
Abstract
Morphological analyses of hipparionine astragali, calcaneum, third phalanx and metapodials from the late Miocene locality of Maragheh, Iran, was carried out using bivariate plots and log10 ratio diagrams. The results, together with previous studies on cranial and dental material, have allowed us to characterise and define the following equid species in the Maragheh assemblage: Hipparion gettyi, aff. Hippotherium brachypus, Cremohipparion aff. moldavicum, Cremohipparion matthewi and Hipparion campbelli. These species are arrayed in three successive biostratigraphic intervals: H. gettyi, late Vallesian Lower Maragheh horizon; aff. H. brachypus and Cr. aff. moldavicum, early Turolian Middle Maragheh levels; and H. campbelli, late early Turolian Upper Maragheh interval. The small hipparion, Cr. matthewi, would appear to range throughout all biostratigraphic intervals of Maragheh. The proposed taxonomic and biostratigraphic resolution here is slightly different from previous studies especially in the recognition of aff. H. brachypus instead of Hipparion prostylum. The Maragheh hipparion assemblages are well correlated to those from Turkey, Greece, the Balkans and Black Sea region and are clearly different from those of Central and Western Europe, central Asia, China and Africa. Maragheh Cremohipparion is related to species of Cremohipparion from Greece, China and the Siwaliks. A clear niche differentiation based on palaeodiet studies has been revealed in Maragheh hipparions, indicative of different environmental adaptations amongst these species.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
History of investigations at Maragheh
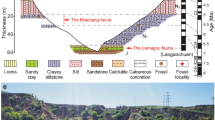
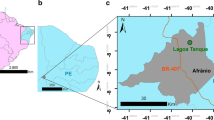
The fossil localities of Maragheh are located in the eastern Azarbaijan Province, northwest Iran. The main fossiliferous layers are located near the villages of Kopran, Mordagh (Mirduq), Karajabad (Kherjabad), Shalilvand (Sholl’avand) and Ilkhchi, between 37° 20′–37° 25′ N latitude and 46° 16′–46° 37′ E longitude. However, fossil sites exist within the present extension of Maragheh city (e.g. south of Jahangir) and also towards its west (e.g. Ahaq) (Fig. 1). The Maragheh fauna, along with Samos and Pikermi sites in Greece, has long been considered one of the three most preeminent Western Eurasian late Miocene Pikermian faunas. As with Pikermi and Samos, Maragheh is a true “Lagerstätte” because of the shear abundance and diversity of its fauna across a great geographic extent, which provides a virtually endless supply of well-preserved fossil specimens. Maragheh is also unique amongst the three classical Pikermian faunas in its clear stratigraphic display and layer-cake stratigraphy with several, laterally continuous pumices and volcanic ashes that are readily suitable for stratigraphic and geochronologic correlation (Mirzaie Ataabadi et al. 2013).

Geographic location of fossil localities in the Maragheh Basin, N.W. Iran. 1 Kopran localities, 2 Varjoy localities, 3 Aliabad localities, 4 Mordagh (Mirduq, Mordaq) localities, 5 Dareh Gorg (Gort Daresi) localities (including new MMTT and INSPE localities), 6 Karajabad (Kherjabad) localities, 7 Sumu Daresi locality, 8 E. Mordagh localities, 9 Shalilvand (Sholl’avand) localities, 10 Ghartavol localities, 11 N. E. Shalilvand (Sholl’avand) localities, 12 Khermejavand locality, 13 Ilkhchi localities and 14 Ahagh (Ahaga), W. Maragheh localities. |--A - H---| corresponds to the position of stratigraphic columns (modified after Mirzaie Ataabadi et al. 2013)
Fossil bones of Maragheh have apparently been known by local people for a long time. They were considered the remains of pre-flood animals or those of mysterious/monstrous creatures. Khanikoff, a Russian explorer, is credited for the first collection of Maragheh fossils in 1840. However, the Maragheh fauna was not initially studied before the latter half of the nineteenth century (Abich 1858; Brandt 1870; Grewingk 1881). As these early works provided data on Maragheh’s similarity to Pikermi, attention of additional scientists was attracted to this site. The Austrian palaeontologist Pohlig made the first comprehensive collection and geological study of Maragheh (Pohlig 1886). He was apparently invited by a local merchant to visit this locality in 1884. His trip was facilitated by Austrians who had major positions in the government of Iran at that time. Pohlig explored the Maragheh basin extensively and sampled fossils from most of the Maragheh section. He preserved locality information for much of its collection, now stored in the Naturhistorisches Museum, Wien. This extraordinary data in an early collection facilitates understanding of stratigraphic provenance of those fossils. Rodler and Kittl, two other Austrian palaeontologists, visited Maragheh and made an extensive collection of fossils, which were later published by Kittl (1887), Rodler (1890), Rodler and Weithofer (1890) and Schlesinger (1917). Damon acquired a small collection of Maragheh fossils for the British Museum of Natural History, London, which was briefly communicated by Lydekker in 1886.
In 1897, the French palaeontologist Marcellin Boule secured permission through the de Morgan agreement with the Iranian government to conduct a palaeontological expedition to Maragheh. This French expedition to Maragheh in 1904 was organised at a grand scale for this time in palaeontology. French palaeontologists assisted by 12 local workmen excavated a large sample of Maragheh fossils from different localities. This massive collection, apparently weighing about 20 tons, was transported from Maragheh to Istanbul on camels and then shipped by boat to France, eventually being deposited in the Museum National d’Histoire Naturelle, Paris, where they reside today. These fossils were first studied preliminarily and later published in the form of short summary papers and eventually a comprehensive monograph (Mecquenem 1924–25).
More than 50 years elapsed before other reported expeditions to Maragheh occurred. Professor Takai of Tokyo University collected a small sample of Maragheh fossils (Takai 1958) and Robert Savage of Bristol University also visited Maragheh in the early 1960s and collected a significant number of fossils. Tobien from the Johannes Gutenberg University, Mainz, accompanied by geologists from the geological survey of Iran and National Iranian Oil Company made important excavations in the middle portion of the Maragheh sequence in the late 1960s (Tobien 1968). During the 1970s, three scientific groups conducted research at Maragheh: a combined Dutch-German group led by Erdbrink and assisted by University of Tehran staff (Erdbrink et al. 1976), a joint University of Kyoto-Geological Survey of Iran led by Kamei (Kamei et al. 1977) and the Lake Rezaiyeh Expedition (LRE) led by Professor Campbell (Campbell et al. 1980) and supported by the Iranian National Museum of Natural History (MMTT), Tehran and the LSB Leakey Foundation, Berkeley, California.
Bernor was responsible for the study of vertebrate fauna for the LRE, which resulted in his PhD (1978) and several manuscripts on the faunal composition and their biostratigraphical and zoogeographic relationships. The fieldwork undertaken in 1974, 1975 and 1976 by LRE had three important outcomes: (a) collection of fossils with attention to their stratigraphic provenance (which was usually neglected by old collectors)—this led to the establishment of the first biostratigraphy of the Maragheh fauna; (b) study of old collections to better understand the taxonomy and diversity of the mammalian fauna; and (c) application of various geochronologic tools to obtain well-resolved ages for the Maragheh section and its faunas. The latest was a single crystal argon study of the LRE volcanic ash samples by Swisher (1996).
After a few decades of inactivity in the Maragheh basin, Iran’s Department of Environment (DOE) and Iranian National Museum of Natural History (MMTT) initiated and sponsored new excavations in the area, which resulted in the nomination of 10 km2 of the Maragheh fossiliferous area as a national protected zone and in the establishment of a field museum and research station in the Dareh Gorg (Gort Daresi) area. Recently, the MMTT-University of Helsinki-Japanese initiative known as the International Sahand Paleoenvironment Expedition (INSPE) was also initiated (Mirzaie Ataabadi et al. 2013 and this issue’s papers). This programme undertook field activities between 2007 and 2009, discovering several new localities and fossils as well as investigating several key geological horizons (Sawada et al. 2016, this issue) and sedimentological and palaeoenvironmental study of the main fossiliferous area in Dareh Gorg (Sakai et al. 2016, this issue). This programme has further reinitiated the study of the mammalian fauna with the intention of bringing them into a contemporary taxonomic context for comparative palaeoecological and palaeobiogeographic studies (present work and Solounias and Danowitz 2016, this issue).
History of investigations on the Hipparion fauna
Mecquenem (1908) described the Paris collection of the Maragheh fauna, including the hipparionine horses and referred them to the species Hipparion gracile. Gabunia (1959) described a new species from Maragheh, Hipparion urmiense, based on material in Georgian National Museum, Tbilisi. The nomen urmiense is derived from Lake Urmia which is located immediately to the west of the town of Maragheh. These fossils were apparently found in the Maragheh fossil horizons near the village Kurtevul (Ghartavul), but no further geographic or stratigraphic documentation is available (Bukhsianidze 2015). Forsten (1968) proposed the existence of two major groups of hipparionine horses in the Palaeartic region, including Maragheh.
Woodburne and Bernor (1980) studied the superspecific groupings of Old World hipparionine horses. They presented four supraspecific groups of hipparionine horses based on the preorbital fossa (POF) morphology, its depth and its location in the facial region. They identified all four groups in the Maragheh area, confirming that there is a well-diversified hipparion fauna at Maragheh. Bernor et al. (1980) also used these data to study the chronology of Maragheh and some other Old World Miocene faunas. Forsten (1983) also studied the POF and its significance as a taxonomic character in the study of Old World hipparions. She used preorbital bar length/P2-orbit distance diagrams to discriminate hipparion species in Maragheh as well as some other Greek localities. These species conform to Woodburne and Bernor (1980), who recognised lineages with long preorbital bars, groups 1 and 3, versus a short preorbital bar, groups 2 and 4. Bernor et al. (1996) subsequently identified the following lineages: group 1 = Hippotherium, groups 2 and 4 = Cremohipparion (larger versus smaller taxa) and group 3 = Hipparion s.s.
Bernor (1985) studied the systematics of Maragheh hipparionine horses based on old museum specimens and the material excavated by LRE. He described two new species from this region, one from the lower horizons of Maragheh (Hipparion gettyi) and one from its upper levels (Hipparion campbelli). Watabe and Nakaya (1991a) studied the phylogenetic significance of the postcranial hipparionine material from Maragheh. They distinguished three morphotypes in the material based on the size and proportion of third metapodials which corresponded to the three skull types present in the collection of Maragheh in Kyoto. They also studied the cranial skeleton of hipparions from Maragheh (Watabe and Nakaya 1991b) and recognised four species, including H. gettyi, Hipparion prostylum, Hipparion moldavicum and H. urmiense. Watabe and Nakaya (1991b) believed that their H. urmiense is the senior synonym of H. campbelli; however, Bernor has noted that the type of H. urmiense does not have an intact premaxilla and that the premaxilla discriminates between Samos Hipparion dietrichi and Maragheh H. campbelli and is morphologically critical at the species level. Moreover, the Middle Maragheh H. “prostylum” was also believed to be a member of Hipparion s.s. and is likewise morphologically similar to H. urmiense. We show herein that this is not the case: Middle Maragheh hipparion is not a member of Hipparion s.s. Therefore, the nomen H. urmiense is a problematic taxon: there is no stratigraphic provenance for the type and the type is insufficient to recognise a species or the lineage that the species belongs to on its own.
Systematic conventions
The nomen Hipparion has been used in a variety of ways by different authors. We follow characterisations and definitions for hipparionine horses recently provided in Bernor et al. (1996, 1997). Hipparion monographs by Gromova (1952) and Gabunia (1959) are cited after the French and English translations. The taxon Hipparion has been applied in a variety of ways by different authors. We utilise the following definitions in this work:
-
Hipparionini—a tribe of Equidae with an isolated protocone on maxillary premolar and molar teeth and, as far as known, tridactyl feet, including species of the following genera: Cormohipparion, Neohipparion, Nannippus, Pseudhipparion, Hippotherium, Cremohipparion, Hipparion, Sivalhippus, Eurygnathohippus (= a senior synonym of Stylohipparion), Proboscidipparion and Plesiohipparion. These lineages have recently been reviewed by Qiu et al. (1987), Bernor and White (2009), Bernor et al. (2010, 2013, 2014), Armour-Chelu and Bernor (2011), Wolf et al. (2013) and Bernor and Sun (2015).
-
Hipparion s.s.—the name is restricted to a specific lineage of hipparionine horses with the facial fossa positioned dorsally high on the face (MacFadden 1980, 1984; Woodburne and Bernor 1980; Woodburne et al. 1981; MacFadden and Woodburne 1982; Bernor 1985; Bernor and Hussain 1985; Bernor et al. 1987, 1989; Woodburne 1989 and the authors of this contribution). The posterior pocket becomes reduced and eventually lost and confluent with the adjacent facial surface (includes group 3 of Woodburne and Bernor 1980). Bernor’s definition departs from some investigators in not recognising North American species of Hipparion s.s. Bernor (1985) and Bernor (in Bernor et al. 1989) have argued that any morphologic similarity between North American “Hipparion” and Old World Hipparion s.s. is due to homoplasy. This issue is found to be even more complex in our current analysis of the Maragheh hipparions in that there has, to date, been an inadequate accounting of the genotype species, H. prostylum de Christol postcranial size and proportions. We find here that previous referrals of Middle Maragheh H. prostylum to be questionable.
-
“Hipparion”—several distinct and separate lineages of Old World hipparionine horses once considered to be referable to the genus Hipparion (re: Bernor et al. 2011). We emphasise here the need to avoid confusion of well-defined hipparionine lineages with poorly characterised taxa of “Hipparion” sensu lato.
-
Hippotherium—a discrete genus of Western Eurasian hipparionine horses known from Central Europe, Italy, Greece, Turkey, Bulgaria, Macedonia, Ukraine and Iran. Species belonging to this genus include Hippotherium primigenium, Hippotherium intrans, Hippotherium microdon, Hippotherium kammerschmittae, Hippotherium malpassi, Hippotherium brachypus and perhaps Hippotherium giganteum (re: Bernor et al. 2011).
Metric procedures
Measurements are all given in millimetres and rounded to 0.1 mm. Measurement numbers (M1, M2, M3, etc.) refer to those published by Eisenmann et al. (1988) and Bernor et al. (1997) for the skulls and postcrania. Tooth measurement numbers refer to those published by Bernor et al. (1997) and Bernor and Harris (2003).
Bernor and Armour-Chelu (1999), Bernor and Harris (2003), Bernor and Scott (2003), Bernor et al. (2004, 2005, 2010, 2013), Gilbert and Bernor (2008), Bernor and Haile Selassie (2009) and Bernor and White (2009) have compared African hipparions to an extensive series of late Miocene-Pleistocene Eurasian and African assemblages. Bernor and Sun (2015) have recently reviewed cheek tooth ontological stages in Chinese Plesiohipparion and Proboscidipparion.
In various studies, Eisenmann (see Eisenmann 1995 for a comprehensive summary) has used log10 ratio diagrams to evaluate differences in hipparion metapodial proportions as a basis for recognising taxa and their evolutionary relationships. Bernor et al. (2003), Bernor and Harris (2003) and Armour-Chelu and Bernor (2011) have used multiple statistical tests, including univariate, bivariate and multivariate statistics as well as log10 ratio diagrams, to evaluate and resolve the alpha systematics of hipparionine horses. Bernor et al. (2005) used log10 ratio diagrams together with multivariate statistics to evaluate metapodial and first phalangeal evidence for postcranial evolution in Ethiopian hipparions. We incorporate these previously used methodologies in this work. Our statistical analysis uses the skeletal population from Höwenegg (Hegau, southern Germany, 10.3 Ma; Bernor et al. 1997) for calculating 95 % confidence ellipses used in bivariate plots, and log10 mean standard values for all log10 ratio diagrams (MPIIIs).
Abbreviations
We use the following abbreviations in this manuscript:
Ma: mega-annum in the geochronologic time scale. Ages in m.y. are based on radioisotopic analyses or magnetostratigraphic analyses. North Africa: localities may be referred to the MN biochronologic time scale. Western Eurasia: Vallesian, Turolian and Ruscinian; intervals of the European land mammal age sequence, commonly termed units (sensu Fahlbusch 1991).
Measurement table abbreviations—sex: M = male; F = female; ? = unknown.
Sex can be defined by the size of a canine tooth, male being large, female being small.
Side: lt. = left; rt. = right.
Cranial abbreviations: IOF = infraorbital foramen; POB = preorbital bar; POF = preorbital fossa.
Element abbreviations: CALC = calcaneum; AST = astragalus; MCIII = metacarpal III; MTIII = metatarsal III; MPIII = metapodial III; 1PHIII = first phalanx III (central digit) of either the anterior or posterior limb, which are difficult to distinguish in hipparion; tx = maxillary tooth; tm = mandibular tooth; mand = mandible.
M1-M38 refers to measurements as described by Eisenmann et al. (1988) and Bernor et al. (1997). Statistical plots include abbreviations by locality. These abbreviations are as follows:
- A:
-
USA
- C:
-
China—Plesiohipparion or Proboscidipparion
- c:
-
China—Chinese taxa other than Plesiohipparion or Proboscidipparion
- D:
-
Abu Dhabi
- E:
-
Eppelsheim, Germany
- G:
-
Langebaanweg, South Africa
- H:
-
Höwenegg, Germany
- K:
-
Pikermi, Greece
- L:
-
Çalta, Turkey
- O:
-
Samos, Greece
- M:
-
Maragheh, Iran
- P:
-
Potwar Plateau, Pakistan
- Q:
-
Saloniki, Greece
- S:
-
Sahabi, Libya
- T:
-
Sinap and Esme Akçakoy, Turkey
- U:
-
Mt. Luberon, France
- Z:
-
Akkaşdagi, Turkey
Museum collections included
We sample the following museum and institutional vertebrate collections for this study:
- AMNH:
-
American Museum of Natural History, New York
- BMNH:
-
Natural History Museum, London
- BSPG:
-
Bayerische Staatssammlung für Paläontologie und Geologie, Munich (formerly Tobien collection, University of Mainz)
- GIU (now IvAU):
-
Geological Institute, Utrecht (now Instituut voor Aardwetenschappen Utrecht)
- MMTT:
-
Muze Melli Tarikh Tabeie, Tehran
- MNHN:
-
Museum National d’Histoire Naturelle, Paris
- NHMW:
-
Naturhistorisches Museum, Wien
Maragheh bivariate plot symbols
-
1.
MMTT collection, circle (O)
-
2.
MNHN collection, X
-
3.
NHMW collection, plus sign (+)
-
4.
BSPG collection, upright triangle (▲)
-
5.
AMNH, upside down triangle (▼)
-
6.
BMNH, sideways triangle, apex to the left (◄)
-
7.
GIU, sideways triangle, apex to the right (►)
Statistical analysis
ESM Table 1 provides measurements of the Maragheh specimens used in this analysis. These measurements were taken by R.L. Bernor, M. Mirzaie Ataabadi, K. Meshida, M. Watabe and D. Wolf. Comparative measurements were taken from Bernor’s unpublished equid database. Analyses of the Maragheh hipparions include bivariate plots of CALC, AST, MCIII, MTIII and 1PHIII. All of these skeletal elements are clearly differentiated with the exception of 1PHIII. While anterior and posterior 1PHIII can be clearly differentiated in living Equus, the same cannot be said for all hipparions (see, e.g. Wolf et al. 2013). The Höwenegg hipparion skeletons were found in articulation and thus the anterior versus the posterior 1PHIII are known. However, the statistical differences between these phalanges are minor at best in the Höwenegg sample, which has led us to analyse all 1PHIII together. Some advanced African hipparions may in fact differ significantly in anterior versus posterior 1PHIII dimensions (Bernor et al. 2010).
Bivariate plots
Bivariate plots will include three parallel sets of plots: one of our entire analytical samples; a second of the Maragheh specimens plotted indicating the seven different assemblage samples (AMNH, BMNH, BSPG, GIU, MMTT, MNHN and NHMW); and a third showing an analysis of Pikermi, Samos, Saloniki and Mt. Luberon. These three-partite statistical analyses are intended to reveal which Maragheh taxa occur in which museum collection, in which stratigraphic intervals and between Maragheh and other closely relevant late Miocene localities. Furthermore, this analysis is intended to provide new insights into the occurrence through time and space of hipparion superspecific lineages at Maragheh and other Pikermian age localities.
We analyse CALC, AST, MCIII, MTIII and 1PHIII below. In each plot, a 95 % ellipse representing the Höwenegg population standard of H. primigenium is used as the species standard. The array of comparative Hipparion measurements is heuristic for demonstrating the range of variation in size and proportion of the element/sample combination being plotted.
We plot CALC maximum length (M1) versus distal maximum width (M6) herein (Fig. 2a–c):

Bivariate plot of CALC maximum length (M1) versus distal maximum width (M6) comparing a all studied specimens, b Maragheh specimens and c Pikermian specimens
Figure 2a includes four Eppelsheim Hippotherium, seven Pakistan Sivalhippus, six Maragheh Hippotherium, two Pikermi H. brachypus and one Sahabi Sivalhippus specimen within the ellipse or on the lower border of the ellipse. There are two large Sivalhippus Pakistan specimens and one large Samos H. aff. brachypus specimens above the ellipse. The smallest specimens are two from Samos and two from Maragheh: these are referable to Cremohipparion matthewi. There are a great number of specimens that plot from below the ellipse including Maragheh (M), Pakistan (P), Sahabi (S), Langebaanweg (G), Mt. Luberon (U), Samos (O) and Sinap (T). These are likely a mix of taxa including Hipparion s.s. (Samos, Mt. Luberon and Maragheh), Cremohipparion (Samos, Maragheh and possibly Mt. Luberon), Eurygnathohippus hooijeri (Langebaanweg) and primitive “Hipparion” spp. (Sinap).
Figure 2b includes four MNHN Maragheh specimens that plot in the ellipse and several just below. MMTT has one specimen just below, three specimens well below the ellipse and two far below the ellipse. The two MMTT specimens furthest below the ellipse are referable to Cr. matthewi and the larger four below the ellipse are likely H. campbelli. There are six NHMW specimens that plot close to the upper four MMTT Maragheh specimens that are plausibly related to H. campbelli.
Figure 2c includes two Pikermi (K) specimens that are found within the ellipse, best referable to H. brachypus. One Samos specimen (O) lies above the ellipse, probably representing the large representative of the H. brachypus lineage from Samos. There is an extensive cluster of 10 Mt. Luberon (U) and 3 Saloniki (Q) specimens below the ellipse best referred to H. prostylum (Hipparion s.s.). There is a single specimen from Pikermi (K) in this cluster that may either be Cremohipparion mediterraneum or H. cf. prostylum. There are two very small Saloniki specimens (Q) that are the size of Maragheh Cr. matthewi and this may be a valid referral.
We next plotted AST maximum length (M1) versus distal articular width (M5) (Fig. 3a–c):

Bivariate plot of AST maximum length (M1) versus distal articular width (M5) comparing a all studied specimens, b Maragheh specimens and c Pikermian specimens
Figure 3a plots the staggering number of astragali in our sample and most measurements fall along a regression line through the Höwenegg ellipse, suggesting that these measurements reflect the size (= body weight) of the animal. The largest specimens are from Pakistan (P) Sivalhippus spp. and the smallest specimens from Maragheh (M) and Saloniki (Q) Cr. matthewi.
Figure 3b exhibits a great dispersion of points. Three MMTT Maragheh specimens plot within the lower portion of the ellipse. There are likewise a large number of specimens from MNHN and NHMW within the ellipse and these are referable to the Middle Maragheh robust-limbed taxon aff. Hippotherium sp. The vast majority of MMTT specimens plot below the ellipse, although there are also some to the left of the ellipse, as well. There are two very small MMTT specimens that plot with a single AMHN specimen referable to Cr. matthewi. The majority of specimens from MMTT, MNHN and NHMW below the ellipse are likely referable to either Cremohipparion moldavicum and/or H. campbelli.
Figure 3c has seven Pikermi (K) astragali specimens that plot within the Höwenegg ellipse and three Mt. Luberon specimens that plot at the bottom of the ellipse. The Pikermi specimens are referable to H. brachypus, as are potentially those specimens from Mt. Luberon. There are three Pikermi specimens below the ellipse and a number of Mt. Luberon and Saloniki specimens below the ellipse. This cluster below the ellipse varies considerably in length (M1) versus width (M5) and represents a great deal of size variability. Some may be due to ontogeny (juvenile astragali, which are smaller but cannot be easily distinguished), but some variability may also be due to actual species diversity. Mt. Luberon (U) and Saloniki (Q) likely have H. prostylum, but Cr. matthewi might also be represented by the smallest one or two specimens.
We plotted MCIII maximum length (M1) versus distal maximum width (M11) herein (Fig. 4a–c):

Bivariate plot of MCIII maximum length (M1) versus distal maximum width (M11) comparing a all studied specimens, b Maragheh specimens and c Pikermian specimens
Figure 4a demonstrates the extensive array of MCIII dimensions above, below and to the left of the Höwenegg ellipse. There are also a few large specimens to the right of the ellipse. The largest specimen is of Chinese Plesiohipparion (C) that plots well above all other specimens. The specimens with the greatest width are from Samos and likely represent a large member of the H. brachypus lineage. The smallest specimen is from China (c), and the narrowest specimens are from Samos (O) and Maragheh (M) and are likely referable to Cr. matthewi (sensu lato). There are many specimens from Maragheh (M), Pikermi (K), Samos (O), Pakistan (P) and Sinap (T) that plot within the Höwenegg ellipse; M, K and O are referable to Hippotherium, while P is referable to Sivalhippus and T is “Hipparion” sp.
Figure 4b plots Maragheh MCIII specimens from five museum and institutional collections. There is a single MMTT 13 specimen, referable to H. campbelli that plots to the left of the Höwenegg ellipse. This specimen is in the middle of the range of a series of MNHN specimens that are referable to Cr. moldavicum (larger, longer specimens) and perhaps Cr. matthewi (smaller, shorter specimens). There are likewise small and narrow specimens from the AMNH (upside down triangles) that are also best referred to the small taxon Cr. matthewi. There are a number of specimens that plot within the Höwenegg ellipse, mostly from the MNHN collection, but also from the NHMW and the BSPG collection (triangle). The MNHN and BSPG collections have good skull material that was previously referred (MNHN) to H. prostylum. This allocation is now rejected because of the associated robust limbs that are directly comparable to Höwenegg Hippotherium, suggesting a possible referral to that genus. We explore this further in the “Systematics” section below.
Figure 4c compares previous MCIII plots with our sample from Samos, Pikermi, Mt. Luberon and Saloniki. There are more than a dozen Pikermi specimens that plot within the Höwenegg ellipse, and these specimens are referable to H. brachypus. Samos has eight specimens that plot within or just outside the top of the ellipse and two large specimens to the right of the top of the ellipse that are all likely referable to the large member of the H. brachypus clade. Pikermi has specimens that plot to the left of the ellipse that are likely referable to Cr. mediterraneum. Samos has a number of specimens that plot along the axis of Pikermi Cr. mediterraneum but include individuals that are longer and shorter. There is a constellation of Samos taxa featured here that could include H. dietrichi, Cremohipparion proboscideum, Cr. aff. mediterraneum, Cr. aff. matthewi and Cremohipparion nikosi. The small Mt. Luberon sample casts some light on the taxonomy, having one specimen that plots on the lower left corner of the ellipse and two specimens that plot to the left of the lower left corner of the ellipse. These Mt. Luberon specimens are best referred to H. prostylum s.s. and are somewhat shorter than MMTT 13 H. campbelli.
We plot MTIII maximum length (M1) versus distal maximum width (M11) herein (Fig. 5a–c):

Bivariate plot of MTIII maximum length (M1) versus distal maximum width (M11) comparing a all studied specimens, b Maragheh specimens and c Pikermian specimens
Figure 5a includes a massive amount of MTIII data in our sample. The longest MTIII is of Plesiohipparion from China. Other than Chinese Plesiohipparion, the longest specimens are from Samos and are likely members of the Cr. proboscideum clade. The widest specimens are also from Samos and Pakistan and likely belong to advanced H. brachypus and Sivalhippus, respectively. The smallest specimens are from Samos and Maragheh and Pakistan; Samos and Maragheh are likely referable to Cr. matthewi. The very small Pakistan specimens are of unknown taxonomic affinity.
Figure 5b plots at least 20 MNHN Maragheh specimens within the Höwenegg ellipse which are referable to a species of aff. Hippotherium. There is a single specimen from the AMNH that plots just outside the upper border of the ellipse that is most similar to this MNHN sample. There is an extensive array of MNHN specimens to the left of the Höwenegg ellipse and subdivided into an upper, larger group and a smaller, lower group. It cannot be absolutely determined with this simple analysis if two taxa are represented by this extended “slender” cluster, but it is possible: the specimens in the larger cluster may be Cr. moldavicum while those in the smaller cluster may be Cr. matthewi. There is a single AMNH specimen that stands out as being the smallest individual well below the two MNHN “slender clusters” which should be referable to Cr. matthewi. The MMTT sample is elongate and slender and groups to the left and in between the two slender MNHN clusters; these MMTT specimens are referable to H. campbelli.
Figure 5c plots approximately 20 specimens from Pikermi within the Höwenegg ellipse referable to aff. H. brachypus. Pikermi also has a number of slender specimens that plot to the left of the Höwenegg ellipse referable to Cr. mediterraneum. Mt Luberon includes five specimens at the lower limit and just outside the lower left corner of the Höwenegg ellipse and another five specimens well to the left of the ellipse comparable in proportions to the lower slender cluster of the MNHN Maragheh sample. The more slender specimens from Mt. Luberon are likely referable to H. prostylum s.s. and are comparable in their proportions to Maragheh H. campbelli. There are two specimens from Saloniki that plot below the Maragheh and Mt. Luberon sample which could be members of the Hipparion s.s. clade based on Saloniki’s skulls, resembling H. prostylum and H. campbelli in their facial morphology and dimensions.
Log10 ratio diagrams (Figs. 7 and 8) comparing MCIII and MTIII of these Saloniki and Mt. Luberon specimens with Maragheh H. campbelli specimens show clearly a great similarity amongst these elements. Maragheh H. campbelli compares very well with H. prostylum from Mt. Luberon and Salonique Hipparion sp.? All elements show the sharp contrast of mid-shaft width (M3) versus depth (M4) proportions, revealing their slender-elongate metapodials and their possible convergence with Cremohipparion in this feature.
Samos has the greatest dispersion of plotted points of any of these localities. Most specimens are along the slender axis being very elongate and narrow to very short and narrow, and these specimens are probably referable to H. dietrichi and multiple species of Cremohipparion (Cr. proboscideum, Cr. aff. mediterraneum and Cr. nikosi). There are also Samos specimens that plot within the Höwenegg ellipse that could be related to Hippotherium. There are very large Samos specimens to the right of the Höwenegg ellipse, larger than all Pikermi specimens that are likely referable to a robust member of the H. brachypus clade.
We plotted 1PHIII maximum length (M1) versus proximal articular width (M4) herein (Fig. 6a–c):

Bivariate plot of 1PHIII maximum length (M1) versus proximal articular width (M4) comparing a all studied specimens, b Maragheh specimens and c Pikermian specimens
Figure 6a includes a large number of 1PHIII specimens in our database. There are many specimens from our localities within, below and to the left of our ellipse. The longest specimens are from Pakistan and are referable to Cremohipparion antelopinum (sensu Wolf et al. 2013). Saloniki also has very long and relatively slender 1PHIIIs that are referable to Hipparion s.s. The smallest two specimens are from Samos and Mt. Luberon and likely referable to Cr. matthewi.
Figure 6b exhibits a great range of variability in the Maragheh 1PHIII sample. The MNHN and NHMW have a great number of specimens within the Höwenegg ellipse that are possibly related to Hippotherium. There are a number of specimens below and to the left of the ellipse from the MNHN, NHMW and AMNH collections. The MMTT sample includes specimens that are elongate and slender placed to the left of the middle to upper portion of the ellipse that are referable to H. campbelli and other smaller specimens that are possibly evolutionarily related to H. campbelli. There are two very small 1PHIII specimens from the MNHN sample, and one from the NHMW sample that are referable to Cr. matthewi.
Figure 6c includes one Samos 1PHIII specimen plotting within and two plotting outside the upper left border of the ellipse. There are two Saloniki specimens that are very long compared to the rest of the sample and may be members of the Hipparion s.s. clade. Mt. Luberon (U) has specimens at the bottom and just below the Höwenegg ellipse as well as five specimens well below the ellipse in a cluster with Samos and two Saloniki specimens. There is probably a mix of Hipparion s.s. and Cremohipparion taxa represented by this cluster.
At the lowest part of the cluster, there is one small specimen from Samos and another one from Mt. Luberon probably related to Cr. matthewi.
Log 10 ratio analyses—MCIIIs
Figures 7a–c and 8a
Figure 7a plots Maragheh Hippotherium specimens that fall within the Höwenegg ellipse for the maximum length (M1) versus distal articular width (M11) dimensions. There are two log10 ratio patterns herein. The first one plots very closely to Pikermi H. brachypus (re: solid black line). Both the length and flat mid-shaft width (M3) and depth (M4) trajectories as well as the proximal and distal articular proportions are similar. The second pattern is more similar to Sinap Cormohipparion sinapensis (broken black line) and contrasts with Hippotherium in having a relatively slender mid-shaft width (M3) with corresponding deep mid-shaft depth (M4) dimensions. This second group includes individuals with some of the smallest dimensions.

Log10 ratio diagrams of MCIIIs comparing Maragheh a aff. H. brachypus, b Cr. aff. moldavicum and c Cr. matthewi with Co. sinapensis and Cr. mediterraneum means

Log10 ratio diagrams of MCIIIs a comparing Maragheh H. campbelli to Mt. Luberon and Saloniki hipparions and MTIIIs comparing Maragheh b aff. H. brachypus to EsmeA_Mean, Pikermi_Mean and AMNH27813 and c Cr. aff. moldavicum, with Co. sinapensis, H. brachypus and Cr. mediterraneum means
Figure 7b plots Maragheh MNHN Cremohipparion aff. moldavicum specimens in comparison to Sinap Cormohipparion sinapensis (broken black line) and Pikermi Cr. mediterraneum (solid black line). All specimens except MNHN Mar. 39/648 compare closely with primitive Co. sinapensis. The MEAN Pikermi Cr. mediterraneum has a similar plot projection as Maragheh Cr. aff. moldavicum but is slightly smaller in proximal and distal articular proportions. Overall, it is of considerable interest how close Cremohipparion is to Sinap Cormohipparion, its plausible ancestor.
Figure 7c plots MNHN Cr. matthewi specimens compared to Co. sinapensis (solid black line) and Cr. mediterraneum (broken black line) means. These FAM (AMNH) and MNHN specimens are smaller than Cr. moldavicum and range from being slightly to much smaller in size compared to Sinap Co. sinapensis and Pikermi Cr. mediterraneum. Maragheh Cr. matthewi exhibits very sharply contrasting mid-shaft width (M3) versus depth (M4) proportions revealing that these were very slender-elongate cursorial small horses (re: Bernor et al. 2003).
Figure 8a plots Maragheh MMTT 13 H. campbelli and MNHN H. aff. campbelli specimens as well as Mt. Luberon H. prostylum and Salonique Hipparion sp. compared to Sinap Co. sinapensis (solid black line) and Pikermi Cr. mediterraneum (dashed black line) means. These specimens are smaller than Co. sinapensis and mostly also smaller than Cr. mediterraneum. Maragheh MMTT 13/2552 H. campbelli (solid black line with ×) tracks closely with H. prostylum from Mt. Luberon and Saloniki. All specimens exhibit very sharply contrasting mid-shaft width (M3) versus depth (M4) proportions, revealing that these horses had very slender-elongate metacarpals. The extreme slenderness with striking M3 versus M4 dimensions are apparently convergent characters shared between the Cremohipparion and Hipparion s.s. lineages.
Log10 ratio analyses—MTIIIs
Figures 8b, c and 9a–c
Figure 8b is an MTIII log10 ratio diagram comparing the Maragheh aff. H. brachypus hypodigm to Esme Akçakoy Co. sinapensis (solid black line) and Pikermi MEAN log10 values of H. brachypus (broken black line) (Koufos 1987). The Esme Akçakoy sample tracks relatively closely to specimens with narrow mid-shaft width (M3) and moderately deep mid-shaft depth (M4) dimensions and has amongst the smallest distal supraarticular width (M10), articular width (M11), sagittal keel (M12) and condylar depth (M13 and M14) dimensions. Pikermi H. brachypus plots with the larger Maragheh specimens, featuring, like another single specimen, MNHN Mar. 35/661, the widest mid-shaft width (M3) dimensions of the sample. H. brachypus also has elevated proximal articular depth (M6) dimensions only exceeded by AMNH 27813, the longest specimen in our sample. While these Maragheh specimens’ maximum length and distal width dimensions fall within the Höwenegg ellipse, there would appear to be two groups represented in this log10 ratio plot: two specimens that have distinctly wider and deeper mid-shaft dimensions (AMNH 27813 and MNHN Mar. 34/660) and the majority of other specimens that have relatively more slender mid-shaft width (M3) and depth (M4) dimensions than the Höwenegg sample. All Maragheh specimens, except MNHN Mar. 3125, exhibit an enhanced “Esme Akçakoy” effect (Bernor et al. 2003) in that there is a striking contrast between the relatively narrow (M3) and deep (M4) mid-shaft dimensions for MTIII. This plot illustrates the possibility that there may be more than one taxon represented in this sample of MTIIIs.

Log10 ratio diagrams of MTIIIs comparing Maragheh a Cr. sp., b Cr. matthewi and c H. campbelli with Co. sinapensis, H. brachypus and PIK87 Cr. mediterraneum means
Figure 8c is a log10 ratio diagram comparing the Maragheh Cr. aff. moldavicum hypodigm to Esme Akçakoy Co. sinapensis (solid black line) and Pikermi (broken black line) MEAN log10 values of Cr. mediterraneum (Koufos 1987). Cr. aff. moldavicum clearly has more elongate (M1) and slender mid-shaft (M3) specimens than the Höwenegg sample. Maragheh Cr. aff. moldavicum has the most specimens with M1 dimensions greater than in Pikermi Cr. mediterraneum but is very similar in its mid-shaft width (M3) versus depth (M4) proportions to Pikermi Cr. mediterraneum. Cr. mediterraneum plots at the bottom of the Maragheh Cr. aff. moldavicum range for maximum length (M1) and proximal (M5, M6) and distal articular (M10, M11, M12, M13 and M14) dimensions. Esme Akçakoy Co. sinapensis plots generally within the middle of the Maragheh Cr. aff. moldavicum plots but is distinguished by having less of a contrast in mid-shaft width (M3) versus depth (M4) dimensions and it is shorter in its maximum length than the entire Cr. aff. moldavicum sample.
Figure 9a is an MTIII log10 ratio diagram comparing the Maragheh sample of MNHN Cremohipparion sp. specimens to Esme Akçakoy Co. sinapensis (solid black line) and Pikermi (broken black line) mean log10 values of Cr. mediterraneum (Koufos 1987). These specimens mostly have the length of the Esme Akçakoy Co. sinapensis sample but have the striking contrast between mid-shaft width (M3) versus depth (M4). At the same time, this sample exhibits size and proportions similar to Pikermi Cr. mediterraneum. There is no evidence from the total Maragheh cranial material that Cr. mediterraneum occurred at Maragheh. We refer this sample then to Cr. aff. moldavicum.
Figure 9b is an MTIII log10 ratio diagram comparing the Maragheh Cr. aff. matthewi hypodigm to Esme Akçakoy Co. sinapensis (solid black line) and Pikermi (broken black line) mean log10 values of Cr. mediterraneum (Koufos 1987). These specimens have the length of Pikermi Cr. mediterraneum but are much more slender in their build, again showing the sharp contrast in mid-shaft width (M3) versus mid-shaft depth (M4) measurements typical of Cremohipparion and in particular the Cr. aff. moldavicum group.
Figure 9c is an MTIII log10 ratio diagram comparing Maragheh MMTT 13 H. campbelli specimens (MMTT 13/1153 and MMTT 13/1622) and MNHN Mt. Luberon H. prostylum and Saloniki H. cf. prostylum to Co. sinapensis (solid black line) and Pikermi Cr. mediterraneum means (broken black line). The MMTT, Mt. Luberon and Saloniki specimens are closely similar to one another and at the same time are more slenderly constructed than Esme Akçakoy Co. sinapensis. The Maragheh, Mt. Luberon and Saloniki specimens exhibit a sharper contrast in mid-shaft width versus depth than Pikermi Cr. mediterraneum and are for the most part more slenderly built than Cr. mediterraneum. This analysis supports the assertion that Maragheh H. campbelli is a clade in common with the genotype species of H. prostylum s.s. from Mt. Luberon and Saloniki (re: Woodburne and Bernor 1980).
Systematics
Order Perissodactyla Owen, 1848
Suborder Hippomorpha Wood, 1937
Family Equidae Gray, 1821
Tribe Hipparionini Quinn, 1955
“Hipparion” gettyi Bernor, 1985
Hipparion sp. 3 Bernor et al. (1979b), p. 94, fig. 1; Campbell et al. (1980), p. 841
Hipparion sp. (Kopran horse) Woodburne et al. (1981), p. 1343, fig. 5d; Bernor et al. (1980), p. 723, fig. 8; Bernor and Hussain (1985), p. 36, fig. 3d
Hipparion gettyi Bernor 1985, pp. 194–198, fig. 10
Holotype: A skull NHMW 2015/0238/0001 (KNHM-RLB 8401 of Bernor 1985), preserved by the Naturhistorisches Museum, Vienna.
Type locality: Kopran, Maragheh.
Age: Circa 9–8.7 Ma, latest Vallesian, MN10.
Referred specimens: Besides the type specimen, we recognise KNHM no number, an old adult palate with right P3-M3 and left P3-M3; KNHM A4866, young adult female mandible with i1-m3; right i1-p2 (I3’s just erupting); MMTT 41/2224, lt. p2; MMTT 41/2464, lt. M1; MMTT 41/2211, lt. P4; MMTT 41/2220, lt. M2; MMTT 41/2248, lt. M1; MMTT 41/2519, lt. p4; MMTT 41/2234, lt. p4; MMTT 36/2022, rt. P2; MMTT 41/2213, rt. dP3 or 4; MMTT 41/2216, rt. dP3 or 4; MMTT 36/2027, lt. p2; 41/2225, rt. m2; 43/2489, rt. m1; 43/2484, lt. m1.
Diagnosis (modified from Bernor 1985 ): A medium-sized species of “Hipparion” with a long POB; lacrimal does not invade POF; POF is anteroposteriorly long, egg-shaped and anteroposteriorly oriented, dorsoventrally deep, with strong posterior pocketing, medially deep with a faint anterior rim and moderately expressed peripheral outline. Nasal notch is placed above the P2 parastyle. Adult cheek teeth have complex, moderately thickly banded plications of the pre- and postfossettes; pli caballins are persistently double; hypoglyphs are deeply incised; protocones are moderately elongate; P2 anterostyle is elongate. An uncertainly associated MPIII from Kopran is slender and elongate.
Description: Bernor (1985) named and described the Vienna and MMTT material referred to H. gettyi which we briefly summarise herein. The type specimen is a skull, NHMW 2015/0238/0001 (Fig. 10a–c). This specimen is of a moderately large-sized old “Hipparion” with a small canine, indicating that it was female. The skull is virtually complete except for the posterior orbits and posterior and inferior cranium (Fig. 10a). The snout is moderately long, the nasals are shallowly retracted to the P2 parastyle. The POB is long with the distal most lacrimal being well posterior to the POF. The POF is relatively large, elongate, egg-shaped, anteroposteriorly directed, has a moderately well-delineated peripheral rim with a weakly expressed anterior rim, is medially deep, is deeply pocketed posteriorly and placed well above the facial-maxillary crest. In dorsal view (Fig. 10b), the skull reveals that it is narrow and the POF is medially deep. The ventral view (Fig. 10c) reveals that the type skull has all teeth preserved and at least one half worn; this clearly is an older individual. The incisors have well-defined infundibula that are irregularly shaped and fill the central portion of the occlusal surface. Both canines are complete and small, indicating that this individual is a female. P2 has a moderately elongate anterostyle. All cheek teeth have moderately complex fossette plications, pli caballins are consistently double, hypoglyphs are moderately deep and protocones are oval-shaped (Fig. 10c).

The type specimen of H. gettyi in a lateral, b dorsal and c ventral views
Bernor (1985) described an unnumbered NHMW fragmentary maxilla of an old individual from Kopran, Maragheh, which has very worn cheek teeth that are not easily referred to any species. Also, there is a young adult female mandible in the NHMW collection from Kopran with the following salient characters: elongate anterostylid, metaconid and metastylid subequal in size and with rounded to slight angular shape, pli caballinids absent, linguaflexids shallow on all teeth.
The Lake Rezaiyeh Expedition collected a number of maxillary and mandibular cheek teeth from Kopran. Bernor (1985) reported the following salient features for this sample: maximum crown height is 50 mm; protocone shape in early wear is elongate, labially rounded and lingually flattened with a generally elongate-oval shape; pre- and postfossette plications are complex and the bands are thick (not thin); hypoglyph is deeply incised. Mandibular cheek teeth have the following salient features: maximum recorded crown height is 44 mm; pli caballinid is absent, ectoflexid is generally not deep; linguaflexid is moderately deep; metaconid and metastylid vary in shape, being generally rounded to squared.
Remarks: The type specimen of “H.” gettyi was collected by Pohlig from Kopran in the later half of the nineteenth century. Kopran I and II occur in the westernmost portion of the Maragheh Basin where the local exposures are currently encroached upon by the expanding town of Maragheh. The Kopran localities are the lowest fossil levels of the section (150–120 m below loose chippings of Campbell et al. 1980). Bernor (1985) referred maxillary cheek teeth to “H.” gettyi, these being collected by the Lake Rezaiyeh Expedition from the 150- to 70-m levels. Mirzaie Ataabadi et al. (2013) revised the biostratigraphy and chronology of the Maragheh sequence, indicating that Kopran I occurs at the −150-m level and Kopran II occurs at the −120-m level, and they have interpolated ages of 8.967 and 8.7216, respectively. These ages are correlative with Europen MN10, the late Vallesian.
Bernor (1985) provided an extensive differential diagnosis between “H.” gettyi and mostly more primitive members of the early concept of “group 1” hipparions of Woodburne and Bernor (1980) and Bernor et al. (1980). Essentially, there are two genera that are included in “group 1”: Hippotherium (Bernor et al. 1997 and 2011) and Cormohipparion (sensu Bernor et al. 2003) or “Cormohipparion” (sensu Bernor et al. 2004 and Bernor and White 2009). H. primigenium (Bernor et al. 1997) is best known from the 16 skeletons that have been excavated from Höwenegg, Germany (10.3 Ma., Swisher 1996; Woodburne et al. 1996). Bernor et al. (2011) described a late assemblage of H. malpassi from Baccinello, Italy (ca. 7.1 Ma), and demonstrated the genus’ geographic extension into Italy, Greece (H. brachypus) and Turkey (H. brachypus, a large variety) in the early to middle Turolian (ca. 8.7–7.1 Ma.). Cormohipparion is known from MN 9 (early Vallesian) levels of Sinap and Esme Akçakoy, Turkey (Co. sinapensis), while the more advanced form “Cormohipparion” (“Cormohipparion” africanum) is known from Bou Hanifia, Algeria, and Chorora, Ethiopia (Suwa et al. 2015).
“H.” gettyi differs from more primitive Hippotherium in the following characteristics: generally smaller size; having a POF less deep dorsoventrally and medially with a less distinct anterior rim; a nasal notch that is slightly more retracted to above the P2 parastyle; cheek tooth fossettes which are less complexly plicated; shorter and rounder protocones; somewhat less deeply incised hypoglyphs; mandibular cheek tooth pre- and postflexids being less complexly plicated. “H.” gettyi is approximately the same size as Co. sinapensis, has a slightly less well-developed POF than C. sinapensis and has cheek teeth that have less elongate protocones than Co. sinapensis. There are a number of cranial and dental features that unite “H.” gettyi with these superspecific clades, including a lack of deeply incised nasal bones, a long POB, a medially deep and distally deeply pocketed POF, maxillary cheek teeth with relatively complexly plicated fossettes and persistently double pli caballins. MPIIIs and 1PHIIIs are thus far unknown for “H.” gettyi, which makes comparisons to species of Hippotherium, Cormohipparion and “Cormohipparion” untenable. Morphologically, “H.” gettyi seems to be a taxon transitional between Cormohipparion and Hippotherium on the one hand and Hipparion s.s. on the other. Collection of postcrania from Kopran I and Kopran II is essential for elucidating “H.” gettyi’s superspecific taxonomy.
aff. Hippotherium brachypus
?Hipparion urmiense, Gabunia, 1959, p. 166–176; pl. VI; figs. 2 and 3
Hipparion Group 3, Woodburne and Bernor, 1980; tab. 3
Hipparion cf. prostylum, Bernor et al., 1980, pg. 736, tab. 2
Hipparion prostylum (s.l.), Bernor, 1985, pg. 198; fig. 11a–g
Age: Middle Maragheh, ca. 8.6–8.2 Ma.
Referred specimens: Skulls including KNHM A4847, KNHM 4844, JGUM (currently BSPG) MB67, JGUM (currently BSPG) MB102, MNHN Mar. 71, MNHN Mar. 18; MNHN Mar. 465; MNHN Mar. 359, AMNH 27807, AMNH 27809. Metacarpals and metatarsals III cited in Supplementary table (Online resource) have been identified from bivariate and log10 ratio plots herein.
Diagnosis: A hipparion species with a long POB, a lacrimal placed well posterior to the POF and the POF being reduced compared to other members of the Hippotherium clade, moderate to short in length, faintly egg- to C-shaped, anteroposteriorly oriented, shallow to very shallow dorsoventrally, with slight to no posterior pocketing, moderate to slight medial depth, the anterior rim being faint to absent and the peripheral rim being moderately to weakly expressed. The nasal notch is placed either just anterior to or above the P2. In middle stage-of-wear, maxillary cheek teeth have moderately complex plications of the pre- and postfossettes; pli caballins are usually single but may occasionally be double; hypoglyphs are moderately deeply incised; protocones are variable in morphology but most commonly may have some slight lingual flattening and are rounded. The P2 anterostyle is elongate. Metapodials are relatively robust as in Hippotherium primigenium, H. malpassi and H. brachypus.
Description: Bernor (1985) described nine skulls and partial skulls which he referred to Hipparion prostylum (s.l.) largely based on the derived, shallow POF which was believed to be related to H. prostylum Gervais, 1849: KNHM A4847; KNHM A4844; JGUM (now BSPG) MB67 (Fig. 11a, b); also, herein JGUM (now BSPG; 1985, fig. 11B) and JGUM (now BSPG) MB 102 (Fig. 12a, b, added here to the original 9 described by Bernor 1985); Mar. 71 (Fig. 13; also Bernor 1985, fig. 11C); MNHN Mar. 18 ([also RLB 1474] Fig. 14a, b [Bernor 1985, fig. 11D)]), Mar. 359 ([also RLB 1475]; Fig. 15a, b [Bernor 1985, fig. 11F)]); Mar. 465 (Bernor 1985, fig. 11E) and AMNH 27807 and 27809.

The aff. H. brachypus specimen (BSPG-MB67) in lateral (a) and dorso-lateral (b) views

The aff. H. brachypus specimen (BSPG-MB102) skull with postcrania showing relatively robust limbs

The aff. H. brachypus specimen (MNHN Mar. 71)

The aff. H. brachypus specimen (MNHN Mar. 18 (also RLB 1474) in lateral (a) and ventral (b) views

The aff. H. brachypus specimen (MNHN Mar. 359 (also RLB 1475) in lateral (a) and ventral (b) views
JGUM (now BSPG) MB67 (Fig. 11a, b) was collected by Tobien in the late 1960s from Middle Maragheh in the vicinity of MMTT 1 (Tobien, personal communication to Bernor). It was collected together with a block that included axial and postcranial skeletal elements including a robust MTIII (Fig. 12). MB67 is a sub-adult specimen, virtually complete except for the posterior orbital rims and cranial vault and cavity. Its dentition is complete with right and left I1-2, dI3, C, P2-M2 (fully erupted) and M3 (nearly completely erupted). The canine is large, indicating that this is a male individual. The nasals are complete and reflected just to the anterior margin of the P2 (Fig. 11a). The POF is moderately long, extending anteriorly to a level above the P3 mesostyle. The POF has moderate dorsoventral dimensions, medial depth and peripheral rim morphology and has an egg shape with medial depth and a distinct posterior rim with no posterior pocketing (Fig. 11b). The lacrimal is not well delineated but enough is preserved to observe that it did not invade the POF. The P2-M2 are erupted but insufficiently worn to reveal middle stage-of-wear occlusal morphology. As a result, protocones are elongate and bucco-lingually compressed and hypoglyphs are deeply incised on M1-2.
The MNHN skulls include two more complete specimens (MNHN Mar. 18 [Fig. 14a, b] and Mar. 359 [Fig. 15a, b]; Bernor 1985, figs. 11D and 11F, respectively) and two more fragmentary specimens (MNHN Mar. 71 and MNHN Mar. 465; Bernor 1985, figs. 11C and 11E, respectively). In all of these skulls, the facial and nasal bone morphology is very similar to BSPG MB67 and MB102, and in MNHN Mar. 359 and Mar. 18, the lacrimal is placed well posterior to the POF (Bernor 1985). Of all the MNHN individuals, Bernor (1985) identified MNHN Mar. 71 as having cheek teeth in a stage-of-wear that preserves occlusal morphology: the pre- and postfossettes are moderately complex on all cheek teeth; pli caballins are faintly bifid on P3-M1 and M3 while being single on P2 and M2; hypoglyphs are shallowly incised on P2-3, moderately deeply incised on P4-M2 and very deeply incised on M3; the protocone is elongate on all cheek teeth (Bernor 1985).
The AMNH has two skull fragments of this taxon, AMNH 27807 (Bernor 1985, fig. 11G) and AMNH 27809. NHMW has another skull, KNHM A4847 (Bernor 1985, fig. 11A) that is very similar to AMNH 27807 and the BSPG and MNHN specimens described above. Cheek teeth of these individuals add no new information because they are either too early in wear or too late in wear to accurately discriminate middle stage-of-wear adult morphology.
Discussion: Bernor (1985) cited the complicated systematics of H. prostylum as it existed in the 1980s. The genotype Hipparion was named by Christol (1832) for material collected from Mt. Luberon (= Cucuron), France. Christol noted that this horse was characterised (in part) by three toes on each foot and an isolated protocone. Gervais (1849) designated three Mt. Luberon species, H. prostylum, Hipparion diplostylum and Hipparion mesostylum, based on the position and frequency of accessory mandibular cheek tooth stylids. Later, Gervais (1859) reunited all of these species into H. prostylum, recognising the usual variability in these characters. Gaudry (1873), Osborn (1918) and Skinner and MacFadden (1977) followed Gervais (1859) in referring all of the Mt. Luberon material to H. prostylum. Woodburne (personal communication to Bernor) has noted that Gaudry (1873, Pl. 6, fig. 1) illustrated the first material from Mt. Luberon that shows the morphology of the face anterior to the orbit. This specimen was re-illustrated by Skinner and MacFadden (1977, fig. 3A). A formal type has not been designated by any of these authors. Sondaar (1974) stated that the specimen figured by Gervais (1859, Pl. 19, fig. 2, a maxilla fragment with P3-M2) should be considered to be the holotype of H. prostylum. Bernor (1985) argued that this was an informal taxonomic recommendation and suggestive of a possible lectotype.
An additional complication arises as a result of Sondaar’s (1974) correct observation that there is likely a second, rare species of “Hipparion” occurring at Mt. Luberon. Woodburne (personal communication) agreed with this observation, noting the occurrence of a taxon with more robust metapodials and slightly more complex plications of the upper and lower molars. Bernor and Hussain (1985) continued in their observation of species heterogeneity in the Mt. Luberon assemblage, citing BMNH M22617 as having a larger fossa and more complex plications of the pre- and postfossettes than other Mt. Luberon specimens. Bernor (1985) nominated a skull, BMNH M33603 (fig. 7J of Woodburne and Bernor 1980) as the Lectotype of H. prostylum. This is a virtually complete skull with P2-M3, lacking the premaxilla and posterior-most cranium and conforming to Woodburne and Bernor’s (1980) diagnosis of H. prostylum.
All previous work on H. prostylum has been dominated by skull, facial and maxillary cheek tooth morphology. While Sondaar (1974) did address postcranial heterogeneity in the Mt. Luberon sample, there has not been a critical evaluation of postcranial morphology. Our analysis of postcrania is revealing. The MNHN Maragheh sample has a number of individuals that plot within the Höwenegg ellipse for MCIII (Fig. 4b), MTIII (Fig. 5b) and 1PHIII (Fig. 6b); the MPIIIs are the most convincing. The log10 ratio diagrams for MCIII (Fig. 7a) and MTIII (Fig. 8b) reveal plots very close to Pikermi H. brachypus mean values.
We no longer are able to refer the Middle Maragheh hipparion sample to H. prostylum sensu lato. Our analysis reveals that the elongate-slender distal limb elements of Mt. Luberon H. prostylum relate closely to H. campbelli from Upper Maragheh (Figs. 8a and 9c), not to the Middle Maragheh hipparions that have Hippotherium-like MPIIIs. Amongst species of Hippotherium, the Middle Maragheh hipparions compare most closely with Pikermi H. brachypus but do not have the flat log10 plot of H. brachypus (Pikermi). They agree particularly well in the postcranial elements. The facial morphology of Maragheh aff. H. brachypus has a further reduced POF, particularly in its decreased medial depth and posterior pocketing like “H.” urmiense Gabunia, 1959. However, in itself, Pikermi H. brachypus has a less dorsoventrally extensive and less posteriorly pocketed fossa compared to Central European Vallesian Hippotherium and Italian H. malpassi. If our hypothesis proves correct and the Middle Maragheh robust-limbed hipparion is a member of the Hippotherium clade, this observation extends the geographic range of Hippotherium from Central and Western Europe to Greece (Pikermi [ca. 8 Ma], H. brachypus), Turkey (Akkasdagi [7.1 Ma], H. brachypus large variety) now as far east as Maragheh, Iran (aff. H. brachypus (with “H.”? urmiense being a possible junior synonym [8.6–8.2 Ma]). Possible occurrence of this clade (aff. H. brachypus) was also recently reported from a new locality in northwest Iran (Mirzaie Ataabadi et al. 2011).
We realise that our recognition of Middle Maragheh aff. H. brachypus is in conflict with previous conventions about the primacy of facial morphology for the recognition of “Hipparion” clades. We can defend the recognition of Maragheh aff. H. brachypus by noting the great similarity between it and other members of the clade in its MCIII and MTIII morphologies. Reduction of the POF no doubt is related to the increased crown height in H. brachypus maxillary cheek teeth: as crown height increased, POF became reduced in dorsoventral and medial depth in order to avoid the cheek tooth roots from penetrating the ventral extent of the POF. As in all Hippotherium, H. brachypus and aff. H. brachypus retained primitive proportions of the MCIII and MTIII and unretracted nasal bones while evolving higher crowned cheek teeth that forced the dorsoventral reduction of the POF. It is clear from our analyses that Hipparion s.s. is a clade of hipparion including H. prostylum, H. dietrichi, H. campbelli and possibly Chinese Hipparion hippidiodus that evolved more slender-elongate limbs that were convergent with species of Cremohipparion. It is not yet certain that Chinese “H.” hippidiodus is a genuine Hipparion s.s.; the facial-maxillary cheek tooth morphology is very similar to Hipparion s.s., but the postcranial anatomy is currently unknown.
Hipparion campbelli Bernor, 1985
Hipparion dietrichi Wehrli (1941), sensu Bernor (1978), p. 50, fig. 2; Bernor et al. (1979b), p. 94; Woodburne and Bernor (1980), pg. 1344, figs. 7a, 7b; Bernor et al. (1980) (in part), p. 724 fig. 8; Bernor and Hussain (1985), p. 40, fig. 5
Hipparion campbelli Bernor (1985), p. 206, figs. 13A–D, 14A–D and 15A–B
Holotype: A skull with an associated lower jaw MMTT 13/1342 (Bernor 1985, fig. 13).
Type locality: MMTT 13 near Sholl’avand, Upper Maragheh, Iran.
Age: 7.88724 Ma interpolated age anchored by single crystal argon ages, MN12, medial Turolian (Swisher 1996; Mirzaie Ataabadi et al. 2013).
Referred specimens (following Bernor 1985 ): The following skull material has been referred to H. campbelli: MMTT 13/1342 (type specimen), an adult female skull; MMTT 13/1343, a juvenile skull; MMTT 13/1291, a juvenile skull fragment; MMTT 13/1333, a juvenile skull fragment. The following mandibular material is referred to H. campbelli: MMTT 13/1342 (associated with the type skull); MMTT 13/1736, mandible with very worn teeth of a senile individual; MMTT 13/2353, juvenile mandible with dentition; isolated teeth and bones referred to H. campbelli are listed in the ESM Table 1.
Geographic range: Iran.
Diagnosis (after Bernor 1985 ): A medium-sized hipparion species with an elongate and slender snout. POF is markedly reduced, egg-shaped and anteroposteriorly directed, with a small vestigial posterior rim and slight medial depression; preorbital bar is moderately long; the lacrimal bone is placed well posterior to the POF; nasal notch is slightly retracted to the mesostyle of the P2; middle adult wear maxillary cheek teeth have moderately complex and very thinly banded plications of the pre- and postfossettes, pli caballins are single, hypoglyphs are moderately deeply incised; protocones are oval shaped; the P2 anterostyle is elongate; mandibles have an elongate, slender symphysial region; canines are placed immediately distal to i3, the mental foramen is placed approximately one half the distance between i3 and p2; in middle adult wear, the p2 anterostylid is elongate, ectoflexids do not separate metastylids and metaconids in the premolars, but do so in the molars; pli caballinids and ectostylids are absent; linguaflexids are shallow; metaconids and metastylids are rounded. MCIIIs and MTIIIs are elongate and slender; 1PHIIIs are elongate and slender (re: ESM Table 1 and bivariate and log10 ratio diagrams above).
Description (following Bernor 1985 ): Bernor (1985) reported that H. campbelli is the most abundantly represented hipparionine in the MMTT Lake Rezaiyeh Expedition sample. This is due to the extensive quarrying activity at locality MMTT 13. The most complete specimen is the adult female skull and mandible, MMTT 13/1342 (Fig. 16a–e). MMTT 13 has also yielded juvenile skulls MMTT 13/1343 (Fig. 17a–c), 13/1291 and 13/1333. There is a mandible associated with the type skull that was numbered at the time MMTT 13/1341, and we currently recognise this as MMTT 13/1342 because it is directly associated with the type skull. Other mandible specimens include MMTT 13/1289, 13/2352 and 13/1726 as well as a number of isolated teeth (re: ESM Table 1, herein). MMTT 13 associated postcrania is moderately extensive (ESM Table 1); MCIIIs, MTIIIs and 1PHIIIs are elongate and slender. They compare well with Mt. Luberon H. prostylum.

Hipparion campbelli skull in lateral (a), dorsal (b) and ventral (c) views and its mandible in lateral (d) and ventral (e) views

Hipparion campbelli juvenile skull paratype in a dorsal, b lateral and c ventral views
The type specimen, MMTT 13/1342, is an adult female skull and mandible (Bernor 1985, figs. 13A, 14A; Fig. 16a–e here). The skull is of moderate size and virtually complete except for the occipital portion of the cranium, the anterior most nasal bones, left I3 and C. The snout is elongate, narrow and with a sharply arcuate incisor arcade. The POF is moderately distant from the orbit and placed high on the face; it has a faint egg shape and anteroposterior orientation but is clearly vestigial with a tiny posterior rim and no posterior pocketing; it is short anteroposteriorly and dorsoventrally and shallow medially. The infraorbital foramen is large and located at the anteroventral border of the POF and above the interstitial contact of P3-P4. The canine is a small peg-like structure indicative of a female individual. The cheek teeth are not yet in a middle stage-of-wear and are generally elongate in shape. P2 has a well-developed, elongate anterostyle; enamel plications are faintly expressed due to early wear but show a degree of complexity with thinly banded fossette plications; the hypoglyph is moderately deeply incised; the protocone is oval shaped. P3 and P4 are in earlier wear than the P2 and as a result have very faintly delimited, thinly banded enamel plications; the distal borders of the prefossettes are touching the mesial borders of the postfossettes; the hypoglyphs are moderately deeply incised; protocones are slightly oval with each one having a vestigial anterolabial enamel pli. M1 and M2 are substantially smaller and more square shaped; fossette enamel plications are faintly expressed; protocones are round to oval in shape. M3 is not completely erupted, with only its mesial one half beginning to wear. Pli caballins are single on all teeth.
The MMTT 13/1342 type mandible (Bernor 1985; fig. 15A and B; Fig. 16d, e here) is nearly complete, only missing the superior two thirds of the ascending ramus. The mandible is also broken just posterior to the canines. As with the maxillary dentition, i3 and m3 are not fully erupted. The mental foramen is positioned slightly closer to p2 than i3. The mandibular symphysis is elongate and the incisor arcade is strongly arcuate. The canines are small, rounded and peg-like and placed immediately distal to i3. The premolars are elongate and substantially larger than the molars. Mandibular p2 mirrors P2 in its strongly extended and anteriorly rounded anterostylid; p2’s anterostylid is elongate; metaconid and metastylid are angular to rounded with the metaconid being subequal in size to metastylid. Mandibular p3 and p4 are identical in occlusal morphology to p2, saving their lack of an anterostylid and that metaconid and metastylid are more rounded. Mandibular m1-2 exhibit serial size reduction in both length and width measurements with m3 being elongate due to the extended talonid. Unlike in the premolars, the ectoflexid separates the metaconid-metastylid in the molars. All cheek teeth lack protostylids and ectostylids.
MMTT 13/1343 is a virtually complete female juvenile skull with dI3, dP2-4 and M1 with M2s exposed in their crypts. This specimen was excavated in very close proximity to MMTT 13/1342 and is similar to it in skull morphology: the POF is identical; the lacrimal is very well delimited and is well posterior to the POF; the infraorbital foramen is large, located anteroventral to the POF and above dP3, whose lingual border is partially exposed in the maxillary lateral wall. The incisor region is very narrow and I3 remains unerupted. The canines are missing, but their alveoli are perfectly preserved and small suggesting that this was a female individual. The dP2-4 are very worn and preserve no fossette morphological details; protocone is rounded on dP2-4 and connected with the protoloph on dP2-3; the hypocones are very shallowly incised. The anterostyle of dP2 is as in the adult female. M1 is too unworn to reveal any occlusal morphology.
There are two other juvenile skull fragments, MMTT 13/1291 (Bernor 1985, figs. 13C and 14C) and MMTT 13/1333 (Bernor 1985, figs. 13D and 14D), that were also excavated from the Sholl’avand Quarry. The morphology of these two specimens is consistent with MMTT 13/1343.
Bernor (1985) suggested possible associations of MMTT 13 mandibles and skulls beyond the type specimen, MMTT 13/1342, including skull MMTT 13/1333 with mandible MMTT 13/1289 and palate MMTT 13/1291 with mandible MMTT 13/2353. A fourth mandible, MMTT 13/1736, has very worn cheek teeth and belongs to a senile individual; its taxonomic identification is not certain. The morphology of these mandibles does not add significant information for diagnosing H. campbelli.
Discussion: Bernor (1985) nominated the MMTT 13/1342 skull (with mandible) as the type of H. campbelli. In so doing, Bernor (1985) corrected previous referrals to H. dietrichi Wehrli 1941 (Bernor 1978; Bernor et al. 1979; Woodburne and Bernor 1980; Campbell et al. 1980). The known stratigraphic range of H. campbelli is restricted to Upper Maragheh and is the defining taxon for that biostratigraphic interval (Bernor 1985, 1986).
H. campbelli is the sister taxon of H. dietrichi, differing in the following characters: the snout is more slender; the incisors have an arcuate alignment instead of being horizontally aligned; the POF is more restricted, egg-shaped and consistently shallower in its medial dimension. MPIIIs in both H. campbelli and H. dietrichi are elongate and slender. Cheek teeth are also similar between these two taxa. Bernor et al. (1980) suggested that H. dietrichi (and by extension, H. campbelli) might be related to East African late Miocene “Hipparion” turkanense (currently Sivalhippus turkanensis; Wolf et al. 2013), because of their mutual loss of the POF. Bernor and Hussain (1985) showed that multiple Old World hipparionine lineages lost the POF, while Bernor and Harris (2003) and later Wolf et al. (2013) showed that Lothagam Sivalhippus turkanensis had far more robust metapodial IIIs than species of Hipparion s.s.
The current study shows that the close metapodial III morphological resemblance between Mt. Luberon H. prostylum and H. campbelli (and, by extension, H. dietrichi) enables all three taxa to be members of the Hipparion sensu stricto clade. It has yet to be demonstrated based on MPIII morphology that Chinese H. hippidiodus is also a member of the Hipparion s.s. clade.
Cremohipparion Qiu et al., 1987
Cremohippparion aff. moldavicum
Hipparion mediterraneum Roth and Wagner (1855); sensu Forsten (Forsten 1968) (in part), p. 40.
Hipparion species 1, Bernor (1978), fig. 3A; Bernor et al. (1979b), p. 94
Hipparion species 2, Bernor (1978), fig. 3B
Hipparion mediterraneum, Campbell et al. (1980), p. 841; Bernor et al. (1980), p. 728, fig. 8; Bernor and Hussain (1985), p. 39, fig. 4B
“Hipparion” Group 2. Woodburne and Bernor (1980), p. 1329, figs. 6A–D
“Hipparion” aff. moldavicum Bernor (1985), p. 211, figs. 16A–I and 17A–D
Holotype of Cremohipparion moldavicum (Gromova, 1952): P.I.N. no 1256-3639, Academy of Sciences Paleontological Institute of the U.S.S.R., figured by Gromova, 1952, figures 1–3
Type locality: Taraklia, Republic of Moldavia, District of Bender.
Age: Maeotian (= Medial Turolian, MN 12).
Geographic range: Moldavia, Turkey and Iran.
Diagnosis (translated from Gromova, 1952: 154–155): Medium-sized hipparion, basal length of cranium = 271–273 mm; length of cheek teeth (P2-M3) = 121–141 mm; muzzle elongate; index of orbital-facial length = 67; index of anatomical axes = 214.6. Frontals narrow, width index = about 38.2; fronto-basal index = 261.4 mm. Dental series short; index of length to basal width = 33.5; to the premolars, 41.2–46.8; diastemato-dentary index = 66.1–28.9. Upper molars large relative to premolars; molar-premolar index = 82.4–91. A single POF, very long and elevated, index of position relative to orbit = 26.6–37.8; relative to the facial crest, 16.7–64.3. Nasal notch moderately deep, its posterior border is at the level of, or slightly anterior to the anterior border of P2. The diastema is well developed; its index is 60.4–77. Protocone is short and wide, length index of P3-M2 in little or moderately worn individuals is 20.7–37.5, in very worn teeth, 25–43.3; index of form in the same conditions 42.8–78.3 and 57.1–92.3. Enamel plication is moderate in upper cheek teeth, on the posterior wall of the prefossette and anterior wall of the postfossette; when P3, 4 and M1, 2 are very worn or moderately worn, they have 3.5–6.5 to 9.5 plis. Cheek teeth have a height-length index on P3, 4 of 156–195.5; on M1-3, 204.3–232.5; on p3, 4 and m1, 2 159–200. A double knot (metaconid-metastylid) of Hipparion type (rounded with intervening V-shaped linguaflexid). External depression in lower cheek teeth deep, complementary elements little developed. The islette occupies the anterior portion of the I3 crown. The postcranials are gracile and elongate; index of width in the lower articulation relative to the width of MCIII, 14.5–16.3; to the length of MT III, 12.4–14.2. Metapodial length relative to width: MT/T = 74.4. Lateral digits are moderately developed; moderate indices of lower extremities diameters MC II and IV to MC III, 75.8, and MT II and IV to MT III, 66.8; moderate indices of length of the first phalanges of the lateral digits to those of the anterior median digits, 58.2; posterior, 53.8. Extremities recurved to the level of articulations; moderate index of the pisiform bone 114.3 mm; anterior third phalanx narrow; length-width index of posterior third phalanx 71.4–86.6.
Emended diagnosis (modified from Bernor, 1985): A medium-sized hipparionine with an elongate snout. The POF is single, subtriangular shaped, anteroposteriorly oriented and elongate, dorsoventrally and medially deep, with slight posterior pocketing, a distinct anterior rim and strongly expressed peripheral outline. The POB is short with the lacrimal bone invading the posterior aspect of the POF. The nasal notch is incised at a level either just above the mesial border of P2, or slightly mesial to it. Middle wear adult cheek teeth have moderately complex plications of the pre- and postfossettes; protocones are round to oval and show lingual flattening in some individuals. The P2 anterostyle is usually elongate but can be short and rounded in some individuals. MCIIIs and MTIIIs are elongate and slender.
Description: Bernor (1985) identified nine skulls and skull fragments of Maragheh Cr. aff. moldavicum (his “Hipparion” aff. moldavicum) from the BMNH (M3924), NHMW (A4848, RLB 8403, RLB 8404) and the MNHN (Mar. 1477 (= RLB 7914), Mar. 71 (= RLB 7915), Mar. 1476 (= RLB 8001), Mar. 466 (= RLB 8002) and Mar. 3428 (= RLB 8003)). Of these, only BMNH M3924 has a complete snout and is directly comparable to the Taraklia assemblage of Cr. moldavicum as described by Gromova (1952).
BMNH M3924 (Bernor 1985, figs. 16A, 17A; Fig. 18a, b here) is a moderate-sized sub-adult male skull with Cs (erupting) and P2-M2 and with the right M3 erupting. Most of the cranium is complete. The POF is long, subtriangular shaped, anteroposteriorly oriented, dorsoventraly deep, slightly pocketed posteriorly, medially very deep with distinct, concentrically shaped pits, a strongly expressed anterior rim and peripheral rim. The POB is short and the lacrimal bone clearly invades the posterior aspect of the fossa. The nasal notch is retracted to a level above the P2 parastyle. The IOF is placed above the distal one half of P3 and just inferior to the anteroventral rim of the fossa. The cheek teeth are restricted in their height due to the great ventral extension of the POF. The cheek teeth have complex, thinly banded plications of the pre- and postfossettes, irregularly shaped bifid pli caballins (P2-M1), hypoglyphs that are deeply (P2-P4), to moderately deeply (M1) incised; protocones are rounded to elongate shaped (P2-M2); P2 anterostyle is short and rounded. The muzzle is short and slender in proportion. BMNH M3924 differs most distinctly from Cr. moldavicum in its shorter snout; in all other characteristics, it is similar.

Cremohipparion aff. moldavicum skull (BMNH M3924) with a short snout in lateral (a) and ventral (b) views
The Vienna specimens reported by Bernor (1985) included (note that in 1985, Bernor used the KNHM abbreviation for NHMW): KNHM-RLB 8402, KNHM A4848, KNHM-RLB 8403 and KNHM-RLB 8404). Of these, KNHM-RLB 8402 was collected from Ketschawa (Kherjabad = MMTT localities 1A, 1B and 1 from Middle Maragheh) and is a palate fragment of a very old adult containing P2-M3, the right preorbital bar and ventral two thirds of the facial fossa. This morphology is as in BMNH M3924 and the entire Maragheh hypodigm of Cr. aff. modavicum. KNHM-RLB 8403 (Bernor 1985, fig. 16C) and KNHM-RLB 8404 (Bernor 1985, fig. 16D) likewise were collected from Ketschawa and are Middle Maragheh correlative. A fourth KNHM specimen, A4848 (Bernor 1985, fig. 16B), was collected from Zad Baschi (of uncertain stratigraphic provenance) and again agrees well with the Cr. aff. moldavicum facial morphology.
Bernor (1985) reported a single old adult mandible from Zad Baschi in the Vienna collection, KNHM A4845, that compares closely in size, occlusion and preservation with KNHM A4848. This specimen is the only known Maragheh mandible that can reliably be referred to Cr. aff. moldavicum. The salient morphologic features of this specimen include the following: cheek teeth are large and rather square in occlusal outline; premolar ectoflexids do not separate metaconid/metastylid; pli caballinids and ectostylids are absent; linguaflexid is shallow on p2 and V-shaped on p3-4; metaconids and metastylids are round on all of the premolars; molar ectoflexids separate metaconids and metastylids; pli caballinids and ectostylids are absent on all molars; linguaflexids are U-shaped on all molars and metaconids and metastylids are angular on all molars. The p2 anterostylid is elongate.
Of the five MNHN specimens of Cr. aff. moldavicum, MNHN Mar. 1477 (= RLB 7914), “the black skull”, is the largest (Bernor 1985, fig. 16E; Fig. 19a, b here). This specimen is lacking the snout and posterior rim. The salient morphological features are as in Cr. moldavicum: POB is of moderate length with the lacrimal invading the posterior aspect of the POF; POF is very large, subtriangular in shape, medially and dorsoventrally very deep, only slightly pocketed posteriorly; P2 absent; P3-4 have complex plications of pre- and postfossettes, protocone is rounded to oval with some lingual flattening, hypoglyph is moderately deeply incised on P4; molars are as in P4 except for the distal tapering of M3. MNHN Mar. 1476 (= RLB 8001; Bernor 1985, fig. 16G; Fig. 20a, b here) is very similar to MNHN Mar. 1477, but somewhat smaller. Salient morphological features are as in Mar. 1477 including POB short with the lacrimal invading the posterior aspect of the POF; POF with large subtriangular shape, being dorsoventrally and medially deep; POF posterior pocketing being slight; cheek teeth being largely obscured with matrix and glue but reveal P4-M3 with oval-shaped protocones; bifid pli caballins on P3-4, these being single on M1-M2; pre- and postfossettes appear to be complex; hypoglyphs are shallow. As in the type Cr. moldavicum material, there is no intermediate fossa as is characteristically found in Pikermi Cr. mediterraneum.

Cremohipparion aff. moldavicum skull (MNHN 1477) and (RLB 7914) without snout in a lateral and b ventral views

Cremohipparion aff. moldavicum skull (MNHN 1476) and (RLB 7915) without snout in a lateral and b occlusal maxillary teeth view
The remaining specimens of MNHN Maragheh Cr. aff. moldavicum Mar. 466 (= RLB 8002) and Mar. 3428 (= RLB 8003) add no further morphologic information to our characterisation given above. Full descriptions of all this material and the NHMW material can be found in Bernor (1985). The Paris collection of Cremohipparion has abundant complete MPIII material that we refer to Cr. aff. moldavicum. This material has MCIIIs and MTIIIs that are very elongate and slender as in the type Cr. moldavicum material described by Gromova (1952).
The problem is that in terms of MPIII bivariate dimensions M1 versus M11, H. campbelli plots in the middle of the Cremohipparion range (Figs. 4b and 5b). Moreover, Cr. aff. moldavicum and Cr. matthewi overlap in their ranges.
The genus Cremohipparion included an extensive suite of species that ranged across much of Eurasia and presumptively into North Africa (Bernor et al. 1996, 2012). Cr. moldavicum is one of the most primitive members of the clade and it is found from Moldavia to Greece, Turkey (Koufos and Vlachou 2005) and Iran in the medial Turolian. Koufos and Vlachou (2016) have reported the occurrence of a smaller Cremohipparion, Cremohipparion macedonicum, from a series of Greek localities of late Vallesian–Turolian age. Cr. mediterraneum is one of two typical taxa for Pikermi, Greece. Samos, Greece has the larger taxon Cr. proboscideum along with the small taxon, C. “matthewi” and the tiny taxon Cr. nikosi (Bernor and Tobien 1989). “Hipparion” periafricanum is believed to be related to Cr. nikosi and is arguably the smallest Old World hipparion (Bernor et al. 1996). Eastward from Iran, there occurs Cr. antelopinum from “Dhok Pathan levels” (ca. 8 Ma) of the Potwar Plateau, Pakistan (Wolf et al. 2013). In China, there are two taxa, the Baodean (late Miocene, late Turolian correlative, ca. 7 Ma) Cremohipparion forstenae (former Cr. richthofeni; Bernor et al. 1990) and the Pliocene age Yushe Basin highly derived form Cremohipparion licenti (Qiu et al. 1987; Bernor et al. 1996). Where known, these taxa are united by the following salient characters: facial morphology includes short POB, large, dorsoventrally extensive POF lacking significant posterior pocketing; some species exhibit strong nasal retraction (Cr. proboscideum, Cr. forstenae and especially Cr. licenti) and elongate MCIIIs and MTIIIs. Also, Cremohipparion includes a radicle of small to very small taxa including Cr. macedonicum, Cr. matthewi, Cr. nikosi and Cr. periafricanum. Cremohipparion would appear to be a late Vallesian derivative of Cormohipparion or possibly early Hippotherium that reduced the length of the POB and lost its deep posterior pocketing and at the same time evolved more elongate, slender limbs adapted for open country running (re: Bernor et al. 2003). The large, medially deep POF with strong anterior rim and peripheral outline is primitive for Old World hipparions.
Cremohipparion matthewi Kormos, 1911
“Hipparion”? matthewi Bernor (1985), p. 219, figs. 18A and B, 19A and B; Fig. 21a–c here

Cremohipparion matthewi skull (GIU P100-1958 (cast)) in lateral (a), dorsal (b) and oblique (c) views
Holotype: OK-557 Hungarian Geological Institute, Budapest, Hungary.
Type locality: Samos, Greece.
Referred specimens: GIU P100-1958 (cast), a skull fragment with rt. and lt. p2-m3; KNHM-RLB 8405, lt. mandible fragment with p2-m2; NHMW-RLB 8407, rt. mandible with m1-2; NHMW-RLB 8408, lt. mandible with m1-2; NHMW-RLB 8408, lt. mandible with m1-2; NHMW A4837, MCIII. MMTT specimens: 18/1438, rt. P2; 18/1447, lt. dP3; 18/1443, lt. dP3; 18/1440, rt. M1; 37/2089, lt. dP3; 37/2093, lt. P4; 37/2049, rt. M1; 25/1635, unerupted dP2-3; 24/1732, rt. M1; 24/1682, right calcaneum; 26/1828, rt. dPx; 26/2563, lt. dP2; 26/2562, lt. dP3; 26/1874, lt. P4; 26/1827, rt. p3; 26/1645, lt. M1-2, rt. M2; 26/1591, rt. m2; 26/1640, lt. m2; 26/1677, lt. astragalus; 26/1682, rt. calcaneum; 26/1684, articulated distal lt. metapodial, astragalus and navicular.
Geographic range: S.W. Asia and Greece.
Age: Medial–late Turolian.
Diagnosis: The OK-557 type specimen of Cr. matthewi is a complete skull of a small senile male hipparion with a complete dentition. Salient morphological features include small hipparion with nasal bones that are retracted to the mesial border of P2; POB is short with the lacrimal invading the posterior border of the POF; POF is relatively large, ovoid, with a weak peripheral rim, medially shallow with a ventrally deep extension as is found in Cr. moldavicum, with no posterior pocketing; P2 has a short anterostyle; protocones are oval to round, fossettes have relatively simple complexity and hypoglyphs are shallow due to their late stage-of-wear. There is no intermediate POF.
Description: Paul Sondaar provided Bernor with a cast of a skull fragment of a small, sub-adult hipparion, GIU P100-1958 (Bernor 1985, figs. 18A and B; here, Fig. 21a–c) collected from an unknown Maragheh locality. This specimen includes the middle part of the skull lacking the nasals and the cranium posterior to the anterior medial half of the orbits, right P2-M2 and left P3-M2; M3s are emerging in their crypts. This specimen compares very closely with the type specimen of Cr. matthewi, a cast of which was graciously provided by Professor Laszlo Kordos (Hungarian Geological Institute, Budapest, Hungary). Salient morphological features include POB slightly longer than the type specimen, with the lacrimal invading the posterior portion of the POF; POF is relatively large, ovoid, with a weak peripheral rim, medially shallow with a ventrally deeper extension than found in the type specimen and as is found in Cr. moldavicum, with no posterior pocketing; P2 has a broken anterostyle crown, but the anterostyle is believed to have been relatively short; premolar protocones are rounded even though in early wear, while M1 and M2 protocones are more oval-shaped due to early wear and M3s are just emerging from their crypts; pre- and postfossette plications are relatively simple, pli caballins are single; premolar hypoglyphs are deeply incised while being shallowly incised on M1. Both GIU P100-1958 and the type specimen of Cr. matthewi exhibit a remarkable similarity to Maragheh Cr. aff. moldavicum, essentially being smaller and with a reduction of the POF medial depth and peripheral rim expression.
Bernor (1985) reported a number of isolated maxillary cheek teeth that he referred to “H.” cf. matthewi that we refer to the present taxon, Cr. matthewi. These teeth have the following salient features: relatively buccolingually narrow and anteroposteriorly elongate; the posterior border of the prefossette and the anterior border of the postfossette show the strongest plications; the protocone is rounded or flattened lingually until late wear; the hypoglyph is moderately deep. These teeth compare closely with GIU P100-1958.
Mandibular cheek teeth from Maragheh attributed to Cr. matthewi are consistently small, buccolingually narrow and anteroposteriorly elongate; they tend not to have pli caballinids; ectoflexids do not separate metaconid and metastylid in the premolars, while doing so in the molars; metaconids and metastylids are rounded, linguaflexids shallow. The best-preserved mandibular specimen is MMTT 26/1573, a left mandible fragment with p4-m3 (Bernor 1985, fig. 19A and B).
The NHMW collection has a number of small mandibular teeth and a small MCIII referable to Cr. matthewi. Mandibular remains include NHMW-RLB 8405, 8406 and 8407. NHMW-RLB 8405 is an adult left mandible fragment with p2-m3 with the following salient features: cheek teeth are small and have an elongate, somewhat bucco-lingually compressed occlusal profile; pli caballinids are absent; linguaflexids are shallow on p2, U-shaped on p3-m1 and V-shaped on m2; metaconids and metastylids are rounded on all of the cheek teeth. NHMW-RLB 8406 is a left mandible with p4-m2; cheek teeth are more square-shaped due to advanced wear; ectoflexids separate metaconid and metastylid in all of the cheek teeth; pli caballinids are as in 8405. NHMW-RLB 8407 is a right mandible with m1-2 and NHMW-RLB 8408 is a left mandible fragment with m1-2; both are of senile individuals. NHMW-RLB 8408 is from Kopran II and represents the oldest occurrence of Cr. matthewi at Maragheh: Lower Maragheh, 8.7216 Ma, early Turolian, MN11 (Mirzaie Ataabadi et al. 2013).
The Ilkhchi MCIII, NHMW A4837 is complete. Bernor (1985) reported it as being the smallest MCIII he had seen from Maragheh. It is relatively elongate and very gracile in shape; the sagittal keel is sharply extended; the caudal surface is deeply set between two ridges; the distal cranial pit is very deep, accentuated by a raised proximal rim. Our analysis herein (Fig. 4b) shows that there are even smaller specimens of Maragheh Cr. matthewi from the AMNH and MNHN. We provide a list of Maragheh Cr. matthewi postcrania in ESM Table 1 herein.
Discussion: Bernor (1985) reported that the nomen “Hipparion” matthewi has traditionally been used for dwarf species from later Turolian levels of the eastern Mediterranean, southwest Asia and the Ukraine. Bernor’s (1978) original assignment of the small hipparionine material from Maragheh to H. matthewi was based primarily on a size comparison of the Lake Rezaiyeh collections’ fragmentary jaws, teeth and postcrania to a very well-preserved skull from Samos housed in the NHMW (NHMW A4742; Bernor and Hussain 1985, fig. 6A). Bernor (1985) followed Woodburne and Bernor (1980) and Bernor et al. (1980) in citing “Group 2” Old World Hipparion affinities: short POB with lacrimal invading posterior aspect of POF; relatively large POF dimensions of length and dorsoventral extent; slight posterior pocketing.
Bernor and Tobien (1989) recognised the nomen Hipparion (Cremohipparion) of Qiu et al. (1987) as being valid at the genus rank and named a new dwarf form with retracted nasals from Samos, Cr. nikosi. Bernor et al. (1996) as well as we here recognise 10 Cremohipparion species, including Cr. moldavicum, Cr. macedonicum, Cr. mediterraneum, Cr. proboscideum, Cr. matthewi, Cr. nikosi, Cr. periafricanum, Cr. antelopinum, Cr. forstenae and Cr. licenti. These taxa range from Spain to China and Indo-Pakistan and may also occur in North Africa. Chronologically, the Cremohipparion clade ranges from the late Vallesian (Cr. moldavicum and Cr. macedonicum) to the early Ruscinian of China (Cr. licenti). Maragheh Cremohipparion includes two species, Cr. moldavicum and Cr. matthewi. Skull, cheek tooth and postcranial material from Maragheh suggest that Cr. matthewi could be a descendant of Cr. moldavicum; Cr. matthewi evolved into a smaller form, with slender metapodials and a reduced POF beginning in the later Vallesian.
Biostratigraphy and biogeography
Biostratigraphy
Here (Fig. 22), we amend the biostratigraphic range charts of the hipparionine horses from Maragheh provided by Bernor (1985, 1986) and Mirzaie Ataabadi et al. (2013). We define the biostratigraphic units and their characteristic taxa as:

Lower Maragheh: “Hipparion” gettyi and Cremohipparion matthewi
Middle Maragheh: aff. Hippotherium brachypus, Cremohipparion aff. moldavicum and Cremohipparion matthewi
Upper Maragheh: Hipparion campbelli and Cremohipparion matthewi
Biogeography
The Old World “Hipparion” radiation occurred shortly after the introduction of North American Cormohipparion into Eurasia and Africa and was appropriately termed the “Hipparion Datum” in the hallmark publication by Berggren and Van Couvering (1974). That Cormohipparion was the initial progenitor and likely the sole genetic stock of the “Hipparion Datum” is of little doubt (Bernor et al. 1997, 2010; Woodburne 2007). Cormohipparion is recorded from Sinap, Turkey (Bernor et al. 2003), and the Potwar Plateau, Pakistan (Wolf et al. 2013), at ca. 10.7 Ma. The taxonomy of the oldest and most primitive hipparions from the Pannonian C horizons of the Vienna Basin is very primitive in some dental characters but shows shared derived characters of the maxillary cheek teeth with H. primigenium. The morphology, taxonomy, geological and palaeoecological contexts of these hipparions are under study by Bernor, Göhlich and Harzhauser (in progress).
The most primitive and oldest Maragheh hipparion is “H.” gettyi. Bernor (1985) believed that this was the most primitive member of the Hipparion s.s. clade biostratigraphically displayed through the Maragheh section; “H.” gettyi was used to characterise and define Lower Maragheh, “H.” prostylum characterised and defined Middle Maragheh and H. campbelli characterised and defined Upper Maragheh. However, the new systematics that we have adopted here does not support this previous interpretation in all regards.
“H.” gettyi exhibits shared primitive characters of the cranium that reveal either a Cormohipparion or Hippotherium relationship, in particular in the primitive retention of a large, dorsoventrally deep and posteriorly pocketed POF and retention of bifid pli caballins on the maxillary cheek teeth. MPIIIs are unknown for “H.” gettyi. “H.” gettyi has not been reported elsewhere, but its cranial and dental morphology is not greatly different from Co. sinapensis, which is a plausible relative. “H.” gettyi’s age at between 9.0 and 8.5 Ma is significantly less than the 10.7–10 Ma Co. sinapensis’ range in Turkey. This limited information suggests that “H.” gettyi’s biogeographic range was centred in Turkey and Iran.
The MNHN collection has a suite of skulls that Bernor (1985) referred to “H.” prostylum. These were accompanied by some MPIIIs that were more robust than the Cremohipparion assemblage plotted herein (Fig. 5b bivariate plot). Tobien’s collection of two skulls and postcrania cited earlier here in (MB67 and MB102 block) from Middle Maragheh are virtually identical to the MNHN collection. We refer this more robust lineage herein to aff. H. brachypus and recognise two morphs, not distinguishable at the species level. These morphs are represented in the MNHN, NHMW and BSP assemblages. These two morphs are readily recognisable in the MCIII sample. Morph 1 is distinguished by relatively broader dimensions of mid-shaft width (M3; Fig. 7a) exhibited in the following specimens: MNHN Mar. 3648, MNHN 3649, NHMW A4838, and MNHN Mar. 55/631. This morphology is reminiscent of Pikermi H. brachypus, which is clearly illustrated in Fig. 7a (MCIII log10 ratio) and reinforced by a bivariate plot (Fig. 4b). The second morph exhibits the “Esme Akçakoy Effect” (sensu Bernor et al. 2003) whereby M3 is relatively gracile (narrow) while M4 contrasts in its relatively greater depth (versus M3). The following specimens (Fig. 7a) exhibit this morphology: MNHN Mar. 48/643, BSPm7, MNHN Mar. 46/640, MNHN Mar. 47/649, MNHN Mar. 53/639, NHMW A4840, MNHN Mar. 3110, MNHN Mar. 54/636. We do not know of another taxon that has this combination of cranial and postcranial characters and believe, once again, that this taxon likely had a short biogeographic range of Iran and possibly Turkey. This Maragheh hipparion is not referable to H. prostylum (re: Bernor 1985).
Our description and analyses of H. campbelli demonstrate it to be a genuine member of the Hipparion s.s. clade. This is supported by its absence of a POF (except for a remnant posterior rim), cheek tooth morphology and demonstrably elongate-slender limbs closely comparable to the Mt. Luberon and Saloniki samples of H. prostylum s.s. (Figs. 4b, 5b, 8a and 9c). Likewise, H. campbelli is demonstrably the sister taxon of Samos H. dietrichi which likewise has elongate slender limbs. If Chinese H. hippidiodus from Baode proves to have comparable MPIIIs to these species of Hipparion s.s., then it would represent the furthest eastern extension of the Hipparion s.s. clade. Hipparion s.s. then would have a potential medial Turolian range of southern France, through Greece and Iran to North China. The only other Hipparion clade that has such a demonstrably extensive east–west Eurasian geographic range is Pliocene Plesiohipparion (Bernor and Sun 2015; Bernor et al. 2015).
The genus Cremohipparion is represented at Maragheh by two taxa, Cr. aff. moldavicum and Cr. matthewi. Cr. aff. moldavicum compares very closely to the former Western USSR taxon Cr. moldavicum, apparently differing only in its shorter snout (as represented by only a single specimen, BMNH M3924). The cranium characteristically has a very large, dorsoventrally and medially deep POF placed close to the orbit, with the lacrimal invading the POF. These are hallmark characters of Cremohipparion (Woodburne and Bernor 1980 [their “Group 2”]; Bernor et al. 1980; Bernor et al. 1996). Cr. aff. moldavicum is known to occur in Middle Maragheh and is characterised as having elongate-slender MPIIIs; bivariate plots (M1 versus M11; Fig. 5b) plot all specimens to the left of the Höwenegg ellipse and log10 plots (Fig. 7b) compare very closely to the mean for Sinap Co. sinapensis as well as Pikermi Cr. mediterraneum.
Cr. matthewi is very similar in its MPIII morphology to Cr. aff. moldavicum, but smaller, and would appear to be closely related phylogenetically. Cr. matthewi may well have evolutionarily diverged from Cr. moldavicum. The bivariate plot (Fig. 5b) exhibits very small hipparions that we refer to Cr. matthewi. Log10 ratio plots (Fig. 7c) clearly have a number of FAM (=AMNH) and MNHN specimens that are referable to Cr. matthewi. Maragheh Cr. matthewi has plotted points that are similar in their profile to Sinap Co. sinapensis and Pikermi Cr. mediterraneum but of smaller and more gracile build. Cr. matthewi is known to occur in Lower to Upper Maragheh ranging in age from 8.6 to 7.4 Ma (Mirzaie Ataabadi et al. 2013). There is possibly more than one small clade of hipparions in the Western USSR, Iran, Turkey and the Circum-Mediterranean. Cr. nikosi is an extremely small hipparion from Samos that would appear to vary in the expression of its POF but has extremely long and slender MPIIIs (Bernor and Tobien 1989; Bernor et al. 1996).
Cremohipparion underwent an extensive evolutionary radiation across Eurasia and includes Chinese species Cr. forstenae (late Miocene) and Cr. licenti (early Pliocene), Siwalik Cr. antelopinum (late Miocene, best known from the 8.0 Ma levels), Western USSR-Iran Cr. moldavicum, Greek Cr. macedonicum, Cr. mediterraneum, Cr. proboscideum, Cr. aff. mediterraneum, Cr. nikosi and possibly peri-Mediterraneum Cr. periafricanum (Gromova 1952; Bernor et al. 1996; Koufos 1987). Cremohipparion first occurs in the late Vallesian, ca. 10 Ma (Cr. moldavicum and Cr. macedonicum). All other taxa arise in the Turolian except for the extremely derived Cr. licenti that has highly retracted nasal bones and multiple facial fossae. There are no known Cremohipparion after the early Pliocene of China.
Palaeoecology
Mirzaie Ataabadi et al. (2013) used genus-level faunal resemblance index (GFRI) and ungulate tooth crown height to show the similarity and contrast between Pikermian-type biomes including Maragheh-Pikermi-Samos triad. They found that in the early Vallesian, the Western and Central European localities show little similarity to Maragheh due to them representing forested environments with warm climates and low seasonality. The hypsodonty in these faunas was only due to the occurrence of the invasive species of hipparionine horses (Hippotherium). They showed an adaptation to more closed environments with a significant amount of browse in their diets (Bernor et al. 2003). In remarkable contrast was the later Vallesian “proto-Pikermian” Sinap 12 locality, showing higher similarity and stronger mesodonty and hypsodonty signals. The classical Turolian “Pikermian Chronofaunas” of Samos and Pikermi in Greece show high similarity with Maragheh and had more stable and evenly distributed community structures including brachydonts, mesodonts and hypsodonts. However, Pikermi has a lower percentage of hypsodont forms, which reflects the warm temperate (more) wooded settings at this locality (Solounias et al. 1999). Kostopoulos (2009) also showed that the palaeoecological profile in Pikermi included the tree-dwelling semi-terrestrial primate Mesopithecus, diversified felids and mustelids and browse-dependent proboscideans, rhinos, giraffes and bovids. Abundance of hipparionines like H. brachypus and Cr. mediterraneum supports this environmental setting. These species are more adapted to closed habitats with possibly more browse in their diets. In contrast to Pikermi, Samos has no primate or browse-dependent taxa and has diversified and abundant hipparionine horses including H. brachypus, H. dietrichi, Cr. proboscideum, Cr. aff. matthewi and Cr. nikosi, thus showing many similarities with the Western Asian forms (Vlachou and Koufos 2009; taxa cited herein sensu Bernor, personal observation). It also includes gazelles with grazing rhinos, giraffes and bovids (Kostopoulos 2009). The Maragheh palaeoecological profile is remarkable in this context in having a mixture of Samos and Pikermi characters. Not only primates are present in Maragheh, diversified felids, mustelids and hipparionine horses occur, as well. Browsing and grazing proboscideans, rhinos, giraffes and bovids were also present in Maragheh. Therefore, although Maragheh is more similar to Samos than it is to Pikermi, environmentally, it has more wooded settings in its dominant grass/bushy vegetation than Samos.
Few studies concerned with the palaeodiet of Maragheh hipparionine horses support such environmental settings. Bernor et al. (2014) explored the palaeoecology of Maragheh fauna using mesowear analysis on ungulate maxillary molars present in the MMTT-UCR collection at Howard University, Washington, DC as well as European collections cited herein. The Maragheh hipparions in this study show a spectrum of dietary abrasiveness. Cr. aff. moldavicum and Cr. moldavicum had attritional (browse-dominated) wear features typical of browsers. More intermediate abrasion wear, typical of more balanced mixed feeding diet, was seen in H. prostylum (currently, aff. H. brachypus) and H. campbelli. Some species including H. gettyi, Hipparion sp. and Cr. matthewi exhibited a preference towards increased grass in their diet. The Maragheh bovid species also displayed a range of dietary abrasiveness, whereas the giraffids and chalicotheres had fairly attritional mesowear signals.
Despite small sample sizes for some taxa, their results were consistent with long-held taxon-based interpretations of Maragheh palaeoecology as a warm temperate woodland habitat. This is consistent with those found by Solounias et al. (2010) for Pikermi. Based on the predominance of browsing and mixed feeding types of mesowear signatures, they suggest a mosaic habitat with woodland and some open savanna, rather than a purely grassland or purely closed habitat.
Yamada et al. (2016, this issue) also undertook a mesowear analysis on Maragheh fossil equids and bovids stored in Kyoto University, Japan. They find that hipparionine horses from this collection clustered with extant grazing ruminants and show a narrow and less diverse diet, primarily including grass. They report relatively high percentages of high and round cusps and an absence of blunt cusps in species like H. “prostylum” (= our aff. H. brachypus) and Cr. aff. moldavicum, the two best represented hipparionine horses in the Kyoto collection (Watabe and Nakaya 1991b). These results are somewhat different from previous mesowear studies (Bernor et al. 2014). This apparent inconsistency could be attributed to analytical methodology or simply fossil sampling. The Kyoto collection was excavated from single or close horizons in the middle Maragheh interval. Therefore, local conditions might have characterised these parts of the Maragheh Formation and its fauna. Although they showed no signals of forest vegetation and segregation of dietary niches in the Maragheh fauna, their results do not exclude the existence of wooded habitats in the Maragheh region during the late Miocene. The postcranial anatomy of fossil hipparions from Maragheh also agrees with this palaeoenvironmental interpretation. While species like aff. H. brachypus are characterised by less cursorial, large and robust metapodials suitable for living in more closed habitats, the others mostly have long and slender metapodials adapted for sustained running in more open environments.
A recent study of carbon and oxygen isotopes has shed some new, albeit preliminary results on Maragheh ungulate diet and environments through the 9–7.4-Ma interval (Biasatti et al. 2015). Based on 15 bulk (whole tooth) and 211 serial tooth enamel samples from three species of Maragheh hipparionines (H. gettyi, H. campbelli and Cr. matthewi), it was found that there is a shift in carbon isotope values from C4 to C3 ecosystems and that there is a positive correlation between δ13C and δ18O values prior to ~8.2 Ma. This new evidence suggests that the Iranian Plateau was rising during the interval of time recorded by the Maragheh strata. More work is needed on the entire MMTT Maragheh ungulate enamel and associated palaeosols to better resolve this hypothesis.
Conclusions
Morphological analyses of postcranial hipparionine material from Maragheh along with previous and current studies on cranial and dental material have allowed us to characterise the following species in the Maragheh assemblage: “H.” gettyi, aff. H. brachypus, H. campbelli, Cr. aff. moldavicum and Cr. matthewi. These species are arranged in three successive biostratigraphic intervals with clear chronological differences: H. gettyi defines the late Vallesian Lower Maragheh horizon, aff. H. brachypus and Cr. aff. moldavicum define early Turolian Middle Maragheh levels and H. campbelli defines the late early Turolian Upper Maragheh interval. Cr. matthewi is present through all biostratigraphic intervals at Maragheh. The proposed taxonomic and biostratigraphic resolution here is slightly different from previous studies especially in the recognition of aff. H. brachypus instead of H. prostylum. The Maragheh hipparion assemblages are correlated with those from Turkey, Greece, the Balkans and Black Sea region. At the same time, they are clearly very different from those of central Asia and China. The Maragheh hipparionine horses indicate a clear niche differentiation based on palaeodiet studies indicative of different environmental adaptations.
References
Abich, H. W. (1858). Tremblement de terre observe a Tebriz en Septembre 1856, notices physiques et geographiques de M Khanykof sur 1’Azerbeidjan. Bulletin de 1’Académie (Imperiale) des Sciences de Saint Petersbourg, Classe Physico-Mathmatique, 382 XVI-22 notes 27,339-352.
Armour-Chelu, M., & Bernor, R. L. (2011). Equidae. In T. Harrison (Ed.), Geology and paleontology of Laetoli (pp. 295–326). New York: Springer.
Berggren, W. A., & Van Couvering, J. A. (1974). The late Neogene: biostratigraphy, geochronology and paleoclimatology of the last 14 million years in marine and continental sequences. Palaeogeography, Palaeoclimatology, Palaeoecology, 16(1–2), 1–216.
Bernor, R. L. (1978). The mammalian systematics, biostratigraphy and biochronology of Maragheh and its importance for understanding Late Miocene hominoid zoogeography and evolution. Los Angeles: University of California dissertation.
Bernor, R. L. (1985). Systematic and evolutionary relationships of the hipparionine horses from Maragheh, Iran (Late Miocene, Turolian age). Palaeovertebrata, 15(1985), 173–269.
Bernor, R. L. (1986). Mammalian biostratigraphy, geochronology and zoogeographic relationships of the late Miocene Maragheh fauna, Iran. Journal of Vertebrate Paleontology, 6(1), 76–95.
Bernor, R. L., & Armour-Chelu, M. (1999). Toward an evolutionary history of African hipparionine horses. In T. Brommage, & F. Schrenk (Eds.), African biogeography, climate change and early hominid evolution, Wenner-Gren Foundation Conference, Livingstonia Beach Hotel, Salima Malawi (pp. 189–215). Oxford: Oxford.
Bernor, R. L., & Haile Selassie, Y. (2009). Equidae. In Y. Haile-Selassie & G. Woldegabriel (Eds.), Ardipithecus kadabba: late Miocene evidence from the Middle Awash, Ethiopia (pp. 397–428). Berkeley: University of California Press.
Bernor, R. L., & Harris, J. (2003). Systematics and evolutionary biology of the Late Miocene/Early Pliocene hipparionine horses from Lothagam, Kenya. In M. Leakey & J. Harris (Eds.), Lothagam: the dawn of humanity in eastern Africa (pp. 387–438). New York: Columbia University Press.
Bernor, R. L., & Hussain, S. T. (1985). An assessment of the systematic, phylogenetic and biogeographic relationships of Siwalik hipparionine horses. Journal of Vertebrate Paleontology, 5(1), 32–87.
Bernor, R. L., & Scott, R. S. (2003). New interpretations of the systematics, biogeography and paleoecology of the Sahabi hipparions (latest Miocene) Libya. Geodiversitas, 25(2), 297–319.
Bernor, R. L., & Sun, B. (2015). Morphology through ontogeny of Chinese Proboscidipparion and Plesiohipparion, observations on their Eurasian and African relatives. Vertebrata Pal Asiatica, 53(1), 73–92.
Bernor, R. L., & Tobien, H. (1989). Two small species of Cremohipparion (Mammalia Equidae) from Samos, Greece. Bayerische Staatssammlung fur Paläontologie und Historische Geologie Mitteilungen, 29(1989), 207–226.
Bernor, R. L., & White, T. D. (2009). Systematics and biogeography of “Cormohipparion” africanum, Early Vallesian (MN 9 ca 105 Ma) of Bou Hanifia, Algeria. In L. Barry Albright III (Ed.), Papers on geology, vertebrate paleontology and biostratigraphy in honor of Michael O Woodburne (pp. 635–658). Arizona: Museum of Natural History.
Bernor, R. L., Tobien, H., & Van Couvering, J. A. (1979). The mammalian biostratigraphy of Maragheh. Annales Géologie des Pays Hellenica, 1(1979), 91–99.
Bernor, R. L., Woodburne, M. O., & Van Couvering, J. A. (1980). A contribution to the chronology of some Old World faunas based on hipparionine horses. Geobios, 13(5), 25–59.
Bernor, R. L., Qiu, Z., & Tobien, H. (1987). Phylogenetic and biogeographic bases for an Old World hipparionine horse geochronology. Proceedings of the VIII International Congress of the Regional Committee on Mediterranean Neogene Stratigraphy. Annales Instituti Geologici Publici Hungarici, 70(1987), 43–53.
Bernor, R. L., Tobien, H., & Woodburne, M. O. (1989). Patterns of Old World hipparionine evolutionary diversification. In E. Lindsay, V. Fahlbusch, & P. Mein (Eds.), European Neogene mammal chronology (pp. 263–319). New York: Plenum Press.
Bernor, R. L., Qiu, Z., & Hayek, L. A. (1990). Systematic revision of Chinese Hipparion species described by Sefve, 1927. American Museum Novitates, 2984(1990), 1–60.
Bernor, R. L., Koufos, G., Woodburne, M., & Fortelius, M. (1996). The evolutionary history and biochronology of European and Southwest Asian Late Miocene and Pliocene hipparionine horses. In R. L. Bernor, V. Fahlbusch, & H. W. Mittmann (Eds.), The evolution of Western Eurasian Neogene mammal faunas (pp. 7–46). New York: Columbia University Press.
Bernor, R. L., Tobien, H., Hayek, L. A., & Mittmann, H. W. (1997). The Höwenegg hipparionine horses: systematics, stratigraphy, taphonomy and paleoenvironmental context. Andrias, 10(1997), 1–230.
Bernor, R. L., Scott, R. S., Fortelius, M., Kappelman, J., & Sen, S. (2003). Systematics and evolution of the Late Miocene hipparions from Sinap, Turkey. In M. Fortelius, J. Kappelman, S. Sen, & R. L. Bernor (Eds.), The geology and paleontology of the Miocene Sinap Formation, Turkey (pp. 220–281). New York: Columbia University Press.
Bernor, R. L., Kaiser, T. M., & Nelson, S. V. (2004). The oldest Ethiopian hipparion (Equinae, Perissodactyla) from Chorora: systematics, paleodiet and paleoclimate. Courier Forschungsinstitut Senckenberg, 246(2004), 213–226.
Bernor, R. L., Scott, R. S., & Haile-Selassie, Y. (2005). A contribution to the evolutionary history of Ethiopian hipparionine horses: morphometric evidence from the postcranial skeleton. Geodiversitas, 27(1), 133–158.
Bernor, R. L., Armour-Chelu, M., Gilbert, H., Kaiser, T., & Schulz, E. (2010). Equidae. In L. Werdelin & B. Sers (Eds.), Cenozoic mammals of Africa (pp. 685–721). Berkeley: University of California Press.
Bernor, R. L., Kaiser, T. M., Nelson, S. V., & Rook, L. (2011). Systematics and paleobiology of Hippotherium malpassi n. sp from the latest Miocene of Baccinello V3 (Tuscany Italy). Bolletino della Societa Paleontologica Italiana, 50(3), 175–208.
Bernor, R. L., Boaz, N. T., & Rook, L. (2012). Eurygnathohippus feibeli (Perissodactyla: Mammalia) from the late Miocene of Sahabi (Libya) and its evolutionary and biogeographic significance. Bolletino della Societa Paleontologica Italiana, 51(1), 39–48.
Bernor, R. L., Gilbert, H., Semprebon, G., Simpson, S., & Semaw, S. (2013). Eurygnathohippus woldegabrieli sp. nov. (Perissodactyla: Mammalia) from the Middle Pliocene of Aramis, Ethiopia (4.4 Ma). Journal of Vertebrate Paleontology, 33(6), 1472.
Bernor, R. L., Semprebon, G., & Damuth, J. (2014). Maragheh ungulate mesowear: interpreting paleodiet and paleoecology from a diverse fauna with restricted sample sizes. Annales Zoologici Fennici, 51(1–2), 201–208.
Bernor, R. L., Sun, B., & Chen, Y. (2015). Plesiohipparion shanxiense n. sp. from the Early Pleistocene (Nihowanian) of E Shanxi, China. Bolletino della Societa Paleontologica Italiana, 54(3), 197–210.
Biasatti, D. M., Bernor, R. L., & Cooper, L. W. (2015). Insights on late Miocene climate changes and regional uplift in Maragheh Basin, eastern Azarbaijan Province, northwest Iran revealed by stable oxygen isotope analyses of fossil horse tooth enamel. Society of Vertebrate Paleontology, Program and Abstracts (2015), 89.
Brandt, J. F. (1870). Ueber die von Herrn Magister Adolph Goebel auf seiner persischen Reise bei der Stadt Maragha in der Provinz Azerbeidjan gefundenen Säugethier-Reste. Denkschrift des Naturforscher-Vereins zu Riga, 1–8.
Bukhsianidze, M. (2015). Catalogue of the holotypes of fossil land mammals in the Georgian National Museum. Tbilisi: Georgian National Museum.
Campbell, B. G., Amini, M. H., Bernor, R. L., Dickenson, W., Drake, W., Morris, R., Van Couvering, J. A., & Van Couvering, J. A. H. (1980). Maragheh: a classical late Miocene vertebrate locality in northwestern Iran. Nature, 287(5785), 837–841.
Christol, J. (1832). Description d’Hipparion. Annals Science Industry Midi France, 1(1832), 180–181.
Eisenmann, V. (1995). What metapodial morphometry has to say about some Miocene hipparions. In E. S. Vrba, G. H. Denton, T. C. Partridge, & L. H. Burckle (Eds.), Palaeoclimate evolution with emphasis on human origins (pp. 48–162). New Haven: Yale University Press.
Eisenmann, V., Alberdi, M. T., de Giuli, C., & Staesche, U. (1988). Methodology. In M. Woodburne & P. Y. Sondaar (Eds.), Studying fossil horses (pp. 1–71). Leiden: EJ Brill Press.
Erdbrink, D. P. B., Priem, H. N. A., Hebeda, E. H., Cup, C., Dankers, P. O., & Cloetingh, S. A. P. L. (1976). The bone bearing beds near Maragheh in NW Iran. Proceedings Koninklijke Nederlandse Akademie van Wetenschappen B, 79(2), 85–113.
Fahlbusch, V. (1991). The meaning of MN zonations: considerations for a subdivision of the European continental Tertiary using mammals. Newsletters on Stratigraphy, 24(1991), 159–73.
Forsten, A. (1968). Revision of the Palearctic Hipparion. Acta Zoologica Fennica, 119(1968), 1–134.
Forsten, A. (1983). The preorbital fossa as a taxonomic character in some Old World Hipparion. Journal of Paleontology, 57(4), 686–704.
Gabunia, L. K. (1959). Istorii Gipparionov (po materialam iz neogena SSSR). Tbilisi: USSR Academy of Science.
Gaudry, A. (1873). Animaux fossiles du Mont Léberon (Vaucluse): Etude sur les Vertébrés. Paris: Savy.
Gervais, P. (1849). Note sur la multiplicité des espèces d’ Hipparion (genre de chevaux à trois digits) qui sont enfouis à Cucuron (Vaucluse). Comptes Rendus Academi Sciences Paris D, 29, 284–286.
Gervais, P. (1859). Zoologie et Paléontologie française (Animaux Vertébrés). Paris.
Gilbert, H. & Bernor, R. L. (2008) Equidae. In W. H. Gilbert & B. Asfaw (Eds.) Homo erectus – Pleistocene Evidence from the Middle Awash, Ethiopia (pp. 133–166). Berkeley: University of California Press.
Grewingk, C. (1881). Ueber fossile Saugethiere von Maragha in Persien. Verhandlungen der K. K. Geologischen Reichsanstalt (1881), 296.
Gromova, V. (1952). Le genre Hipparion. Bulletin Recherche Géologie et Minerologie CEDP, 12(1952), 1–288.
Kamei, T. J., Ikeda, H., Ishida, S., Ishida, L., Onishi, H., Partoazar, H., Sasajima, S., & Nishimura, S. (1977). A general report of the geological and paleontological survey in Maragheh area, north-west Iran. Memoirs of the Faculty of Science, Kyoto University, 43(1–2), 131–164.
Kittl, E. (1887). Beiträge zur Kenntniss der fossilen Saugethiere von Maragha in Persien, 1 Carnivoren. Annalen des Naturhistorischen Museums Wien, 11(1887), 317–338.
Kostopoulos, D. S. (2009). The Pikermian event: temporal and spatial resolution of the Turolian large mammal fauna in SE Europe. Palaeogeography, Palaeoclimatology, Palaeoecology, 274(1–2), 82–95.
Koufos, G. D. (1987). Study of the Pikermi hipparions. Bulletin du Muséum national d’Histoire naturelle Paris, (4e sér., 9, sect. 3), 327–363.
Koufos, G. D., & Vlachou, D. (2005). Equidae (Mammalia Perissodactyla) from the late Miocene of Akkasdagi Turkey. Geodiversitas, 27(4), 633–705.
Koufos, G. D., & Vlachou, T. D. (2016). Equidae. In G. D. Koufos & D. S. Kostopoulos (Eds.), Palaeontology of the upper Miocene vertebrate localities of Nikiti (Chalkidiki Peninsula, Macedonia, Greece) (Geobios 49 (pp. 85–118). Lyon: Elsevier Masson SAS.
Lydekker, R. (1886). On the fossil Mammalia of Maragha in northwest Persia. Quarterly Journal of the Geological Society of London, 42(1–4), 173–176.
MacFadden, B. J., & Woodburne, M. O. (1982). Systematics of the Neogene Siwalik hipparions (Mammalia Equidae) based on cranial and dental morphology. Journal of Vertebrate Paleontology, 2(2), 185–218.
MacFadden, B. J. (1980). The Miocene horse Hipparion from North America from the type locality in southern France. Palaeontology, 23(3), 617–635.
MacFadden, B. J. (1984). Systematics and phylogeny of Hipparion, Neohipparion, Nannippus, Cormohipparion (Mammalia Equidae) from the Barstovian (medial Miocene), Texas Gulf Coastal Plain. Journal of Paleontology, 58(2), 185–218.
Mecquenem, R. (1908). Contribution à l’étude du gisement des vertebres de Maragha et de ses environs. Annales d’Histoire Naturelle, 1(1908), 27–79.
Mecquenem, R. (1924–25). Contribution à l’étude des fossiles de Maragha. Annales de Pa1éonto1ogie, 13/14(1924–25), 135–160.
Mirzaie Ataabadi, M., Bernor, R. L., Kostopolus, D., Wolf, D., Orak, Z., Zaree, G., Nakaya, H., Watabe, M., & Fortelius, M. (2013). Recent advances on paleobiological research of the Late Miocene Maragheh fauna, northwest Iran. In X. Wang, M. Fortelius, & L. Flynn (Eds.), Asian Neogene mammal biostratigraphy and chronology (pp. 544–563). New York: Columbia University Press.
Mirzaie Ataabadi, M., Mohammadalizadeh, J., Zhang, Z., Watabe, M., Kaakinen, A., & Fortelius, M. (2011). Late Miocene large mammals from Ivand (northwestern Iran). Geodiversitas, 33(4), 709–728.
Osborn, H. F. (1918). Equidae of the Oligocene, Miocene and Pliocene of North America: iconographic type revision. American Museum of Natural History Memoirs, 2(1918), 1–326.
Pohlig, H. (1886). On the Pliocene of Maragha, Persia and its resemblance to that of Pikermi in Greece; on fossil elephant remains of Caucasia, Persia; on the results of a monograph of the fossil elephants of Germany and Italy. Quarterly Journal of the Geological Society of London, 42(1–4), 177–182.
Qiu, Z. X., Huang, W. L., & Guo, Z. H. (1987). The Chinese hipparionine fossils. Palaeontologica Sinica, C25(1987), 1–243.
Rodler, A. (1890). Ueber Urmiatherium polaki n. gen n. sp., einen neuen Sivatheriiden aus dem Knochenfelde von Maragha. Denkschriften K. Akademie der Wissenschaften, Wien; Mathematisch-Naturwissenschaftliche Klasse, 56(1890), 315–332.
Rodler, A., & Weithofer, K. A. (1890). Die Wiederkauer der Fauna von Maragha. Denkschriften K. Akademie der Wissenschaften, Wien; Mathematisch Naturwissenschaftlichen Klasse, 62(1890), 753–772.
Roth, J. & Wagner, A. (1855) Die fossilen Knochenueberreste von Pikermi in Griechenland. Abhandlungen der bayerischen Akademie der Wissenschafen, Munich, 7, 371–464.
Sakai, T., Zaree, G.R., Sawada, Y., Mirzaie Ataabadi, M. & Fortelius, M. (2016). Depositional environment reconstruction of the Maragheh Formation, East Azarbaijan, northwestern Iran. In M. Mirzaie Ataabadi and M. Fortelius (Eds.) The late Miocene Maragheh mammal fauna; results of recent multidisciplinary research. Palaeobiodiversity and Palaeoenvironments, 96(3). Doi:10.1007/s12549-016-0238-z.
Sawada, Y., Zaree, G.R., Sakai, T., Itaya, T. Yagi, K., Imaizumi, M., Mirzaie Ataabadi, M., & Fortelius, M. (2016). K–Ar ages and petrology of the late Miocene pumices from the Maragheh Formation, northwest Iran. In M. Mirzaie Ataabadi and M. Fortelius (Eds.) The late Miocene Maragheh mammal fauna; results of recent multidisciplinary research. Palaeobiodiversity and Palaeoenvironments, 96(3). Doi:10.1007/s12549-016-0232-5.
Schlesinger, G. (1917). Die Mastodonten des k. k. Naturhistorischen Hofmuseums. Morphologische- Phylogenetische Untersuchungen. Denkschriften K. Akademie der Wissenschaften, Wien; Mathematische-Naturwissenschaftliche Klasse, 1(1917), 1–230.
Skinner, M. F., & MacFadden, B. J. (1977). Cormohipparion n. gen. (Mammalia, Equidae) from the North American Miocene (Barstovian- Clarendonian). Journal of Paleontology, 51(4), 912–926.
Solounias, N., Plavcan, J. M., Quade, J., & Witmer, L. (1999). The paleoecology of the Pikermian biome and the Savanna myth. In J. Agustí, L. Rook, & P. Andrews (Eds.), The evolution of Neogene terrestrial ecosystems in Europe (pp. 436–453). Cambridge: Cambridge University Press.
Solounias, N., Rivals, F., & Semprebon, G. M. (2010). Dietary interpretation and paleoecology of herbivores from Pikermi and Samos (late Miocene of Greece). Paleobiology, 36(1), 113–136.
Solounias, N., & Danowitz, M. (2016). The Giraffidae of Maragheh and the identification of a new species of Honanotherium. In M. Mirzaie Ataabadi and M. Fortelius (Eds.) The late Miocene Maragheh mammal fauna; results of recent multidisciplinary research. Palaeobiodiversity and Palaeoenvironments, 96(3) doi:10.1007/s12549-016-0230-7.
Sondaar, P. Y. (1974). The Hipparion of the Rhone valley. Geobios, 7(2), 289–306.
Suwa, G., Beyene, Y., Nakaya, H., Bernor, R. L., et al. (2015). Newly discovered cercopithecid, equid and other mammalian fossils from the Chorora Formation, Ethiopia. Anthropological Science, 123(1), 19–39.
Swisher, C. C., III. (1996). New 40Ar/39Ar dates and their contribution toward a revised chronology for the Late Miocene of Europe and West Asia. In R. L. Bernor, V. Fahlbusch, & H. W. Mittmann (Eds.), The evolution of Western Eurasian Neogene mammal faunas (pp. 64–77). New York: Columbia University Press.
Takai, F. (1958). Vertebrate fossils from Maragha. Institute of Oriental Culture, University of Tokyo, 26 (1958), 7–11.
Tobien, H. (1968). Palaeontologische Ausgrabungen nach Jungtertiären Wirbeltieren auf der Insel Chios (Griechenl) und bei Maragheh (NW Iran). Jahrbuch der Vereiningung “Freunde der Universität Mainz”, (1968), 51–58.
Vlachou, T. D., & Koufos, G. D. (2009). The Late Miocene mammal faunas of the Mytilinii Basin, Samos Island, Greece: new collection, 11 Equidae. Beiträge zur Paläontologie, 31(2009), 207–281.
Watabe, M., & Nakaya, H. (1991a). Phylogenetic significance of the postcranial skeletons of the hipparions from Maragheh (late Miocene), NW Iran. Memoirs of the Faculty of Sciences, Kyoto University, 56(1–2), 11–53.
Watabe, M., & Nakaya, H. (1991b). Cranial skeletons of Hipparion (Perissodactyla, Mammalia) from Maragheh (Turolian late Miocene), NW Iran. Memoirs of the Faculty of Sciences, Kyoto University, 56(1–2), 55–125.
Wolf, D., Bernor, R. L., & Hussain, S. T. (2013). Evolution, biostratigraphy and geochronology of Siwalik hipparionine horses. Palaeontographica, 300(2013), 1–118.
Woodburne, M. O., & Bernor, R. L. (1980). On superspecific groups of some Old World hipparionine horses. Journal of Paleontology, 54(6), 1319–1348.
Woodburne, M. O., MacFadden, B. J., & Skinner, M. (1981). The North American “Hipparion Datum” implications for the Neogene of the Old World. Geobios, 14(1), 1–32.
Woodburne, M. O. (1989). Hipparion horses: a pattern of endemic evolution and intercontinental dispersal. In D. R. Prothero & R. M. Schoch (Eds.), The evolution of perissodactyls (pp. 197–233). New York: Oxford University Press.
Woodburne, M. O., Theobald, G., Bernor, R. L., Swisher, C. C., III, König, H., & Tobien, H. (1996). Advances in the geology and stratigraphy at Höwenegg, southwestern Germany. In R. L. Bernor, V. Fahlbusch, & H. W. Mittmann (Eds.), The evolution of Western Eurasian Neogene mammal faunas (pp. 106–123). New York: Columbia University Press.
Woodburne, M. O. (2007). Phyletic diversification of the Cormohipparion occidentale Complex (Mammalia: Perissodactyla, Equidae) Late Miocene North America and the origin of the Old World Hippotherium datum. Bulletin of the American Museum of Natural History, 306(2007), 1–138.
Yamada, E., Hasumi, E., Miyazato, N., Akahoshi, M., Watabe, M., & Nakaya, H. (2016). Mesowear analyses of sympatric ungulates from the late Miocene Maragheh, Iran. In M. Mirzaie Ataabadi and M. Fortelius (Eds.) The late Miocene Maragheh mammal fauna; results of recent multidisciplinary research. Palaeobiodiversity and Palaeoenvironments, 96(3). doi:10.1007/s12549-016-0237-0.
Acknowledgements
MMA’s visits to MNHN, Paris, and Howard University were partially supported by a grant from the NSF RHOI (Revealing Hominid Origin Initiative; BCS-0321893) project funded to Professors F. Clark Howell and Tim White. Bernor and Wolf also wish to acknowledge the support from RHOI and NSF grants EAR1113175, EAR1138908 and EAR1333230 for this research undertaking. We thank Pascal Tassy and Christine Argot and the MNHM staff for their help during our visits to Paris. Bernor thanks Fred Roegl and Fritz Steininger for their assistance studying the Maragheh material in Vienna. MMA also thank Mahito Watabe (former Hayashibara Museum, Okayama), Hideo Nakaya (Kagoshima University), Ursula Göhlich (NHMW), Gertrud Rössner (BSPG) and Wilma Wessels (IvAU) for facilitating his visits. We thank two anonymous reviewers who improved our work.
Author information
Authors and Affiliations
Corresponding author
Additional information
This article is a contribution to the special issue “The late Miocene Maragheh mammal fauna; results of recent multidisciplinary research”
Electronic supplementary material
Below is the link to the electronic supplementary material.
ESM 1
(XLSX 45 kb)
Rights and permissions
About this article

Cite this article
Bernor, R.L., Mirzaie Ataabadi, M., Meshida, K. et al. The Maragheh hipparions, late Miocene of Azarbaijan, Iran. Palaeobio Palaeoenv 96, 453–488 (2016). https://doi.org/10.1007/s12549-016-0235-2
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s12549-016-0235-2