
Abstract
Continued excavations during the last decade have yielded large quantities of tetrapod remains from the Middle Triassic (Ladinian) Erfurt Formation (Lower Keuper) in Germany. The temnospondyl dental morphotypes are highly variable but represent low taxonomic diversity. This is in contrast to the reptilian tooth morphotypes, which comprise a minimum of 26 distinct types, only some of which can be referred to taxa based on diagnostic skeletal material. The assemblage includes a taxonomically diverse range of semi-aquatic or aquatic faunivores, in addition to large terrestrial carnivores and many smaller-sized forms that may have subsisted on invertebrates and small vertebrates. With only two taxa known to date, tetrapods with dentitions suitable for oral processing of plant material form the least common faunal element. The Lower Keuper assemblages are dominated by diapsid reptiles, especially archosauriforms. Unlike in the more or less coeval tetrapod communities from Gondwana, gomphodont cynodonts are represented only by a single molariform tooth to date, whereas avemetatarsalian archosaurs are entirely absent. Most remarkable is the virtually total absence of medium- to large-sized herbivores (rhynchosaurs, dicynodont synapsids).
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
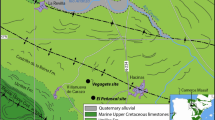
The Middle Triassic (Ladinian) Lower Keuper was one of the first Mesozoic continental formations to yield substantial skeletal material of tetrapods (Jaeger 1824, 1828; Alberti 1834; Meyer and Plieninger 1844). Apart from abundant temnospondyls (especially Mastodonsaurus and plagiosaurids), it also produced finds of enigmatic archosauriform reptiles (Zanclodon laevis, Z. crenatus: Plieninger 1847; Quenstedt 1885; ‘Z.’ schuetzii: Fraas 1900). Subsequent examination of this material by Huene (1905, 1922) revealed that the tetrapod material from the Gaildorf locality was quite diverse. Later, the extensive excavation at Kupferzell in 1977 yielded abundant and taxonomically diverse finds of tetrapods from all size classes (Witzmann and Schoch 2018). Since the late 1990s, focused excavations in quarries near Vellberg (Fig. 1) have provided much additional material, including skeletons of numerous tetrapods either entirely new to science or formerly known only from isolated bones and teeth (Schoch 1997, 2006, 2008; Gower 1999; Hellrung 2003; Witzmann et al. 2008). Among these, the most surprising discovery to date was that of the stem-turtle Pappochelys rosinae (Schoch and Sues 2015, 2017). These finds have shown that the Lower Keuper forms one of the richest sources of vertebrate fossils of late Middle Triassic age anywhere in the world (Schoch and Seegis 2016).

Left, location of the fossiliferous sites yielding Lower Keuper tetrapods, with outcropping areas of the Middle–Upper Triassic Keuper sequence shaded. Right, occurrence of particular tooth types mapped onto range chart
Although recovery and preparation of the vast quantities of tetrapod skeletal remains from Kupferzell and Vellberg are ongoing, a preliminary report detailing the known tetrapod diversity is much needed. This subject may be best approached by a study of the numerous dental morphotypes because isolated teeth are by far the most common tetrapod remains in the Lower Keuper strata. The objective of this report is to describe and illustrate all currently identified dental morphotypes of tetrapods from the Lower Keuper of southern Germany and assess their affinities. From the Lower Keuper of northern Germany, a number of isolated tetrapod teeth have already been documented from particular localities (Dorka 2002; Löffler et al. 2007). Our study aims to build a framework for future work on Triassic tetrapod assemblages as well as stimulate further search for additional fossils, particularly from the Middle and Late Triassic of Europe. It also contributes to a better understanding of the diversity and spatiotemporal distribution of Triassic microvertebrate assemblages, which have received increasing attention in recent years (Heckert 2004; Heckert et al. 2012; Sues and Fraser 2010).
Material and localities
The bulk of the material examined here was collected at two localities, Kupferzell and Vellberg in the southern German state of Baden-Württemberg (Fig. 1). These two sites form the two richest of a large number of Lower Keuper fossil Lagerstätten, which are widely distributed across southern and central Germany. The Lower Keuper sequence, ranging in thickness some 20–30 m, is usually best exposed at the top of the Muschelkalk limestones, which are commercially quarried in many locations between Stuttgart, Würzburg and Nürnberg. Highway construction south of Kupferzell-Bauersbach exposed fossiliferous mudstones (top section of Untere Graue Mergel) for about 3 months in 1977, and emergency salvage excavations led to the recovery of both skeletal remains of large tetrapods and numerous small vertebrate fossils (Wild 1980; Schoch and Wild 1999). The other site, a commercial limestone quarry near the town of Vellberg, has produced much additional vertebrate material from the Untere Graue Mergel since the early 1980s (Schoch 2002; Schoch and Seegis 2016). Smaller samples, collected at temporary exposures created by construction activities near Michelbach an der Bilz and Wolpertshausen, have also added to the picture (Hagdorn et al. 2015).
Institutional abbreviations
SMNS, Staatliches Museum für Naturkunde, Stuttgart; MHI, Muschelkalkmuseum Hagdorn, Ingelfingen, Germany.
Description
Terminology
In this survey, we employ a range of anatomical terms that require definitions (see also Edmund 1969; Smith and Dodson 2003). ‘Crown’ refers to the portion of the tooth exposed above the gum line, whereas ‘root’ is any substantial part by which the tooth is attached to the jaw. In this sense, archosauriform teeth have roots, whereas temnospondyl teeth do not. We follow Berkovitz and Shellis (2017) in the definition of the following general types of tooth attachment. ‘Thecodont’ refers exclusively to tooth implantation where elongated roots (usually longer than the crown is tall) are set in well-defined sockets in jaw elements and held by ligaments. ‘Acrodont’ refers to teeth fused to the apical margin of a jawbone. ‘Pleurodont’ refers to the attachment of a tooth to the labial wall of a jawbone (Fig. 2).

General features of tetrapod teeth, exemplified by different tooth types occurring in the Lower Keuper. a, e Labyrinthodont tooth with enamel and dental infolding, resulting grooves, and cross-section (Mastodonsaurus). b, f Bicarinate ziphodont tooth with rectangular serrations pointing to the side (Batrachotomus). c Putative herbivorous tooth with denticles pointing crownwards (archosauriform). d, g Piscivorous tooth (Nothosaurus). h–j Modes of tooth emplacement: h labyrinthodont (fangs and marginal teeth), i ziphodont (‘thecodont’), j pleurodont
Apical: Towards the tip (apex) of the tooth crown.
Basal: Towards the crown-root transition on the tooth.
Mesial: The tooth surface facing towards the mandibular symphysis or midline of the cranium. In most cases, mesial refers to the same direction as anterior, but, near the midline, the direction of the surface often changes from anterior to medial.
Distal: The opposite of mesial, namely the tooth surface facing towards the jaw joint. In most cases, distal refers to the same direction as posterior.
Lingual: The tooth surface facing towards the oral cavity/tongue.
Labial: The tooth surface facing towards the lip.
Carinate: The presence of a keel or cutting edge. Often, keels are located on two opposing sides of the tooth crown, typically on the mesial and distal surfaces of archosauriform tooth crowns. Keels may be continuous, forming a single cutting edge, or discontinuous (serrated). Carinae are common on many types of tetrapod teeth (Berkovitz and Shellis 2017). We refer to teeth with a single carina as ‘unicarinate’ and those with two carinae as ‘bicarinate’.
Serrations and denticles: Any carina that is subdivided into discrete units (serrations or denticles), each of which functions as a miniature cutting edge. Serrations are common on tooth crowns of non-archosaurian archosauromorphs, pseudosuchians and avemetatarsalians, but they are absent on all known temnospondyl teeth (Fig. 2; Schoch and Milner 2000, 2014).
Ziphodont: This type of tooth is found in many probably carnivorous reptiles (Fig. 2b–i). The crown is labiolingually flattened, often (but not always) recurved, and typically has serrated mesial and distal carinae (Langston 1975). Among extant tetrapods, ziphodont tooth crowns are present only in some varanid lizards (Auffenberg 1981; D’Amore and Blumenschine 2009, 2012).
Labyrinthodont: In most early tetrapods, the teeth are anchored to the slender jaw bones not by means of elongated roots, but by a tight attachment of their petaloid basal surfaces (‘pinch-and-swell’ structure in cross-section) (Preuschoft et al. 1991). Parallel grooves at the base of the tooth, extending parallel to the apicobasal axis of the crown, are found in all known Triassic temnospondyl teeth (Fig. 2a–h). They reflect the deep infolding of dentine and enamel, a condition termed labyrinthodont or plicidentine (Owen 1841; Bystrow 1938; Schultze 1969). Plicidentine teeth evolved convergently in some teleostean fishes, tetrapodomorphs, some parareptiles, ichthyopterygians and varanid lizards (Maxwell et al. 2011a, b).
Ridges: The petaloid structure of labyrinthodont teeth must not be confused with the apicobasally extending superficial ridges on the enamel covering of tooth crowns (Fig. 2d). Such ridges are present in a wide variety of tetrapod taxa including eosauropterygians, tanystropheid archosauromorphs and archosauriforms (see below).
Locality data
Most tooth types described here co-occur in the Vellberg locality and within the same horizon (bed E6 of Schoch and Seegis 2016). This makes them likely to belong to a true fauna, rather than a mere assemblage. The only tooth types not found in Vellberg are the traversodontid type S1. Among the labyrinthodont teeth, only type A7 (plagiosternine teeth) was not found in the same bed as the others but occurs in lagoonal deposits (Plagiosternum: bed E4 of Schoch and Seegis 2016) or channel sandstones (Megalophthalma: Hauptsandstein unit of the Lower Keuper).
Amphibian (temnospondyl) teeth
Type A1: Mastodonsaurus
As the most abundant large tetrapod, Mastodonsaurus giganteus is commonly represented by its enormous, up to 14-cm-tall fangs in numerous horizons of the Lower Keuper (Meyer and Plieninger 1844; Hagdorn and Simon 1985; Schoch 1999). Whereas the largest teeth are undoubtedly referable to this taxon (Fig. 3a–c), smaller fangs of similar shape cannot be unambiguously assigned to Mastodonsaurus (see discussion for A2).

Temnospondyl tooth types (all labyrinthodont). a–d Mastodonsaurus giganteus (a largest fang, SMNS 55911; b narrow cone, SMNS 82013; c wide cone, SMNS 84001; d marginal tooth, SMNS 91622). e, f Callistomordax kugleri (e marginal teeth, SMNS 96934; f fang, SMNS 90520). g Marginal tooth of Trematolestes hagdorni (SMNS 91546). h–j Marginal teeth of plagiosaurids (h Gerrothorax pulcherrimus, SMNS 83842; i Plagiosternum granulosum, SMNS 90720; j Plagiosuchus pustuliferus, SMNS 57921)
Size: 60–100 mm crown height (fangs), with some teeth attaining heights ranging from 120 to 140 mm.
Tooth shape: Conical, straight or gently curved lingually, usually pointed, but often with apical wear due to tooth-food contact. Marginal teeth have transversely oval to almost rectangular sockets and markedly curved crowns with pointed apices facing lingually.
Cross-section: Round to oval for fangs, transversely subrectangular for marginal teeth.
Carinae: Only occasionally present near the tips of individual marginal teeth, carinae aligned along mesial and distal surfaces.
Serrations: None.
Basis for assignment: Numerous skulls and jaws referable to Mastodonsaurus giganteus (Schoch 1999) document the morphology and variability of teeth in this taxon.
Diagnostic feature: Labyrinthine infolding is reflected by deep external grooves, which aligned parallel to the apicobasal axis of the tooth crown and extend from the base to at least mid-height of the crown. Well-preserved teeth show a change in colouration (dark brown to lighter colours) in the apical third of the crown or even alternating darker and lighter ribbons (Moser and Schoch 2007).
Variation: There is a wide variation in length-to-cross-section ratio, form and wear of tip, as well as the depth and number of grooves. Quite common are fangs with ‘stepped’ flanks.
Morphotypes: Two distinct morphotypes of large temnospondyl fangs are common in Lower Keuper strata, here labelled as A1a and A1b. The more common type (A1a) is a wide cone with a wide base, relatively widely spaced grooves, and often stepped flanks at mid-crown level. The degree of crown curvature is usually slight. The ratio between base diameter and crown height ranges from 0.4 to 0.45. The majority (70%) of known A1 teeth fall into this category. The second type (A1b) is a narrow cone with straight flanks, only gentle curvature and tightly spaced grooves. The grooves at the base of the tooth are set at regular distances and extend along at least three quarters of the height of the crown. The two types grade into one another in large samples (> 50 teeth). Referral of the entire range of A1 teeth to a single taxon, Mastodonsaurus giganteus is suggested by the considerable diversity in fang shape and size in the available cranial material. However, pending larger samples, it cannot be entirely ruled out that the two tooth types indicate the existence of more than one species.
Wear: Worn tips are common, and wear often extends basally along the flank of a crown. Tooth-food contact resulted in wear at the tip of the crown (which can attain wart- or teat-like shapes, hence the Greek Mastodonsaurus meaning ‘teat-toothed reptile’). Articulated skulls reveal that, in life, the vomerine and palatine fangs were in close proximity to the lingual wall of the lower jaw. In some specimens, both the fangs and mandibular bone show traces of such contact. A second type of wear occurs on the apices and flanks of the symphyseal fangs, which may have either been either too large for accommodation by the premaxillary openings or indicate the existence of two generations of fangs at the same time.
Type A2: Kupferzellia
Kupferzellia wildi is a second, smaller-sized capitosauroid temnospondyl from the Lower Keuper. It has a wide parabolic skull, small orbits and a more delicate palate compared to Mastodonsaurus (Schoch 1997). At some sites, it may be as common as Mastodonsaurus, with skull lengths ranging from 20 to 40 cm. Isolated fangs of Kupferzellia differ in size but not in shape from the highly variable ones of Mastodonsaurus. Adult fangs of Kupferzellia attain only half the maximum height of those in Mastodonsaurus, but the fangs and especially marginal teeth of juveniles of the two taxa cannot be readily distinguished. Here, we describe only the adult fangs associated with diagnostic cranial remains of Kupferzellia (Schoch 1997).
The distinction of tooth types A1 and A2 is not always clear-cut, and referral of type A2 to Kupferzellia must remain tentative because most known skulls of the latter taxon do not preserve complete fangs.
Size: 20–40 mm crown height.
Tooth shape: Conical, straight, and pointed fangs, and transversely oval and lingually curved marginal teeth.
Cross-section: Round to oval for fangs, transversely subrectangular for marginal teeth.
Carinae: None.
Serrations: None.
Basis for assignment: A number of well-preserved skulls of Kupferzellia wildi document the morphology and variability of teeth in this taxon.
Type A3: Trematolestes
The gracile, slender-skulled Trematolestes hagdorni was the first trematosaurid temnospondyl for which complete skeletons were found (Schoch 2006).
Size: 4–15 mm crown height (fangs); palatal teeth relatively large, 4–8 mm high; marginal teeth with 2–5 mm crown height.
Tooth shape: Labiolingually compressed, slender, tip curved lingually, pointed (Fig. 3g).
Cross-section: Sagittally oval for fangs, round for marginal teeth; at mid-crown level labiolingually compressed, lenticular.
Carinae: Both fangs and marginal teeth bicarinate; fangs with carinae aligned mesiolingually and distolabially, on marginal teeth more mesially and distally.
Serrations: None.
Basis for assignment: Several skulls of this trematosaurid representing a considerable size range have this tooth morphotype (SMNS 81790, 90022; MHI 1715).
Diagnostic feature: Labyrinthine infolding resulting in numerous deep, equidistant grooves, extending almost to the apex of the crown.
Type A4: Callistomordax
Unlike Trematolestes, Callistomordax is a short-snouted temnospondyl with a long, anguilliform body and a disproportionately long trunk (Schoch 2008).
Size: 10–14 mm crown height (fangs).
Tooth shape: Crowns of both fangs and marginal teeth labiolingually compressed, slender and pointed (Fig. 3e, f); fangs with constriction at base and mesial and distal expanded crown with prominent carinae. Compared to A3, fangs are more flattened and the tip of the crown is not curved lingually. Marginal teeth very small, straight at base, turning abruptly triangular in labial view and strongly compressed labiolingually.
Cross-section: At the base of the crown, round to oval for fangs and transversely elongated for marginal teeth bases; lenticular from mid-crown level on towards tip in both fangs and marginal teeth.
Carinae: Always bicarinate, with carinae aligned mesially and distally.
Serrations: None.
Basis for assignment: Several well-preserved skulls of Callistomordax representing different sizes show the structure and variability of the teeth (SMNS 55384, 82035, 90516, 90700; MHI K4).
Type A5: Plagiosuchinae
Plagiosuchus pustuliferus, an abundant temnospondyl in the Lower Keuper, had tiny marginal teeth and lacks fangs (Damiani et al. 2009).
Size: 3–5 mm crown height.
Tooth shape: Conical, markedly curved lingually, pointed (Fig. 3j).
Cross-section: Round to oval at base.
Carinae: None.
Serrations: None.
Basis for assignment: The dentition of this taxon is currently documented only by a single complete skull and mandible (SMNS 57921).
Diagnostic feature: Labyrinthine infolding only at the base, enamel surface of the crown otherwise smooth.
Type A6: Plagiosaurinae
The distinctive plagiosaurid Gerrothorax pulcherrimus is readily identified by the pustular ornamentation of the bones of its strongly dorsoventrally flattened skull (Hellrung 2003). Like Plagiosuchus, it lacks fangs, but its marginal teeth are quite different from those of other known plagiosaurids (Schoch and Witzmann 2012).
Size: 4–5 mm crown height.
Tooth shape: Conical, apical portion markedly curved lingually, pointed (Fig. 3h).
Cross-section: Transversely oval to mesiodistally compressed at base, oval at mid-crown level.
Carinae: Mesial and distal carinae present in some teeth.
Serrations: None.
Surface: Covered by numerous ridges near the tip. These are not parallel but diverge from a point close to the tip in radial fashion. The apical half of the crown is covered by an unusually thick layer of enamel.
Basis for assignment: Four good skulls and numerous jaw fragments document the structure and variability of these teeth (SMNS 83070, 83625, 83866, 91354).
Diagnostic feature: Labyrinthine infolding simple but producing well-developed, equidistant grooves extending from the base of the crown apically to mid-crown level.
Type A7: Plagiosterninae
Two closely related taxa share this tooth type, which differs in the height of the crowns but not in their structure. The much better-known taxon is Plagiosternum granulosum, which is represented by several skulls and mandibles. Megalophthalma ockerti (from the Hauptsandstein of Vellberg) has a relatively longer skull and even larger orbits (Schoch et al. 2014). Both taxa share relatively long and slender, lingually curved marginal teeth and the absence of fangs.
Size: 6–9 mm crown height (Plagiosternum, Fig. 3i) and 9–11 mm crown height (Megalophthalma).
Tooth shape: Slender conical, tip strongly curved lingually, pointed (in both taxa).
Cross-section: Round at base, oval at mid-crown level, lenticular near tip (in both taxa).
Carinae: Mesial and distal carinae extending from mid-crown level to tip (Plagiosternum).
Serrations: None.
Surface: Covered by distinct, fine ridges near tip.
Basis for assignment: Well-preserved cranial and mandibular remains are available for both plagiosternines (SMNS 90698, 90720, MHI 2).
Diagnostic feature: Labyrinthine infolding with well-developed, equidistant grooves extending from base to at least mid-crown level.
Reptilian teeth
Type R1: Batrachotomus
The most common large reptilian teeth can be assigned to this tooth type, which is represented by a broad range of crown sizes and shapes, but with consistent metric patterns of serration. Although it cannot be ruled out that more than one taxon was present, the large teeth probably belonged to the up to 5-m-long paracrocodylomorph archosaur Batrachotomus kupferzellensis Gower, 1999. This tooth morphotype was already described by Fraas (1900) as ‘Zanclodon’ schuetzii (Hagdorn and Mutter 2011).
Size: 10–50 mm crown height; root length 120–140% of crown height.
Tooth shape: Recurved, wide at the base of the crown (Fig. 4a–c).

Large reptile tooth types. a–c Type R1, Batrachotomus kupferzellensis (a SMNS 55911, reversed; b SMNS 82013; c SMNS 84001). d Type R2, Zanclodon laevis (SMNS 6045). e Type R3 (SMNS 96933). f Type R4 (SMNS 96935). g, h Archosauriform type R5 (SMNS 81798). i, j Type R6, doswelliid archosauriform Jaxtasuchus salomoni (SMNS 91083). k Pseudosuchian type R7 (SMNS 96936). l Archosauriform type R8 (SMNS 91312). m, n Herbivorous archosauriform type R9 (SMNS 91343). o Type R10 (SMNS 91427). p Type R25, Nothosaurus sp. (SMNS 92554). q Type R24, Tanystropheus sp. (SMNS 91385)
Cross-section: Labiolingually compressed, oval at base.
Carinae: Bicarinate.
Serrations: Always serrated; individual serrations subquadrangular and equidimensional along carina; 3 or 4 serrations per mm of carina.
Basis for assignment: Several skeletons of Batrachotomus kupferzellensis preserve teeth in their jaw elements (Gower 1999; Gower and Schoch 2009). The single isolated tooth of Zanclodon schuetzii (SMNS 53538, Fraas 1900; Huene 1907–1908) falls within the variational range of the samples for Batrachotomus (size and shape of tooth crown; 3 serrations per mm).
Variation: A wide range of crown shapes and sizes, depending on relative position in the jaws and possibly ontogenetic status. The longest teeth are located in the mid-portion of the maxilla and anterior part of the dentary and the smallest ones close to the anterior and posterior ends of the maxilla. Tooth bases vary in cross-section from oval to almost round. The apices of the crowns are often worn and rounded by tooth-on-food contact, exposing more lightly coloured deeper parts of enamel or even the dentine core. A number of tooth crowns show spalling of long flakes, probably due to forceful tooth-to-bone contact during predation (Schubert and Ungar 2005; D’Amore and Blumenschine 2009).
Type R2: Zanclodon laevis
Teeth of this type are slightly smaller than R1 and apparently much rarer (Fig. 4d). The only unambiguous record is the holotypic maxilla of Zanclodon laevis Plieninger, 1847, which is diagnostic but unfortunately of little help for determining the phylogenetic relationships of this taxon (Schoch 2011b). A second specimen, a single tooth, was reported by Huene (1907–1908 from the Lower Keuper of Zuffenhausen (Stuttgart suburban area). A diagnostic feature of the jaw in Zanclodon laevis is the presence of a solid lingual alveolar wall rather than discrete interdental plates, unlike the condition in most known non-archosaurian archosauriforms and pseudosuchians (Nesbitt 2011).
Size: 15–30 mm crown height.
Tooth shape: Recurved to straight, base moderately wide.
Cross-section: Labiolingually strongly flattened.
Carinae: Weakly carinate or rounded along the distal edge.
Serrations: Absent.
Basis for assignment: The holotype of Zanclodon laevis (Plieninger 1847; Schoch 2011b) is an incomplete maxilla with several teeth and an attached neural arch.
Variation: Schoch (2011b) reported some positional variation in the degree of the curvature of the tooth crowns.
Affinities: At present, Zanclodon laevis can only be identified as Archosauriformes incertae sedis (Schoch 2011b).
Type R3: partially serrated ziphodont
These teeth are generally very similar to slender morphs of type R1. The principal difference is the restriction of the serrated carinae to the apical half or two-thirds of the tooth crown (Fig. 4e).
Size: 10–20 mm crown height.
Tooth shape: Recurved, base moderately wide.
Cross-section: Anteroposteriorly oval at base, more rounded towards the tip.
Carinae: Slightly curved and not strictly parallel mesially and distally.
Serrations: Subquadrangular, 5 to 6 per mm of carina.
Basis for assignment: Only isolated teeth are known, leaving their affinities unresolved.
Variation: Length and alignment of carinae differ considerably in the known samples.
Affinities: Referral to Batrachotomus kupferzellensis cannot be entirely ruled out but is unlikely because all known cranial remains (Gower 1999; Gower and Schoch 2009) have only teeth of type R1.
Type R4: unicarinate ziphodont
This dental morphotype is rare but quite distinct from all other known reptilian teeth from the Lower Keuper. A few teeth of this type have been recovered from the Vellberg locality. They are distinguished by the presence of only a serrated mesial carina (Fig. 4f).
Size: 10–15 mm crown height.
Tooth shape: Gently recurved, base narrow; root unknown.
Carinae: Only along the mesial surface.
Serrations: Subquadrangular, 5–6 per mm.
Basis for assignment: Only isolated tooth crowns known.
Variation: Unknown.
Affinities: Unknown.
Type R5: slender lingually curved ziphodont
In the samples from Kupferzell and Vellberg, this morphotype ranks second in frequency among the larger reptilian teeth (Fig. 4g, h). Although distinctive in the irregularly aligned ridges on the distal flank of the crowns, it overlaps in size with type R6. Teeth may therefore be confused with the latter in cases of poor preservation. Clearly thecodont with long roots.
Size: 13–22 mm crown height.
Tooth shape: Gently curved lingually or almost straight, base very narrow.
Cross-section: Oval at base, lenticular at mid-crown level.
Carinae: Always bicarinate.
Serrations: Denticles present on both mesial and distal carinae; 6 per mm of carina regardless of crown height.
Surface: Smooth except for ridges, but lacking crenulations or grooves.
Basis for assignment: Only isolated teeth known.
Variation: Degree of lingual and posterior curvature as well as general outline and symmetry highly variable.
Diagnostic features: Prominent, parallel vertical ridges on lingual and labial crown surfaces. The ridges on the lingual flank are more clearly defined and aligned in parallel, whereas those of the labial flank are not strictly parallel, often discontinuous and variable in width. The enamel between the ridges is extremely finely wrinkled.
Wear: Common, usually extending basally from the tip along the lingual flank of the crown.
Affinities: Unknown. The resemblance to type R6 is noteworthy but may only reflect similar modes of feeding. A second possibility is that this tooth type represents an ontogenetically advanced stage of R6, but there is substantial size overlap between the two types, which does not support this hypothesis. In 2008, isolated teeth of type R5 were found in close proximity to elements of a small paracrocodylomorph (maxilla, braincase, vertebrae). These remains possibly belong to the same taxon, probably a pseudosuchian that is clearly distinct from Batrachotomus in features of the maxilla and vertebrae (Schoch 2011b). However, association of tooth type R5 with the aforementioned type of maxilla must remain conjectural.
Type R6: Jaxtasuchus salomoni
This tooth type is not as common as R5. It has been found in jaws of the doswelliid archosauriform Jaxtasuchus salomoni Schoch and Sues, 2014. The teeth have deeply thecodont implantation. There are no interdental plates.
Size: 6–12 mm crown height.
Tooth shape: Only gently recurved distally or straight, base narrow. Crown tall apicobasally (Fig. 4i, j).
Cross-section: Oval at base, lenticular at mid-crown level.
Carinae: Distinct but smooth mesial and distal carinae.
Serrations: None.
Surface: Covered by numerous closely set ridges (mid-crown level to tip).
Basis for assignment: One specimen of Jaxtasuchus (SMNS 91083) preserves both maxillae with several teeth still in their sockets and others fallen out but remaining associated with the cranial remains.
Variation: Within the jaw, there is substantial positional variation: alveolar outlines reveal that the teeth continuously increased in size from the anterior to the posterior up to a point at about two thirds the length of the maxilla and then the height of the tooth crowns decreased again more posteriorly.
Diagnostic features: Prominent, parallel vertical ridges extend along the labial and lingual surfaces of the crowns. Most of these extend to the tip of the crown, but they become more faint and numerous apically. On some teeth, shorter ridges extend from the base of the crown between the longer ridges for only part of the height of the crown.
Affinities: Jaxtasuchus was an 80–120-cm-long, possibly semi-aquatic doswelliid archosauriform (Schoch and Sues 2014). Its distinctive rectangular dorsal osteoderms are common in Lower Keuper strata and bear a central eminence (tubercle) and ornamentation composed of pits and polygonal ridges.
Type R7: archosauriform with keeled armour plates
Based on a single disarticulated skeleton (SMNS 96936), this type of tooth has been identified as belonging to an archosauriform that attained a total length of 100 to 120 cm. Despite similar size, this taxon clearly differs from Jaxtasuchus in many skeletal features (cranial elements, vertebrae and limb proportions). Its most distinctive feature is the structure of the osteoderms, which are tear-drop-shaped and strongly keeled, with an entirely smooth dorsal surface. These elements were briefly described and figured by Schoch (2011b), prior to the discovery of a more complete specimen that includes a maxilla with deep alveoli.
Size: 5–6 mm crown height.
Tooth shape: Clearly recurved, base bulbous, crown stout but pointed, relatively low compared to tooth crowns of Jaxtasuchus or Batrachotomus.
Cross-section: Oval at base.
Carinae: Mesial and distal carinae near the tip, but two-thirds of the crown lack any keels.
Serrations: None.
Basis for assignment: One partial skeleton includes a maxilla with two teeth.
Variation: Unknown.
Diagnostic features: Parallel ridges on labial aspect of crown prominent, differing from those of other taxa in that they are separated by grooves rather than emplaced on a smooth enamel surface.
Affinities: The sigmoidal curvature of and presence of a fourth trochanter on the femur suggest archosauriform affinities, whereas the humerus and scapula resemble those of basal pseudosuchians. Compared to Jaxtasuchus, this taxon had a more robust skeleton and proportionately larger skull.
Type R8: archosauriform: tiny recurved teeth
This type is probably common but is not often recognised due to its minute size (Figs. 4l and 5d). It has been identified in articulated specimens of a small archosauriform (15–25 cm body length). This taxon has highly distinctive osteoderms with (1) an oval to rhomboidal outline, (2) serrated lateral and medial margins, (3) a parasagittal ridge and (4) pitted ornamentation. A second type or osteoderm, probably from the ventral region, comprises rectangular plates with broad anterior bars for the overlap by the preceding plate. The skull of this small archosauriform was lightly built and relatively low and had a long snout. The teeth are small and numerous in relation to jaw size. Tooth implantation is thecodont, with roots longer than crowns and well-developed interdental plates on the lingual side of the maxilla.

Small reptile tooth types. a, b Type R11 (a maxilla, SMNS 91547; b dentary; SMNS 91111). c Type R12, maxilla (SMNS 91376). d Type R8, dentary (SMNS 91118). e, f Type R13, stem-turtle Pappochelys rosinae (e maxilla, SMNS 91431; f dentary, SMNS 92066). g, h Type R14, lepidosauromorph (g maxilla, SMNS 91319; h dentary, SMNS 92079). i, j Type R15, choristodere-like diapsid (i maxilla, j dentary, both SMNS 91552). k Type R16, maxilla (SMNS 91590). l, m Rhynchocephalians: l type R17, dentary (SMNS 92072), m type R18, maxilla (SMNS 91313). n Type R19, dentary (SMNS 91379). o Type R22, maxilla (SMNS 92102). p Type R20, dentary (SMNS 96944)
Size: 2.5–3.5 mm crown height.
Tooth shape: Apex of crown markedly recurved.
Cross-section: Round to oval at base, and oval at mid-crown level.
Carinae: Some carinate teeth in the premaxilla (both mesial and distal sides) and mandible (distal side). When present, the carinae are confined to the region of the apex.
Serrations: None.
Basis for assignment: Teeth of this type are present in the jaws of several specimens with the diagnostic osteoderms (SMNS 90505, 91118, 91121, 91312).
Variation: Both carinate and non-carinate teeth are present in the same maxilla, indicating positional variation. In addition, even neighbouring teeth vary with respect to curvature, crown size and cross-sectional outline.
Diagnostic features: Crowns always recurved, especially towards the apex. Fine parallel stripes along the labial surface, following the general curvature. The roots are long.
Affinities: A basal archosauriform possibly related to certain other osteoderm-bearing taxa, such as doswelliids and Vancleavea. Work on the anatomy and relationships of this archosauriform is in progress by the senior author and Randall Irmis (University of Utah).
Type R9: archosauriform: teeth with pointed serrations parallel to apex
This tooth type is highly variable with respect to position and size. Tooth implantation is thecodont, with long roots. There are no interdental plates.
Size: 1.5–7 mm crown height. Teeth at mid-level in each premaxilla and maxilla are tallest, those at anterior and posterior ends of the jaw elements shortest.
Tooth shape: Distinctly heterodont, ranging from asymmetrically recurved to symmetrically lancet-shaped.
Cross-section: Labiolingually compressed, oval at base, lenticular at mid-level.
Carinae: Always bicarinate, with mesial and distal carinae.
Serrations: Always present, pointed, proportionately large, with serrations aligned parallel or close to the apicobasal axis of the crown. Remarkably similar to tooth crowns in basal sauropodomorph dinosaurs (e.g. Plateosaurus).
Basis for assignment: Two partial, disarticulated skeletons of an archosauriform (SMNS 91343) are known in which these teeth are associated with premaxillae, maxillae and dentaries.
Variation: Pronounced positional variation in tooth size and structure: premaxillary teeth conical; maxillary teeth largest teeth at mid-length of maxilla, with posteriormost teeth on maxilla smallest.
Diagnostic features: The shape of the tooth crowns and the orientation of the serrations are unique among the Lower Keuper tetrapods. Similar teeth are known in extant iguanid lizards (Iguana iguana, Montanucci 1968) and sauropodomorphs (Plateosaurus, Huene 1907–1908).
Affinities: A basal archosauriform lacking osteoderms, a relatively plesiomorphic femur (only gently S-shaped shaft, no fourth trochanter) and a proportionately large head. The robust jaws and large, serrated teeth are diagnostic for this taxon, which is currently under study by the senior author, Hans-Dieter Sues, and Julia Desojo.
R10: large chisel-shaped teeth
This tooth type is known only from a single jaw fragment (SMNS 91427) but is unique among tetrapods from the Triassic of Germany.
Size: 6–8 mm crown height.
Tooth shape: Tall crown, chisel-shaped, with parallel and straight mesial and distal margins.
Cross-section: Oval at base.
Carinae: Sharp carinae along the mesial edge and apex, but not distally.
Serrations: None.
Basis for assignment: Teeth preserved in jaw fragment (dentary?).
Variation: Positional variation minor.
Diagnostic features: The chisel-shaped crowns and orientation of carinae make this type very distinctive. The teeth are densely set in the jaw, lack labyrinthine infolding or ridges, but have pronounced carinae along the mesial and dorsal edge.
Affinities: Unknown. Probably an amniote because there are no traces of labyrinthodont infolding, but even this feature does not necessarily rule out more basal tetrapods. The tooth shape resembles that in the Early Permian anthracosaur Archeria (Holmes 1989) and the Early Carboniferous lepospondyl Adelogyrinus (Andrews and Carroll 1991).
With few exceptions, the following types of teeth are too small to be recovered as isolated teeth by quarrying. They are usually found only in situ in jaws but are also recovered by screen-washing of fossiliferous matrix.
R11: strong conical teeth with striae and lingual keel
This type is known from six disarticulated skeletons that include a dentary and associated postcranial bones (SMNS 91111, 91547).
Size: 0.8–1 mm crown height.
Tooth shape: Tall crown, tip recurved (or lingually curved), 3–5 striae on the lingual side, intercalating with a short lingual keel near tip.
Cross-section: Oval at base and mid-crown level.
Carinae: A single lingual one.
Serrations: None.
Basis for assignment: Jaws associated with numerous postcranial bones (ilia, vertebrae, limbs). The ilium has a slender dorsal process similar to that in lepidosauromorphs.
Variation: Positional variation considerable, with anterior maxilla teeth tallest and most strongly recurved, the posterior ones being shortest and more bulbous; dentary teeth intermediate in size and degree of curvature. However, lingual striae and keels are throughout present.
Implantation: Pleurodont.
Diagnostic features: The striated and lingually keeled tips, pleurodonty, and close packing of teeth are highly distinctive features.
Affinities: Small lepidosauromorph diapsid (10–15 cm snout-vent length).
R12: robust recurved teeth
Known from a partial skull that includes a maxilla (SMNS 91376) (Fig. 5c).
Size: 1.2–1.5 mm crown length.
Tooth shape: Very robust base, some tooth crowns slightly recurved, with pointed tip.
Implantation: Probably thecodont.
Cross-section: Oval at base, labiolingually compressed at mid-crown level.
Carinae: Apparently none.
Serrations: None.
Basis for assignment: Maxilla associated with vertebrae.
Variation: Size of teeth decreasing posteriorly.
Diagnostic features: The markedly recurved, thick-based tooth crowns are roughly triangular in outline in side view and widely spaced, with the maxilla accommodating up to 12 teeth.
Affinities: Probably an archosauriform, based on the presence of an antorbital fenestra.
R13: Pappochelys: straight peg-like teeth in tall maxilla
The stem-turtle Pappochelys rosinae is known from a suite of partial skeletons (SMNS 90013, 91431, 92063, 92066) that were described by Schoch and Sues (2015, 2017). The premaxilla bears four teeth, the maxilla up to 17 (Fig. 5e), and the dentary at least 29 (Fig. 5f).
Size: 0.6–0.9 mm crown height.
Tooth shape: Robust base, tall and straight crown, pointed tip.
Implantation: Apparently thecodont with teeth sitting in shallow sockets.
Cross-section: Oval at base and at mid-crown level.
Carinae: None.
Serrations: None.
Variation: Teeth becoming slightly larger posteriorly.
Diagnostic features: In both upper and lower jaws, the more posterior teeth are smaller and somewhat more robust.
R14: cylindrical peg-shaped teeth
Several sets of disarticulated skeletal remains of a distinctive diapsid reptile preserve teeth of this type. This taxon has a tall maxilla and a relatively short antorbital region of the skull, in contrast with the taxon represented by tooth type R13. The outline of the maxilla is similar to that of the taxon with R13, but the teeth are more robust and heterodonty is less pronounced (Fig. 5g, h). Known from six partial, disarticulated skeletons (SMNS 91110, 91270, 91319, 91374, 91376, 91399) and numerous isolated dentaries.
Size: 0.4–0.7 mm crown height.
Tooth shape: Cylindrical, peg-shaped; crowns straight and slender crowns, with tapering to rounded tips.
Implantation: Pleurodont.
Cross-section: Oval at base and at mid-crown level.
Carinae: None.
Serrations: None.
Variation: The size of teeth becomes smaller and they have rounded tips posteriorly.
Diagnostic features: The long maxilla and dentary house numerous small teeth that are clearly heterodont: peg-shaped teeth markedly taller anteriorly but very short near the posterior end of the maxilla.
Affinities: Diapsid; maxillary outline resembling that of R13.
R15: choristoderan-like diapsid: strongly recurved and pointed teeth
This common taxon is represented by isolated jaw bones and disarticulated skeletons (SMNS 91347, 91552) (Fig. 5i, j).
Size: 0.9–1.2 mm crown height.
Tooth shape: Distinctly recurved, especially near tip, some with faint labyrinthodont infolding at base.
Implantation: Thecodont.
Cross-section: Oval at base and at mid-crown level.
Carinae: None.
Serrations: None.
Variation: Anteriormost teeth are longest with tips most clearly recurved.
Diagnostic features: The markedly recurved, thick-based teeth are roughly triangular in side view and widely spaced, with the maxilla accommodating up to 14 teeth. In general, this tooth type closely resembles that of Prolacerta in shape and size (Modesto and Sues 2004), but the placement is less regular. The dentary is short, posteriorly framing a large meckelian fenestra.
Affinities: Based on derived features in the ilium, humerus and femur, this taxon is a diapsid that resembles choristoderans in various features. The skull was apparently lightly built with gracile, strut-like elements framing large temporal fenestrae.
R16: tiny slender and recurved teeth
This tooth type is so far known only from a single, 13-mm-long disarticulated skull (SMNS 91590) (Fig. 5k).
Size: 0.4–0.6 mm crown height.
Tooth shape: Slender, markedly recurved, closely set.
Implantation: Unknown.
Cross-section: Oval at base, labiolingually compressed at mid-crown level.
Carinae: Apparently none.
Serrations: None.
Variation: Positional variation minimal.
Diagnostic features: At the present stage, the tiny size in combination with the slender recurved shape and close packing are sufficient to distinguish this type from all others.
Affinities: Diapsid reptile, possibly lepidosauromorph.
R17: rhynchocephalian: triangular acrodont teeth
This type of tooth is present in jaws of a tiny reptile (SMNS 91060, 91321, 91322). A dentary is only about 12 mm long (Fig. 5l).
Size: 0.3–1.0 mm crown height.
Tooth shape: Triangular, robust, lingually and labially bulbous, pointed tip with faint striations parallel to apicobasal axis of crown.
Implantation: Acrodont.
Cross-section: Oval at base, labiolingually compressed at mid-crown level.
Carinae: None.
Serrations: None.
Basis for assignment: Only dentary teeth are currently known.
Diagnostic features: The teeth are all similar in shape but increase in size posteriorly, with the last tooth by far the largest.
Affinities: Rhynchocephalian that closely resembles Diphydontosaurus from the Upper Triassic of England. First reported by Jones et al. (2013).
R18: rhynchocephalian: heterodont with acrodont implantation
Based on disarticulated skull and postcranial remains, this tooth type appears to represent a taxon distinct from that with teeth of type R17 (SMNS 91061, 91313, 91323) (Fig. 5m). The dentary is about 20 mm long and substantially deeper than that of the taxon with tooth type R17, but the teeth are not larger than in the latter.
Size: 0.5–1.0 mm crown height.
Tooth shape: Distinctly heterodont; from slender recurved crowns (anterior portion of maxilla) to bulbous triangular (posterior portion of dentary and maxilla). Some teeth in the posterior part of the maxilla have a markedly thickened base.
Implantation: The teeth on the dentary and the more posterior portion of the maxilla are acrodont, whereas the teeth on the premaxilla and anterior portion of the maxilla are pleurodont.
Cross-section: Oval at base, slightly labiolingually compressed at mid-crown level.
Carinae: None.
Serrations: None.
Basis for assignment: Two partial skeletons preserve jaws with teeth, skull elements and trunk vertebrae.
Diagnostic features: The pronounced heterodonty is diagnostic, especially the grading from slender recurved to bulbous triangular.
Affinities: At least the posterior teeth are acrodont and similar to those of rhynchocephalians. Together with type R17, these teeth were first reported by Jones et al. (2013). More complete material is needed before this taxon can be diagnosed; further studies should clarify whether R17 and R18 merely represent different ontogenetic stages belonging to the same taxon.
R19: stout conical with pointed cusp
A single isolated dentary preserves a series of distinctive, markedly bulbous teeth (SMNS 91379) (Fig. 5n).
Size: 1.0–1.9 mm crown height.
Tooth shape: Stout conical, slightly recurved, with single pointed cusp.
Implantation: Unclear.
Cross-section: Round at base, oval at mid-crown level.
Carinae: None.
Serrations: None.
Basis for assignment: None.
Diagnostic features: The teeth are throughout similar in their stoutness, the presence of a single pointed cusp, and marked zones of preserved colouration (light brown at base, dark at mid-crown level and reddish-brown at apex). The surface of the teeth is smooth.
Affinities: Tetrapod of uncertain position.
R20: stout conical with rounded cusp
A further single jaw fragment (SMNS 91380) has bulbous teeth that differ from those of tooth type R19 in the size and structure of the cusps (Fig. 5p).
Size: 0.7–1.0 mm crown length.
Tooth shape: Stout conical with single rounded or slightly edged cusp.
Implantation: Unclear, probably in sockets on top of jaw.
Cross-section: Round at base, oval at mid-crown level.
Carinae: None.
Serrations: None.
Basis for assignment: None.
Diagnostic features: The teeth form straight cones and appear to be set in shallow sockets. One tooth has a broad base, with a concave central part, suggesting a bifurcating root. The larger teeth have parallel striations near the apex. Preserved colour is a uniform light brown.
Affinities: Tetrapod of uncertain position.
R21: Colognathus: bulbous and pointed teeth
Three fragments of lower jaws with remarkably large teeth have this type of tooth (SMNS 91315, 91428, 91429) (Fig. 6a, b) (Schoch 2011a; Sues and Schoch 2013).

Small reptile tooth types. a, b Type R21, Colognathus sp. (a lateral view, SMNS 91315; b two associated teeth; SMNS 91429). c, d Type R26, procolophonid teeth from labial (c) and lateral (d) (SMNS 94999). e–i Type R23, tricuspid teeth (e tooth mesial view, SMNS 91387; f dentary with three teeth in situ, SMNS 91385; g tooth in occlusal view, SMNS 91387; h, i teeth in mesial view; h SMNS 91388; i SMNS 91389)
Size: 3–3.5 mm crown length.
Tooth shape: Stout conical crown with a single pointed cusp, triangular in labial view, lingual and labial flanks bulbous.
Implantation: Acrodont.
Cross-section: Rounded at base, labiolingually oval at mid-crown level.
Carinae: None.
Serrations: None.
Basis for assignment: None.
Diagnostic features: The cusps bear fine, parallel ridges (usually five or six on the labial aspect of the crown) near the apex. In all three known mandible fragments, only two very large teeth are present at the posterior end of the dentary. There may have been one or two additional teeth anterior to these, but the dentary is very thin there and they could not have been as large as the two posterior ones. The tooth crowns have distinct zones of colouration: dark at base, light brown at mid-crown level.
Affinities: The jaws and teeth closely resemble those of the enigmatic tetrapod Colognathus obscurus from the Upper Triassic of North America and have been described by Sues and Schoch (2013).
R22: small peg-like teeth in tall maxilla
Based on a single maxilla (15 mm long) with 11 teeth (SMNS 91430; Fig. 5o).
Size: 0.5–0.8 mm crown length.
Tooth shape: Peg-like with pointed crown; the anteriormost teeth point posteriorly, whereas those at mid-level are straight. The posteriormost tooth is much more robust with broad base.
Implantation: Unknown.
Cross-section: Probably round.
Carinae: None.
Serrations: None.
Basis for assignment: None.
Diagnostic features: Teeth relatively small, only one fourth the height of the maxilla. Prominent anterior premaxillary ramus, external naris framed by anterodorsally extending alar process.
Affinities: Possibly diapsid.
R23: tricuspid teeth
A wide range of tricuspid (triconodont) teeth are found in Lower Keuper sediments, especially mudstones. These are consistent with the teeth of juveniles of the archosauromorph Tanystropheus longobardicus (Peyer 1931; Wild 1973; Nosotti 2007). Usually these teeth are found isolated. To date, only one tiny dentary fragment bearing such teeth has been discovered (Fig. 6f).
Size: 0.5–3.5 mm crown length.
Tooth shape: Stout conical with three pointed cusps, consisting of a large central cusp and two small accessory ones mesially and distally.
Implantation: Thecodont with relatively short root and thin bony lingual wall of the alveolus.
Cross-section: Elongate oval at base and at mid-crown level.
Carinae: None.
Serrations: None.
Basis for assignment: Only a single dentary with three tricuspid teeth in place, separated by wide gaps (SMNS 91385).
Diagnostic features: The small accessory cusps are usually tiny and located at mid-level or slightly crownwards of mid-level. In apical view, the base is elongate oval (mesiodistally) and the three cusps are aligned in a single mesiodistal axis. The labial and lingual flanks are bulbous and may bear parallel crenulations in large teeth. As noted by Wild (1973), wear surfaces are usually perpendicular to the long axis of the crown, affecting only the apex, which is often rounded rather than pointed.
Affinities: Diagnostic cervical vertebrae of Tanystropheus occur in the same horizons as the tricuspid teeth. Furthermore, the small dentary from Vellberg preserves the anterior end, which closely resembles that in juvenile Tanystropheus longobardicus (Wild 1973; Nosotti 2007). Based on juvenile specimens of Tanystropheus longobardicus, the small tricuspid teeth from the Lower Keuper represent individuals with total lengths ranging from 60 to 100 cm. Yet, it is by no means certain that all tricuspid teeth from the Lower Keuper belong to tanystropheids as similar teeth are present in basal pterosaurs and some cynodont synapsids.
R24: Tanystropheus
Widely known from Muschelkalk deposits, the tall conical teeth of Tanystropheus spp. are also found in brackish or shallow marine deposits of the Lower Keuper. At the Kupferzell locality, a few teeth possibly referable to Tanystropheus were also discovered in strata that yielded diverse temnospondyls.
Size: 25–35 mm crown height.
Tooth shape: Slender conical, markedly curved lingually, usually with lingually worn tip (Fig. 4q).
Implantation: Thecodont with relatively long root in alveoli with a thin lingual wall.
Cross-section: Round at base and at mid-crown level.
Carinae: None.
Serrations: None.
Basis for assignment: Tanystropheus longobardicus is known from numerous articulated partial to complete skeletons from the Anisian-Ladinian Grenzbitumenzone of Monte San Giorgio, Switzerland (Wild 1973), and Besano, Italy (Nosotti 2007).
Diagnostic features: Distinct, often rounded ridges extend apically on all surfaces of the crown from slightly below mid-level. These ridges are not very tall and vary in distance as well as height. They also do not extend strictly parallel and may bifurcate or merge along the apicobasal axis of the crown. The ridges resemble those on teeth of type R5, although that type has a very different cross-section. The enamel between the ridges is very finely and irregularly corrugated. Smaller, probably juvenile tooth crowns are more flattened laterally, their base being enlarged anteriorly and posteriorly to form incipient lateral cusps (Dorka 2002: morphotype 13).
R25: Nothosaurus
Nothosaurus spp. have very distinctive teeth, which resemble those of Tanystropheus at first glance. Closer inspection reveals the equidistant arrangement and high regularity of ridges on these teeth (Fig. 4p).
Size: 20–50 mm crown height (probably representing different species of Nothosaurus).
Tooth shape: Slender, conical, strongly curved lingually, with pointed tip.
Implantation: Thecodont with relatively short root.
Cross-section: Round throughout the height of the crown.
Carinae: None.
Serrations: None.
Basis for assignment: Known from many dissociated and articulated skeletal remains, especially from the Muschelkalk of southern Germany (Rieppel 2000).
Diagnostic features: Apicobasal ridges tall, sharp-edged, regular in length and distance from each other, extending from the base of the crown almost to the tip.
R26: Procolophonidae
Size: 14–18 mm crown height (Fig. 6c, d).
Tooth shape: Crown labiolingually broad, tapering towards apex in labial/lingual view.
Implantation: Subthecodont.
Cross-section: More or less elliptical.
Carinae: None.
Serrations: None.
Basis for assignment: A partial dentary with teeth (SMNS 94999).
Diagnostic features: Strong base, rounded cusp, no striations or carinae.
Synapsid teeth
S1: traversodontid cynodont
A single lower postcanine tooth from a desiccation crack filling at Michelbach an der Bilz (SMNS 51962) represents a traversodontid cynodont, Nanogomphodon wildi, and was described by Hopson and Sues (2006).
Tetrapod diversity and palaeoecology
The diversity of tetrapod teeth is relatively high in the Lower Keuper, which is not surprising given the wide range of depositional environments represented by this mixed shallow marine to terrestrial sequence (Beutler et al. 1999; Etzold and Schweizer 2005; Nitsch 2015). In addition, the position of southern Germany at low palaeolatitude during the Middle Triassic suggests subtropical conditions, which is consistent with the rich vegetation recorded from Lower Keuper strata (Heunisch 1999, 2015; Kelber 2015). Recent excavations have added substantially to our knowledge of tetrapod diversity, now including numerous small diapsid reptiles. Among the isolated teeth, the largest ones of which represent non-archosaurian archosauriforms and pseudosuchians, whereas the small and tiny teeth belong to a wide range of taxa including archosauriforms, tanystropheids, a choristodere-like reptile, a stem-turtle, rhynchocephalians, basal lepidosauromorphs, and possibly additional diapsid groups. It is noteworthy that cynodonts are virtually absent with the exception of a single isolated lower postcanine tooth of a traversodontid cynodont (Hopson and Sues 2006). This is in stark contrast to stratigraphically slightly younger tetrapod assemblages from the Upper Triassic of Argentina and Brazil (Bonaparte 1978; Martínez et al. 2013) or the Newark Supergroup of eastern North America (Sues and Fraser 2010). It also differs from the Middle Triassic assemblages reported from the southern Urals, Russia (Shishkin et al. 2000). Most surprisingly, remains of avemetatarsalian archosaurs and rhynchosaurs appear to be entirely absent in the Lower Keuper localities studied to date.
General palaeoecological features and problems
The palaeoenvironments represented by the Lower Keuper deposits are fairly well understood (Beutler et al. 1999; Nitsch 2015) and range from shallow marine (dolomites, mudstones) to brackish-lacustrine-paludal (dolomites, marlstones, mudstones) and to deltaic or fluvial depositional settings (sandstones, siltstones, coal seams). Much less common than peat-forming lakes or lagoons were freshwater lakes. They are best exemplified by a locally restricted mudstone horizon (bed E6 of Schoch 2002) at the Vellberg locality. This stratum contains a diverse autochthonous assemblage of lacustrine vertebrates (elasmobranchs, actinopterygians, dipnoans, coelacanths, temnospondyls, choristodere-like diapsids) and an allochthonous assemblage of mainly terrestrial tetrapods (chroniosuchians, paracrocodylomorph pseudosuchians, non-archosaurian archosauriforms, various lepidosauromorph or related diapsids, stem-turtle, procolophonoids). The reptilian dental morphotypes described here primarily represent the latter assemblage.
Nothosaurus and Tanystropheus form the largest and most common marine and brackish-water inhabitants, possibly in lagoonal environments, whereas pseudosuchians (mainly Batrachotomus) are abundant in terrestrial settings. The most common freshwater tetrapods are temnospondyls, particularly Mastodonsaurus and Gerrothorax, and the abundance of large isolated fangs of the former (types A1a–b) is noteworthy. Mastodonsaurus was present almost everywhere, forming the apex predator in freshwater settings; very rarely does it co-occur with Nothosaurus, however, suggesting different habitat preferences for these two taxa.
Smaller lacustrine taxa comprise diapsid reptiles with snout-vent lengths ranging from 15 to 40 cm, especially a 25–40-cm-long choristodere-like form (R15). Furthermore, two armoured archosauriforms are especially common in the Vellberg lake deposit: a 20–30-cm-long, long-skulled form (R8) and the 80–120-cm-long doswelliid Jaxtasuchus (R6, Schoch and Sues 2014). Finally, numerous remains of the stem-turtle Pappochelys (R13) have been found in the lacustrine deposits at Vellberg, suggesting this taxon may have lived close to or along lakeshores (Schoch and Sues 2015, 2017). The majority of small reptilian remains have tooth types resembling those found among extant terrestrial reptiles, notably forms feeding on invertebrates. These finds contribute substantially to our knowledge of terrestrial palaeoecosystems, because so far the Vellberg deposit is the first to yield such tetrapod remains in greater quantity and better quality of preservation, to be elucidated in the next section.
The palaeoecology of the Lower Keuper vertebrate communities is still poorly understood. Some clarification may be expected from the forthcoming studies of tetrapod taxa that are now represented by extensive skeletal remains and probably will reveal information concerning the mode of life of these animals. Some dental morphotypes described here may turn out to be merely parts of distinctly heterodont dentitions once more complete cranial material becomes available. Thus, it is generally inadvisable to base new taxa solely on isolated teeth at this point in time. In many cases, a single tooth cannot even be identified at a lower taxonomic level.
Palaeoecological significance of tooth types
The only palaeoecological considerations possible with the given set of data are comparisons of Lower Keuper tetrapod teeth with analogous tooth morphotypes in other extinct and extant taxa. Furthermore, physical parameters of tooth crowns can be broadly related to broader dietary categories (Lucas 2004). Naturally, such comparisons can only be at a rather coarse scale and focus on a few readily apparent functional properties of the teeth. To gain more reliable information, multiple articulated skeletons with gut contents are required. However, some studies have even shown that gut contents and tooth form are not always directly correlated (e.g. Munk and Sues 1993). Furthermore, research of feeding behaviour in extant lizards has shown that similar teeth may be used for different diets. For instance, iguanas predominantly subsist on plants but will occasionally also consume animal protein (Hotton 1955; Barrett 2000). However, most tooth morphotypes in extant tetrapods indicate the principal components of the diet. Below we will give a brief discussion of how the tetrapod teeth of the Lower Keuper compare to those of extant taxa and what may be inferred from dental resemblances.
With the exception of tooth types R26 and S1, representing a traversodontid cynodont, the known tooth morphotypes from the Lower Keuper show little evidence for oral food processing. Teeth with conical crowns are suitable for impaling and killing prey, much as those in extant crocodylians. The interdigitating teeth with slender, conical and often procumbent crowns and pointed tips of Nothosaurus closely resemble those of many plesiosaurs, which have been interpreted as puncturing teeth suitable for seizing slippery prey such as cephalopods and fish (Massare 1987; Rieppel 2002). Ziphodont teeth with serrated carinae are mechanically suited for slicing through muscle fibre (Farlow et al. 1991; Abler 1992; D’Amore and Blumenschine 2009, 2012) and are generally considered indicative of carnivorous habits. Tooth type R9 closely resembles the teeth of extant herbivorous iguanid lizards and basal sauropodomorph dinosaurs, which probably also fed primarily on plants.
Temnospondyls: Although most known temnospondyls were aquatic and are considered primarily piscivorous, only a few specimens preserve gut contents (Schoch 2009, 2014). For the entire Lower Keuper, such information has not yet been reported, despite some promising accumulations of articulated skeletal material of temnospondyls discovered in the last decade (Callistomordax, Gerrothorax, Plagiosuchus, Trematolestes). Common attributes of temnospondyl teeth are their conical shape, the presence of plicidentine, the close spacing of marginal teeth and the presence of fangs in the larger-sized capitosaurs and trematosaurs. There are no close analogues to the latter teeth among present-day tetrapods. Nevertheless, these features suggest that all known temnospondyl taxa were carnivorous, presumably mainly feeding on fish. Divergent tooth structures and sizes likely reflect different prey preferences or modes of prey acquisition. The high diversity of aquatic tetrapods, in addition to numerous fish taxa, suggests that the larger temnospondyls may not have confined themselves to piscivory. In fact, a crushed skull of a Mastodonsaurus with bite marks suggests that this individual fell victim to a much larger conspecific (Schoch and Seegis 2016).
Based on its body size and the structure of its jaws and dentition, Mastodonsaurus (3–5 m long) was certainly able to feed on most vertebrates in the 1–3-m length range (Schoch 2015a). Its enormous fangs indicate that large or struggling prey could be quickly immobilised or killed (Schoch 1999). The archosauriform skeletons are usually only partially preserved, and predation or scavenging by aquatic carnivores may account at least in part for this taphonomic phenomenon. Smaller diapsids are typically represented by oval or longitudinal accumulations of closely packed, disassociated bones, which are likely regurgitates or disintegrated coprolites (Schoch and Seegis 2016). The same pattern is found among juvenile dipnoans, which are common in these freshwater deposits (Hagdorn et al. 2015).
Mastodonsaurus and Kupferzellia probably preyed on animals of different sizes, but the close similarity between the dentitions indicates comparable modes of feeding (Schoch and Milner 2000). By contrast, Trematolestes and Callistomordax share carinate fangs and marginal teeth, which were probably used in a different way than those of capitosaurs. Both taxa were probably able swimmers and hunted prey actively, but the nature of the prey remains unknown. Plagiosaurids have rather small teeth, which are confined to the anterior margins of the jaws, and lack fangs altogether. They may have been ambush predators feeding on small fish and invertebrates. The smallest plagiosaurid, Gerrothorax, was a pioneer taxon present across a wide range of habitats that often had few other tetrapods (Schoch and Witzmann 2012; Sanchez and Schoch 2013).
Bulbous teeth: Tooth types R19–R21 are difficult to attribute to a particular tetrapod clade, but probably represent otherwise still undocumented amniotes, based on examination of tooth surface and jaw structure. R21 is referable to the enigmatic tetrapod Colognathus (Heckert et al. 2012; Sues and Schoch 2013) based on tooth shape. Comparable teeth are present in various extant scincid and teiid lizard species that specialise on snails and other hard-shelled food items that require crushing (Estes and Williams 1984) The much less robust teeth of R19 and R20 resemble those of the xantusiid lizard Xantusia, which feeds on termites and spiders (Zweifel and Lowe 1966).
Acrodont teeth: Tooth types R17 and R18 represent rhynchocephalians (Jones et al. 2013). The acrodont teeth of the extant tuatara (Sphenodon punctatus) are thought to exert greater biting force than the pleurodont dentition of most squamates (Borsuk-Białynicka 1996; Jones 2008). The tuatara feeds on insects, especially large grasshoppers and beetles, and spiders. Adult tuataras will occasionally prey on juvenile seabirds (Cree 2014).
Ziphodont recurved teeth: Tooth types R1 and R3–R4 are ziphodont, with distally recurved and labiolingually flattened tooth crowns, which bear serrated carinae at least on one side. Ziphodonty is today restricted to some varanid lizards among extant reptiles but was widespread among Mesozoic archosauriform reptiles. In the Lower Keuper, the best-known ziphodont taxon is Batrachotomus, which, at the Kupferzell site, left diagnostic tooth marks on limb bones and ribs of Mastodonsaurus (Schoch and Wild 1999). This is also supported by finds of large temnospondyl ribs and even fangs at the Vellberg locality, which preserve cut marks generated by serrated teeth matching the size and structure of ziphodont tooth type R1 (Schoch and Seegis 2016: fig. 10c). A feature shared by tooth types R1–R4 is the distal curvature of the tooth crown (although some teeth in the holotypic maxilla of Zanclodon laevis are straight), which closely resembles the condition in Varanus komodoensis (D’Amore and Blumenschine 2009, 2012). All extant varanids with labiolingually compressed and recurved teeth are large-sized carnivores that prey on mammals, and V. komodoensis has been reported to bring down even water buffaloes (Auffenberg 1981). The fact that all large tetrapod teeth (other than labyrinthodont and sauropterygian types) are ziphodont indicates that archosauriforms were the dominant terrestrial predators in the Lower Keuper.
However, the frequency of types R1 to R5 is very different, suggesting that the preferred habitat of Batrachotomus is more frequently preserved or collected than that of Zanclodon. Batrachotomus is known from lacustrine and brackish lagoonal deposits (Vellberg, Kupferzell, Ilshofen), whereas the only known specimen of Zanclodon laevis was recovered from a coal seam (Gaildorf) (Schoch 2015b). As argued for metriorhynchid crocodyliforms (de Andrade et al. 2010), the different distributional patterns of denticles likely reflect different feeding preferences or modes of prey processing in the Lower Keuper ziphodonts, with Batrachotomus forming the apex terrestrial predator.
Lingually curved ziphodont teeth: One of the more peculiar dentitions is the enigmatic tooth type R5, which is still known only from isolated teeth. Although it resembles Jurassic thalattosuchian crocodyliforms (e.g. Steneosaurus bollensis), its combination of serrated carinae, longitudinal ridges, and the pronounced lingual curvature of the crown is distinctive. This curvature could indicate piscivory by analogy with crocodyliforms. However, this conclusion must remain tenuous because thalattosuchians either lack identifiable food residues in their stomach contents (Steneosaurus; Mateer 1974) or have mixed diets composed of cephalopods, crustaceans and fish (Metriorhynchus: Martill 1986; Cricosaurus: Günter Schweigert, pers. comm.).
Potential piscivores: Nothosaurus has long been considered a fish eater, able to conduct rapid snapping bites with its characteristic ‘pincer’ jaws (Rieppel 2002). Both the functional properties of the nothosaurid dentition and its close similarity to those of the extant crocodylians Tomistoma schlegelii and Gavialis gangeticus support this inference. The teeth of the doswelliid Jaxtasuchus (R6) differ considerably from all dental morphotypes in the Lower Keuper but share with R5 the presence of marked ridges and distinct mesial and distal carinae. The pronounced ridges are similar to those on teeth of adult tanystropheids, but there are no extant analogues known to us. As for tanystropheids, Wild (1973) reported hooklets of cephalopod arms from gut contents of adult Tanystropheus longobardicus, and it is likely that this archosauromorph, at least as an adult, preyed on a range of smaller marine animals (Fig. 7).

Palaeoecological significance of temnospondyl and reptile tooth morphotypes from the Lower Keuper as analysed in this paper. Hypothesised prey preferences inferred from extant taxa with similar dentition. The large faction of putative fish eaters and the paucity of diagnostic herbivores form the most striking features of this assemblage
It is possible that some of the other presumed carnivores from the Lower Keuper also preyed on fish. Most extant crocodylians subsist on fish as well as tetrapods and have stout conical teeth without carinae and serrations.
Teeth of choristoderan-like diapsid: Teeth of the type R15 are associated with skeletons of an aquatic choristodere-like diapsid. Except for the absence of venom grooves, they resemble the teeth of the gila monster (Heloderma suspectum), which preys on arthropods and small tetrapods. A relatively large skeleton of the Vellberg taxon (SMNS 90696) preserves numerous scales of a small actinopterygian (Dipteronotus sp.) between the ribs and gastralia of its trunk region.
Teeth of Pappochelys: The stem-turtle Pappochelys has closely set, peg-like teeth similar to those of the collared lizard (Crotaphytus), which feeds on insects and small vertebrates.
Other morphotypes of recurved teeth: The smallest archosauriform taxon in the Lower Keuper has recurved non-ziphodont teeth, which have carinae only in the apical portion of the crown. Similar teeth are known in various extant lizards (Lialis, Xenopeltis, Celestus, Shinisaurus; e.g. Mahler and Kearney 2006) as well as in other archosauriforms (Proterosuchus) and non-archosauriform archosauromorphs (Prolacerta) (Welman 1998; Modesto and Sues 2004). The extant analogues prey on other lizards (Lialis burtonensis), amphibians (Xenopeltis unicolour) and even small mammals (Celestus enneagrammus). The more robust teeth of types R11–R13 appear to be variants of such dentitions and may have been used for similar purposes.
Cylindrical teeth: The much weaker dentition of type R14 (diapsid) resembles teeth of the lizards Cricosaura typica and Eublepharis macularis, which feed on insect larvae and spiders (Hotton 1955).
Serrations parallel towards apex: This relatively small type of tooth (R9) is the most distinctive of all Lower Keuper tetrapods known to date. Despite its considerable positional variation, the teeth all share coarse serrations aligned obliquely towards the apex of the crown (‘Spitzkerbung’ sensu Huene 1907–1908). Such teeth are present in various extant iguanids, which are almost exclusively herbivorous (Montanucci 1968; Iverson 1982).
Procolophonid teeth: The teeth closely resemble the more posterior dentary teeth in leptopleuronine procolophonid parareptiles (e.g. Scoloparia; Sues and Baird 1998). Unfortunately, the apical portions of the tooth crowns cannot be examined in occlusal view in the single known jaw fragment from Vellberg for further comparison with other known leptopleuronines.
Conclusions
-
1.
The majority of tooth morphotypes of tetrapods from the Lower Keuper indicates faunivory. With the exception of some clear-cut cases (e.g. large ziphodont carnivores, ‘pincer’-jawed piscivores), most of these dentitions cannot be unequivocally linked to a particular dietary habit. This is highlighted by some extant iguanid lizards, which almost exclusively subsist on plant matter but will occasionally consume animal protein. In the Lower Keuper environments, many small- and medium-sized diapsids were probably faunivorous.
-
2.
The number of potential piscivores in the Lower Keuper was substantially higher and more groups evolved this mode of feeding than formerly thought. This reflects the abundance and diversity of fish in and the depositional environments of the Lower Keuper (Böttcher 2015; Schoch and Seegis 2016). In addition to sauropterygians, archosauriforms (Jaxtasuchus and the larger taxon bearing R5 type teeth) and the choristodere-like reptile (R15) probably fed on fish. The frequency and diversity of fish and fish eaters is not surprising, given the abundance of biologically productive lacustrine or lagoonal environments recorded by the various Lower Keuper sedimentary deposits (Nitsch 2015).
-
3.
As for the many smaller diapsids, the question to be addressed is not so much what exactly the taxa fed on, but what kind of prey their teeth could have seized and processed by analogy to extant reptiles or based on biomechanical considerations. When the range of tetrapod dentitions present in the Lower Keuper is compared to extant tetrapod teeth, the resemblances are close and sometimes striking. In most cases, the extant analogues fall into entirely different groups (usually squamates) than their Lower Keuper analogues (mostly archosauromorphs).
-
4.
In the Lower Keuper, evidence of herbivores that used their teeth for oral processing of fodder is so far restricted to two dental morphotypes (R9, R26), suggesting either that other taxa possibly fed on plants but had no dental features indicative of herbivory or that herbivorous tetrapods were not a common faunal element in the depositional environments of the Lower Keuper. In the Lower Keuper, vegetation was at least locally abundant and diverse (Kelber 2015) and could have supported a diversified herbivore community. Surprisingly, however, the typical herbivorous synapsids and reptiles of the Middle and Late Triassic found elsewhere—dicynodonts, gomphodont cynodonts and rhynchosaurs—are virtually absent from the Lower Keuper. The available evidence indicates that a combination of both causes may explain the absence of evidence on herbivores. For instance, taxa with bulbous or multicuspid teeth could have included both herbivores and omnivores by analogy with extant lizards. Still, the absence of large herbivores, especially dicynodonts, remains a consistent feature of Lower Keuper strata (Maisch et al. 2009). The discovery of a new deposit may change this picture. Until recently, this was considered the case for all 40-million-year-spanning Keuper deposits, but the discovery of a partial dicynodont mandible in Carnian conglomerates proved that these large-bodied herbivores were present during at least part of this time interval (Schoch 2012).
References
Abler, W. L. (1992). The serrated teeth of tyrannosaurid dinosaurs, and biting structures in other animals. Paleobiology, 18, 161–183.
Alberti, F. V. (1834). Beitrag zu einer Monographie des Bunten Sandsteins, Muschelkalks und Keupers, und die Verbindung dieser Gebilde zu einer Formation (p. 366). Stuttgart: Verlag der J.G. Cotta’schen Buchhandlung.
Andrade, M. B. de, Young, M. T., Desojo, J. A., & Brusatte, S. L. (2010). The evolution of extreme hypercarnivory in Metriorhynchidae (Mesoeucrocodylia: Thalattosuchia) based on evidence from microscopic denticle morphology. Journal of Vertebrate Paleontology, 30, 1451–1465.
Andrews, S. M., & Carroll, R. L. (1991). The Order Adelospondyli: carboniferous lepospondyl amphibians. Transactions of the Royal Society of Edinburgh Earth Sciences, 82, 239–275.
Auffenberg, W. (1981). The behavioral ecology of the Komodo monitor (p. 406). Gainesville: University Presses of Florida.
Barrett, P. M. (2000). Prosauropods and iguanas: speculations on the diets in extinct reptiles. In H.-D. Sues (Ed.), Evolution of herbivory in terrestrial vertebrates: perspectives from the fossil record (pp. 42–78). Cambridge: Cambridge University Press.
Berkovitz, B., & Shellis, P. (2017). The teeth of non-mammalian vertebrates (p. 342). London: Academic Press.
Beutler, G., Hauschke, N., & Nitsch, E. (1999). Faziesentwicklung des Keupers im Germanischen Becken. In N. Hauschke & V. Wilde (Eds.), Trias. Eine ganz andere Welt (pp. 129–174). Munich: Verlag Dr. Friedrich Pfeil.
Bonaparte, J. F. (1978). El Mesozoico de América del Sur y sus tetrapodos. Opera Lilloana, 26, 1–596.
Böttcher, R. (2015). Fische. In H. Hagdorn, R.R. Schoch & G. Schweigert (Eds.), Der Lettenkeuper – Ein Fenster in die Zeit vor den Dinosauriern. Palaeodiversity Sonderband, 141–202.
Borsuk-Białynicka, M. (1996). The Late Cretaceous lizard Pleurodontagama and the origin of tooth permanency in Lepidosauria. Acta Palaeontologica Polonica, 41, 231–252.
Bystrow, A. P. (1938). Zahnstruktur der Labyrinthodonten. Acta Zoologica (Stockholm), 19, 387–425.
Cree, A. (2014). Tuatara: biology and conservation of a venerable survivor (p. 564). Canterbury: Canterbury University Press.
Damiani, R., Schoch, R. R., Hellrung, H., Werneburg, R., & Gastou, S. (2009). The plagiosaurid temnospondyl Plagiosuchus pustuliferus from the Middle Triassic of Germany: anatomy and functional morphology of the skull. Zoological Journal of the Linnean Society, 155, 348–373.
D’Amore, D. C., & Blumenschine, R. J. (2009). Komodo monitor (Varanus komodoensis) feeding behavior and dental function reflected through tooth marks on bone surfaces, and the application to ziphodont paleobiology. Paleobiology, 35, 525–552.
D’Amore, D. C., & Blumenschine, R. J. (2012). Using striated tooth marks on bone to predict body size in theropod dinosaurs: a model based on feeding observations of Varanus komodoensis, the Komodo monitor. Paleobiology, 38, 79–100.
Dorka, M. (2002). Tetrapod teeth from an Upper Ladinian bone bed, Schöningen (Lower Saxony, Germany). Paläontologische Zeitschrift, 76, 283–296.
Edmund, A. G. (1969). Dentition. In C. Gans (Ed.), Biology of the Reptilia, Vol. 1: morphology A (pp. 117–200). New York: Academic Press.
Estes, R., & Williams, E. E. (1984). Ontogenetic variation in the molariform teeth of lizards. Journal of Vertebrate Paleontology, 4, 96–107.
Etzold, A. & Schweizer, V. (2005). Der Keuper in Baden-Württemberg. In Deutsche Stratigraphische Kommission (Eds.), Stratigraphie von Deutschland IV. Keuper. Bearbeitet von der Arbeitsgruppe Keuper der Subkommission Perm-Trias der DSK. Courier Forschungsinstitut Senckenberg, 253, 215–258.
Farlow, J. O., Brinkman, D. L., Abler, W. L., & Currie, P. J. (1991). Size, shape, and serration density of theropod dinosaur lateral teeth. Historical Biology, 16, 161–198.
Fraas, E. (1900). Zanclodon schützii n. sp. aus dem Trigonodusdolomit von Hall. Jahreshefte des Vereins für vaterländische Naturkunde in Württemberg, 56, 510–513.
Gower, D. J. (1999). The cranial and mandibular osteology of a new rauisuchian archosaur from the Middle Triassic of southern Germany. Stuttgarter Beiträge zur Naturkunde, B, 280, 1–49.
Gower, D. J., & Schoch, R. R. (2009). The postcranial skeleton of the rauisuchian Batrachotomus kupferzellensis. Journal of Vertebrate Paleontology, 29, 103–122.
Hagdorn, H., & Simon, T. (1985). Geologie und Landschaft des Hohenloher Landes (p. 186). Sigmaringen: Thorbecke.
Hagdorn, H., & Mutter, R. (2011). The vertebrate fauna of the Lower Keuper Albertibank (Erfurt Formation, Middle Triassic) in the vicinity of Schwäbisch Hall (Baden-Württemberg, Germany). Palaeodiversity, 4, 223–243.
Hagdorn, H., Schoch, R.R., Seegis, D. & Werneburg, R. (2015). Wirbeltierlagerstätten im Lettenkeuper. In H. Hagdorn, R.R. Schoch & G. Schweigert (Eds.), Der Lettenkeuper – Ein Fenster in die Zeit vor den Dinosauriern. Palaeodiversity Sonderband, 325–358.
Heckert, A. B. (2004). Late Triassic microvertebrates from the lower Chinle Group (Otischalkian-Adamanian: Carnian), southwestern U.S.A. New Mexico Museum of Natural History and Science Bulletin, 27, 1–170.
Heckert, A. B., Mitchell, J. S., Schneider, V. P., & Olsen, P. E. (2012). Diverse new microvertebrate assemblage from the Upper Triassic Cumnock Formation, Sanford subbasin, North Carolina, USA. Journal of Paleontology, 86, 368–390.
Hellrung, H. (2003). Gerrothorax pustuloglomeratus, ein Temnospondyle (Amphibia) mit knöcherner Branchialkammer aus dem Unteren Keuper von Kupferzell (Süddeutschland). Stuttgarter Beiträge zur Naturkunde, B, 330, 1–130.
Heunisch, C. (1999). Die Bedeutung der Palynologie für Biostratigraphie und Fazies in der Germanischen Trias. In N. Hauschke & V. Wilde (Eds.), Trias – eine ganz andere Welt (pp. 207–220). Munich: Verlag Dr. Friedrich Pfeil.
Heunisch, C. (2015). Die Palynoflora des Lettenkeupers. In H. Hagdorn, R.R. Schoch & G. Schweigert (Eds.), Der Lettenkeuper – Ein Fenster in die Zeit vor den Dinosauriern (pp. 101–106). Palaeodiversity Sonderband.
Holmes, R. (1989). The skull and axial skeleton of the Lower Permian anthracosauroid amphibian Archeria crassidisca Cope. Palaeontographica A, 207, 161–206.
Hopson, J. A., & Sues, H.-D. (2006). A traversodontid cynodont from the Middle Triassic (Ladinian) of Baden-Württemberg. Paläontologische Zeitschrift, 80, 124–129.
Hotton III, N. (1955). A survey of adaptive relationships of dentition to diet in the North American Iguanidae. American Midland Naturalist, 53, 88–114.
Huene, F. (1905). Über die Nomenklatur von Zanclodon. Centralblatt für Mineralogie, Geologie und Paläontologie, 1905, 10–12.
Huene, F. (1907–1908). Die Dinosaurier der europäischen Triasformation mit Berücksichtigung der aussereuropäischen Vorkommnisse. Geologische und Paläontologische Abhandlungen Supplement, 1, 1–419.
Huene, F. (1922). Beiträge zur Kenntnis der Organisation einiger Stegocephalen der schwäbischen Trias. Acta Zoologica (Stockholm), 3, 395–460.
Iverson, J. B. (1982). Adaptations to herbivory in iguanine lizards. In G. M. Burghardt & A. S. Rand (Eds.), Iguanas of the world: their behavior, ecology and conservation (pp. 60–76). Park Ridge: Noyes Publications.
Jaeger, G. F. (1824). De Ichthyosauri sive Proteosauri fossilis speciminibus in agro Bollensi in Wirtembergia repertis (p. 14). Stuttgart: Verlag der J.G. Cotta’schen Buchhandlung.
Jaeger, G. F. (1828). Über die fossile [sic] Reptilien, welche in Württemberg aufgefunden worden sind (p. 48). Stuttgart: Verlag der J.B. Metzler’schen Buchhandlung.
Jones, M. E. (2008). Skull shape and feeding strategy in Sphenodon and other Rhynchocephalia (Diapsida: Lepidosauria). Journal of Morphology, 269, 945–966.
Jones, M. E., Anderson, C. L., Hipsley, C. A., Müller, J., Evans, S. E., & Schoch, R. R. (2013). Integration of molecules and new fossils supports a Triassic origin for Lepidosauria (lizards, snakes, and tuatara). BMC Evolutionary Biology, 13, 208.
Kelber, K.P. (2015). Die Makroflora des Lettenkeupers. In H. Hagdorn, R.R. Schoch & G. Schweigert (Eds.), Der Lettenkeuper – Ein Fenster in die Zeit vor den Dinosauriern. Palaeodiversity Sonderband, (pp. 51–100).
Langston Jr., W. (1975). Ziphodont crocodiles: Pristichampsus vorax (Troxell), new combination, from the Eocene of North America. Fieldiana Geology, 33, 291–314.
Löffler, T., Thies, D., Vespermann, J., & Zellmer, H. (2007). Fauna, Sedimentologie und Geochemie des Unteren Keupers (Erfurt Formation) von Schöningen. Berichte der Naturhistorischen Gesellschaft Hannover, 149, 3–39.
Lucas, P. W. (2004). Dental functional morphology: how teeth work (p. 355). Cambridge: Cambridge University Press.
Mahler, L. D., & Kearney, M. (2006). The palatal dentition in squamate reptiles: morphology, development, attachment, and replacement. Fieldiana Zoology, New Series, 182, 1–61.
Maisch, M. W., Vega, C. S., & Schoch, R. R. (2009). No dicynodont in the Keuper – A reconsideration of the occurrence of aff. Dinodontosaurus in the Middle Triassic of southern Germany. Palaeodiversity, 2, 271–278.
Martill, D. M. (1986). The diet of Metriorhynchus, a Mesozoic marine crocodile. Neues Jahrbuch für Geologie and Paläontologie Monatshefte, 1986, 621–625.
Martínez, R. N., Apaldetti, C., Alcober, O. A., Colombi, C. E., Sereno, P. C., Fernandez, E., Malnis, P. S., Ciorrea, G. A., & Abelin, D. (2013). Vertebrate succession in the Ischigualasto Formation. Society of Vertebrate Paleontology Memoir, 12, 10–39.
Massare, J. A. (1987). Tooth morphology and prey preference of Mesozoic marine reptiles. Journal of Vertebrate Paleontology, 7, 121–137.
Mateer, N. J. (1974). Three Mesozoic crocodiles in the collections of the Palaeontological Museum, Uppsala. Bulletin of the Geological Institute University of Uppsala, New Series, 4, 53–72.
Maxwell, E. E., Caldwell, M. W., & Lamoureux, D. O. (2011a). The structure and phylogenetic distribution of amniote plicidentine. Journal of Vertebrate Paleontology, 31, 553–561.
Maxwell, E. E., Caldwell, M. W., Lamoureux, D. O., & Budney, L. A. (2011b). Histology of tooth attachment tissues and plicidentine in Varanus (Reptilia: Squamata), and a discussion of the evolution of amniote tooth attachment. Journal of Morphology, 31, 553–561.
Meyer, H. V., & Plieninger, T. (1844). Beiträge zur Paläontologie Württemberg’s enthaltend die fossilen Wirbelthierreste aus den Triasgebilden mit besonderer Rücksicht auf die Labyrinthodonten des Keupers (p. 132). Stuttgart: E. Schweizerbart’sche Verlagsbuchhandlung.
Modesto, S. P., & Sues, H.-D. (2004). The skull of the Early Triassic archosauromorph reptile Prolacerta broomi and its phylogenetic significance. Zoological Journal of the Linnean Society, 140, 335–351.
Montanucci, R. R. (1968). Comparative dentition in four iguanid lizards. Herpetologica, 24, 305–315.
Moser, M., & Schoch, R. (2007). Revision of the type material and nomenclature of Mastodonsaurus giganteus (Jaeger) (Temnospondyli) from the Middle Triassic of Germany. Palaeontology, 50, 1245–1266.
Munk, W., & Sues, H.-D. (1993). Gut contents of Parasaurus (Pareiasauria) and Protorosaurus (Archosauromorpha) from the Kupferschiefer (Upper Permian) of Hessen, Germany. Paläontologische Zeitschrift, 67, 169–176.
Nesbitt, S. J. (2011). The early evolution of archosaurs: relationships and the origin of major clades. Bulletin of the American Museum of Natural History, 352, 1–292.
Nitsch, E. (2015). Fazies und Ablagerungsräume. In H. Hagdorn, R.R. Schoch & G. Schweigert (Eds.), Der Lettenkeuper – Ein Fenster in die Zeit vor den Dinosauriern. Palaeodiversity Sonderband, 285–324.
Nosotti, S. (2007). Tanystropheus longobardicus (Reptilia, Protorosauria): re-interpretations of the anatomy based on new specimens from the Middle Triassic of Besano (Lombardy, northern Italy). Memorie della Società Italiana d Scienze Naturali e del Museo Civico di Storia Naturale di Milano, 35(3), 1–88.
Owen, R. (1841). On the teeth of species of the genus Labyrinthodon (Mastodonsaurus salamandroides and Phytosaurus) of Jaeger, from the German Keuper and the sandstone of Warwick and Leamington. Proceedings of the Geological Society London, 3, 357–360.
Peyer, B. (1931). Die Triasfauna der Tessiner Kalkalpen II. Tanystropheus longobardicus Bass. sp. Abhandlungen der Schweizerischen Paläontologischen Gesellschaft, 50, 5–110.
Plieninger, T. (1847). Über ein neues Sauriergenus und die Einreihung der Saurier mit flachen, schneidenden Zähnen in eine Familie. Jahreshefte des Vereins für vaterländische Naturkunde in Württemberg, 2, 148–154.
Preuschoft, H., Reif, W. E., Loitsch, C., & Tepe, E. (1991). The function of labyrinthodont teeth: big teeth in small jaws. In N. Schmitt-Kittler & K. Vogel (Eds.), Constructional morphology and evolution (pp. 151–171). Heidelberg: Springer.
Quenstedt, F. A. (1885). Handbuch der Petrefaktenkunde (p. 982). Tübingen: Laupp.
Rieppel, O. (2000). Sauropterygia I: Placodontia, Pachypleurosauria, Nothosauroidea, Pistosauroidea. In P. Wellnhofer (Ed.), Handbuch der Paläoherpetologie, Teil 12A (p. 134). Munich: Verlag Dr. Friedrich Pfeil.
Rieppel, O. (2002). Feeding mechanics in Triassic stem group sauropterygians: the anatomy of a successful invasion of Mesozoic seas. Zoological Journal of the Linnean Society, 135, 33–63.
Sanchez, S., & Schoch, R. R. (2013). Bone histology reveals a high environmental and metabolic plasticity as a successful evolutionary strategy in a long-lived homeostatic Triassic temnospondyl. Evolutionary Biology, 40, 627–647.
Schoch, R. R. (1997). A new capitosaur amphibian from the Lower Lettenkeuper (Triassic: Ladinian) of Kupferzell (southern Germany). Neues Jahrbuch für Geologie und Paläontologie Abhandlungen, 203, 239–272.
Schoch, R. R. (1999). Comparative osteology of Mastodonsaurus giganteus (Jaeger, 1928) from the Middle Triassic (Lettenkeuper: Longobardian) of Germany (Baden-Württemberg, Bayern, Thüringen). Stuttgarter Beiträge zur Naturkunde, B, 178, 1–173.
Schoch, R. R. (2002). Stratigraphie und Taphonomie wirbeltierreicher Schichten im Unterkeuper (Mitteltrias) von Vellberg (SW-Deutschland). Stuttgarter Beiträge zur Naturkunde, B, 318, 1–30.
Schoch, R. R. (2006). A complete trematosaurid amphibian from the Middle Triassic of Germany. Journal of Vertebrate Paleontology, 26, 29–43.
Schoch, R. R. (2008). A new stereospondyl from the Middle Triassic of Germany, and the origin of the Metoposauridae. Zoological Journal of the Linnean Society, 152, 79–113.
Schoch, R. R. (2009). The evolution of life cycles in early amphibians. Annual Review of Earth and Planetary Sciences, 37, 135–162.
Schoch, R. R. (2011a). A procolophonid-like tetrapod from the Middle Triassic of Germany. Neues Jahrbuch für Geologie und Paläontologie Abhandlungen, 259, 251–255.
Schoch, R. R. (2011b). New archosauriform remains from the German Lower Keuper. Neues Jahrbuch für Geologie und Paläontologie Abhandlungen, 260, 87–100.
Schoch, R. R. (2012). A dicynodont mandible from the Triassic of Germany forms the first evidence of large herbivores in the Central European Basin. Neues Jahrbuch für Geologie und Paläontologie Abhandlungen, 263, 119–123.
Schoch, R. R. (2014). Amphibian evolution: the life of early land vertebrates (p. 288). Chichester: Wiley.
Schoch, R.R. (2015a). Amphibien und Chroniosuchier. In H. Hagdorn, R.R. Schoch & G. Schweigert (Eds.), Der Lettenkeuper – Ein Fenster in die Zeit vor den Dinosauriern. Palaeodiversity Sonderband, 203–230.
Schoch, R.R. (2015b). Reptilien. In H. Hagdorn, R.R. Schoch & G. Schweigert (Eds.), Der Lettenkeuper – Ein Fenster in die Zeit vor den Dinosauriern. Palaeodiversity Sonderband, (pp. 231–264).
Schoch, R. R., & Milner, A. R. (2000). Stereospondyli. In P. Wellnhofer (Ed.), Handbuch der Paläoherpetologie (Vol. 3B, p. 203). Munich: Verlag Dr. Friedrich Pfeil.
Schoch, R. R., & Milner, A. R. (2014). Temnospondyli I. In H. D. Sues (Ed.), Handbuch der Paläoherpetologie, Vol. 3A2 (p. 150). Munich: Verlag Dr. Friedrich Pfeil.
Schoch, R. R., & Seegis, D. (2016). A Middle Triassic palaeontological gold mine: the vertebrate deposits of Vellberg (Germany). Palaeogeography Palaeoclimatology Palaeoecology, 459, 249–267.
Schoch, R. R., & Sues, H.-D. (2014). A new archosauriform reptile from the Middle Triassic (Ladinian) of Germany. Journal of Systematic Palaeontology, 12, 113–131.
Schoch, R. R., & Sues, H.-D. (2015). A Middle Triassic stem-turtle and the evolution of the turtle body plan. Nature, 523, 584–587.
Schoch, R.R. & Sues, H.-D. (2017). Osteology of the Middle Triassic stem-turtle Pappochelys rosinae and the evolution of the turtle skeleton. Journal of Systematic Palaeontology. https://doi.org/10.1080/14772019.2017.1354936.
Schoch, R. R., & Wild, R. (1999). Die Saurier von Kupferzell – der gegenwärtige Forschungsstand. In N. Hauschke & V. Wilde (Eds.), Die Trias – eine ganz andere Welt (pp. 409–418). Munich: Verlag Dr. Friedrich Pfeil.
Schoch, R. R., & Witzmann, F. (2012). Cranial morphology of the plagiosaurid Gerrothorax pulcherrimus as an extreme example of evolutionary stasis. Lethaia, 45, 371–385.
Schoch, R. R., Milner, A. R., & Witzmann, F. (2014). Skull morphology and phylogenetic relationships of a new Middle Triassic plagiosaurid temnospondyl from Germany, and the evolution of plagiosaurid eyes. Palaeontology, 57, 1045–1058.
Schubert, B. W., & Ungar, P. S. (2005). Wear facets and enamel spalling in tyrannosaurid teeth. Acta Palaeontologica Polonica, 50, 93–99.
Schultze, H.-P. (1969). Die Faltenzähne der rhipidistiiden Crossopterygier, der Tetrapoden und der Actinopterygier-Gattung Lepisosteus, nebst einer Beschreibung der Zahnstruktur von Onychodus (struniiformer Crossopterygier). Palaeontologia Italiana, 65, 63–137.
Shishkin, M. A., Ochev, V. G., Lozovskii, V. R., & Novikov, I. V. (2000). Tetrapod biostratigraphy of the Triassic of Eastern Europe. In M. J. Benton, M. A. Shishkin, D. M. Unwin, & E. N. Kurochkin (Eds.), The age of dinosaurs in Russia and Mongolia (pp. 120–139). Cambridge: Cambridge University Press.
Smith, J. B., & Dodson, P. (2003). A proposal for a standard terminology of anatomical notation and orientation in fossil vertebrate dentitions. Journal of Vertebrate Paleontology, 23, 1–12.
Sues, H.-D., & Baird, D. (1998). Procolophonidae (Reptilia: Parareptilia) from the Upper Triassic Wolfville Formation of Nova Scotia, Canada. Journal of Vertebrate Paleontology, 18, 525–532.
Sues, H.-D., & Fraser, N. C. (2010). Triassic life on land: the great transition (p. 236). New York: Columbia University Press.
Sues, H.-D., & Schoch, R. R. (2013). First record of Colognathus (?Amniota) from the Middle Triassic of Europe. Journal of Vertebrate Paleontology, 33, 998–1002.
Welman, J. (1998). The taxonomy of the South African proterosuchids (Reptilia, Archosauromorpha). Journal of Vertebrate Paleontology, 18, 340–347.
Wild, R. (1973). Die Triasfauna der Tessiner Kalkalpen. XXIII. Tanystropheus longobardicus (Bassani) (Neue Ergebnisse). Schweizerische Paläontologische Abhandlungen, 95, 1–162.
Wild, R. (1980). The fossil deposits of Kupferzell, southwest Germany. Mesozoic Vertebrate Life, 1, 15–18.
Witzmann, F. & Schoch, R.R. (2018). Skull and postcranium of the bystrowianid Bystrowiella schumanni from the Middle Triassic of Germany, and the position of chroniosuchians within Tetrapoda. Journal of Systematic Palaeontology16, 711-739. https://doi.org/10.1080/14772019.2017.1336579.
Witzmann, F., Schoch, R. R., & Maisch, M. W. (2008). A relict basal tetrapod from the Middle Triassic of Germany. Naturwissenschaften, 95, 67–72.
Zweifel, R. G., & Lowe, C. H. (1966). The ecology of a population of Xantusia vigilis, the desert night lizard. American Museum Novitates, 2247, 1–57.
Acknowledgements
The senior author thanks Werner Kugler, Traugott and Ute Haubold, Alfred Bartholomä, Michael Salomon, Christoph Straub, Fabian Mohl and Tobias Mann for donating numerous specimens to the SMNS and providing invaluable help during field work and the Schumann family for granting access to their property and their continued support. Norbert Adorf, Isabell Rosin, Achim Lehmkuhl, Marit Kamenz, Martina Battenstein and Kim Krämer are thanked for their skillful fossil preparation and Ronald Böttcher, Julia Desojo, Susan Evans, Randall Irmis, Sterling Nesbitt, Erin Maxwell, Johannes Müller and Marc Jones for fruitful discussions. We thank the two reviewers, Sterling Nesbitt and Ralf Werneburg, for their helpful suggestions.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
The authors declare that they have no conflict of interest.
Rights and permissions
About this article

Cite this article
Schoch, R.R., Ullmann, F., Rozynek, B. et al. Tetrapod diversity and palaeoecology in the German Middle Triassic (Lower Keuper) documented by tooth morphotypes. Palaeobio Palaeoenv 98, 615–638 (2018). https://doi.org/10.1007/s12549-018-0327-2
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s12549-018-0327-2