
Abstract
A rich and diverse ichthyofauna is described from the upper Oligocene (Egerian) sands of Máriahalom, Hungary. The site is dominated by brackish molluscs that are preserved together with rare marine and terrestrial vertebrates. Based on the isolated elasmobranch and bony fish remains, eight sharks, four rays and seven teleost taxa were identified from Máriahalom. The ichthyofauna represents a nearshore marine subtropical community dominated by odontaspidid and carcharhinid sharks and by euryhaline durophagous sparids and by sciaenid teleost fishes. Epibenthic feeders were common, whereas piscivorous taxa included barracudas, billfishes and numerous sharks, including the macropredatory Otodus angustidens. Palaeontological, sedimentological and stratigraphical data indicates a tide-influenced and fluvial-influenced nearshore palaeoenvironment with brackish lagoons and normal marine littoral habitats. The remains of terrestrial, brackish and nearshore marine invertebrates and vertebrates accumulated in a tidal channel. The systematics of the ichthyofauna is consistent with the palaeogeography inferred from invertebrates, i.e. a Central Paratethys connected to the Mediterranean area during the Egerian.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
The Paratethys was a predominantly epicontinental sea separated from the Mediterranean area by the Alpine orogeny during the early Oligocene and initially occupying the modern-day area between the Western Alps and the Aral Lake (Fig. 1a). The Central Paratethys covered the Intra-Carpathian region and was bordered by the Western Paratethys in the Alpine Foreland and the Eastern Paratethys in the present Ponto-Caspian area (Popov et al. 2004; Piller et al. 2007). In general, the evolution of the Paratethys was characterised by a west to east up-filling and the alternating periods of isolation and re-connection from and towards the Mediterranean and Boreal Province (Piller et al. 2007). Isolated phases led to low-salinity (brackish) conditions and the establishment of euryhaline and often endemic faunas when isolation persisted for a sufficiently long time (Báldi 1989; Rögl 1998). Periods with oceanic connections, on the other hand, allowed for the invasion of more global, normal saline faunas (Rögl 1998; Popov et al. 2004).

Following an early Oligocene isolation and poorly oxygenated phase, normal marine conditions were re-established in the Paratethys by the late early Oligocene (Báldi 1989; Rögl 1998; Popov et al. 2004). This was due to a newly formed seaway between the Central Paratethys and the Mediterranean area that extended across the present area of Slovenia (Rögl 1998; Popov et al. 2004). Faunal exchanges between the two marine realms are already well documented in the Kiscell Clay Formation (Báldi 1983; Nagymarosy 1990), and in younger marine formations of the upper Oligocene (Egerian stage). The dispersal of Mediterranean and Boreal invertebrates into the Central Paratethys is well-recognised (Rögl and Steininger 1984), while marine vertebrates have been poorly investigated due to the lack of well-documented fossil records. Here, we report on a Central Paratethyan marine vertebrate fauna from the upper Oligocene of Hungary and test whether its composition is consistent with a Central Paratethys that was open towards the Mediterranean area.
Although the Oligocene of the Intra-Carpathian area is rich in elasmobranch and bony fishes, only a limited number of studies deal with them in detail (e.g. Weiler 1933, 1938; Böhm 1942; Solt 1988; Szabó and Kocsis in press). Moreover, a study of fish otoliths from the Eger Formation is the only work from the latest Oligocene (Egerian; Nolf and Brzobohaty 1994). We present a detailed taxonomic description of the elasmobranch and actinopterygian fishes from the late Oligocene Egerian Máriahalom site. Máriahalom is particularly interesting because it has yielded a predominantly brackish invertebrate fauna with an underrepresentation of normal marine taxa (Báldi and Cságoly 1975; Janssen 1982; Báldi 1983) during a period when normal marine conditions characterised the Central Paratethys. The systematic and palaeoecological evaluation of the vertebrate fauna together with the sedimentological and stratigraphical assessment provided herein shed further light on the palaeoenvironmental conditions at the area.
Locality and stratigraphy
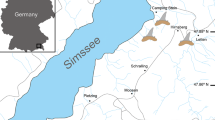
The upper Oligocene Máriahalom locality is a sandpit located 47 km north-west from Budapest (Hungary), along the south-western side of the road between the villages of Úny and Máriahalom (Fig. 2a). During the early Oligocene, a large part of the Transdanubian Range was uplifted and the consequent denudation removed the top part of the Eocene sections, while restricted, anoxic shales (Tard Clay) were deposited in the deep basin to the east (Telegdi-Roth 1927; Báldi 1983; Kázmér et al. 2003). Due to renewed subsidence, the later part of the Kiscellian stage (late Rupelian; Piller et al. 2007) was characterised by freshwater to deep marine deposition. To the west, the alluvial Csatka Formation and the littoral to neritic Hárshegy Formation with local bauxite, kaolinite and coal accumulations were formed (Sztanó et al. 1998; Nagymarosy 2012). In the deep basin to the E–SE, open marine circulation became restored and the Kiscell Clay accumulated as a bathyal deposit over a vast region (Báldi 1980). During the Egerian stage, sediment input from the west increased and long-term normal regression resulted in the interfingering alluvial Csatka and the marginal marine to neritic Törökbálint Formations (Korpás 1981; Nagymarosy and Gyalog 1997; Sztanó et al. 1998) (Fig. 1b). The Máriahalom locality is part of the Mány Member of the Törökbálint Formation. The Törökbálint Formation includes deposits from different palaeoenvironments including brackish to normal marine conditions (Korpás 1981; Nagymarosy 2012). The Mány Member, with a predominantly brackish mollusc community (e.g. Pirenella, Polymesoda, Potamides, Brotia; Báldi and Cságoly 1975), represents a transition between alluvial and normal marine conditions and is interpreted as a product of lagoonal environments (Báldi 1967; Nagymarosy and Gyalog 1997; Nagymarosy 2012). However, since the Egerian normal regression is documented at several nearby sections (e.g. Báldi 1983; Sztanó et al. 1998), the brackish lagoon was possibly part of a delta plain environment. The Mány Member of the Törökbálint Formation consists dominantly of calcareous silt and sandstone with coal string and variegated clay intercalations deposited in a brackish, shallow water environment. It corresponds to the Egerian stage based on mollusc biostratigraphy of more than 50 boreholes (Báldi 1967, 1973; Báldi and Cságoly 1975; Nagymarosy and Gyalog 1997; Báldi et al. 1999).

Locality map (a) and schematic stratigraphic section of the Máriahalom vertebrate site (b)
The age of the basal and lower part of the Egerian stage appears to correspond to the late and terminal Chattian, while the upper part of the Egerian can be correlated to the early Miocene (Aquitanian or Aquitanian/Burdigalian boundary) based on biostratigraphy and preliminary Sr-isotope stratigraphy data (e.g. Báldi et al. 1999; Piller et al. 2007; Less et al. 2015). The Paleogene/Neogene boundary is difficult to detect in the Egerian stage since the index fossils for the Aquitanian (Paragloborotalia kugleri) are absent in the Central Paratethys region (e.g. Báldi et al. 1999; Piller et al. 2007). However, the Máriahalom site most probably belongs to the lower part of the Egerian stage (probably equivalent to the upper Oligocene part of the Egerian) because the presence of the anthracotheriid mammal Microbunodon minimum (Cuvier, 1822) at the site (Rabi and Botfalvai 2008) constrains the age to the MP28–MP30 zone of the late Oligocene (late Chattian; Russell et al. 1982; Lihoreau et al. 2004; Scherler et al. 2013).
The rich mollusc fauna of Máriahalom was classified as a typical Potamides-Pirenella community, possibly indicating a brackish lagoon environment although normal marine taxa (e.g. Glycymeris latiradiata, Mytilus aqutanicus) are also present in lower abundance (Báldi and Cságoly 1975; Janssen 1982). The section predominately consists of medium-grained sand, sandy clay and interbedding sandstone layers. The highly matured quartz sand is moderately sorted, with very low carbonate content. The lower part of the Máriahalom section contains 10–12% of clay, while in the upper part, it is reduced to 2–4%. The most common sedimentary structure at the upper part of the section of Máriahalom is the trough cross-stratification that ranges in thickness from 150 to 200 cm, while mud drapes are common at the lower part of the section indicating some (meso?) tidal influence (Fig. 2b). According to Báldi and Cságoly (1975), the water depth did not exceed 2–5 m even at high tides based on the Potamides-Pirenella community, whereas the size of cross-bedding points to a minimum depth of 7–15 m and probable channelization.
The Máriahalom locality yielded fossils of at least 26 vertebrate taxa, including sharks, rays, bony fishes, turtles, crocodilians, birds and large mammals, making it the oldest diverse vertebrate assemblage from the Cenozoic of Hungary. The Máriahalom vertebrate assemblage is dominated by isolated teeth and bones of aquatic and semi-aquatic animals, including several marine taxa, whereas terrestrial mammals coming from the same bonebed are underrepresented.
All the vertebrate fossils are found isolated, and they have been collected from almost the entire area of the sandpit area. However, increased abundance of vertebrate remains is linked to 2–5-m-wide, 1–1.5-m-thick coquina accumulations, which are composed almost entirely of transported, abraded and mechanically sorted mollusc shells (Rabi and Botfalvai 2008). Remains of shallow marine, freshwater and terrestrial vertebrates are found together in these lenses, and this fossil accumulation may have been interpreted as a lag deposit of a tidal channel.
The marked trough cross-stratifications, the coarser grained sediment, the low clay content and the coquina-like fossil accumulation (including the skeletal elements of terrestrial and marine animals) indicate that the depositional environment was characterised by high-energy conditions in a shallow water environment, where bones of animals from different habitats were washed into concentrated fossil lenses before the final burial.
Materials and methods
While the remarkably abundant and well-preserved mollusc assemblage of the Máriahalom sandpit has received considerable attention (Báldi 1967, 1973; Báldi and Cságoly 1975; Janssen 1982), the vertebrate fossils remained undiscovered until the 1980s, when private collectors began to visit the locality. The study of the vertebrate remains began to intensify thanks to excavations conducted between 1999 and 2006 (Rabi and Botfalvai 2008). All of these specimens, consisting of hundreds of isolated bones and teeth, have been donated to the vertebrate palaeontological collection of the Hungarian Natural History Museum (MTM). All of the 565 fish remains (173 shark, 94 ray and 298 teleost fossils) described herein have been found isolated, mostly by screening at the Máriahalom sandpit. Taxonomy of chondrichthyans follows Cappetta (2012) and Reinecke et al. (2001, 2005, 2014).
Systematic palaeontology
Classis: Chondrichthyes Huxley, 1880
Ordo Squatiniformes De Buen, 1926
Familia Squatinidae Bonaparte, 1838
Genus Squatina Duméril, 1806
Squatina sp.
Fig. 3

Squatina sp. teeth from the Egerian of Máriahalom (Hungary). a Anterolateral tooth (VER 2016.2593.) in apical view, b in lingual view, c in labial view and d in basal view. e Anterior tooth (VER 2016.2592.) in apical view, f in lingual view, g in labial view and h in basal view. Scale bar 10 mm
Referred material: Five teeth (MTM VER 2016.2591., VER 2016.2592., VER 2016.2593., VER 2016.2601.)
The referred teeth from Máriahalom exhibit a labiolingually weakly bent main cusp without lateral cusplets (Fig. 3). The main cusp is pointed, narrow and convex on both sides. The cutting edges are smooth. The flattened, weakly bent or nearly straight root has a triangle-shaped contour in basal view. The root is widened mesiodistally and bears a large central foramen on its basal side. The Máriahalom teeth are indistinguishable from other Squatina teeth reported from other Oligocene localities in Europe (e.g. Müller 1996; Baut and Génault 1999; Reinecke et al. 2014); however, the Máriahalom material also shows affinities with the species Squatina angeloides. On the other hand, the conservative evolution of Squatina dentition makes tooth-based separation of the species often problematic (Cappetta 2012).
Ordo Lamniformes Berg, 1958
Familia Odontaspididae Müller and Henle, 1839
Odontaspididae indet.
Referred material: 54 teeth (MTM VER 2016.2589., VER 2016.2605., VER 2016.2607., VER 2016.2613., VER 2016.2744.)
Many teeth are assigned to odontaspid sharks, based on the sigmoid shape of the main cusp (in case of anterior files), the smooth cutting edges and the bifurcated root (see the taxa in Cappetta 2012). However, due to their poor preservation (missing root and/or missing lateral cusplets, etc.), these cannot be identified below family level. The teeth originate from various positions of the jaw. It remains unclear whether these teeth belonged to any of the odontaspidid taxa described below or some of them represent separate taxa.
Genus Araloselachus Glikman, 1964
Araloselachus cuspidatus (Agassiz, 1843)
Fig. 4

Araloselachus cuspidatus teeth from the Egerian of Máriahalom (Hungary). a Anterior tooth (VER 2016.2586.) in lingual view, b in profile view and c in labial view. d Upper lateral tooth (VER 2016.2587.) in lingual view, e in mesial view and f in labial view. g Distal tooth (VER 2016.2561.) in lingual view and h in labial view. i Parasymphyseal tooth (VER 2015.2585.) in lingual view, j in profile view and k in labial view. l Parasymphyseal tooth (VER 2016.2570.) in lingual view, m in profile view and n in labial view. Scale bar 10 mm
Synonymy
1843 Lamna cuspidata n. sp.—Agassiz; p. 290, Pl. 37a, figs. 43–50
1903 Lamna (Odontaspis) cuspidata Ag.—Koch; p. 32, Pl. 1, figs. 13a–13c
1978 Odontaspis (Synodontaspis) cuspidata cuspidata (Ag.)—Brzobohatý and Schultz; p. 443, Pl. 1, fig. 11
1993 Carcharias cuspidata (Agassiz, 1843)—Baut; p. 4, figs. 22 and 24
1995 Synodontaspis cuspidata (Agassiz, 1844)—Holec et al.; p. 40–41, Pl. 11, figs. 2, 4–6 and Pl. 12, fig. 1
1999 Carcharias cuspidata (Agassiz, 1844)—Baut and Génault; p. 15–16, Pl. 3, figs. 3–6
2001 Carcharias cuspidatus (Agassiz, 1843)—Holec; p. 121–123, 123; Pl. 1, figs. 5 and 6a and Pl. 2, fig. 1
2003 Carcharias cuspidata Agassiz, 1843—Antunes and Balbino; p. 142, Pl. 2, fig. 6
2007 Carcharias cuspidatus (Agassiz, 1843)—Kocsis; p. 32, Pl. 4, figs. 12–13
2010 Carcharias cuspidata (Agassiz, 1843)—Hovestadt et al.; p. 60, figs. 5–7
2012 Araloselachus cuspidatus (Agassiz, 1843)—Cappetta; p. 191, fig. 180
2014 Araloselachus cuspidatus (Agassiz, 1843)—Reinecke et al.; p. 17–20, Pl. 16–18 (with all figures)
Referred material: 42 teeth (MTM VER 2016.2561., VER 2016.2570., VER 2016.2571., VER 2016.2572., VER 2016.2573., VER 2016.2574., VER 2016.2575., VER 2016.2576., VER 2016.2579., VER 2016.2580., VER 2016.2581., VER 2016.2582., VER 2016.2583., VER 2016.2584., VER 2016.2585., VER 2016.2586., VER 2016.2587., VER 2016.2590., VER 2016.2602., VER 2016.2604., VER 2016.2606., VER 2016.2609., VER 2016.2610., VER 2016.2611., VER 2016.2612., VER 2016.2743.)
Remarks: Araloselachus has been considered as a synonym of Carcharias, but Cappetta (2012) separated it as a different genus.
A. cuspidatus is the most abundant shark species at the Máriahalom locality. The teeth are strongly convex on their lingual side, whereas they are weakly convex or nearly flat on their labial side. The cutting edges are non-serrated, and the profile of the cusp is rather straight (see Fig. 4b). The wide lateral cusplets of anteriors are low, wide and roundish in cross section, while those of laterals are also wide, but flattened labiolingually with rounded contour. The root is thick and massive on all teeth and has two lobes with a well-developed bulge and transversal groove. Anterior teeth (see Fig. 4a–c) are high, symmetrical and straight, while upper laterals are distally bent (see Fig. 4d–f).
We distinguished these teeth from other Máriahalom odontaspid teeth by their proportions (teeth of A. cuspidatus are much more robust than those of any other local odontaspids; the anteriors are wider and more triangular in labial view), the small and simple lateral cusplets, and by the anterior files that are not sigmoidal in profile (see Cappetta 2012).
A. cuspidatus is widely known from the Oligocene and Miocene teeth localities of Europe and North America (e.g. Cappetta 1987; Holec et al. 1995; Kocsis 2007; Reinecke et al. 2014).
Genus Carcharias Rafinesque, 1810
Carcharias cf. acutissima (Agassiz, 1843)
Fig. 5

Carcharias cf. acutissima teeth from the Egerian of Máriahalom (Hungary). a Anterior tooth (VER 2016.2569.) in lingual view, b in profile view and c in labial view. d Upper lateral tooth (VER 2016.2563.) in lingual view, e in profile view and f in labial view. g Lower anterolateral-lateral tooth (VER 2016.2566.) in lingual view, h in profile view and i in labial view. Scale bar 10 mm
Synonymy
1843 Lamna (Odontaspis) acutissima sp. nov.—Agassiz; p. 294, Pl. 37a, figs. 33 and 34
1993 Carcharias acutissima (Agassiz, 1844)—Baut; p. 4, figs. 20, 23 and 25
1999 Synodontaspis acutissima (Agassiz, 1844)—Baut and Génault; p. 16, Pl. 3, figs. 1 and 2
2001 Carcharias acutissimus (Agassiz, 1844)—Reinecke et al.; p. 11–13, Pl. 11–15 and Pl. 23, figs. a and b
2010 Carcharias acutissima (Agassiz, 1843)—Hovestadt et al.; p. 60, figs. 3 and 4
Referred material: 24 teeth (MTM VER 2016.2563., VER 2016.2564., VER 2016.2566., VER 2016.2567., VER 2016.2569., VER 2016.2578., VER 2016.2588., VER 2016.2608., VER 2016.2740., VER 2016.2755.)
These sand tiger shark teeth are different from those of Carcharias gustrowensis (see below) in having a striated lingual face on the main cusp. They are also much smaller and more slender than those of A. cuspidatus. However, the anterior teeth of C. acutissima bear hook-like lateral cusplets that cannot be observed in the Máriahalom teeth. The teeth are therefore referred to as C. cf. acutissima until additional specimens will be discovered.
Carcharias gustrowensis (Winkler, 1875)
Fig. 6

Carcharias gustrowensis teeth from the Chattian of Máriahalom (Hungary). a Anterior tooth (VER 2016.2562.) in lingual view, b in profile view and c in labial view. d Anterior tooth (VER 2016.2565.) in lingual view, e in profile view and f in labial view. Scale bar 10 mm
Synonymy
1875 Odontaspis gustrowensis n. sp.—Winkler; p. 98, Pl. 2, figs. 1 and 2
1999 Synodontaspis acutissima (Agassiz, 1844)—Baut and Génault; p. 16, Pl. 3, fig. 1
2005 Carcharias gustrowensis (Winkler, 1875)—Reinecke et al.; p. 19–23, Pl. 13–14 (with all figures)
2010 Carcharias gustrowensis (Winkler, 1875)—Hovestadt and Hovestadt-Euler; p. 88–90, fig. 6
2010 Carcharias gustrowensis (Winkler, 1875)—Hovestadt et al.; p. 60, fig. 33C–U
2014 Carcharias gustrowensis (Winkler, 1875)—Reinecke et al.; p. 15–17, Pl. 12–15 (with all figures) and Pl. 19, figs. 1–8
Referred material: three teeth (MTM VER 2016.2562., VER 2016.2565., VER 2016.2577.)
The lack of striae on the lingual face of the crown has been mentioned by several authors as a character of C. gustrowensis (e.g. Haye et al. 2008; Cicimurri and Knight 2009; Hovestadt and Hovestadt-Euler 2010; Hovestadt et al. 2010), and in this feature, the herein referred teeth differ from those of C. acutissima (e.g. see Kocsis 2007; Hovestadt et al. 2010). The teeth bear one or two pairs of pointed, spike-like lateral cusplets that are nearly straight in labiolingual view. Due to the latter feature, it is also easy to distinguish the remains from those of C. acutissima, which are characterised by shorter and hooked cusplets. The cusplets diverge from the main cusp. When two pairs of cusplets are present, the first pair is larger than the second, external pair. The cusplets of anterior teeth are labiolingually bent. The root is strongly bifurcated with a large nutritive groove and central bulge. The anterior teeth are symmetrical or nearly symmetrical (Fig. 6), with a slender main cusp, sigmoid in lateral view.
C. gustrowensis was widely distributed in the Paratethys and North Sea Basin from the late Oligocene to the early–middle Miocene, and it seems to occur in the Chattian—early Aquitanian of Virginia (USA) as well (Reinecke et al. 2005, 2014; Haye et al. 2008).
Familia Lamnidae Müller and Henle, 1838
Genus Carcharoides Ameghino, 1901
Carcharoides catticus (Philippi, 1846)
Fig. 7a–c

Lamnid and otodontid teeth from the Egerian of Máriahalom (Hungary). a Carcharoides catticus upper lateral tooth (VER 2016.2568.) in lingual view, b in mesial view and c in labial view. d Cosmopolitodus sp. upper anteriolateral/lateral tooth (VER 2016.2756.) in lingual view, e in mesial view and f in labial view. g Otodus (Carcharocles) angustidens distal tooth (VER 2016.2560.) in lingual view and h in labial view. Scale bars 10 mm
Synonymy
1846 Otodus catticus n. sp.—Philippi; p. 24, Pl. 2, figs. 5–7
1879 Otodus debilis n. sp.—Probst; p. 155, Pl. 2, figs. 78–81
1903 Lamna cfr. compressa, Ag.—Koch; p. 33–34, Pl. 1, fig. 17
1934 Lamna cattica Philippi—Rocabert; p. 84–87, Pl. 1, fig. 17
1995 Carcharoides catticus (Philippi, 1846)—Holec et al.; p. 42, Pl. 12, fig. 2
2005 Carcharoides catticus (Philippi, 1846)—Reinecke et al.; p. 28–30, Pl. 19, figs. 3–6
2007 Carcharoides catticus (Philippi, 1851)—Kocsis; p. 33, figs. 5.1–5.3
2011 Carcharoides catticus (von Philippi, 1846)—Vialle et al.; p. 246, fig. 2.10
2012 Carcharoides catticus (Philippi, 1846)—Otero et al.; p. 188–189, figs. 3:I-K
2013 Carcharoides catticus (Philippi, 1846)—Otero et al.; p. 18, figs. 3:16–21
2014 Carcharoides catticus (Philippi, 1846)—Reinecke et al.; p. 20, Pl. 20, figs. 1–6
Referred material: two teeth (MTM VER 2016.2568., VER 2016.2741.)
The teeth of C. catticus from Máriahalom have a nearly flat labial face, while the lingual face is slightly convex on both specimens. The enamel surface does not bear any striation. The labiolingually flattened, pointed main crown shows smooth cutting edges running from the apex to the root-crown boundary both on the mesial and distal side. The flat, thin root is bifurcated and bears a well-developed central bulge with a transversal groove on the lingual side. The cusplets of the upper laterals are mostly triangular in shape; however, this could vary intraspecifically (see Verwey 2013). C. catticus can be distinguished from Carcharoides totuserratus by its distribution and by its smooth cutting edges (Cappetta 2012).
All features of the upper lateral teeth of C. catticus are clearly visible in MTM VER 2016.2568, and of those of C. catticus anterior files in MTM VER 2016.2741. We assign these teeth to the species C. catticus based on their smooth cutting edges. This species has been reported as “Lamna cattica” from the Rupelian of the Buda Hills (Weiler 1933, 1938; Földváry 1988; Szabó and Kocsis in press), representing one of the oldest records of the species (according to Reinecke et al. 2014). C. catticus has been reported almost exclusively from the lower Oligocene–middle Miocene deposits of western Africa and Europe (Cappetta 1987; Reinecke et al. 2014); however, this species has been reported also from the middle to late Eocene of Chile (Otero et al. 2012, 2013).
Genus Cosmopolitodus Glikman, 1964
Cosmopolitodus sp.
Fig. 7d–f
Referred material: four teeth (MTM VER 2016.2756., VER 2016.2757., VER 2016.2758., VER 2016.2759.)
The teeth are triangular in labiolingual view, with pointed tip and smooth cutting edge both mesially and distally; the tip is pointed. The crown is weakly convex on the lingual side, while flat on the labial side. The surface of the crown is smooth, without any striation-like pattern. The root runs mesiodistally; a nutritive foramen is visible. The teeth of the upper jaw are wider at the base and straighter in mesiodistal view, while the lower teeth are narrower and weakly bent labiolingually (dignathic heterodonty). The upper teeth have weekly bilobate roots, compared to those of lower teeth that are notably bilobate. Upper and lower anteriors are symmetrical, whereas the crowns of anterolateral to distal teeth gradually bend distally (Cappetta 2012).
Several mako and thresher shark species are known from the Oligocene of Europe, for which the characters of the root and the presence or absence of lateral cusplets represent relevant diagnostic features (Cappetta 2012). The extinct species formerly referred to the genus Isurus have been separated into different genera: Cosmopolitodus, Isurus (both Lamnidae) and Anotodus (Alopiidae) (Cappetta 2012). Since the morphology of the crown of the Máriahalom teeth is consistent with that of the European Oligocene-Miocene reports of Cosmopolitodus, they are tentatively referred to Cosmopolitodus.
Familia Otodontidae Glikman 1964
Genus Otodus Agassiz, 1843
Subgenus Otodus (Carcharocles) Jordan and Hannibal, 1923
Otodus (Carcharocles) angustidens (Agassiz, 1843)
Fig. 7g, h
Synonymy
1843 Carcharodon angustidens n. sp.—Agassiz; p. 255, Pl. 28, figs. 20–25 and Pl. 30, fig. 3
1993 Carcharocles angustidens (Agassiz, 1843)—Baut; p. 5, figs. 12–15
1999 Carcharocles angustidens (Agassiz, 1843)—Baut and Génault; p. 25–26, figs. 12–13 and Pl. 4, fig. 11
2001 Carcharodon angustidens (Agassiz, 1843)—Gottfried and Fordyce; p. 731–735, figs. 3A–R
2001 Carcharocles angustidens (Agassiz, 1843)—Reinecke et al.; p. 19–20, Pl. 28–30
2005 Carcharocles angustidens (Agassiz, 1843)—Reinecke et al.; p. 35–36, Pl. 20, figs. 7–9
2014 Otodus (Carcharocles) sp.—Reinecke et al.; p. 23, Pl. 22, fig. 2
Referred material: one tooth (MTM VER 2016.2560)
The teeth of this species are composed of a massive root, an enlarged main crown and two lateral cusplets. The lingual face is convex, while the labial face is typically flat. The cutting edges are irregularly serrated; they extend from the apex to the base of the crown, and sometimes fuse with those of the cusplets with a notch-like boundary. The main crown of the anterior teeth has a high, nearly symmetric, triangular contour, while that of the laterals is strongly bent distally. The distal teeth are the apicobasally lowest teeth in the dentition. The shape of the serrated lateral denticles varies from triangular to semi-circular (see Yabumoto 1987, pl. 4; Gottfried and Fordyce 2001, fig. 4). The root is thick and bifurcated in the anterior to lateral teeth, but less developed and rather bulbous-like in the distal teeth.
Based on the irregularly serrated cutting edge, we refer specimen MTM VER 2016.2560 to O. (Carcharocles) angustidens. Despite its incompleteness, the single collected specimen from Máriahalom exhibits all the typical features of the distal teeth, particularly that of the right lower distal described by Gottfried and Fordyce (2001).
This species has been previously reported from Hungary, from the Kiscell Clay (Rupelian) of the Buda Hills (Hungary) (Weiler 1933, 1938; Földváry 1988; Szabó and Kocsis in press). Otodus (Carcharocles) becomes uncommon in the upper Oligocene. The subgenus includes many species, among them O. (Carcharocles) angustidens, which persists till the base of the Miocene (Cappetta 2012). O. (Carharocles) angustidens is a well-known shark species also documented by associated skeletal remains (Gottfried and Fordyce 2001). This species is known from the Oligocene to the Miocene (Cappetta 2012; Reinecke et al. 2014).
Ordo Carcharhiniformes Compagno, 1973
Familia Carcharhinidae Jordan and Evermann, 1896
Carcharhinidae indet.
Referred material: Five teeth (MTM VER 2016.2594.)
These tooth crowns belong to the family Carcharhinidae, most probably to a species with a close relation to Carcharhinus, because of the simple, pointed crown with nearly flat lingual and smoothly convex labial face. However, due their missing root lobes and enamel shoulders, they cannot be identified at the generic and specific level because of the large variability of the dentition of the carcharhinid genera (see details in Cappetta 2012).
Genus Carcharhinus Blainville, 1826
Carcharhinus elongatus (Leriche, 1910)
Fig. 8a–j

Carcharhinid remains from the Egerian of Máriahalom (Hungary). a Carcharhinus elongatus anterolateral tooth (VER 2016.2603.) in lingual view and b in labial view. c Carcharhinus elongatus anterolateral tooth (VER 2016.2598.) in lingual view and d in labial view. e Carcharhinus elongatus anterior tooth (VER 2016.2600.) in lingual view and f in labial view. g Carcharhinus elongatus anterolateral tooth (VER 2016.2599.) in lingual view and h in labial view. i Carcharhinus elongatus anterolateral tooth (VER 2016.2597.) in lingual view and j in labial view. k Carcharhinus sp. vertebra (VER 2016.2615.) in the view of the articulation surface and l in dorsal view. m Carcharhinus sp. vertebra (VER 2016.2614.) in the view of the articulation surface and n in dorsal view. Scale bars 10 mm
Synonymy
1910 Sphyrna elongata n. sp.—Leriche; p. 300, Pl. 19, figs. 26–30
1993 Carcharhinus elongatus (Leriche, 1910)—Baut; p. 5–6, fig. 29
1999 Carcharhinus elongatus (Leriche, 1910)—Baut and Génault; p. 31–32, Pl. 6, fig. 4
2001 Carcharhinus elongatus (Leriche, 1910)—Reinecke et al.; p. 32–33, Pl. 50–53 (with all figures)
2005 Carcharhinus elongatus (Leriche, 1910)—Reinecke et al.; p. 52–53, Pl. 38–39 (with all figures) and Pl. 44, figs. 1 and 2
Referred material: 31 teeth (MTM VER 2016.2595., VER 2016.2596., VER 2016.2597., VER 2016.2598., VER 2016.2599., VER 2016.2600., VER 2016.2603., VER 2016.2742.)
Since the morphology of the lower teeth is rather conservative within the genus Carcharhinus, the taxonomy at the species level is primarily based on the elements of their upper dentition (see, e.g. Kocsis 2007; Marsili et al. 2007).
All the Máriahalom requiem shark teeth exhibit a convex lingual and a flat labial face and are characterised by a pointed main crown without lateral cusplets. The teeth are mostly wider than high or sometimes their height nearly equals their width (especially in the anterior teeth). The root is projected mesiodistally, and it bears a transversal groove and clearly visible enamel shoulders. The cutting edges are strongly serrated on the enamel shoulders with the serration gradually disappearing towards the tip of the crown. The upper anterior teeth have a narrow, triangular main cusp, whereas the main crown bends distally from lateral teeth to distal teeth. The lower dentition includes teeth with slightly bent, narrow main cusp.
The C. elongatus teeth from Máriahalom are variable both in size and shape and therefore represent both the lower and the upper dentition in several positions. The crowns of the teeth are somewhat wider and more robust than in Carcharhinus gibbesi, which is another common species found in the European Oligocene and thought to be a transitional species towards the common Miocene Carcharhinus priscus (Reinecke et al. 2014). C. elongatus has been mainly reported from Rupelian deposits of north-western Europe (Reinecke et al. 2014).
Carcharhinus sp.
Fig. 8k–n
Referred material: two vertebrae (MTM VER 2016.2614., VER 2016.2615.)
Two shark vertebrae, assigned to Carcharhinus sp., have been collected from the Máriahalom site. They show diagnostic features of the centra of Carcharhinus defined by Kozuch and Fitzgerald (1989). The centra are aseptate, covered by scattered large pores on the sides of the foramina. The dorsal and ventral foramina are oval to circular and do not extend to the rims of the centra. The concave articular surfaces show visual concentric annuli. Both specimens show a cylindrical shape in dorsal view and an overall rounded outline in the view of the articular surface (the latter feature is notably different in the vertebrae of Galeocerdo).
Similar shark centra have been reported by Purdy et al. (2001), and referred to Carcharhinus.
Ordo Myliobatiformes Compagno, 1973
Myliobatiformes indet.
Referred material: 56 teeth (MTM VER 2016.2622., VER 2016.2623., VER 2016.2624., VER 2016.2626., VER 2016.2627., VER 2016. 2628., VER 2016.2629., VER 2016.2630., VER 2016.2631., VER 2016.2634., VER 2016.2636., VER 2016.2637., VER 2016.2640., VER 2016.2642., VER 2016.2643., VER 2016.2644., VER 2016.2645., VER 2016.2646., VER 2016.2647., VER 2016.2650., VER 2016.2651., VER 2016.2652., VER 2016.2653., VER 2016.2657., VER 2016.2658., VER 2016.2660., VER 2016.2661., VER 2016.2662., VER 2016.2663., VER 2016.2664., VER 2016.2667., VER 2016.2669., VER 2016.2670., VER 2016.2671., VER 2016.2672., VER 2016.2674., VER 2016.2675., VER 2016.2677., VER 2016.2678., VER 2016.2682., VER 2016.2684., VER 2016.2687., VER 2016.2688., VER 2016.2689., VER 2016.2690., VER 2016.2691., VER 2016.2692., VER 2016.2693., VER 2016.2694., VER 2016.2697., VER 2016.2699., VER 2016.2700., VER 2016.2701., VER 2016.2702., VER 2016.2703., VER 2016.2704.), 6 tail-spine fragments (VER 2016.2616., VER 2016.2617., VER 2016.2618., VER 2015.2619., VER 2016.2711., VER 2016.2738.).
Due to inadequate preservation, these remains cannot be properly identified and are referred to as indeterminate myliobatiform tooth remains, based on their grinding-type morphology, their hexagonal occlusal face and their polyaulacorhize root (Cappetta 2012). It remains unclear whether these teeth belonged to any of the myliobatiform families described below or some of them represent other taxa.
Familia Myliobatidae Bonaparte, 1838
Genus Aetomylaeus Garman, 1908
Aetomylaeus sp.
Fig. 9a–f

Myliobatiform tooth plates from the Egerian of Máriahalom (Hungary). a Aetomylaeus sp. lower medial tooth (VER 2016.2620.) in occlusal view, b in lingual view and c in basal view. d Aetomylaeus sp. upper medial tooth (VER 2016.2735.) in occlusal view, e in labial view and f in basal view. g Myliobatis sp. medial tooth (VER 2016.2656.) in occlusal view, h in lingual view and i in basal view. j Myliobatis sp. medial tooth (VER 2016.2655.) in occlusal view, k in basal view, l in lingual view and m in cross-sectional view. n Rhinoptera schultzi lateral tooth (VER 2016.2648.) in occlusal view, o in basal view, p in lateral view and q in lingual view. r Rhinoptera studeri medial tooth (VER 2016.2695.) in occlusal view, s in lingual view, t in labial view and u in lateral view. Scale bars 10 mm
Referred material: 11 teeth (MTM VER 2016.2620., VER 2016.2621., VER 2016.2625., VER 2016.2633., VER 2016.2649., VER 2016.2654., VER 2016.2659., VER 2016.2686., VER 2016.2735., VER 2016.2736., VER 2016.2737.)
The genera of Aetomylaeus and Pteromylaeus have been traditionally regarded as separate (e.g. Cappetta 2012); however, a recent systematic analysis suggests that Pteromylaeus is a junior synonym of Aetomylaeus (White 2014). Since their teeth are very similar, the latter hypothesis is followed herein. The dentition of Aetomylaeus is also similar to that of Myliobatis, and significant morphological differences can be better observed in the lower dentitions. Aetomylaeus typically has seven anteroposterior tooth rows: one medial (or central) and three-three laterals on both sides (Cappetta 2012). The medial files are arched (not as in Myliobatis). The root and the crown of the medial files are thicker medially than laterally, and it is oblique in lateral view. The labial side bears a rich ornamentation, which consists of a dense network of ridges and depressions. The lateral series include asymmetrical teeth with polygonal contour (four to six sides) in occlusal view. The backward directed root is not very high; it has oblique labial and lingual faces.
The genus Aetomylaeus is known from the middle Miocene, but its appearance in the fossil record could have been much earlier, since their teeth have been often confused with those of Myliobatis spp. (Cappetta 2012).
Genus Myliobatis Cuvier, 1816
Myliobatis sp.
Fig. 9g–m
Referred material: Nine teeth (MTM VER 2016.2638., VER 2016.2639., VER 2016.2641., VER 2016.2655., VER 2016.2656., VER 2016.2665., VER 2016.2680., VER 2016.2681., VER 2016.2683.)
Many tooth plates of Myliobatis have been discovered in the Máriahalom site. The medial teeth are straight and mediolaterally widened, with angled lateral edges. The lateral teeth are longer than broad, with an occlusally hexagonal contour. Most externals are pentagonal to triangular in occlusal view. The central teeth are only weakly convex in labial view (Cappetta 2012). The root consists of numerous, mediolaterally flattened, parallel lobes. These teeth were distinguished from those of Rhinoptera (see below) by having a relatively lower crown, different root lobe proportions and tooth connection structure.
Remains of the genus Myliobatis are rather common fossils of Cenozoic marine deposits worldwide (Cappetta 2012). Myliobatis has been reported from several Oligocene localities in Europe (see Baut 1993; Baut and Génault 1999; Hovestadt et al. 2010; Reinecke et al. 2001, 2005, 2014).
Familia Rhinopteridae Jordan and Evermann, 1896
Genus Rhinoptera Cuvier, 1829
Rhinoptera cf. schultzi Hiden, 1995
Fig. 9n–q
Synonymy
1995 Rhinoptera schultzi n. sp.—Hiden; p. 76–77., Pl. 9, figs. 1 and 2
2015 Rhinoptera schultzi Hiden, 1995—Reinecke and Radwański; p. 16, Pl. 8, figs. C–G
Referred material: nine teeth (MTM VER 2016.2632., VER 2016.2635., VER 2016.2648., VER 2016.2666., VER 2016.2668., VER 2016.2673., VER 2016.2679., VER 2016.2685., VER 2016.2698.)
Among other characters, teeth of Rhinoptera differ from those of Myliobatis in the absence of a distinct shelf at the crown base and in having more regular hexagonal outline in occlusal view (Reinecke and Radwański 2015).
The Máriahalom teeth assigned to R. cf. schultzi are hexagonal in occlusal view; the occlusal surface is labiolingually flat and shows a weakly granulated ornamentation. The lateral ends are weakly concave. The root is somewhat lower apicobasally than the crown. Due to the preservation, the number of the root lobes cannot be counted (the holotype bears 25 lobes; Hiden 1995). More specimens are necessary to confirm the specific assignment of the material.
R. schultzi differs from Rhinoptera studeri (the other Rhinoptera species unearthed at the Máriahalom site, see below) in having a much thinner crown, smaller width/length ratio and in the absence of ornamentation of the labial and lingual face (Hiden 1995). According to the data of Hiden (1995), R. schultzi has teeth smaller than those of R. studeri (this can be seen in the Máriahalom material as well). The differences between the proportions of the teeth of the two taxa can be easily observed in lateral view (see Fig. 9p, u—and Hiden 1995, text—Fig. 11).
Rhinoptera cf. studeri (Agassiz, 1843)
Fig. 9r–u
Synonymy
1843 Zygobates studeri n. sp.—Agassiz; p. 329, Tab. R, figs. 3–5
1927 Rhinoptera studeri—Leriche; p.42, Taf.6, fig. I-6
1995 Rhinoptera studeri (Agassiz, 1843)—Hiden; p. 77–78., Taf. 3, fig. 9
1970 Rhinoptera cf. studeri—Cappetta; Pl. 24, figs. 3–5
Referred material: Three teeth (MTM VER 2016.2676., VER 2016.2695., VER 2016.2696.)
The labial and lingual sides of the teeth are ornamented with wrinkles. Their crown is high in labiolingual and lateral views, and the lateral edges are angled in occlusal view. The massive and rounded lingual basal ridge is similar to that of R. schultzi. In the lateral teeth, the greatest height of the crown changes from the middle part of the tooth to its lateral margins.
R. studeri has been described from the lower Miocene deposits of Switzerland, France and Austria (Hiden 1995). A further possible occurrence has been mentioned from the upper Oligocene Chandler Bridge Formation from South Carolina, USA (Cicimurri and Knight 2009); however, these teeth have much lower tooth crowns compared to R. studeri. The genus is known since the Paleocene, and it is still living today in tropical seas (Cappetta 2012). The species R. schultzi was described by Hiden (1995) from the Austrian Styrian Basin, where it occurred sympatrically with R. studeri.
Class Osteichthyes sensu Nelson, 2006
Division Percomorphacea Wiley and Johnson, 2010
Order Perciformes Bleeker, 1859
Family Moronidae Jordan and Evermann, 1896
aff. Morone sp.
Fig. 10a

Teleost remains from the Egerian of Máriahalom (Hungary). a aff. Morone sp. saccular otolith (VER 2016.2750.), inner face. b Sparidae gen. et sp. indet. 1, left premaxilla (VER 2016.2728.) in medial view and c left dentary (VER 2016.2729.) in medial view. d Sparidae gen. et sp. indet. 2, right premaxillary (VER 2016.2732.) in medial view. e, f Sciaena sp. saccular otoliths, inner faces (e VER 2016.2747., f VER 2016.2746.). g Sphyraena sp. anterior tooth (VER 2016.2726.). h–k Aglyptorhynchus sp. rostral fragments: h VER 2016.2707. in lateral view, i in occlusal view and j in anterior view; k VER 2016.2745. in posterior view; l Aglyptorhynchus sp. preural vertebra (VER 2016.2739.) in lateral view and m in anterior view. n Trichiuridae gen. et sp. indet. tooth (VER 2016.2722.). Scale bars: a 3 mm; b–d, h, i, l, m 10 mm; e–g, j, k, n 5 mm
Referred material: One saccular otolith (MTM VER 2016.2750).
The otolith is moderately elongate, with a well-developed rostrum and no antirostrum. The ventral margin is regularly curved. The dorsal margin is characterised by a wide predorsal angle and a rounded postdorsal angle. The posterior tip is blunt. The sulcus is supramedian, with the ostium being two times as wide as the cauda. The tip of the cauda is moderately downturned. The dorsal depression is poorly developed.
The overall morphology of the otolith is in many ways reminiscent to that of certain species of the genus Morone (see Schwarzhans 2010; Nolf 2013).
Family Sparidae Bonaparte, 1832
Gen. et sp. indeterminate 1
Fig. 10b, c
Referred material: 25 premaxillae (MTM VER 2016.2728., VER 2016.2734.), 19 dentaries (MTM VER 2729., VER 2016.2730., VER 2016.2764.).
The best preserved specimens consist of a partially complete left premaxilla lacking the distal portion of the ascending process (MTM VER 2016.2728.; Fig. 10b) and a partially complete left dentary (MTM VER 2016.2729., Fig. 10c). Specimen MTM VER 2016.2728. is characterised by a relatively short, laterally expanded and obliquely oriented alveolar process that terminates distally in a pointed tip. The ascending (plus coalesced articular) process is robust and bears a shallow medial crest. Rounded molariform teeth of various sizes are primarily represented by their sockets. A single and very large kidney-shaped tooth is located at the posteromedial corner of the dentigerous portion of the alveolar process.
Like the premaxilla, the dentary (best preserved specimen: MTM VER 2016.2729.; Fig. 10c) is relatively short with a compact dentigerous portion. The dentigerous horizontal arm exhibits a distally curved posterior tip. The dentary teeth are consistent with those of the premaxillary, including the single large kidney-shaped tooth; the latter occupies the posteromedial corner of the dentigerous area.
The overall morphology of the jaw bones as well as the dentition pattern are in many ways reminiscent of those of certain extant and extinct species of the genus Sparus (see, e.g. Jonet 1975; Schultz et al. 2010). However, the incompleteness of the specimens, including the absence of most of the teeth, does not allow a precise taxonomic determination. In any case, as demonstrated by Santini et al. (2014), durophagy appeared multiple times as a result of convergent radiations since the Oligocene, thereby suggesting that the morphology of jaws and teeth cannot be considered as diagnostic without additional osteological information (see also Marsili et al. 2007).
Gen. et sp. indeterminate 2
Fig. 10d
Referred material: One premaxilla (MTM VER 2016.2732.).
The available specimen (MTM VER 2016.2732.; Fig. 10d) consists of a single alveolar process of a right premaxilla bearing a number of sockets of molariform teeth of variable size approximately arranged into three parallel rows. The dentigerous area is well developed. The alveolar process terminates posteriorly in a blunt tip.
As discussed above, the incompleteness of the fossil, including the absence of the teeth, does not allow a precise taxonomic assignment. The specimen is therefore referred to an indeterminate species of the family Sparidae.
Family Sciaenidae Cuvier, 1829
Genus Sciaena Linnaeus, 1758
Sciaena sp.
Fig. 10e, f
Referred material: 22 saccular otoliths (MTM VER 2016.2710., VER 2016.2746., VER 2016.2747., VER 2016.2748.).
The otoliths are compressed with a remarkably blunt anterior margin. The ostium is considerably large, and the cauda is steeply curved. The Oligocene otoliths described herein differ from those of other extant and fossil species of the genus Sciaena in having the tip of the cauda extending ventrally at the same level as the ventralmost margin of the ostium. Moreover, the maximum depth of the otoliths is located at the level of the ostial-caudal interspace or of the downturned portion of the cauda (see Schwarzhans 1993; Nolf 2013).
A single otolith belonging to an indeterminate species of the genus Sciaena has been reported by Nolf and Brzobohaty (1994) from the upper Oligocene Eger Formation.
Family Sphyraenidae Rafinesque, 1815
Genus Sphyraena Röse, 1793
Sphyraena sp.
Fig. 10g
Referred material: 25 isolated teeth (MTM VER 2016.2725., VER 2016.2726., VER 2015.2727.).
The teeth are nearly triangular, laterally compressed and with a distal cutting edge. The crown exhibits discontinuous vertical striae and delicate crenulations. The anterior premaxillary teeth (Fig. 10g) bear a poorly developed postapical barb (see Nishimoto and Ohe 1982).
A large part of the extant barracuda lineages diversified during the Oligocene (Santini et al. 2015). The morphology of the teeth seems to be strongly conservative within the genus Sphyraena. As a consequence, it is not possible to define the affinity of the fossils documented herein at the species level.
Family Palaeorhynchidae Günther, 1880
Genus Aglyptorhynchus Casier, 1966
Aglyptorhynchus sp.
Fig. 10h–m
Referred material: three incomplete rostra, lacking both the proximal and distal portions (MTM VER 2016.2707., VER 2016.2708., VER 2016.2745.) 1 preural vertebra (VER 2016.2739.).
Billfishes of the genus Aglyptorhynchus comprise about ten species with a mobile rostrum that occupied a variety of environmental contexts from the early Eocene to the middle Miocene (e.g. Fierstine 2006). At least six Aglyptorhynchus species are known from the Oligocene of North America and Europe (Fierstine and Weems 2009).
The rostra (fused premaxillae; Fig. 10h–k) contain two pairs of large nutrient canals; the size of the nutrient canals is reduced anteriorly where they are almost fused into a single canal with an irregular profile. In cross section, the rostra are horseshoe shaped both proximally and distally, with a depth/width ratio close to 1. The dorsal and lateral surface of the premaxillae is characterised by a number of longitudinal ridges, separated from each other by narrow sulci. The ventral surface of each premaxilla bears a band of villiform denticles, in many cases solely represented by their sockets; each band comprises four to seven denticles (or their sockets) through its width. A relatively broad and medially convex edentulous area separates the two contralateral dentigerous bands.
The single available caudal vertebra (Fig. 10l, m) is very similar to the fourth preural vertebra described by Fierstine and Weems (2009) from the late Oligocene Chandler Bridge Formation, South Carolina, which was assigned to an indeterminate species of Aglyptorhynchus. The centrum is anteroposteriorly shortened and dorsoventrally compressed. The dorsal and ventral prezygapophyses are massive; the dorsal one considerably is larger. Postzygapophyses are absent. The neural spine is incomplete, but certainly shorter than the haemal spine. The distal tip of the haemal spine is blunt. The outer surface of the vertebrae is finely ornamented by delicate ridges and furrows. Two small circular fossae can be observed along the lateral surface of the vertebral centrum.
The taxonomic status of these Aglyptorhynchus remains is difficult to define. The overall physiognomy of the rostra (particularly the size and morphology of the bands of villiform denticles along the ventral surface of each premaxilla) is very similar to that of the Oligocene species Aglyptorhynchus denticulatus from Belgium (see Leriche 1909; Fierstine 2005; Fierstine and Weems 2009), from which they differ in having four large rather than two small nutrient canals. However, additional comparative information would be necessary to conclusively demonstrate the separate taxonomic status of the specimens documented herein. For this reason, we prefer to postpone the definition of a new taxon until better preserved, and more complete specimens will be available.
Family Trichiuridae Rafinesque, 1815
Gen. et sp. indeterminate
Fig. 10n
Referred material: Three isolated teeth (MTM VER 2016.2721., VER 2016.2722.).
The available material consists of almost straight, elongate, slender and laterally slightly compressed premaxillary fangs with sagittate tips; the postapical barb of these fangs is always well developed.
Elongate fangs with barbs are present in several cutlassfish species of the genera Anenchelum, Eupleurogrammus, Lepidopus, Lepturacanthus and Trichiurus (see, e.g. Nakamura and Parin 1993; Bannikov and Parin 1995; Gago 1997).
Discussion
Based on the available sedimentological (e.g. shallow water, high-energy conditions, possible tidal influence), palaeontological and taphonomical data (e.g. brackish, normal marine and terrestrial animals are found together in the same horizon), the Máriahalom site probably represents a channel fill (Fig. 11). It might have been formed on a tidal flat or a delta plain embayment where the tidal influence was an important factor. This tidally influenced lagoon might have been partially enclosed by barrier islands on its seaward side (Báldi and Cságoly 1975) but might have been connected to a delta on its landward side (based on the statigraphical position and sedimentological data). Interpreting the broader palaeoenvironment is challenging at the moment and requires detailed sedimentological studies.

Palaeoenvironmental reconstruction of the Máriahalom vertebrate site
The Máriahalom locality yielded a moderately diverse, normal marine littoral fish fauna, including eight shark, four batoid and at least seven bony fish species. The shark genera Araloselachus, Carcharhinus, Carcharias, Carcharoides and Otodus (Carcharocles) are medium-sized to large-sized sharks of the nearshore neritic zone (Reinecke et al. 2014). The most common species may have been the odontaspidid A. cuspidatus. Smaller sized predators included C. elongatus which may have been the second most common taxon based on specimen numbers. Some modern representatives of the Carcharhinidae (Carcharhinus leucas, Glyphis spp.) can live in freshwater for extended periods (Compagno 1984; Martin 2005). Based on its body size, O. (Carcharocles) angustidens was most probably the top predator of the Máriahalom ecosystem. Epibenthic feeders, such as rays and angel sharks (Squatina), were diverse and common.
The Máriahalom selachians are typical and common faunal elements in Oligocene marine deposits worldwide (see, e.g. Müller 1983; Woydack 1997; Gottfried and Fordyce 2001; Reinecke et al. 2001, 2005, 2014; Parmley et al. 2003; Fitzgerald 2004). On the other hand, the ray R. schultzi has been so far only known from the Alpine and Carpathian foredeeps (Badenian and Eggenburgian of Austria and Poland respectively), and now from the Central Paratethys (Schultz 2013; Pollerspöck and Straube 2016). The other Rhinoptera species, R. studeri, exhibits a broad distribution during most of the Neogene (Cappetta 2012), but Máriahalom may be the first documented record from the European late Oligocene.
The absence of some typical Oligocene elasmobranch taxa may be related to the depositional environment. For example, the absence of deep-water hexanchids and squalids and of epipelagic and mesopelagic Alopias and Isurus (Cappetta 2012; Reinecke et al. 2014) at Máriahalom is not surprising. On the other hand, the absence of tiger sharks (Physogaleus and Galeocerdo) is possibly related to sampling bias considering the wide distribution and habitat range of these taxa (Compagno 1984). Moreover, screen washing may potentially lead to the discovery of small-sized taxa in the future (e.g. Scyliorhinidae, Dasyatis). It is important to remark that among the recovered fishes, nine taxa (C. gustrowensis, C. catticus, Cosmopolitodus sp., O. (C.) angustidens, Rhinoptera cf. studeri, Sparidae gen et sp. indet. 2, aff. Morone sp., Aglyptorhynchus sp., Trichiuridae gen. et sp. indet.) are represented by less than five specimens each, which suggests that continued excavations may recover additional specimens and/or taxa.
The moderately diverse teleost assemblage is strongly dominated by marine euryhaline taxa of the families Sciaenidae (Sciaena sp.) and Sparidae (Gen. et spp. indeterminate), representing more than three fourths of the bony fish remains. Remains of barracudas, trichiurids, moronids and billfishes of the extinct family Palaeorhynchidae are less common, possibly reflecting an actual minor abundance in the original palaeobiotope.
Sparids are marine euryhaline littoral and inshore fishes that commonly visit brackish biotopes in tropical to temperate contexts worldwide (Nelson 2006). The durophagous members of this clade (similar to the taxa present at Máriahalom) are voracious predators of hard-shelled benthic invertebrates, including crustaceans, molluscs and sea urchins (e.g. Bauchot and Hureau 1986). Modern representatives of the genus Sciaena inhabit marine coastal and brackish biotopes with sandy bottoms, where they feed on small fishes and crustaceans (e.g. Chao 1986). Among barracudas, extant species of the genus Sphyraena are coastal epipelagic piscivorous fishes, primarily living in the water column (e.g. Ben-Tuvia 1986). Trichiurids are marine carnivorous benthopelagic fishes living on the shelf and continental slope where they catch prey by waiting in ambush (Nakamura and Parin 1993). Moronids are restricted to coastal and brackish waters in temperate and cold regions where they feed on a variety of fishes and invertebrates (e.g. Tortonese 1986). As far as the extinct palaeorhynchid genus Aglyptorhinchus is concerned, circumstantial evidences seem to suggest that it was able to tolerate broad salinity variation. Aglyptorhinchus remains have been found in deposits documenting a variety of palaeobiotopes and palaeotemperatures (Fierstine 2006). Late Oligocene Aglyptorhinchus remains from South Carolina have been collected from the Chandler Bridge Formation (see Fierstine and Weems 2009) which originated in a marine coastal environment (Weems and Lewis 2002) affected by periodic salinity variations (see Sanders et al. 1982).
Conclusion
The present contribution is the first study on sharks and rays and the second on bony fishes from the Egerian of the Central Paratethys. The fauna represents a subtropical, nearshore marine community with a great diversity of piscivorous predators and epibenthic feeders. Euryhaline taxa (sparid and sciaenid bony fishes and possibly requiem sharks) were particularly common, and these likely visited the brackish environment indicated by previous studies of molluscs. The small sample size of some taxa indicates that taxonomic diversity of the Máriahalom will likely increase with further fieldwork. The taxonomic composition is typical for other late Oligocene faunas outside the Paratethys and is therefore consistent with an open Paratethys towards the Mediterranean area during the Egerian. The high number of normal marine taxa and individuals contrasts with the predominantly brackish invertebrate fauna of Máriahalom and sheds new light on the palaeoenvironmental reconstruction. Stratigraphical, sedimentological and palaeontological data indicate a tide-influenced and fluvial-influenced nearshore palaeoenvironment consisting of a tidal flat or delta plain with associated brackish lagoons and normal marine littoral habitats. The depositional environment might have been a tidal channel, or a tide-influenced delta channel where fossils of marine, brackish and terrestrial vertebrates and invertebrates were accumulated and preserved together.
References
Agassiz, L. (1833-43). Recherches sur les poissons fossiles. Tome III – Atlas.Neuchâtel 1–432., 1–83 Table.
Antunes, M. T., & Balbino, A. C. (2003). Upper Miocene Lamniform Selachians (Pisces) from the Alvalade Basin (Portugal). Ciências de Terra (UNL), 15, 141–154.
Báldi, T. (1967). A Mány-Zsámbéki-medence felső-oligocén makrofaunája. Földtani Közlöny, 97, 437–446.
Báldi, T. (1973). Mollusc fauna of the Hungarian Upper Oligocene (Egerien) (511 p). Budapest: Akadémiai Kiadó.
Báldi, T. (1980). The early history of the Paratethys. Földtani Közlöny, 110(3–4), 456–472.
Báldi, T. (1983). Magyarországi oligocén és alsómiocén formációk. In P. Polyánszky (Ed.), Akadémia Kiadó, Budapest, 292 p.
Báldi, T. (1989). Tethys and Paratethys through Oligocene times. Remarks to a comment. Geologicky Zborník (Geologica Carpathica), 40(1), 85–99.
Báldi, T., & Cságoly, É. (1975). Faziostratotypus: Máriahalom sand pit. In T. Báldi & J. Seneš (Eds.), OM – Egerien. Die Egerer, Pouzdraner, Puchkirchener Schichtengruppe und die Bretkaer Formation Chronostratigraphie und Neostratotypen (Vol. 5, pp. 134–137). Bratislava: VEDA.
Báldi, T., Less, G., & Mandic, O. (1999). Some new aspects of the lower boundary of the Egerian stage (Oligocene, chonostratigraphic scale of Paratethyan area). Abhandlungen der Geologischen Bundesanstalt, 56, 653–668.
Bannikov, A. F., & Parin, N. V. (1995). Description of a new species of scabbardfish (Aphanopinae: Trichiuridae) from the Middle Eocene of the Northern Caucasus, with restoration of the validity of the fossil genus Anenchelum. Journal of Ichthyology, 35, 183–192.
Bauchot, M.-L., & Hureau, J.-C. (1986). Sparidae. In P. J. P. Whitehead, M.-L. Bauchot, J.-C. Hureau, J. Nielsen, & E. Tortonese (Eds.), Fishes of the North-eastern Atlantic and the Mediterranean (FNAM) (Vol. Vol. II, pp. 883–907). Paris: Unesco.
Baut, J.-P. (1993). Contribution à l’étude des Élasmobranches oligocènes du bassin de Paris. 1: Révision des Élasmobranches du Stampien (Oligocène inférieur) de la région d’Étampes, Essonne, France. Cossmanniana, 2, 1–12.
Baut, J.-P., & Génault, B. (1999). Les Elasmobranches des Sables de Kerniel (Rupélien), à Gellik, Nord Est de la Belgique. Memoirs of the Geological Survey of Belgium, 45, 1–61.
Ben-Tuvia, A. (1986). Siganidae. In P. J. P. Whitehead, M.-L. Bauchot, J.-C. Hureau, J. Nielsen, & E. Tortonese (Eds.), Fishes of the north-eastern Atlantic and the Mediterranean (Vol. Volume 2, pp. 964–966). Paris: UNESCO.
Böhm, B. (1942). Adatok a magyarországi harmadkori halfaunához. Geologica Hungarica Series Paleontologica, 16, 6–42.
Brzobohatý, R., & Schultz, O. (1978). Die Fischfauna des Badenien. In J. Seneš (Ed.), Chronostratigraphie und Neostratotypen, Miozän der Zentralen Paratethys. Bd. 4, M4 Badenien (pp. 441–465). Bratislava: Verlag der Slowakischen Akademie der Wissenschaften.
Cappetta, H. (1970). Les Sélaciens du Miocéne de la région de Montpellier. Palaeovertebrata, Mémorie extraordinaire, 139 p., 27 pl.
Cappetta, H. (1987). Handbook of Paleoichthyology, Vol.3B: Chondrichthyes II. Stuttgart: Gustav Fischer Verlag 193 p.
Cappetta, H. (2012). Handbook of Paleoichthyology, Vol.3E: Chondrichthyes II – Mesozoic and Cenozoic Elasmobranchii: Teeth. München: Verlag Dr. Friedrich Pfeil 512 p.
Casier, E. (1966). Fauna ichthyologique du London Clay. British Museum Natural History London, 1966, 1–496.
Chao, L. N. (1986). A synopsis on zoogeography of the Sciaenidae. In T. Uyeno, R. Arai, T. Taniuchi, & K. Matsuura (Eds.), Indo-Pacific fish biology: Proc. Second Int’l Conf. Indo-Pacific Fishes (pp. 570–589). Tokyo: Ichthyological Society of Japan July-August 1985.
Cicimurri, D. J., & Knight, J. L. (2009). Late Oligocene sharks and rays from the Chandler Bridge Formation, Dorchester County, South Carolina, USA. Acta Palaeontologica Polonica, 54(4), 627–647.
Compagno, L. J. V. (1984). FAO species catalogue. Vol 4: Sharks of the world. FAO Fisheries Synopsis No. 125, 4, 1–633.
Fierstine, H. L. (2005). A new Aglyptorhynchus (Perciformes: Scombroidei) from the Lincoln Creek Formation (Late Oligocene, Washington, U.S.A.) Journal of Vertebrate Paleontology, 25, 288–299.
Fierstine, H. L. (2006). Fossil history of billfishes (Xiphioidei). Bulletin of Marine Science, 79, 433–453.
Fierstine, H. L., & Weems, R. E. (2009). Paleontology of the Oligocene Ashley and Chandler Bridge formations of South Carolina, 5: analysis and new records of billfishes (Perciformes: Xiphioidei). Palaeo Ichthyologica, 11, 43–88.
Fitzgerald, E. M. G. (2004). A review of the tertiary fossil Cetacea (Mammalia) localities in Australia. Memoirs of the Museum of Victoria, 61(2), 183–208.
Földváry, G. Z. (1988). Geology of the Carpathian region. Singapore: World Scientific Pub.Co 571 p.
Gago, F. J. (1997). Osteology and phylogeny of the cutlassfishes (Scombroidei: Trichiuridae). Contributions in Science, Natural History Museum of Los Angeles County, 476, 1–79.
Glikman, L. S. (1964). Sharks of Paleogene and their stratigraphic significance. Moscou: Nauka Press 229 p, (in Russian).
Gottfried, M. D., & Fordyce, R. D. (2001). An associated specimen of Carcharodon angustidens (Chondrichthyes, Lamnidae) from the Late Oligocene of New Zealand, with comments on Carcharodon interrelationships. Journal of Vertebrate Paleontology, 21(4), 730–739.
Günther, A. (1880). An introduction to the study of fishes (720 pp). New Delhi: Today & Tomorrow’s Book Agency.
Harzhauser, M., & Piller, W. E. (2007). Benchmark data of a changing sea—palaeogeography, palaeobiogeography and events in the Central Paratethys during the Miocene. Palaeogeography, Palaeoclimatology, Palaeoecology, 253, 8–31.
Haye, T., Reinecke, T., Gürs, K., & Piehl, A. (2008). Die Elasmobranchier des Neochattiums (Oberoligozän) von Johannistal, Ostholstein, und Ergänzungen zu deren Vorkommen in der Ratzeburg-Formation (Neochattium) des südöstlichen Nordseebeckens. Palaeontos, 14, 55–97.
Hiden, H. R. (1995). Elasmobranchier (Pisces, Chondrichthyes) aus dem Badenium (Mittleres Miozän) des Steirischen Beckens (Österreich). Mitteilungen der Abteilung für Geologie und Paläontologie am Landesmuseum Joanneum, 52(53), 41–110.
Holec, P. (2001). Miocénne drsnokozce a kostnaté ryby (Chondrichthyes et Osteichthyes, Vertebrata), z viedenskej panvy pri Bratislave (Slovensko). Mineralia Slovaka, 33, 111–134.
Holec, P., Hornácek, M., & Sykora, M. (1995). Lower Miocene shark (Chondrichthyes, Elasmobranchii) and whale faunas (Mammalia, Cetacea) near Mucin, Southern Slovakia. Geologické práce, 100, 37–52.
Hovestadt, D. C., & Hovestadt-Euler, M. (2010). A partial skeleton of Carcharias gustrowensis (Winkler, 1875) (Chondrichthyes, Odontaspididae) including embryos, a chimaeriod dorsal fin spine and a myliobatoid tailspine from the Oligocene of Germany. Cainozoic Research, 7, 83–97.
Hovestadt, D. C., Hovestadt-Euler, M., & Micklich, N. (2010). A review of the chondrichthyan fauna of Grube Unterfeld (Frauenweiler) clay pit. Kaupia, Darmstädter Beiträge zur Naturgeschichte, 17, 57–71.
Janssen, A. W. (1982). Late Oligocene molluscs from a sand-pit near Máriahalom (Hungary). A preliminary study. Annales Universitates Scientarium Budapestiensis de Rolando Eötvös Nominate, Sectio Geologica, 14, 109–150.
Jonet, S. (1975). Notes d’Ichthyologie Miocene Portugaise. VI – Les Sparidae. Boletim da Sociedade Geológica de Portugal, 19, 135–172.
Kázmér, M., Dunkl, I., Frisch, W., Kuhlemann, J., & Ozsvárt, P. (2003). The Paleogene forearc basin of the Eastern Alps and Western Carpatians: subduction erosion and basin evolution. Journal of the Geological Society, 160, 413–428.
Koch, A. (1903). Tarnócz Nógrád megyében.Mint kövült czápafogaknak új gazdag lelőhelye. Földtani Közlöny, 33, 22–44.
Kocsis, L. (2007). Central Paratethyan shark fauna (Ipolytarnóc, Hungary). Geologica Carpathica, 58(1), 27–40.
Korpás, L. (1981). A Dunántúli-középhegység oligocén – alsó-miocén képződményei. (Oligocene – Lower Miocene Formations of the Transdanubian Central Mountains in Hungary). Magyar Állami Földtani Intézet Évkönyve 1981, 64(1), 1–80 (in Hungarian).
Kozuch, L., & Fitzgerald, C. (1989). A guide to identifying shark centra from southeastern archaeological sites. Southeastern Archaeology, 8(2), 146–157.
Leriche, M. (1909). Note préliminaire sur des poisons nouveaux de l’Oligocène belge. Bulletin de la Société Belge de Géologie, de Paléontologie et d’Hydrologie, procès-verbaux, 22, 378–384.
Leriche, M. (1910). Les poissons tertiaires de la Belgique. III. Les poissons oligocénes. Mémoriesdu Musée Royal d’Historie Naturelle de Belgique, 5(2), 229–363.
Leriche, M. (1927). Les Poissons de la Molasse suisse. Mémoires de la Société Paléontologique Suisse, 46, 1–55.
Less, Gy., Frijia, G., Filipescu, S., Holcová, K., Madic, O., & Sztanó, O. (2015). New Sr-isotope stratigraphy (SIS) age-data from the Central Paratethys. 2nd International congress on Stratigraphy. Abstract volume, 223.
Lihoreau, F., Blondel, C., Barry, J., & Brunet, M. (2004). A new species of the genus Microbunodon (Anthracotheriidae, Artiodactyla) from the Miocene of Pakistan: genus revision, phylogenetic relationships and palaeobiography. Zoologica Scripta, 33, 97–115.
Marsili, S., Carnevale, G., Danese, E., Bianucci, G., & Landini, W. (2007). Early Miocene vertebrates from Montagna della Maiella, Italy. Annales de Paléontologie, 93, 27–66.
Martin, R. A. (2005). Conservation of freshwater and euryhaline elasmobranchs: a review. Journal of the Marine Biological Association of the United Kingdom, 85, 1049–1073.
Müller, A. (1983). Fauna und Palökologie des marinen Mitteloligozäns der Leipziger Tieflandsbucht (Böhlener Schichten). Altenburger Naturwissenschaftliche Forschungen, 2, 1–152.
Müller, A. (1996). Die Ichthyofauna des Oberoligozäns der Hessischen Senke (Raum Kassel, Deutschland). Leipziger Geowissenschaften, 2, 31–115.
Nagymarosy, A. (1990). Paleogeographical and paleotectonical outlines of some intra-Carpatian Paleogene basins. Geologický zborník (Geologica Carpathica), 41, 259–274.
Nagymarosy, A. (2012). Paratethys evolution and its consequences for the Paleogene-Neogene Chronostratigrapic Framework. In J. Haas (Ed.), Geology of Hungary (pp. 81–99), Springer.
Nagymarosy, A., & Gyalog, L. (1997). Mány Formation. In L. Császár (Ed.), Basic Litostratigraphic Units of Hungary (pp. 14–15). Budapest: The Geological Institute of Hungary.
Nagymarosy, A., & Sztanó, O. (2014). Oligocene lithostratigraphic units. Unpublished report of the Oligocene Committee on Stratigraphy, p.12.
Nakamura, I., & Parin, N. V. (1993). Snake mackerels and cutlassfishes of the world (families Gempylidae and Trichiuridae). FAO Fisheries Synopsis, 125(15), 1–136.
Nelson, S. J. (2006). Fishes of the world (4th ed.). New York: John Wiley & Sons, Inc 601 p.
Nishimoto, H., & Ohe, F. (1982). Teeth of fossil Sphyraena of the Miocene Mizunami Group, central Japan. Bulletin of the Mizunami Fossil Museum, 9, 85–102.
Nolf, D. (2013). The diversity of fish otoliths, past and present. Bruxelles: Royal Belgian Institute of Natural Sciences 222 p.
Nolf, D., & Brzobohaty, R. (1994). Fish otoliths from the Late Oligocene (Eger and Kiscell Formations) in the Eger area (northeastern Hungary). Bulletin de l’Institut Royal des Sciences Naturelles de Belgique, Sciences de la Terre, 64, 225–252.
Otero, R. A., Torres, T., Le Roux, J. P., Hervé, F., Fanning, C. M., Yury-Yáñez, R. E., & Rubilar-Rogers, D. (2012). A Late Eocene age proposal for the Loreto Formation (Brunswick Peninsula, southernmost Chile), based on fossil cartilaginous fishes, paleobotany and radiometric evidence. Andean Geology, 39(1), 180–200.
Otero, R. A., Oyarzún, J. L., Soto-Acuña, S., Yury-Yáñez, R. E., Gutierrez, N. M., Le Roux, J. P., Torres, T., & Hervé, F. (2013). Neoselachians and Chimaeriformes (Chondrichthyes) from the latest Cretaceous-Paleogene of Sierra Baguales, southernmost Chile. Chronostratigraphic, paleobiogeographic and paleoenvironmental implications. Journal of South American Earth Sciences, 48, 13–30.
Parmley, D., Cicimurri, D. J., & Campbell, B. (2003). Late Eocene sharks of the Hardie Mine local fauna of Wilkinson County, Georgia. Georgia Journal of Science, 61(3), 153–179.
Philippi, R. A. (1846). Tornatella abbreviata, Otodus mitis, Otodus catticus, und Myliobatis testae. Palaeontographica, 1, 23–25.
Piller, W. E., Harzhauser, M., & Mandic, O. (2007). Miocene Central Paratethys statigraphy—current status and future directions. Stat, 4, 151–168.
Pollerspöck, J., & Straube, N. (2016). www.shark-references-com, World Wide Web electronic publication, Version 2016.
Popov, S. V., Rögl, F., Rozanov, A. Y., Steininger, F. F., Shcherba, I. G., & Kovác, M. (2004). Lithological-paleogeographic maps of Paratethys, 10 maps Late Eocene to Pliocene. Courier Forschungsinstitut Senckenberg, 250, 1–46.
Probst, J. (1879). Beiträge zur Kenntniss der fossilen Fische aus der Molasse von Baltringen. Hayfische. Jahreshefte des Vereins für vaterländische Naturkunde in Württemberg, 35, 127–191.
Purdy, R. W., Schneider, V. P., Applegate, S. P., McClellan, J. H., Meyer, R. L., & Slaughter, B. H. (2001). The Neogene sharks, rays, and bony fishes from the Lee Creek Mine, Aurora, North Carolina. In C. E. Ray, D. J. Bohaska (Eds.), Geology and Paleontology of the Lee Creek Mine, North Carolina (pp. 71–202). III, Smithsonian Contributions to Paleobiology, No. 90.
Rabi, M., & Botfalvai, G. (2008). A preliminary report of the Late Oligocene vertebrae fauna from Máriahalom, Hungary. Hantkeniana, 6, 177–185.
Reinecke, T., & Radwański, A. (2015). Fossil sharks and batoids from the Korytnica-clays, early Badenian (Langhian, Middle Miocene), Fore-Carpathian basin, central Poland—a revision and updated record. Palaeontos, 28, 5–37.
Reinecke, T., Stapf, H., & Raisch, H. (2001). Die Selachier und Chimären des Unteren Meeressandes und Schleichsandes im Mainzer Becken (Alzey- und Stadecken-Formation, Rupelium, Unteres Oligozän). Palaeontos, 1, 1–73.
Reinecke, T., Moths, H., Grant, A., & Breitkreutz, H. (2005). Die Elasmobranchier des norddeutschen Chattiums, insbesondere des Sternberger Gesteins (Eochattium, Oligozän). Palaeontos, 8, 1–135.
Reinecke, T., Balsberger, M., Beaury, B., & Pollerspöck, J. (2014). The elasmobranch fauna of the Thalberg beds, early Egerian (Chattian, Oligocene), in the Subalpine Molasse Basin near Siegsdorf, Bavaria, Germany. Palaeontos, 26, 3–129.
Rocabert, L. (1934). Contribucó al coneixement de la fauna ictiológica terciària catalana. Butlleti Institució catalana Història Natural, 34, 78–107.
Rögl, F. (1998). Paleogeographic consideration for Mediterranean and Paratethys seaways (Oligocene to Miocene). Annalen des Naturhistorischen Museums in Wien, 99A, 279–310.
Rögl, F., & Steininger, F. F. (1984). Neogene Paratethys, Mediterranean and Indo-Pacific seaways. Implications for the paleobiogeography of marine and terrestrial biotas. In P. Brenchley (Ed.), Fossils and Climate (pp. 171–200). Chichester: John Wiley & Sons Ltd..
Russell, D. E., Hartenberger, J., Pomerol, C., Sen, S., Schmidt-Kittler, N., & Vianey-Liaud, M. (1982). Mammals and stratigraphy: the paleogene of Europe. Palaeovertebrata, Mémoir extraordinaire, 1–77.
Sanders, A. E., Weems, R. E., & Lemon Jr., E. M. (1982). Chandler Bridge Formation—a new Oligocene stratigraphic unit in the lower coastal plain of South Carolina. United States Geological Survey, 1529-H, 105–124.
Santini, F., Carnevale, G., & Sorenson, L. (2014). First multi-locus timetree of seabrems and porgies (Percomorpha: Sparidae). Italian Journal of Zoology, 81, 55–71.
Santini, F., Carnevale, G., & Sorenson, L. (2015). First timetree of Sphyraenidae (Percomorpha) reveals a Middle Eocene crown age and an Oligo-Miocene radiation of barracudas. Italian Journal of Zoology, 82, 133–142.
Scherler, L., Mennecart, B., Hiard, F., & Becker, D. (2013). Evolutionary history of hoofed mammals during the Oligocene-Miocene transition in Western Europe. Swiss Journal of Geosciences, 106, 349–369.
Schultz, O. (2013). Pisces. In W. Piller (Ed.), Catalogus Fossilium Austriae, Bd. 3 (576 pp.). Wien: Verlag der Österreichischen Akademie der Wissenschaften (ISBN 978–3–7001-7238-3).
Schultz, O., Brzobohatý, R., & Kroupa, O. (2010). Fish teeth from the Middle Miocene of Kienberg at Mikulov, Czech Republic, Vienna Basin. Annalen des Naturhistorisches Musem Wien, 112, 489–506.
Schwarzhans, W. (1993). A comparative morphological treatise or recent and fossil otoliths of the family Sciaenidae (Perciformes). Piscium Catalogus: Part Otolithi Piscium, 1, 1–245.
Schwarzhans, W. (2010). The otoliths from the Miocene of the North Sea Basin (352 pp). Leiden: Backhuys Publishers.
Solt, P. (1988). Odontaspis (Synodontaspis) divergens n. sp. a csillaghegyi oligocénből. A Magyar Állami Földtani Intézet Évi Jelentése az 1986-os évről, 519–533.
Szabó, M., & Kocsis, L. (in press). A preliminary report on the early Oligocene (Rupelian, Kiscellian) selachians from the Kiscell Formation (Buda Mts, Hungary), with the re-discovery of Wilhelm Weiler’s shark teeth. Fragmenta Palaeontologica Hungarica, 33, 31–64.
Sztanó, O., Magyari, Á., & Nagymarosy, A. (1998). High-resolution stratigraphy in the Esztergom Basin, northeastern Transdanubia, Hungary: II. Oligocene sequences and their interpretation. Földtani Közlöny, 128(2–3), 455–486 (in Hungarian with English abstract).
Telegdi-Roth, K. (1927). Spuren einer infraoligozanen Denudation am nordwestlichen Range des Transdanubischen Mittelgebirge. Földtani Közlöny, 57, 117–128.
Tortonese, E. (1986). Moronidae. In P. J. P. Whitehead, M.-L. Bauchot, J.-C. Hureau, J. Nielsen, & E. Tortonese (Eds.), Fishes of the north-eastern Atlantic and the Mediterranean (Vol. Volume 2, pp. 793–796). Paris: UNESCO.
Verwey, G. (2013). Carcharoides uit het Churchilldok, Antwerpen. Afzettingen WTKG, 34(2), 26–29.
Vialle, N., Adnet, S., & Cappetta, H. (2011). A new shark and ray fauna from the Middle Miocene of Mazan, Vaucluse (southern France) and its importance in interpreting the paleoenvironment of marine deposits in the southern Rhodanian Basin. Swiss Journal of Palaeontology, 130(2), 241–258.
Weems, R. E., & Lewis, W. C. (2002). Structural and tectonic setting of the Charleston, South Carolina, region: evidence from the tertiary stratigraphic record. Geological Society of America Bulletin, 114, 24–42.
Weiler, W. (1933). Zwei oligozäne Fischfaunen aus dem Königreich Ungarn. Geologica Hungarica, Series Palaeontologica, 11, 1–54.
Weiler, W. (1938). Neue Untersuchungen an mitteloligozänen Fischen Ungarns. Geologica Hungarica, Series Palaeontologica, 15, 1–31.
White, W. T. (2014). A revised generic arrangement for the eagle ray family Myliobatidae, with definitions for the valid genera. Zootaxa, 3860(2), 149–166.
Winkler, T. C. (1875). Beschreibung einiger fossiler Tertiaer-Fischreste, vorzugsweise des Sternberger Gesteins. Archiv des Vereins der Freunde der Naturgeschichte in Mecklenburg, 29, 97–129.
Woydack, A. (1997). Die Ichthyofauna der Tagebaue Cospuden und Espenhain (Unteroligozän der Leipziger Bucht). Leipziger Geowissenschaften, 4, 165–187.
Yabumoto, Y. (1987). Oligocene Lamnid shark of the genus Carcharodon from Kitakyushu, Japan. Bulletin of the Kitakyushu Museum of Natural History, 6, 239–264.
Acknowledgements
We thank the two reviewers, Alexandre F. Bannikov (Borisyak Paleontological Institute, Russian Academy of Sciences, Moscow) and James C. Tyler (National Museum of Natural History, Smithsonian Institution, Washington), for their constructive comments and suggestions that greatly improved the manuscript. We want to thank all the numerous fieldwork participants who assisted MR and GB between 1999 and 2006. Botond Kemény and Ágnes Görög (Eötvös Loránd University, Budapest) are particularly thanked for their assistance in fieldwork and discussions. Excavations were supported by grants from the Pro Renovanda Cultura Hungariae Program and the Hantken Miksa Foundation. We are indebted to the staff of the Department of Palaeontology and the Department of Physical and Applied Geology of the Eötvös Loránd University for their support and help provided through the years. Marian Bosnakoff (Natural History Museum, Hungary), Balázs Szinger (MOL Hungarian Gas and Oil Company, Budapest), András Uhrin (Eriksfiord, Stavanger, Norway), Attila Ősi (Eötvös Loránd University, Budapest) and Thomas Reinecke (Bochum, Germany) are acknowledged for insightful discussions and sharing literature. This project has received funding from the European Union’s Seventh Framework programme for research and innovation under the Marie Skłodowska-Curie grant agreement no. 609402-2020 researchers: Train to Move (T2M) to MR. The research of GC was supported by grants (ex-60% 2015 and 2016) of the Università degli Studi di Torino.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article

Cite this article
Szabó, M., Botfalvai, G., Kocsis, L. et al. Upper Oligocene marine fishes from nearshore deposits of the Central Paratethys (Máriahalom, Hungary). Palaeobio Palaeoenv 97, 747–771 (2017). https://doi.org/10.1007/s12549-017-0285-0
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s12549-017-0285-0