
Abstract
In order to counteract heavy foreland losses in the lower reaches of the Weser estuary, a system of sedimentation fields surrounded by brushwood groynes was constructed in 1958. Over the next forty years, the land accretion process was monitored by surveys of bottom-surface elevation, sedimentology, development of vegetation cover and changes in bottom-fauna composition. From the viewpoint of coastal protection, the sedimentation fields achieved a continuous growth of half a metre within forty years and successfully reduced the regression of the eroding shoreline. The ecological results represent a documentation comprising all succession stages from a bare tidal flat to a vegetated salt marsh. However, though the larger parts of the fields had already exceeded mean high-water level at the final survey of 1997, aquatic elements were still present in the vegetation as well as in the bottom fauna. The lack of a seaward inclination was most likely the reason, why saline water from spring tides and storm surges remained enriched in the soil. With growing elevation and condensing vegetation, each of the fields had developed a branched creek system and a varied geomorphological relief. Their habitat and species diversity proved to be equal to naturally grown mainland salt marshes.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
The reasons for conducting this study go back to the 1950s: A section of shoreline in the lower reaches of the Weser estuary was suffering from severe land losses of an old salt marsh. In some places, an eroding cliff had already been approaching the summer dike as shown in Fig. 1. After a number of base-line studies (Homeier 1958: Historic morphology; Köritz 1958: Hydrology; Müller 1958: Sediments and biology), the Coastal Research Station Norderney recommended protecting the endangered sites by using a system of land reclamation fields, which was implemented already in 1958. The expectation was that the process of land accretion induced by this means would lower the effects of current and wave attack and prevent further erosion.

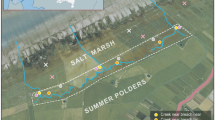
Study area in the lower reaches of the Weser estuary with the land reclamation fields constructed in 1958
From the beginning, it had unanimously been agreed by the coastal engineers and the Coastal Research Station, to use this opportunity to conduct a complete study of the salt marsh formation process until the status of a new salt marsh could be reached. Certainly, nobody had expected that this process would have taken half a century. Two interim reports have been submitted on the state of development until 1961 (Müller 1962) and until 1978 (Grotjahn et al. 1983). The aspects of plant sociology and environment protection (Arens et al. 2003) and of geomorphology (Ladage and Stephan 2007) have been recorded in particular accounts.
To document and to understand the mutual physical and biological effects of a salt marsh formation process were the original research questions. Today, in the face of climate change and accelerated sea-level rise, sedimentation fields are of relevant interest not only for coastal protection but also for nature conservation.
Material and methods
The area concerned is situated on the southern and northern side of the small inlet Cappeler Tief at the eastern shore of the Weser estuary (Fig. 1). The eleven land reclamation fields constructed in 1958 cover a length of 4 km and a surface area of roughly 1 km2. The fields were surrounded by brushwood groynes with an opening in the front groyne allowing tidal waters to flow in and out. After the violent storm surge of 1962 and the destructive ice period of the winter 1962/63, the front groyne was reconstructed by using natural stone. In the first years, a system of draining trenches was maintained. However, since 1962, these activities have been discontinued, enabling the development of a quasi-natural salt marsh.
Information on hydrological conditions is provided by a gauge in the neighbouring harbour Spiekaer Tief. The mean tidal range in the area is about 3.0 m. The mean high water (MHW) in the period 1961–1965 amounted to 1.46 m above mean sea level (MSL) and rose to MSL + 1.61 m between 1991 and 1995. The salinity levels in this area lie within polyhaline ranges (Müller 1962). The nearshore seston contents in the area itself and in the area adjoining to the south amounted to a few hundred mg/l, which reflects a poor to moderate supply (Müller 1962, 1963). The site is exposed to wind and wave action prevailing from the west.
At the beginning of regular assessments conducted in 1960 within and outside the land reclamation fields, an investigation design was established which mostly had been maintained until the final survey of 1997. Bottom-surface elevation was measured in 28 transects, four transects in seven of the eleven fields and additionally on 10 points outside (Fig. 1). In total, from 1960 through 1997, ten surveys were carried out, until 1984 by levelling methods, in 1989 by tachymeter surveys and in 1997 by using GPS-techniques. From the data derived at distances of ten metres, the mean values were calculated for each of the investigated fields, for the groups of the northern (Nos. VI, VIII, X and XI) and southern fields (Nos. I, III and V) and for the total area. The values of ditches and channels, of course, remained unconsidered. After a branched creek system had developed in the 1970s, aerial-photograph analysis was used for mapping the surface structures. The campaigns for measuring topography were carried out independently from seasons, while samplings and surveys of sediments, vegetation and bottom fauna were limited to the period from June to August.
Sedimentology was investigated at 30 permanent plots, one outside in front of each of the fields numbered I to X and two plots inside (Fig. 1). Sediment sampling was not used in the 1970 survey, and the sampling in 1977/78 was limited to the inside of the fields (30 stations different from the permanent plots). Samples of sediments were taken from the surface down to 5 cm depth. Using the Atterberg method, analysis of grain-size distribution started in 1957. However, after 1961, this was changed to the areometer method. Because of unsatisfactory comparability, only the results gained by the latter method are represented as accumulated curves of grain-size distribution. As additional parameters, lime content (CaCO3 after Scheibler) and water content (% of wet weight) were determined, and in 1985 and 1997 also the content of water-soluble chlorides (dried sediment diluted with aquadest 1:5, titration after Mohr-Knudsen) was measured.
Until the end of the 1960s, vegetation was assessed by descriptive methods only. In the course of the 1970s, a branched creek system and a surface relief had developed, which resulted in an increasingly diversified vegetation cover. Therefore, a more thorough assessment was required by increased station numbers and by analysis of aerial photographs. Three series of photographs became available from contemporary projects: from the summer of 1978, infrared-false colour diapositives, from the summer of 1985, a set of black and white photo copies, and from the summer of 1997, colour-infrared diapositives. After standardising and georeferencing the different images, the map contents were transferred to a geographic information system (details given by Arens et al. 2003). The analysis comprised the topographic situation including the creek systems and the foreland border. The colour and the structure characteristics were used to delimit the areas of open water, uncovered tidal flats and vegetation types. Thereafter, the proportions of the different units were determined by planimetry. By these means, the surface area could be subdivided into a number of units, mainly related to geomorphological structures, viz
-
Bare tidal flats without overgrowth of vascular plants
-
Low-lying areas with sparse plant covering less than 50%
-
Plane areas of high elevations with dense plant covering 50 to 100% (with different subunits)
-
Elevated bank ridges
-
Sparsely covered fringe under the eroding cliff
-
Areas developed by technical land reclamation due to the activities of a farmer since the 1980s
The farmer started with digging trenches until nearly to the front groyne of the northern fields VIII and IX. Later, between 1985 and 1988, he extended his activities down to the fields VI and VII and northward to the edge of field X (Arens et al. 2003).
Furthermore, another factor began affecting the vegetation: grazing. The technically reclaimed fields (and also nearshore zones of some other fields) were opened to different intensity of sheep, cattle and horse grazing, as described by Arens et al. (2003).
The ground-truth investigations included the description of localities, the total vegetation cover (%), the coverage of individual plant species (%) and the presence of rare species.
The invertebrate bottom fauna was investigated at the 30 permanent plots inside (20) and in front of the fields (10), but at places of particular interest, additional samples were taken. In his baseline study of 1957, Müller (1958) took samples of 0.25 m2, 30 cm deep, extracting the fauna by sieving with 1 mm mesh size. This procedure was continued until 1965. The survey of 1970 was restricted to qualitative sampling. Since 1977/78, methods have changed because of the increased proportions of small species like oligochaetes and collembolans. Tubes of 177 cm2 cross-sectional area and 30 cm length were used; per station, three samples were taken inside and seven samples outside the fields. Instead of sieving, the fauna was sorted in live state and after species identification and counting the material was frozen. Ash-free dry weight (ADW) was determined after return to the home laboratory.
Results
Surface elevation
The measurement of bottom-surface elevations in 1957, before the construction of land reclamation fields, resulted in values of about 1.0 m above mean sea level, which is approximately half a metre below MHW. In 1960, when the regular measuring programme was started, the mean elevation within the fields had slightly increased to MSL + 1.12 m. The surface was still plane and not yet subdivided by creeks, as shown by the examples of transects 2 and 22 in Fig. 2. The increase of elevation went more or less constantly with average annual rates of 1.6 cm. In the early 1970s, the trend of increasing values for surface elevations (Fig. 3) seemed to have slowed down significantly. However, as evident from the transects of 1970 and 1976 (Figs. 2 and 4), just in this phase, a decisive step of development took place: A branched creek system had developed in each of the fields, subdivided them and established a morphological relief of contrasting elevations (Figs. 4 and 5). As evident from Fig. 3, the southern fields since the 1980s had increasingly surpassed the northern ones, in spite of the rigorous land reclamation efforts in the fields Nos. VI to IX.

Two examples of change in bottom-surface elevation from 1960 to 1997, transects number 2 (field I) and number 22 (field VIII)

Growth of bottom-surface elevation in the land reclamation fields. Separate curves based on mean values of the southern fields, the northern fields and the whole area

Example of a selected transect in field VI: growth of elevation, relief formation and development of vegetation

Surface morphology and vegetation cover in field VII, obtained by analysis of aerial photographs from 1978, 1985 and 1997 and by ground-truth investigation. Increased farming land use changed the morphology drastically between 1985 and 1997
During the period of 1960 till 1997, the growth of elevation resulted in an increase of 0.59 m (average 1960: MSL + 1.12 m; 1997: MSL + 1.71 m). Thus, the average for 1997 had already exceeded the mean tidal high-water mark by about 0.10 m (period 1991–1995: MSL + 1.61 m according to the gauge of Spiekaer Tief). Taking in consideration that this occurred over a period of almost forty years, it has actually to be reduced by the sea-level rise of 0.15 m during this period. Thus, in the average of all fields, the remaining elevation growth amounts to only 0.44 m.
Furthermore, it has to be emphasised that elevation within the fields did not increase evenly. Larger and medium-sized creeks were still lined by low-lying fringes at the beginning (Fig. 5). Later, in the 1980s, they were becoming zones of preferred growth caused by the sediment-catching effect of vegetation. This mechanism gave rise to the origin of elevated bank ridges, which essentially contributed to the existence of a geomorphological relief.
To follow the dynamics of the eroding shoreline (Fig. 6), a topographical map drawn by the Coastal Research Station on a scale of 1:5000 was used, based on levelling of the year 1965. In this map, the trace of the eroding cliff is precisely represented and allows comparisons with later situations based on analysis of aerial photography. The results reveal that until 1976, the cliff in the southern fields suffered an abrasion of 3 to 8 m. In the northern fields, the losses in some wedges intruding the foreland reached up to 22 m. In the following period up to 1997, the changes indicate only small proportions of regressing erosion. By 2006, Ladage and Stephan (2007) realised an annual loss of 0.3 m in the southern fields and a standstill of the break off in the northern fields.

View of the eroding cliff in field I, summer 1978
Sediments
In the situation of 1957, before the set-up of the reclamation fields, the tidal flat bottom was described as well-sorted fine-sandy sediment (Müller 1958). Afterward, sediment composition and related parameters developed differently outside and inside the fields (Fig. 7). While the grain size distribution in front of the fields retained their fine-sandy character, inside the fields, the sediments developed to a sandy, strongly silty mud on the way to a soil, of which the surface layer was densely interspersed with roots of salt marsh plants. Together, with clay and silt also, the lime and water content increased considerably (Fig. 8). As regards water-soluble chlorides (measured only 1985 and 1997), the values inside are remarkably higher than outside the fields:
-
Inside 1985: 868 mg ± 231 mg /100 g
-
Inside 1997: 1412 mg ± 555 mg /100 g
-
Outside 1985: 548 mg ± 100 mg /100 g
-
Outside 1997: 640 mg ± 125 mg /100 g

Change of surface sediments outside and inside the fields. Accumulated curves of grain-size distribution from selected years

Changes in the sediment parameters water content (% of wet weight) and lime content (% CaCO3 of dry weight)
Vegetation
Before construction of the land reclamation fields in 1957, the tidal flats in front of the eroding cliff were free of vascular-plant vegetation except the occasional occurrence of Zostera noltei, Salicornia sp., Spartina anglica and Tripolium pannonicum. Since 1958, a vegetation cover developed gradually inside the newly constructed fields.
The plant species encountered inside and outside the fields are given in Table 1 (only vascular plants), arranged according to their time of appearance. Until 1965, the pioneer vegetation of Salicornia, Spartina and Tripolium predominated in low coverage. In the course of the 1970s, the fields made a rather essential step: Because the condensed vegetation hampered the water from easily flowing over the plains, a system of branched creeks had originated in each of the fields and converged towards the inlets in the seaward front groyne. By the interaction of seston transporting creeks and seston catching vegetation, a distinct morphological surface relief had now developed, which resulted in an increasingly diversified vegetation cover. At the survey of 1977/78, additionally, to the original pioneer species, Puccinellia maritima and Suaeda maritima had appeared and became dominant. Moreover, some first representatives of later succession stages joined the salt marsh community.
Based on the pattern reached by relief formation in the 1970s, the fields continued their further development until the last survey of 1997 (Figs. 9 and 10). Though unexpectedly, due to activities of a farmer since the 1980s, an additional unit had appeared in the northern fields: the areas developed by technical land reclamation (Figs. 5, 9 and 10).
The ‘Bare tidal flats without vascular plants’ sometimes carried a scattered vegetation of benthic algae, predominantly Vaucheria sp., less frequently Enteromorpha sp. The ‘Low-lying areas’ initially (1977/78) were loosely covered by Salicornia and later, with rising surface altitude, became interspersed by Spartina anglica, Puccinellia maritima, Tripolium pannonicum and Suaeda maritima. Until the last survey of 1997, this unit had been shrinking to negligible size (Figs. 9 and 10).

Morphology and vegetation of the land reclamation fields in summer 1997, resulting from aerial-photograph analysis and ground-truth investigation

Composition of geomorphological and vegetation units during the surveys of 1977/78, 1985 and 1997
The ‘Plane areas of high elevation’ since 1977/78 already represented the largest share of the whole area. Supported by morphological criteria, a division into three subunits of different plant communities appears useful:
-
Subunit dominated by Spartina anglica and Puccinellia maritima (Fig. 11). In 1977/78, both species were growing in a rather regular distribution. Accompanying species in low densities were Salicornia sp., Tripolium pannonicum, Suaeda maritima, Triglochin maritima and Cochlearia anglica. Species spectrum and dominance relations remained similar until 1985 and 1997, except the additional appearance of Atriplex prostrata and Spergularia media. Obviously, this subunit represents elements as well of the pioneer zone as of the low marsh
-
Subunit ‘humpbacked’ Puccinellia lane. This habitat, limited to a narrow strip behind the front groyne (Fig. 12), increased faster in elevation than all other areas (s. transects in Figs. 2 and 4) and had already in 1977/78 approached the mean high-water level or even reached it. By its way of growth on small hilltops the dominating grass Puccinellia maritima had created a peculiar surface structure. Accompanying species were Cochlearia anglica, Suaeda maritima, Spergularia media, Tripolium pannonicum and Plantago maritima. It turned out from the surveys of 1985 and 1997 that the stage of a higher salt marsh had been reached, which was indicated by the presence of species like Limonium vulgare, Glaux maritima and Armeria maritima
-
Subunit transition areas to the foreland. In the northern fields, from the middle of field VII to field XI, the border between the accretion area and the foreland is no longer formed sharply by the cliff. It consists of a gradual transition reaching the stage of a high marsh. With rising elevation, areas dominated by Puccinellia are replaced by areas dominated by Festuca with the corresponding accompanying species. Higher up, a Juncus-gerardii lane is reached, associated with Festuca rubra, Elymus athericus and glycophyte species in the top section of Table 1

View of the widely-spread Puccinellia-Spartina vegetation in summer 1985

Fringe of the so-called ‘humpbacked’ Puccinellia lane behind the front groyne, which is characterised by an accelerated accretion. Summer 1978
The ‘Elevated bank ridges’ are a prominent element of the surface relief. In the 1970s, the bank areas lining the newly originating creeks were still low-lying fringes (Fig. 5), but in the 1980s, they started to rise and finally stood out against the surrounding areas. The vegetation on these ridges in certain seasons differed sharply from the adjoining habitats. In spring, the banks were lit up by a white bloom of Cochlearia anglica (Fig. 13). Towards summer, the herb material of Cochlearia decayed and became covered by rampant growth of Suaeda maritima, followed by Tripolium pannonicum and Atriplex prostrata. By 1997, the bank-ridge was dominated by Puccinellia maritima. Plants from surrounding salt marsh zones occurred in small abundances.

Blooming Cochlearia anglica on the bank ridge of a creek, spring 1985
‘Areas developed by technical land reclamation’: In the 1980s, a farmer had begun to reclaim land in the northern fields, the major part of which became converted to an artificial salt marsh until 1997 (Fig. 9). It is clear that the natural structures of the originating salt marsh were completely destroyed by this. The plane parts of the area were dominated by short-grazed Puccinellia maritima accompanied by slightly taller individuals of other salt marsh herbs. In places of uprising sediment material deposited between the trenches, representatives of later succession stages had established, e.g. Elymus athericus and even a number of glycophytes.
Finally, two small-scale habitats have to be mentioned briefly:
-
The ‘Sparsely covered fringe under the eroding cliff’. Being located in the range of the high-water line, the soil is irregularly flooded and intensely drained; only a few members of the Puccinellia community may occasionally grow here
-
In 1985, a strip of ‘Swampy sinks’ was found lining the abovementioned ‘Sparsely covered fringe’ (Fig. 5). In the stagnant water Triglochin maritima, Suaeda maritima and Spartina anglica were thriving
Considering the whole area, a mean surface elevation of MSL + 1.51 m was reached in 1985 and the diversity had increased to a number of 25 species. Puccinellia maritima had now become omnipresent followed by Tripolium pannonicum, Suaeda maritima and Spartina anglica. In the summer of 1997, the mean elevation of MSL + 1.71 m had exceeded the mean tidal high-water level (1991: MSL + 1.61 m), and the major part of the area lay above that height (Fig. 3). Nevertheless, in large parts of the reclamation fields, the aspect of a finally developed high salt marsh vegetation did not exist. The species composition confirms this impression (Table 1). It had increased to 34 taxa (Table 1), when for the first time the two Salicornia species, S. europaea and S. stricta, were distinguished. Though Puccinellia maritima was present and dominant everywhere, due to sufficient water content and increased salinity of the soil, all representatives of the earlier succession stages maintained their dominant positions, for example, Salicornia sp., Spartina anglica, Suaeda maritima and Cochlearia anglica. Figure 9 shows the state of development as assessed by the last survey in 1997.
Bottom fauna
During the period of investigation, 42 species were identified in the bottom fauna, and 6 additional animal taxa are named on higher levels (Tables 2 and 3). Under ecological aspects, the animals can be divided into two groups: first, a low number of tidal flat species, mainly representatives of polychaetes, crustaceans, gastropodes and bivalves. Second, with proceeding growth of surface elevation, specific animals of a lower salt marsh are establishing, mostly halobiontic and halophilous Hexapoda and occasional members of other classes. Their species number, when considering only the endofauna of the bottom, is also low.
As to be expected, the faunal composition outside and inside the land reclamation fields developed differently. Originally, before the set-up of groynes in 1957, twelve species had lived in the tidal flat bordering the eroding cliff (Table 2 and 3). Outside the fields, a constant group of these species remained dominant until the final surveys of 1985 and 1997 (Table 2). Two polychaete species which had spread as neozoa in the entire Wadden Sea appeared later: Tharyx killariensis since the end of the 1960s (Michaelis 1981, cited as Tharyx marioni) and Marenzelleria cf. wireni in the 1970s (Essink and Kleef 1988).
Inside the land reclamation fields, elements of the original tidal-flat fauna disappeared gradually and animals of salt marsh border habitats established (Table 3). The decisive change occurred in the 1970s. In 1976, the mean elevation rose to heights in the range of MSL + 1.30 to MSL + 1.35 m and the mean inundation time decreased to 1.8 h per tide. As first indicators of a salt marsh formation process, the collembolan Archisotoma pulchella, larvae and pupae of the dipterous family Dolichopodidae and the opistobranchiate gastropod Alderia modesta appeared; the latter species lives as a feeding specialist on the benthic alga Vaucheria sp., which itself is specific for brackish and salt marsh bordering habitats. On the final surveys, 1985 and 1997, the animals of lower salt marsh habitats became predominant in species composition. Meanwhile, large parts of the areas exceeded the mean tidal high water level, partly by natural development, in most of the northern fields, by technical activities. The most frequent and characteristic animals of these habitats proved to be unidentified Enchytraeidae (1997: 100%), the larvae of Dolichopodidae (1985: 52%, 1997: 86%) and the oligochaete Tubifex costatus. Furthermore, larvae and pupae of unidentified Diptera and other Insecta had increased considerably in frequency.
As already noted with regard to vegetation, also in the bottom fauna, a group of originally tidal flat animals was still present, when the reclamation fields had reached or exceeded the mean high water level, ten species in 1985 and eight species in 1997. Nereis diversicolor in 1985 occurred with a frequency of 72% and of still 55% in 1997. Also, Hydrobia ulvae (41%), Carcinus maenas (23%) and Macoma balthica (14%) are belonging to this group.
The summarising parameters of bottom fauna, i.e. species number, abundance and biomass, are given in Table 4. The fluctuations until 1963 were similar outside and inside the fields. In the 1970s, the assemblage inside the fields began to pursue a deviating course, as already indicated by the species spectrum. The total species number increased significantly while the number per station dropped to low values. Abundance declined gradually and biomass drastically. Finally outside and inside the fields, two opposite habitats were facing each other, as already shown up by the characteristics of geomorphology, sedimentology and vegetation.
Discussion
On the coast of Lower Saxony, the scientific attention to the questions of salt-marsh formation, development and erosion arose in the 1930s and 1940s with studies on the biota of the Jadebusen shallows (Linke 1939a), on the importance of suspended matter supply (Linke 1939b) and on the role of the newly introduced cordgrass Spartina (Linke 1973). It is a long-established experience of coastal inhabitants that surface elevation is a main variable controlling the distribution of salt-marsh habitats with their plant and animal life. Local techniques of land reclamation and economic use of the marshlands depended on this knowledge. On the border between sea and land, the frequency and duration of tidal inundations delimit the landward distribution of marin-aquatic organisms and the seaward intrusion of terrestrial plants and animals. Numerous - if not innumerable - studies have been dealing with the ecological zonations arising from these conditions (e.g. Bilio 1963; Heydemann 1967; Ranwell 1972; Dijkema 1983; Beeftink 1987; Bockelmann et al. 2002; Dijkema et al. 2005). More recent examples are given by Suchrow and Jensen (2010), based on an extensive data set from the North Sea coast of Schleswig-Holstein, by Hartmann and Stock (2019) in a long-term view on a selected site in that area, by Esselink et al. (2017) in a status report on salt marshes of the entire Wadden Sea, and by Balke et al. (2016) in a global approach to define the lower border of salt-marsh vegetation. The latter authors conclude that an increase of tidal ranges by sea-level rise will not lead to an expansion of salt marsh belts along the seashores.
In the sedimentation fields of Cappeler Tief, the appearance of vascular plant species during forty years is more or less identically reflecting the spatial distribution in which salt-marsh plants are arranged along the gradient of increasing elevation. Certain deviations from this general pattern point to peculiarities in local or regional factor constellations, as shown by examples from other poly- and euhaline sites (Fig. 14). Close to Hilgenriedersiel on the mainland coast existed a hitherto (1970) untouched salt marsh showing a species distribution under natural conditions (Heykena 1970). Limited to the true halophytes from Salicornia sp. and Spartina anglica up to the first occurrences of Juncus gerardii, Atriplex littoralis and Elymus athericus, the spectrum comprised the same 20 species as present in the fields of Cappeler Tief in a similar sequence. Opposite to Hilgenriedersiel, on the southern edge of the island Norderney, a counterexample to Cappeler Tief was studied from 1957 to 1971 (Bätje and Michaelis 1976). Situated in waters with extremely low suspended matter content (less than 100 mg/l in the tidal and seasonal cycle), these fields were bordered by lateral trenches and not protected by brushwood groynes. They experienced permanent alterations in growth and intermittent relapses of elevations and remained in a dynamic equilibrium on the early stage of a lower salt marsh, low in diversity (11 species). The Leybucht, a polyhaline bay in the Ems estuary, is a site where rapid accretion in recent decades resulted in the formation of extended salt marsh areas (Niemeyer 1991). The increase amounted to more than 20 km2 from 1948 until 1999 (Arens and Heiber 1999). Due to a predominating flood current of the tidal regime, the bay receives an excessive suspended matter supply of more than 1600 mg/l (Scherfose 1989). Assigned to annual inundation frequencies, the succession stages from the pioneer zone up to the ‘Festucetum’ (in terms of phytosociological systematics) are represented (Scherfose l. c.). The species spectrum is comprised of about 20 halophytes. In their sequences and dominances, cattle grazing is one of the relevant factors.

The coast of Lower Saxony with the study area and the sites of earlier salt-marsh investigations cited in the discussion. 1 Study area Cappeler Tief; 2 Leybucht: Scherfose (1989), Niemeyer (1991), Arens and Heiber (1999); 3 Hilgenriedersiel: Heykena (1970); 4 Norderney south: Bätje and Michaelis (1976); 5 Norderney south-east: Hobohm (1993); 6 Jadebusen: Linke (1939a, b), Michaelis (1987)
In the other bay of the Lower Saxony coast, the Jadebusen (southernmost part of the Jade Bight), a conspicuously impoverished salt-marsh vegetation, occurred in the 1970s (Michaelis 1987). Only Salicornia sp., Spartina sp., Aster tripolium (= Tripolium pannonicum), Puccinellia maritima, Suaeda maritima, Triglochin maritima and Limonium vulgare are named as being present. Seston supply in the nearshore areas is unknown, for the main channel mean values about 100 mg/l are recorded. Already earlier, before the invasion of Spartina, Linke (1939a) had pointed to the remarkably low species number of the salt-marsh vegetation in this bay, citing Salicornia europaea, Aster tripolium (= Tripolium pannonicum) and Festuca thalassica (= Puccinellia maritima) as the only species occurring. In contrast to the highly dynamic Leybucht, the Jadebusen is constrained in stable boundaries by a law from 1886 in order to maintain the bay as a reservoir for flushing and preserving the depth of the waterway to Wilhelmshaven (Michaelis l. c.). The comparison of the Leybucht and the Jadebusen demonstrates that morphological variability is an important prerequisite for the plant species diversity of a salt marsh. Consequently, certain dynamics and a number of additional halophytes returned into the bay (Jadebusen), when in recent decades, many pits had been excavated in the vegetated areas to gain bottom material for coastal protection measures (Arens, unpublished observations).
Examples of high species richness are represented by some sandy salt marshes of the East Frisian barrier islands. Apparently, these are the main sites, where natural salt marshes are still increasing (Bunje and Ringot 2003). Esselink et al. (2017) affirm this trend also for the Wadden Sea islands of the Netherlands and Denmark. On Norderney, an extended transition area from dunes to tidal flats is situated at the southeastern part, the ‘tail’, of the island (Hobohm 1993). In these naturally structured, varied and dynamic habitats small patches of lower and larger areas of higher salt marshes are irregularly distributed. The number of halophytic salt-marsh species is higher than elsewhere, including a dozen of red-list species as, for instance, two representatives of the genus Centaurium (Hobohm l. c.). This kind of salt marsh does not owe its existence and diversity to a high supply of suspended matter but to accretion by latitudinal wind and water-driven sediment displacement (Bunje and Ringot 2003).
Compared to the relatively natural vegetation of the mainland sites Cappeler Tief, Hilgenriedersiel and Leybucht, the foreland marshes of the Schleswig–Holstein coast exhibit a fairly different species composition and species sequence along the elevation gradient (Suchrow and Jensen 2010). For example, two characteristic representatives, Cochlearia anglica and Limonium vulgare, remained unconsidered because of their low frequencies. These areas in Schleswig-Holstein consist predominantly of artificially reclaimed fields drained by regular trench systems and over long periods exposed to intensive sheep grazing (Stock et al. 2005, Suchrow & Jensen l. c.).
Since 1986, the Wadden Sea area of Lower Saxony has gained the conservation status of a national park and is integrated in the Dutch-German-Danish ‘Trilateral Monitoring and Assessment Programme’ (TMAP). In this framework, the dynamics of salt marshes play an important role. For reasons of trilateral comparability, a classification was developed that separates the succession stages of salt marshes into a number of ‘types’ (Petersen et al. 2014). Arens and Götting (2008) translated the results of the survey from 1997 (Cappeler Tief) into this system, according to which the main part of the area belongs to the Puccinellia maritima type followed by the Spartina anglica type.
Concerning the occurrence of Spartina, in 1928/29 specimens introduced from England had been planted in the Leybucht and at some other sites of the East Frisian coast (s. Nehring and Hesse 2008). Taxonomically, the plants were representatives of the sterile bastard species Spartina townsendii. The plantations became extinguished down to some dozens of surviving specimens, which, however, in the 1940s started with blooming and excessive propagation. In 1948, Linke (1973) performed a survey of the pioneer vegetation along the East Frisian coast, where Spartina meanwhile occupied an area of nearly 1000 ha and had become a dominant member of the flora. Hence, the sterile bastard Spartina townsendii, only capable to vegetative reproduction, must have been replaced by the fertile Spartina anglica already in those early years. In retrospect, it may have been wrong to renounce the discrimination between the two hybrids in the current project. Evidently, S. townsendii is still surviving beside S. anglica, limited to the Wadden Sea regions of Lower Saxony and the Netherlands (Granse et al. 2022). In 1957, Spartina is cited as being present at Cappeler Tief (Müller 1958), and since the 1970s was a dominant element of the vegetation (Grotjahn et al. 1983). Until the surveys of 1997 and 2007, the species had increasingly consolidated its dominating role (Arens and Götting 2008), which proves to be true for the entire Wadden Sea (Granse et al. 2020). According to reviews of Reise (1994), Loebl et al. (2006) and Nehring and Hesse (2008), Spartina anglica is even going to extend worldwide, because it might benefit from global warming.
Of course, in view of the investigators, the activities of technical land reclamation and grazing after the survey of 1985 were regarded as rather disturbing and unwelcome factors. The effects of these impacts, analysed field by field, are given by Arens et al. (2003).
In the bottom fauna, going along with the habitat change from a bare tidal flat to a densely vegetated salt marsh, the marine aquatic animal species have gradually withdrawn and became replaced by the indicators of a salt marsh habitat. The first representatives appearing are still aquatic animals like Tubifex costatus, Paranais litoralis, Enchytraeidae, Alderia modesta and Assiminea grayana. The collembolan Archisotoma pulchella and the larvae of the Dolichopodidae belong to the semiterrestric halophilous insects which intrude into the intertidal zone. The specialist Dr. R. G. Sommer (Kiel University) identified the two species Hydrophorus oceanus Macquart and Maecharium maritimae Hal. from images that had been raised from the pupae in sandy cocoons collected during several summers. To the next succession stage belong the amphipode Orchestia gammarellus and beetles of the genera Bledius, Heterocerus and Dicheirotrichus. Until the final stage of the surveys, three representatives of the original tidal flat fauna, Nereis diversicolor, Hydrobia ulvae and Macoma balthica, maintained a dominant role in one or two of the parameters frequency, abundance and biomass. It has to be explicitly pointed to the fact that the bottom fauna of the investigated category represents a decreasing fraction with the advance of salt marsh formation. The newly originating ecosystem becomes invaded by real armies of other animal groups which are going to populate the surface layer, the detritus layer and the herbal layer. The survey of 1997 resulted in 24 species of the ‘bottom fauna’ in narrow sense. Its loss of ecological relevance is demonstrated by the biomass of the fauna living in the sediment: In 1997, it had dropped to less than two grammes ADW per square metre. That is only 5 to 6% of general tidal flat values, which during summer range between 30 and 40 g ADW in the Wadden Sea (Michaelis and Wolff 2001). However, a parallel study from 1985, dealing with the abovementioned subhabitats (Stiffel 1990), provided proof of excessive numbers of spider and insect species living in the surface, detritus and herbal layers. Numerously represented were the orders Araneae, Acari, Collembola, Hemiptera, Coleoptera, Hymenoptera, Diptera, Lepidoptera, Aphidina and Cicadina. Studies from the west coast of Schleswig-Holstein focussing on the salt marsh fauna resulted in numbers of more than 1500 invertebrate species, as emphasised in summarising reports of Heydemann (1981, 1983/84) and Dijkema and Heydemann (1984).
As mentioned before, there is a certain contrast between bottom surface elevation and biological succession stage in the sedimentation fields. In spite of having long since exceeded the elevation of mean high water level, the vegetation and the faunal assemblages still contained elements of early succession stages, even in dominant roles. Examples to be mentioned are the plant species Salicornia sp., Spartina anglica, Suaeda maritima, Spergularia media and Cochlearia anglica as well as the animals Nereis diversicolor, Hydrobia ulvae and Macoma balthica. An explanation can be found in the fact that major parts of the newly originated salt marsh did not develop a seaward slope, which according to Verhoeven (1983) is a general characteristic of strongly wave-exposed salt marshes. In the present case, sediment accumulation behind the front groyne is considered an additional factor. Therefore, seawater after spring tides and storm surges could not run off completely and water from precipitations was retained. As a consequence, soil humidity and salinity remained sufficiently high - salinity content was even higher than outside the fields - for providing life conditions to the species group mentioned.
Daring a look into the future, the Wadden Sea countries hope for being saved from a worst-case scenario like that on the south-eastern English coasts. There, the rapid decrease of pioneer zone and forelands caused by reckless embanking and sea-level rise is called ‘coastal squeeze’ (Reed 1995; Hughes 2001). In Lower Saxony, when assessing losses and gains of salt marshes until the beginning of this century, a considerable increase was ascertained for the Weser estuary (Ladage and Stephan 2007) as well as for the entire coast (Bunje and Ringot 2003). In general, however, the growth of area was not due to natural accretion but in major parts to coastal protection activities, in the first line by setting up land reclamation fields. In the other Wadden Sea countries, the situation is more or less similar (Stock et al. 2005; Esselink et al. 2017). In Lower Saxony, the lack of a pioneer zone seems particularly conspicuous (Esselink et al. l. c.). In Schleswig-Holstein, on the contrary, a number of foreland sites show remarkable growth of a natural marsh outside, in front of the sedimentation fields, as indicated by Hartmann and Stock (2019) and also by latest information from ongoing studies (Elschot et al. in prep) and from coastal shop talk. Nevertheless, the risk of a loss can only be encountered by intensive land-reclamation works, as currently practised all along the Wadden Sea. This kind of solution may ensure the existence of lower salt marshes for a number of decades. As transitional habitats, however, they have a limited lifetime within their firmly enclosed territories. After attaining a later succession stage, the reclamation fields do not offer space for gaining foothold to new pioneer vegetation. To a limited degree, the reopening of protected higher salt marshes will allow a return of initial stages. The Wadden Sea national park of Schleswig-Holstein is going to act in this way (Stock et al. 2005), and also in Cappeler Tief, two of the fields became connected with higher marsh areas by opening the summer dike (Arens and Götting 2008). Outside of the technical structures, it remains questionable, whether sea-level rise and growing exposure of the coastline will permit a really substantial settlement of a new pioneer zone (Houwing et al. 1999). Thus, the highly specialised plant and animal communities depending on lower salt marsh habitats might lose extensive areas of their environment.
References
Arens S, Götting E (2008) Untersuchungen zur ökologischen Entwicklung naturnaher Lahnungsfelder und ihrer Stellung im Naturhaushalt. Oldenburg: 1–81
Arens S, Heiber W (1999) Development of salt marsh vegetation in the Leybucht from 1948 to 1996. Senckenbergiana maritima 29(Suppl):7–11
Arens S, Heiber W, Kayser C, Michaelis H (2003) Eignung naturnaher Lahnungsfelder im Hinblick auf ihren Nutzen zur Vorlandsicherung und ihre Auswirkungen auf den Naturhaushalt. Report Forschungsstelle Küste Norderney 6 / 2003, 85 pp
Balke T, Stock M, Jensen K, Bouma TJ, Kleyer M (2016) A global analysis of the seaward salt marsh extent: the importance of tidal range. Water Resources Res 52(5):1–12
Bätje M, Michaelis H (1976) Bodenchemische Untersuchungen in Verlandungsfeldern an der niedersächsischen Küste. Jber Forschungsstelle Norderney 27:135–165
Beeftink WG (1987) Vegetation responses to tidal inundations of salt marshes. In: Van Andel J, Bakker JP, Snaydon RW (eds) Disturbance in Grasslands. Junk, Dordrecht, pp 97–117
Bilio M (1963) Die biozönotische Stellung der Salzwiesen unter den Strandbiotopen. Verh. Dt. Zool. Ges. in München. Geest & Portig Verlag, Leipzig
Bockelmann A-C, Bakker JP, Neuhaus R, Lage J (2002) The relation between vegetation zonation, elevation and inundation frequency in a Wadden Sea salt marsh. Aquat Bot 73:211–221
Bunje J, Ringot JL (2003) Lebensräume im Wandel, Flächenbilanz von Salzwiesen und Dünen im niedersächsischen Wattenmeer zwischen den Jahren 1966 und 1997 - eine Luftbildauswertung. Schriftenreihe Nationalpark Niedersächsisches Wattenmeer 7, 48 pp
Dijkema KS (1983) The salt marsh vegetation of the mainland coast, estuaries and Halligen. In Dijkema K S, Wolff W J (eds) Flora and vegetation of the Wadden Sea islands and coastal areas. Report 9 of the Wadden Sea Working Group. Stichting Veth tot Steun aan Waddenonderzoek, Leiden: 185–220
Dijkema KS, Heydemann B (1984) Western European salt marshes – Wadden Sea and southwest Netherlands. In Council of Europe (ed) Salt Marshes in Europe. Nat Env Ser 30:82–103
Dijkema KS, de Jong DJ, Vreeken-Buijs MJ, van Duin WE (2005) Salt marshes in the water Framework Directive; Development of Potential Reference Conditions and of Good Ecological Statuses. Report 2005-020 of ALTERRA, Texel
Esselink P, van Duin WE, Bunje J, Cremer J, Folmer EO, Frikke J, Glahn M, de Groot AV, Hecker N, Hellwig U, Jensen K, Körber R, Petersen J, Stock M (2017) Salt marshes. In: Wadden Sea Quality Status Report. Eds.: Kloepper S et al., Common Wadden Sea Secretariat, Wilhelmshaven, Germany
Essink K, Kleef HL (1988) Marenzelleria viridis (Verril, 1873) (Polychaeta: Spionidae): a new record from the Ems estuary (The Netherlands / Federal Republic of Germany). Zool Bijdr 38:3–13
Granse D, Suchrow S, Jensen K (2020) Long-term invasion dynamics of Spartina increase vegetation diversity and geomorphological resistance against sea level rise. Biol Invasions. https://doi.org/10.1007/s10530-020-02408-0
Granse D, Motta MR, Suchrow S, von Schwartzenberg K, Schnittger A, Jensen K (2022) The overlooked hybrid: geographic distribution and niche differentiation between Spartina cytotypes (Poaceae) in Wadden Sea salt marshes. Estuaries Coasts 45:1409–1421. https://doi.org/10.1007/s12237-021-00985-4
Grotjahn M, Michaelis H, Obert B, Stephan H-J (1983) Höhenentwicklung, Sediment, Vegetation und Bodenfauna in den Landgewinnungsfeldern beiderseits des Cappeler Tiefs (1957 bis 1978). Jber Forschungsstelle Norderney 34:63–94
Hartmann K, Stock M (2019) Long-term change in habitat and vegetation in an ungrazed, estuarine salt marsh: man-made foreland compared to young marsh development. Estuar Coast Shelf Sci 227:106348
Heydemann B (1967) Die biologische Grenze Land – Meer im Bereich der Salzwiesen. Steiner-Verlag, Wiesbaden
Heydemann B (1981) Biological consequences of diking on salt marshes and mud flats. In Helwig Ovesen C, Tougaard S (eds). Environmental problems in the Waddensea-region. Esbjerg: 31–71
Heydemann B (1983/84) Das Ökosystem “Küstensalzwiese” - ein Überblick. Faun. Ökol Mitt 5:249–279
Heykena A (1970) Die Vegetation des Vorlandes bei Hilgenriedersiel. Jber Forschungsstelle Norderney 20:87–104
Hobohm C (1993) Die Pflanzengesellschaften von Norderney. Arbeiten aus der Forschungsstelle Küste. Norderney 12:1–202
Homeier H (1958) Die morphologische Entwicklung der Cappeler Außendeichsgebiete. Jber Forschungsstelle Norderney 9:112–116
Houwing EJ, Willem E, Smit-van der Waaij Y, Dijkema KS, Terwindt JHJ (1999) Biological and abiotic factors influencing the settlement and survival of Salicornia dolichostachya in the intertidal pioneer zone. Mangroves Salt Marshes 3:197–206
Hughes RG (2001) Biological and physiological processes that affect salt marsh erosion and salt marsh restoration: development of hypotheses. In Reise K (ed) Ecological comparisons of sedimentary shores. Ecol Stud 151:173–192
Illies J (1967) (ed.) Limnofauna Europaea. Gustav Fischer Verlag, Stuttgart
Köritz D (1958) Hydrometrische Verhältnisse auf dem Cappeler Watt und bauliche Folgerungen aus der Gesamtuntersuchung. Jber Forschungsstelle Norderney 9:131–144
Ladage F, Stephan H-J (2007) Untersuchungen zur Vorlandsicherung an der Wurster Küste. Report Forschungsstelle Küste Norderney 02/2007, 41 pp
Linke O (1939a) Die Biota des Jadebusenwattes. Helgol Wiss Meeresunters 1 (3)
Linke O (1939b) Bericht über die im Arbeitsjahr 1937/38 im Watt zwischen Juist und dem Festland durchgeführten Sinkstoffuntersuchungen. Internal Report Forschungsstelle Norderney, 21 pp
Linke O (1973) Published under authorship of “Forschungsstelle Norderney” Zur Ausbreitung und Gesellschaftsbildung von Spartina townsendii an der ostfriesischen Küste. Jber Forschungsstelle Norderney 24:219–239
Loebl M, van Beusekom JEE, Reise K (2006) Is spread of the neophyte Spartina anglica recently enhanced by increasing temperatures? Aquat Ecol 40:315–324
Michaelis H (1981) Recent changes in the communities of the Wadden Sea – natural phenomena or effects of pollution? In: Helwig Ovesen C, Tougaard S (eds) Environmental problems of the Waddensea-region. Esbjerg, pp 87–95
Michaelis H (1987) Bestandsaufnahme des eulitoralen Makrobenthos im Jadebusen in Verbindung mit einer Luftbild-Analyse. Jber Forschungsstelle Norderney 38:13–97
Michaelis H, Wolff WJ (2001) Soft-bottom fauna of a tropical (Banc d’Arguin, Mauritania) and a temporate (Juist area, German North Sea coast) intertidal area. In Reise K (ed) Ecological comparisons of sedimentary shores. Springer Verlag: 255–274
Müller CD (1958) Biologische und sedimentologische Untersuchungen des Cappeler Wattes. Jber Forschungsstelle Norderney 9:117–130
Müller CD (1962) Untersuchung des Verlandungsfortschrittes beiderseits des Cappeler Tiefs. Jber Forschungsstelle Norderney 13:151–179
Müller CD (1963) Fauna und Sediment im Wurster Watt von Solthörn bis Dorumer Tief und ihre Beeinflussung durch die Februarsturmflut 1962. Jber Forschungsstelle Norderney 14:143–166
Nehring S, Hesse K-J (2008) Invasive alien plants in marine protected areas: the Spartina anglica affair in the European Wadden Sea. Biol Invasions 10:937–950
Niemeyer HD (1991) Case study Ley Bay: an alternative to traditional enclosure. Proc. 3rd Conf. Coastal & Port Engineer. in Develop. Countries, Mombasa/Kenya: 1–16
Petersen J, Kers B, Stock M (2014) TMAP-typology of coastal vegetation in the Wadden Sea. Wadden Sea Ecosyst 32:1–90
Ranwell DS (1972) Ecology of salt marshes and sand dunes. Chapman & Hall
Reed DJ (1995) The response of coastal marshes to sea-level rise: survival or submergence? Earth Surf Proc Landforms 20(1):39–48
Reise K (1994) Das Schlickgras Spartina anglica: Die Invasion einer neuen Art. In Lozán J L, Rachor E, et al. Warnsignale aus dem Wattenmeer. Blackwell Wissenschafts-Verlag Berlin: 211–214
Rothmaler W (2021) Exkursionsflora von Deutschland, Gefäßpflanzen: Grundband. 22nd edition, Springer Spektrum
Scherfose V (1989) Salzmarsch-Pflanzengesellschaften der Leybucht – Einfluss der Rinderbeweidung und der Überflutunghäufigkeit. Drosera 89(1/2):105–112
Stiffel S (1990) Besiedlungsmuster der Bodenfauna einer Salzwiese am niedersächsischen Wattenmeer. Diploma study, University of Cologne. 225 pp
Stock M, Gettner S, Hagge H, Heinzel K, Kohlus J, Stumpe H (2005) Salzwiesen an der Westküste von Schleswig-Holstein 1988 – 2001. Schriftenreihe des Nationalparks Schleswig-Holsteinisches Wattenmeer 15, 239 pp
Suchrow G, Jensen K (2010) Plant species responses to an elevation gradient in German North Sea salt marshes. Wetlands 30(4):735–746
Verhoeven B (1983) Geomorphology and soil of saltmarshes. In Dijkema KS, Wolff WJ (eds) Flora and vegetation of the Wadden Sea islands and coastal areas. Report 9 of the Wadden Sea Working Group. Stichting Veth tot Steun aan Waddenonderzoek, Leiden: 26–37
Wolff WJ, Dankers N (1981) Preliminary checklist of the zoobenthos and nekton species of the Wadden Sea. In Dankers N, Kühl H, Wolff W J (eds) Invertebrates of the Wadden Sea. Report 4 of the Wadden Sea Working Group. Stichting Veth tot Steun aan Waddenonderzoek, Leiden: 24–60
Acknowledgements
The costs of this investigation have been borne by the federal state of Lower Saxony, the Ministry of Agriculture and Ministry of Environment. The number of persons to whom we are indebted for support is too large for citing them by names. There are the farmer families, who during the summers of the first decades, hosted our teams and left their cattle sheds for use as field laboratories. Later, when work was based on research vessels, we enjoyed the friendly company of fishermen in the Dorumer Tief harbour. They always offered us a calm berth and supplied us with seafood from their daily catch. We owe remarkable amounts of help to generations of students participating in field and laboratory work. Finally, we gratefully remember the persons of our institute involved in the project, the staff members, scientific colleagues and sailors. Many of whom are meanwhile residing in the eternal researching grounds. Finally, we would like to thank two anonymous reviewers for their careful reading of the manuscript and for their very constructive comments that helped improve this paper.
Funding
There was no special funding of the project, data collection was part of permanent monitoring tasks performed by the Coastal Research Station Norderney.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
The authors declare no competing interests.
Ethical approval
All applicable international, national and/or institutional guidelines for animal testing, animal care and use of animals were followed.
Sampling and field studies
The project was carried out in the framework of official tasks of the Coastal Research Station. All necessary permits for sampling and observational field studies have been obtained from relevant authorities.
Data availability
The data generated and analysed in the course of this study is available in figures, tables and maps of this article and in the interim reports of Arens et al. (2003), Grotjahn et al. (1983), Ladage and Stephan (2007) and Müller (1958, 1962), see References.
Author contribution
HM: project coordination, sedimentology, bottom fauna and aerial photograph analysis. SA: vegetation surveys and aerial photograph analysis. FL: geomorphology and data set evaluation of all surveys. All authors read and approved the final manuscript.
Additional information
Communicated by U. Schückel
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
This article is a contribution to the Topical Collection Biodiversity and Ecology of the Wadden Sea under changing environments.
Hermann Michaelis is formerly affiliated to Coastal Research Station (Forschungsstelle Küste), Norderney, Germany, and is now retired.
Rights and permissions
Springer Nature or its licensor (e.g. a society or other partner) holds exclusive rights to this article under a publishing agreement with the author(s) or other rightsholder(s); author self-archiving of the accepted manuscript version of this article is solely governed by the terms of such publishing agreement and applicable law.
About this article

Cite this article
Michaelis, H., Arens, S. & Ladage, F. Forty years of a salt marsh formation in the Wadden Sea: surface elevation, sediments, vegetation and bottom fauna. Mar. Biodivers. 54, 32 (2024). https://doi.org/10.1007/s12526-024-01421-z
Received:
Revised:
Accepted:
Published:
DOI: https://doi.org/10.1007/s12526-024-01421-z