lebedev, O. A., Zakharenko, G. V., Silantiev, V. V., & Evdokimova, I. O. (2018). New finds of fishes in the lower uppermost Famennian (Upper Devonian) of Central Russia and habitats of the Khovanshchinian vertebrate assemblages. Estonian Journal of Earth Sciences, 67(1), 59–75.
Legrand-Blain, M. (1979). Répartition des Productidina et des Spiriferidina dans le Carbonifère du Sahara algérien. Comptes Rendus, 8ième Congrès International Stratigraphie et Géologie du Carbonifère, Moscou, 1975, 6, 203–210.
Legrand-Blain, M. (1985a). North Africa. Iullemedden Basin. In R. H. Wagner, C. F. W. Prins, & I. F. Granados (Eds.), The Carboniferous of the World: 2. Australia, Indian Subcontinent, South Africa, South America, and North Africa (pp. 323–325). IUGS Publication 20.
Legrand-Blain, M. (1985b). North Africa. Taoudenni Basin. In R. H. Wagner, C. F. W. Prins, & I. F. Granados (Eds.), The Carboniferous of the World: 2. Australia, Indian Subcontinent, South Africa, South America, and North Africa (pp. 327–329). IUGS Publication 20.
Legrand-Blain, M. (1997). Strunian-Tournaisian brachiopod faunas from Morvan, N. Massif Central, France. In M. Podemski (Ed.), Proceedings of the XIII International Congress on the Carboniferous and Permian: 28th August-2nd September, 1995, Kraków, Poland (pp. 301–308). Warszawa: Polish Geological Institute.
Lemosquet, Y., Pareyn, C., & Weyant, M. (1980). Transgression dinantienne et mouvements hercyniens précoces sur la bordure septentrionale du craton africain (Bassin de Béchar, Sahara Sud-Oranais, Algérie). Comptes Rendus, 8ième Congrès International Stratigraphie et Géologie du Carbonifère, Moscou, 1975, 6, 199–209.
Lethiers, F. (1982). Les Ostracodes du Dévonien supérieur (Nord de la France, Belgique, Ouest du Canada). Unpublished PhD Thesis, Lille University.
Loboziak, S., & Melo, J. H. G. (2002). Devonian miospore successions of Western Gondwana: update and correlation with Southern Euramerican miospore zones. Review of Palaeobotany and Palynology, 121, 133–148. https://doi.org/10.1016/S0034-6667(01)00098-7.
Loboziak, S., & Streel, M. (1981). Miospores in Middle-Upper Frasnian to Famennian sediments partly dated by conodonts (Boulonnais, France). Review of Palaeobotany and Palynology, 34, 49–66.
Loboziak, S., Streel, M., Caputo, M. V., & Melo, J. H. G. (1992). Middle Devonian to lower Carboniferous miospore stratigraphy in the central Parnaíba Basin (Brazil). Annales de la Société Géologique de Belgique, 115, 215–226.
Loboziak, S., Melo, J. H. G., Steemans, P., & Barrilari, I. M. R. (1995). Miospore evidence for pre-Emsian and latest Famennian sedimentation in the Devonian of the Paraná Basin, south Brazil. Anais da Academia Brasileira de Ciências, 67, 391–392.
Loboziak, S., Melo, H. H., & Streel, M. (1999). Latest Devonian and Early Carboniferous palynostratigraphy of Northern Brazil and North Africa – a proposed integration of Western European and Gondwanan miospore biozonations. Bulletin du Centre de recherches Elf exploration production, 22(1998), 241–259.
Loboziak, S., Caputo, M. V., & Melo, J. H. G. (2000). Middle Devonian-Tournaisian miospore biostratigraphy in the southwestern outcrop belt of the Parnaíba basin, north-central Brazil. Revue de Micro-paléontologie, 43, 301–318.
Loevezijn, G. B. S., van & Raven, J. G. M. (2017). Frasnian carbonate shoals and sequence stratigraphy of the Upper Devonian series from the southern Cantabrian Mountains, northern Spain. Boletín Geológico y Minero, 128, 931–961.
López Gamundi, O. R., & Rossello, E. A. (1993). Devonian-Carboniferous unconformity in Argentina and its relation to the Eo-Hercynian orogeny in southern South America. Geologische Rundschau, 82, 136–147.
Maass, R., & Stoppel, D. (1982). Nachweis von Oberdevon bei Markstein (Bl. Munster, Südvogesen). Zeitschrift der Deutschen Geologischen Gesellschaft, 133(2), 403–408.
Mansy, J.-L., Guennoc, P., Robaszynski, E., Amédro, E., Auffet, L. P., Vidier, J.-P., Lamarche, L., Lefèvre, O., Sommé, L., Brice, D., Mistiaen, B., Prud’homme, A., Rohart, J.-C., & Vachard, D. (2007). Notice explicative, Carte géologique de France (1/50000), feuille Marquise (seconde édition). Orléans: BRGM 213 pp.
Marshall J.A. (2021). A terrestrial Devonian-Carboniferous boundary section in East Greenland. In M. Aretz & C. Corradini (Eds.) Global review of the Devonian-Carboniferous Boundary. Palaeobiodiversity and Palaeoenvironments, 101(1). https://doi.org/10.1007/s12549-020-00448-x. [this issue]
Marshall, J., Reeves, E. J., Bennett, C., Davies, S., Kearsey, T., Millward, D., Smithson, T., & Browne, M. (2019). Reinterpreting the age of the uppermost ‘Old Red Sandstone’ and Early Carboniferous in Scotland. Earth and Environmental Science Transactions of the Royal Society of Edinburgh, 109, 265–278.
Marshall, J. E. A., Lakin, J., Troth, I., & Wallace-Johnson, S. M. (2020). UV-B radiation was the Devonian-Carboniferous terrestrial extinction killmechanism. Science Advances, 6, eaba0768. https://doi.org/10.1126/sciadv.aba0768.
Massa, D., Coquel, R., Loboziak, S., & Taugourdeau-Lantz, J. (1979). Essai de synthèse stratigraphique et palynologique du Carbonifère en Libye. Annales Société géologique du Nord, 99, 429–442.
Matyja, H., Woroncowa-Marcinowska, T., Filipiak, P., Brański, P. & Sobień, K. (2021). The Devonian/Carboniferous boundary interval in Poland: multidisciplinary studies in pelagic (Holy Cross Mountains and Sudetes) and ramp (Western Pomerania) successions. In M. Aretz & C. Corradini (Eds.) Global review of the Devonian-Carboniferous Boundary. Palaeobiodiversity and Palaeoenvironments, 101(2). https://doi.org/10.1007/s12549-020-00442-3. [this issue]
Mawson, R., & Talent, J. A. (1997). Famennian-Tournaisian conodonts and Devonian/Early Carboniferous transgressions and regressions in north-eastern Australia. Geological Society of America Special Paper, 321, 189–233. https://doi.org/10.1130/0-8137-2321-3.189.
McNestry, A. (1988). The palynostratigraphy of two uppermost Devonian–Lower Carboniferous borehole sections in South Wales. Review of Palaeobotany and Palynology, 56, 69–87.
Melo, J. H. G. (2005). Palynostratigraphy of some Palaeozoic rock units of Bolivia: Additional results. Actas del VI Congreso de Exploración y Desarrollo de Hidrocarburos (VI CONEXPLO) (pp. 1–20). Mar del Plata, Argentina. Trabajos Técnicos: CD-Rom.
Melo, J. H. G., & Loboziak, S. (2003). Devonian–Early Carboniferous miospore biostratigraphy of the Amazon Basin, Northern Brazil. Review of Palaeobotany and Palynology, 124, 131–202.
Melo, J. H. G., & Playford, G. (2012). Miospore palynology and biostratigraphy of Mississippian strata of the Amazona Basin, northern Brazil. Part Two. AASP Contribution Series, 47, 93–201.
Meor, H. H., & Lee, C. P. (2003). The Sanai Limestone Member – a Devonian limestone unit in Perlis. Geological Society of Malaysia Bulletin, 46, 137–141.
Mergl, M., & Massa, D. (2000). Chapter 4 - A Palaeontological Review of the Devonian and Carboniferous Succession of the Murzuq Basin and the Djado Sub-Basin. In M. A. Sola & D. Worsley (Eds.), Geological Exploration in Murzuq Basin (pp. 41–88). Elsevier Science B.V.
Metcalfe, I. (2002). Devonian and Carboniferous conodonts from the Kanthan Limestone, Peninsular Malaysia and their stratigraphic and tectonic implications. In L. V. Hills, C. M. Henderson, & E. W. Bamber (Eds.), Carboniferous and Permian of the World: XIV ICCP Proceedings. Memoir of the Canadian Society of Petroleum Geologists 19, 552–579.
Mezhvilk, A. A. (1958). Stratigrafiya Severnogo Kharaulakha [Stratigraphy of the Northern Kharaulakh]. Sovetskaya Geologiya, 7, 43–61. [in Russian]
Milana, J. P., & Di Pasquo, M. (2019). New chronostratigraphy for a lower to upper Carboniferous strike-slip basin of W-Precordillera (Argentina): Paleogeographic, tectonic and glacial importance. Journal of South American Earth Sciences, 96, 102383.
Milani, E. J., Melo, J. H. G., Souza, P. A., Fernandez, L. A., & França, A. B. (2007). Bacia do Paraná. Boletim de Geociências da Petrobras, 15, 265–287.
Millward, D., Davies, S., Brand, P., Browne, M., Bennett, C., Kearsey, T., Sherwin, J. E., & Marshall, J. (2018). Palaeogeography of tropical seasonal coastal wetlands in northern Britain during the early Mississippian Romer's Gap. Earth and Environmental Science Transactions of the Royal Society of Edinburgh, 109, 279–300.
Mistiaen, B., Brice, D., Hubert, B. L. M., & Pinte, E. (2015). Devonian palaeobiogeographic affinities of Afghanistan and surrounding areas (Iran, Pakistan). Journal of Asian Earth Sciences, 102, 102–126.
Molloy, P. T., Talent, J. A., & Mawson, R. (1997). Late Devonian-Tournaisian conodonts from the Eastern Khyber region, North-West Pakistan. Rivista Italiana di Paleontologia e Stratigrafia, 103, 123–148.
Moreno, C., Sierra, S., & Sáez, R. (1996). Evidence for catastrophism at the Famennian-Dinantian boundary in the Iberian Pyrite Belt. In P. Strogen, I. D. Somerville, & G. L. L. Jones (Eds.), Recent Advances in Lower Carboniferous Geology (Vol. 107, pp. 153–162). Geological Society London, Special Publications.
Mory, A. J., & Crane, D. T. (1982). Early Carboniferous Siphonodella (Conodonta) faunas from eastern Australia. Alcheringa, 6, 275–303.
Mory, A.J., & Hocking, R.M. (2011). Permian, Carboniferous and Upper Devonian Geology of the Northern Canning Basin, Western Australia: a field guide (pp. 1–36) Geological Survey of Western Australia, Record 2011/16.
Morzadec, P., & Streel, M. (1980). Remaniement de spores dévoniennes dans le Famennien de la rade de Brest (France). Geobios, 13, 115–119.
Morzadec, P., Paris, F., Pelhate, A., & Streel, M. (1975). Datation par les spores du début de la sédimentation carbonifère dans l'ouest du Synclinorium de Chateaulin, Massif Armoricain. Comptes Rendus Hebdomadaires des Séances de l'Académie des Sciences. Série D, Sciences Naturelles, 281, 1955–1957.
Munkhjargal, A., Königshof, P., Hartenfels, S., Jansen, U., Nzik, A., Carmichael, S. K., Waters, J. A., Gonchigorj, S., Crônier, C., Yarinpil, A., Paschall, O., & Dombrowski, A. (2021a). The Hushoot Shiveetiin gol section (Baruunhuurai Terrane, Mongolia): sedimentology and facies from a Late Devonian island arc setting. Palaeobiodiversity and Palaeoenvironments. https://doi.org/10.1007/s12549-020-00445-0.
Munkhjargal, A., Königshof, P., Waters, J. A., Carmichael, S. K., Gonchigdorj, S., Thassanapak, H., Udchachon, M., & Davaanyam, S. (2021b). The Mandalovoo - Gurvansayhan Terranes in the southern Gobi of Mongolia: new insights from the Bayankhoshuu Ruins section. Palaeobiodiversity and Palaeoenvironments. https://doi.org/10.1007/s12549-020-00471-y.
Naquishbandi, S. F., Sherwani, G. H., & Redha, D. N. (2010). Palynological study of Ora and the upper part of the Kaista Formation in Zakho area, Iraqi Kurdistan region. Kirkuk University Journal, Scientific studies, 5, 50–73.
Narkiewicz, K., Rakociński, M., Corradini, C., & Racki, G. (2017). New conodont data from the Devonian-Carboniferous boundary interval in the Kitab Reserve Area (Uzbekistan). In Fourth International Conodont Symposium. ICOS IV - Progress on Conodont Investigation. (Vol. 22, pp. 183–185). Cuadernos del Museo Geominero.
Narkiewicz, K., Corradini, C., Abdiev, N., & Narkiewicz, M. (2021). Conodont biostratigraphy and biofacies across the Devonian-Carboniferous boundary in the Kule section (Uzbekistan). Geological Quarterly 65, 17. https://doi.org/10.7306/gq.1588.
Nemirovskaya, T. I., Chermnykh, V. A., Kononova, L. I., & Pazukhin, V. N. (1993). Conodonts of the Devonian-Carboniferous boundary section, Kozhim, Polar Urals, Russia. Annales de la Société géologique de Belgique, 115(2), 629–647.
Nicoll, R. S., & Druce, E. C. (1979). Conodonts from the Fairfield Group, Canning Basin, Western Australia. Australia, Bureau of Mineral Resources, Bulletin, 190, 134p.
Niemeyer, H., Urzúa, F., & Rubinstein, C. (1997). Nuevos antecedentes estratigráficos y sedimentológicos de la Formación Zorritas, Devónico-Carbonífero de Sierra de Almeida, Región de Antofagasta, Chile. Revista Geológica de Chile, 24, 25–43.
Nikolaeva, S. V., Kulagina, E. I., Mustapayeva, S. N., Alekseev, A. S., Gatovsky, Y. A., Denayer, J., Ohar, V., Kurilenko, A. V., & Mychko, E. V. (2021). Progress in the study of the Devonian-Carboniferous boundary sections in the Berchogur Depression (Mugodzhary Mountains, Western Kazakhstan). Paläontologische Zeitschrift, (in press).
Okay, A. I., Satir, M., & Siebel, W. (2006). Pre-Alpide Palaeozoic and Mesozoic orogenic events in the Eastern Mediterranean region. In D. G. Gee & R. A. Stephenson (Eds.), European Lithosphere Dynamics (Vol. 32, pp. 389–405). Geological Society of London, Memoirs.
Oliveira, J. T. (1990). Stratigraphy and syn-sedimentary tectonism in the South Portuguese Zone. In R. D. Dallmeyer & E. Martínez Garcia (Eds.), Pre-Mesozoic Geology of Iberia (pp. 334–347). Springer Verlag.
Oliveira, J. T., Relvas, J., Pereira, Z., Matos, J., Rosa, C., Rosa, D., Munhá, J. M., Jorge, R., & Pinto, A. (2006). O Complexo Vulcano-Sedimentar da Faixa Piritosa: estratigrafia, vulcanismo, mineralizações associadas e evolução tectono-estratigráfica no contexto da Zona Sul Portuguesa. In R. Dias, A. Araújo, P. Terrinha, & J. C. Kullberg (Eds.), Geologia de Portugal no contexto da Ibéria (pp. 207–243). Évora: Universidade Évora.
Oliver Jr., W. A., Pedder, A. E. H., Weiland, R. J., & van Ufford, A. Q. (1995). Middle Palaeozoic corals from the southern slope of the Central Ranges of Irian Jaya, Indonesia. Alcheringa, 19(1), 1–15.
Over, D.J. (2021). The Devonian-Carboniferous boundary in the United States. In M. Aretz & C. Corradini (Eds.) Global review of the Devonian-Carboniferous Boundary. Palaeobiodiversity and Palaeoenvironments, 101(2) https://doi.org/10.1007/s12549-020-00428-1. [this issue]
Pareyn, C. (1961). Les Massifs Carbonifères du Sahara Sud-Oranais (pp. 1–326 (vol. 1), pp. 1–324 (vol. 2)). Paris: Publications du Centre de Recherches Sahariennes (Éditions du Centre National de la Recherche Scientifique) Série Géologie.
Paschall, O., Carmichael, S. K., Königshof, P., Waters, J. A., Ta, P. H., Komatsu, T., & Dombrowski, A. (2019). The Devonian-Carboniferous boundary in Vietnam: sustained ocean anoxia with a volcanic trigger for the Hangenberg Crisis? Global and Planetary Change, 175, 64–81.
Pazukhin, V. N., Kulagina, E. I., & Sedaeva, K. M. (2009). Devonian and Carboniferous boundary on the western slope of the southern Urals. In V. N. Puchkov, E. I. Kulagina, S. V. Nikolaeva, & N. N. Kochetova (Eds.), Carboniferous type sections in Russia, potential and global stratotypes. Proceedings of the international scientific conference Ufa – Sibai , 13–18 August , 2009 (pp. 22–33). Ufa: DesignPolygraphServis. [in Russian]
Pereira, Z., Matos, J. X., Fernandes, P. M. C., & Oliveira, J. T. (2007). Devonian and Carboniferous palynostratigraphy of the South Portuguese Zone, Portugal – an overview. Comunicações Geológicas, 94, 53–79.
Pereira, Z., Matos, J. X., Fernandes, P. M. C., & Oliveira, J. T. (2008). Palynostratigraphy and systematic palynology of the Devonian and Carboniferous successions of the South Portuguese Zone, Portugal (pp.1–181). Instituto Nactional de Engenharia, Tecnologia e Inovação.
Perret, M.-F. (1988). Le passage du Dévonien au Carbonifère dans les Pyrénées. Zonation par conodontes. In G. Flajs, R. Feist, & W. Ziegler (Eds.), Devonian-Carboniferous Boundary. (Results of recent studies.), Courier Forschungsinstitut Senckenberg 100, 39–52.
Perret, M. F. (1993). Recherches micropaléontologiques et biostrati-graphiques (conodontes, foraminifères) dans le Carbonifère Pyrénéen. Strata, 21, 1–597.
Petter, G. (1959). Goniatites dévoniennes du Sahara. Publications du Service de la Carte géologique d'Algérie (Nouvelle Série, Paléontologie), 2, 1–313.
Petter, G. (1960). Clymeniés du Sahara. Publications du Service de la Carte géologique d'Algérie (Nouvelle Série, Paléontologie), 6, 53–56.
Playford, G. (1990). Australian Lower Carboniferous miospores relevant to extra-Gondwanan correlations: an evaluation. In P. L. Brenckle & W. L. Manger (Eds.), Intercontinental Correlation and Division of the Carboniferous System. Courier Forschungsinstitut Senckenberg 130, 85–125.
Plodowski, G., & Salanci, A. (1990). Devon-Karbon Greze in Anatolien. Courier Forschungsinstitut Senckenberg, 127, 238–249.
Plotitsyn, A. (2016). Correlation levels in Upper Famennian-Tournaisian deposits of the North of Urals and Chernyshev Swell. Vestnik (IG Komi SC UB RAS), 7, 46–53.
Poletaev, V. I. (1981). Lithostratigraphic dissection of limestone formation of the Lower Carboniferous of Donetsk Basin. Ukrainian Institute of Geological Sciences, 49, 18–34. [in Ukrainian]
Poletaev, V. I., Brazhnikova, N. E., Vasilyuk, N. P., & Vdovenko, M. V. (1990). Local zones and major Lower Carboniferous biostratigraphic boundaries of the Donets Basin (Donbass), Ukraine. U.S.S.R. Courier Forschungsinstitut Senckenberg, 130, 47–59.
Poletaev, V. I., Vdovenko, M. V., Shchoglev, O. K., Boyarina, N. I., & Makarov, I. A. (2011). The stratotypes of the regional subdivisions of Carboniferous and Lower Permian Don-Dnieper Depression and their biostratigraphy, 236 pp. Kyiv: Logos. [in Ukrainian]
Poletayev, V. I., Vdovenko, M. V., Shulga, V. F., Nemirovska, T. I., & Shchogol’ev, N. I. (2013). The Carboniferous System. In P. F. Gozhyk (Ed.), Stratigraphy of Upper Proterozoic and Phanerozoic of Ukraine. Volume 1. Stratigraphy of Upper Proterozoic, Paleozoic and Mesozoic of Ukraine (pp. 247–355). National Academy of Sciences of Ukraine, Institute of Geological Sciences: Kyiv [in Ukrainian].
Poty, E. (1999). Famennian and Tournaisian recoveries of shallow water Rugosa following late Frasnian and late Strunian major crises, southern Belgium and surrounding areas, Hunan (South China) and the Omolon region (NE Siberia). Palaeogeography, Palaeoclimatology, Palaeoecology, 154, 11–26.
Poty, E., Devuyst, F.-X., & Hance, L. (2006). Upper Devonian and Mississippian foraminiferal and rugose coral zonations of Belgium and northern France: a tool for Eurasian correlations. Geological Magazine, 143, 829–857.
Prestianni, C., Rustán, J. J., Balseiro, D., Vaccari, E., Sterren, A. F., Steemans, P., Rubinstein, C., & Astini, R. A. (2015). Early seed plants from Western Gondwana: Paleobiogeographical and ecological implications based on Tournaisian (Lower Carboniferous) records from Argentina. Palaeogeography, Palaeoclimatology, Palaeoecology, 417, 210–219.
Prestianni, C., Sautois, M., & Denayer, J. (2016). Disrupted continental environments around the Devonian-Carboniferous Boundary: introduction of the tener event. Geologica Belgica, 19(1-2), 135–145.
Qie, W., Sun, Y., Guo, W., Ting, N, Chen, B., Song, J., Liang, K., Yin, B., Han, S., Chang, J., Wang, XD. (2021). The Devonian-Carboniferous boundary in China. In M. Aretz & C. Corradini (Eds.) Global review of the Devonian-Carboniferous Boundary. Palaeobiodiversity and Palaeoenvironments, 101(2). https://doi.org/10.1007/s12549-021-00494-z. [this issue]
Rakociński, M., Pisarzowska, A., Corradini, C., Narkiewicz, K., Dubicka, Z., Abdiyev, N. (2021). Mercury spikes as evidence of extended arc-volcanism around the Devonian–Carboniferous boundary in the South Tian Shan (southern Uzbekistan). Scientific Reports 11(1).
Randon, C., Wonganan, N., Caridroit, M., Perret-Mirouse, M.-F., & Degardin, J.-M. (2006). Upper Devonian-Lower Carboniferous conodonts from Chiang Dao cherts, Northern Thailand. Rivista Italiana di Paleontologia e Stratigrafia, 112, 191–206.
Roberts, J., Jones, P. J., & Jenkins, T. B. H. (1993). Revised correlations for Carboniferous marine invertebrate zones of eastern Australia. Alcheringa, 17, 353–376.
Rolet, J., Le Gall, В., Darboux, J. R., Thonon, P., & Gravelle, M. (1986). L'évolution géodynamique dévono-carbonifère de l'extrémité occidentale de la chaîne hercynienne d'Europe sur le transect Armorique-Cornwall. Bulletin de la Société géologique de France, 2(8), 43–54.
Rubinstein, C., Niemeyer, H., & Urzúa, F. (1996). Primeros resultados palinológicos en la Formación Zorritas, Devónico-Carbonífero de Sierra de Sierra de Almeida, Región de Antofagasta, Chile. Revista Geológica de Chile, 23, 81–95.
Rubinstein, C. V., Petus, E., & Niemeyer, H. (2017). Palynostratigraphy of the Zorritas Formation, Antofagasta region, Chile: insights on the Devonian/Carboniferous boundary in western Gondwana. Geoscience Frontiers, 8, 493–506.
Rzhonsnitskaya, M. A. (1967). Devonian of the USSR. In D. H. Oswald (Ed.), International Symposium on the Devonian, Volume I (pp. 331–348). Calgary: Alberta Society of Petroleum Geologists.
Sanz-Lopéz, J. (2002a). Devonian and Carboniferous pre-Stephanian rocks from the Pyrenees. In S. García-López & F. Bastida (Eds.), Palaeozoic conodonts from northern Spain. Eight International Conodont Symposium held in Europe (pp. 367–389). Madrid: Publicaciones del Instituto Geológico y Minero de España (Serie: Cuadernos del Museo Geominero, 1).
Sanz-Lopéz, J. (2002b). Devonian and Lower Carboniferous rocks from the Cadì nappe (eastern Pyrenees). In S. García-López & F. Bastida (Eds.), Palaeozoic conodonts from northern Spain. Eight International Conodont Symposium held in Europe (pp. 419–438). Madrid: Publicaciones del Instituto Geológico y Minero de España (Serie: Cuadernos del Museo Geominero, 1).
Sanz-López, J., García-López, S., Montesinos, J. R., & Arbizu, M. (1999). Biostratigraphy and sedimentation of the Vidrieros Formation (middle Famennian-lower Tournaisian) in the Gildar-Montó unit (northwest Spain). Bollettino della Società Paleontologica Italiana, 37, 145–158.
Saraev, S. V., Yazikov, A. Y., Bakharev, N. K., Izokh, N. G., Sobolev, E. S., & Gonta, T. V. (2013). Sedimentology and biostratigraphy of the Upper Devonian deposits in Arctic Siberia (Lena River Delta), Sedimentary basins, sedimentation and post-sedimentary processes in the geological history. Materials of VII all-Russian lithological meeting (Novosibirsk, 28-31 October 2013) (pp. 69–72). Novosibirsk III: IPGG SB RAS.
Schecoldin, R. A., Sobolev, N. N., & Matveev, V. P. (1994). Carboniferous deposits of the southern area of Novaya Zemlya. Newsletter on Carboniferous Stratigraphy, 12, 17–20.
Seidl, J. K., & Röhlich, E. (1984). Geological map of Libya, 1:250 000. Sheet: Sabha. (NG 33-2), Explanatory Booklet. Tripoli: Industrial Research Centre 138 p.
Shilo, N. A., Bouckaert, J., Afanasjeva, G. A., Bless, M. J. M., Conil, R., Erlanger, O. A., Gagiev, M. H., Lazarev, S. S., Onoprienko, Y. I., Poty, E., Razina, T. P., Simakov, K. V., Smirnova, L. V., Streel, M., & Swennen, R. (1984). Sedimentological and paleontological atlas of the late Famennian and Tournaisian deposits in the Omolon region (NE-USSR). Annales de la Société géologique de Belgique, 107, 137–247.
Shizuya, A., Oba, M., Ando, T., Ogata, Y., Takashiman, R., Nishi, H., Konatsu, T., & Nguyen, P. D. (2020). Variations in trace elements, isotopes, and organic geochemistry during the Hangenberg Crisis, Devonian–Carboniferous transition, northeastern Vietnam. Island Arc, 29e12337, 20 pp. https://doi.org/10.1111/iar.12337.
Simakov, K. V., Bless, M. J. M., Bouckaert, J., Conil, R., Gagiev, M. H., Kolesov, Y. V., Onoprienko, Y. I., Poty, E., Razina, T. P., Shilo, N. A., Smirnova, L. V., Streel, M., & Swennen, R. (1983). Upper Famennian and Tournaisian deposits of the Omolon region (NE URSS). Annales de la Société géologique de Belgique, 106, 335–399.
Spalletta, C., Corradini, C., Feist, R., Korn, D., Kumpan, T., Perri, M.C., Pondrelli, M. & Venturini, C. (2021). The Devonian/Carboniferous Boundary in the Carnic Alps (Austria and Italy). In M. Aretz & C. Corradini (Eds.) Global review of the Devonian-Carboniferous Boundary. Palaeobiodiversity and Palaeoenvironments, 101(2). https://doi.org/10.1007/S12549-019-00413-3. [this issue]
Streel, M., & Theron, J. H. (1999). The Devonian–Carboniferous boundary in South Africa and the age of the earliest episode of the Dwyka glaciation: new palynological result. Episodes, 22, 41–44.
Streel, M., Higgs, K. T., Loboziak, S., Riegel, W., & Steemans, P. (1987). Spore stratigraphy and correlation with faunas and floras in the type marine Devonian of the Ardenne-Rhenish regions. Review of Palaeobotany and Palynology, 50, 211–229.
Streel, M., Paris, F., Riegel, W., & Vanguestaine, M. (1988). Acritarch, chitinozoan and spore stratigraphy from the Middle and Upper Devonian subsurface of Northeast Libya. In A. EI-Arnauti, B. Owens, & B. Thusu (Eds.), Subsurface Palynostratigraphy of Northeast Libya, Benghazi-Libya. (pp. 111–128). Garyounis University Publication.
Streel, M., Caputo, M. V., Loboziak, S., Melo, J. H. G., & Thorez, J. (2000). Palynology and sedimentology of laminites and tillites from the latest Famennian of the Parnaiba basin, Brazil. Geologica Belgica, 3, 87–96.
Streel, M., Caputo, M. V., Melo, J. H. G., & Perez-Leyton, M. (2013). What do latest Famennian and Mississippian miospores from South American diamictites tell us? Palaeobiodiversity and Palaeoenvironments, 93(3), 299–316.
Sudar, M. N., Novak, M., Korn, D., & Jovanović, D. (2018). Conodont biostratigraphy and microfacies of the Late Devonian to Early Carboniferous Milivojevića Kamenjar section (Družetić, NW Serbia). Bulletin of Geosciences, 93(2), 163–183. https://doi.org/10.3140/bull.geosci.1690.
Suttner, T. J., Kido, E., Ariunchimeg, Y., Sersmaa, G., Waters, J. A., Carmichael, S. K., Batchelor, C. J., Ariuntogos, M., Hušková, A., Slavík, L., Valenzuela-Ríos, J. I., Liao, J.-C., & Gatovsky, Y. A. (2020). Conodonts from Late Devonian island arc settings (Baruunhuurai Terrane, western Mongolia). Palaeogeography, Palaeoclimatology, Palaeoecology, 549, 1–122. https://doi.org/10.1016/j.palaeo.2019.03.001.
Swennen, R., Bless, M. J. M., Bouckaert, J., Razina, T. P., & Simakov, K. V. (1986). Evaluation of transgression-regression events in the Upper Famennian-Tournaisian strata of the Southeastern Omolon area (NE-Siberia, USSR). Annales de la Société géologique de Belgique, 109, 237–248.
Talent, J. A., Mawson, R., Andrew, A. S., Hamjlton, P. J., & Whitford, D. J. (1993). Middle Palaeozoic extinction events. Palaeogeography, Palaeoclimatology, Palaeoecology, 104, 139–152.
Talent, J., Mawson, R., Simson, A. J., & Brock, G. A. (2002). Palaeozoics of NE Queensland: Broken River region. Special publication (Macquarie University. Centre for Ecostratigraphy and Palaeobiology); No. 1. Sydney: Geological Society of Australia; Macquarie University Centre for Ecostratigraphy and Palaeobiology.
Termier, G., & Termier, H. (1950). Paléontologie Marocaine. II. Invertébrés de l’ére Primaire. Fascicule III. Mollusques. Notes et Mémoires du Service Géologique, 78, 1–246.
Theron, J. N. (1993). The Devonian-Carboniferous Boundary in South Africa. Annales de la Société géologique de Belgique, 116, 291–300.
Tolokonnikova, Z. A. (2016). New Famennian bryozoans (Upper Devonian) from Azerbaijan and Armenia. Paleontological Journal, 50(3), 245–254.
Tsegab, H., Sum, C. W., Gatovsky, A. Y., Hunter, A. W., Talib, J. A., & Kassa, S. (2017). Higher-resolution biostratigraphy for the Kinta Limestone and an implication for continuous sedimentation in the Paleo-Tethys, Western Belt of Peninsular Malaysia. Turkish Journal of Earth Sciences, 26(5), 377–394.
Vavrdová, M., Isaacson, P. E., Díaz Martínez, E., & Bek, J. (1993). Devonian–Carboniferous boundary at Lake Titikaka, Bolivia, preliminary palynological results. Comptes rendues Douzième Congrès International de la Stratigraphie et Géologie du Carbonifère et Permien, 1, 187–199.
Vavrdová, M., Bek, J., Dufka, P., & Isaacson, P. E. (1996). Palynology of the Devonian (Lochkovian to Tournaisian) sequence, Madre de Dios Basin, northern Bolivia. Věstník Českého geologického Ústavu, 71, 333–349.
Vevel, Ya.A., Zhuravlev, A.V., & Popov, V.V. (2012). Pogranichnye otlozheniya devonskoy i kamennougol'noy sistem v razreze na r. Kamenka (Pechoro-Kozhvinskiy megaval, Timano-Pechorskaya provintsiya) [Deposits of the Devonian and Carboniferous boundary in the Kamenka River section [Pechora-Kozhvinsky megaswell, Timan-Pechora province)]. Neftegazovaya Geologiya. Teoriya I Praktika, 7, [in Russian].
Walliser, O. H., El Hassani, A., & Tahiri, A. (2000). Mrirt: a key area for the Variscan Meseta of Morocco. Notes et Mémoires Service Géologique Maroc, 399, 87–108.
Wang, C. Y., & Minjin, C. (2004). Early Carboniferous age for Arynshand Formation, Mongolian South Gobi, based on conodonts. Alcheringa, 28, 433–440.
Wang, C. Y., Weddige, K., & Minjin, C. (2005). Age revision of some Palaeozoic strata of Mongolia based on conodonts. Journal of Asian Earth Sciences, 25, 759–771.
Webb, G. E. (1998). Earliest known Carboniferous shallow-water reefs, Gudman Formation (Tn1b), Queensland, Australia: Implications for Late Devonian reef collapse and recovery. Geology, 26, 951–954.
Webb, G. E. (2005). Quantitative analysis and paleoecology of earliest Mississippian microbial reefs, lowermost Gudman Formation, Queensland, Australia: not just post-disaster phenomena. Journal of Sedimentary Research, 75, 875–894.
Wendt, J. (1988). Facies pattern and palaeogeography of the Middle and Late Devonian in the eastern Anti-Atlas (Morocco). In N. J. McMillan, A. F. Embry, & D. J. Glass (Eds.), Devonian of the world. Proceedings of the 2nd International Symposium on the Devonian System, Calgary, Canadian Society of Petroleum Geology, Memoir 14, 467–480.
Wendt, J., Aigner, T., & Neugebauer, J. (1984). Cephalopod limestone deposition on a shallow pelagic ridge: the Tafilalt Platform (upper Devonian, eastern Anti-Atlas, Morocco). Sedimentology, 31, 601–625.
Weyant, M. (1976). Une série dévonienne condensée sur la bordure nord du Bassin de Béchar (Sahara sud-oranais, Algérie). 4ème Réunion Annuel Sciences de la Terre, Paris, 398.
Weyant, M. (1985). Conodonts. In R. H. Wagner, C. F. W. Prins, & I. F. Granados (Eds.), The Carboniferous of the World: 2. Australia, Indian Subcontinent, South Africa, South America, and North Africa (pp. 358–361). IUGS Publication 20.
Weyer, D. (1965). Etroeungt im Morvan (Zentralfrankreich). Mitteilungen des zentralen geologischen Instituts, 1, 289–302.
Wicander, R., Clayton, G., Marshall, J. E. A., Troth, I., & Racey, A. (2011). Was the latest Devonian glaciation a multiple event? New palynological evidence from Bolivia. Palaeogeography, Palaeoclimatology, Palaeoecology, 305, 75–83.
Yolkin, E.A., Kim, A.I., & Talent, J.A. (2008). Devonian Sequences of the Kitab Reserve Area. Field Excursion Guidebook. International Conference “Global Alignments of Lower Devonian Carbonate and Clastic Sequences” (SDS/IGCP 499 Project joint field meeting) (pp. 1–98). Kitab State Geological Reserve, Uzbekistan, August 25–September 3, 2008. Novosibirsk: Publishing house of SB RAS.
Zhaimina, V. Y., & Kulagina, E. I. (2019). Features of Devonian and Carboniferous Boundary deposits in the Karamurun Section of the Bolshoi Karatau Mountains. News of the National Academy of Sciences of the Republic of Kazakhstan, Series of Geology and Technical Sciences, 4(436), 42–53.
Zhaimina, V. Y., Buvtyshkin, V. M., Fazylov, E. M., Musina, E. S., Shabalina, L. V., & Prihodko, D. V. (2017). The Upper Devonian- Carboniferous reef structures in the Bolshoi Karatau mountains (South Kazakhstan). Field Trip Guide (pp. 1–186). Almaty-Turkestan: Subcomission on Carboniferous Stratigraphy.
Zhuravlev, A. V. (2020). Trophic position of some Late Devonian-Carboniferous (Mississippian) conodonts revealed on carbon organic matter isotope signatures: a case study of the East European basin. Geodiversitas, 42, 443–453.
Zhuravlev, A. V., & Sobolev, D. B. (2019). Devonian-Carboniferous Boundary in the east of the Pechora Plate (Kamenka River and Vangyr River Sections). Vestnik (IG Komi SC UB RAS), 10, 16–22.
Ziegler, W., & Sandberg, C. A. (1996). Reflexions on Frasnian and Famennian Stage boundary decisions as a guide to future deliberations. Newsletters on Stratigraphy, 33, 157–180.
Acknowledgements
The authors thank the many colleagues who have shared their information and views on the DCB with us. A special thanks goes to those colleagues who attended the meetings of the task group for the Revision of the Devonian-Carboniferous Boundary. The manuscript benefits from the thoughtful comments of the journal editor and reviewer P. Königshof.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
The authors declare that they have no conflict of interest.
Additional information
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
This article is a contribution to the special issue “Global review of the Devonian-Carboniferous Boundary”
Rights and permissions
About this article

Cite this article
Aretz, M., Corradini, C. & Denayer, J. The Devonian-Carboniferous Boundary around the globe: a complement. Palaeobio Palaeoenv 101, 633–662 (2021). https://doi.org/10.1007/s12549-021-00495-y
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s12549-021-00495-y