
Abstract
The present study is a first characterization of benthic communities associated with the intensively-exploited sea urchin Loxechinus albus in Northwest Patagonia. Using video-transects, we sampled 22 L. albus beds, distributed among five sampling zones, defined by oceanographic criteria and geographic proximity. Species composition was characterized and compared among zones using Permutational analysis of variance (PERMANOVA), followed by multiple comparisons and distance-based redundancy analysis. In addition, we used species archetype models (SAMs) to identify and estimate the probability of presence of different groups of species (archetypes) that respond similarly to environmental variables. Most zones were characterized by one or two highly abundant taxa, with Corynactis, Arbacia dufresnii, Antholoba achates, and Pseudechinus magellanicus being the most abundant. Sampling zones were shown to have significant effects upon species composition, reflected in significant differences among most zones. The most informative SAM identified three archetypes and considered substrate effects, linear and quadratic effects of temperature and chlorophyll-a concentration, and linear effects of salinity. Probability of Archetype 1 decreased with latitude, particularly south of the Desertores Sill Constriction. Probability for Archetype 2 seemed related to both latitude and salinity. Archetype 3 presented low probabilities across the whole study area. Considering all results, we propose dividing the study area into three main zones: (1) an estuarine area north from Desertores Sill Constriction, (2) an oceanic area corresponding to the Gulf of Corcovado, the Outer Chonos Archipelago and northern sections of the Inner Chonos Archipelago, and (3) an estuarine area located at the southern section of the Inner Chonos Archipelago.
Similar content being viewed by others
Avoid common mistakes on your manuscript.
Introduction
The Fjord and Channel System of West Patagonia (41° 40′ S–56° 44′ S) is one of the largest, most complex (> 80,000 km of coastline) and least studied estuarine systems of the world (Häussermann and Försterra 2007; Försterra 2009). In terms of biodiversity, the limited information available about its benthic fauna was made evident when important scientific efforts conducted in recent years reported many new species and occurrences (Försterra and Häussermann 2003; Cairns et al. 2005; Häussermann and Försterra 2007). For instance, Häussermann and Försterra (2009) described more than 470 benthic species inhabiting this area, of which > 10% were new to science or lacked proper descriptions prior to their book.
If information about the biodiversity of benthic species in the Fjord and Channel System of West Patagonia is scarce, our knowledge about spatial distribution patterns within this area is even poorer. While available data is mainly limited to site observations (many of them from unverified technical reports), few efforts have been made to integrate such information into regional spatial distribution patterns (Försterra 2009). Addressed as part of the Magellan Province (Spalding et al. 2007), two–three distinct ecoregions have been identified north and south of the Taitao Peninsula (47° S; Viviani 1979). Nevertheless, given the size and environmental complexity of this area and the limited information available, these ecoregions only provide a broad reference with regard to species distribution patterns. A first attempt to define more informative subunits within this system was made by Försterra (2009), who identified 12–14 biogeographical subunits. However, acknowledging this was a preliminary effort, the author called for further verification and analysis.
The ecoregion located north of the Taitao Peninsula, which we will refer to here as Northwest Patagonia (NWP), concentrates most of the severe and increasing anthropogenic impacts affecting the area, such as salmon aquaculture and benthic fisheries, whose ongoing effects on the marine ecosystem are still poorly understood (Molinet et al. 2018; Försterra 2009; Niklitschek et al. 2013). One of these impacts is related to the intensive fishery of the sea urchin Loxechinus albus (Orensanz et al. 2005), which is massively extracted by divers, mainly for exportation purposes (Moreno et al. 2007). Chilean landings averaged 18,077 t between 2010 and 2016, accounting for > 50% of the worldwide landings of sea urchins (FAO 2014) and involving ~4300 divers (SERNAPESCA 2016). Over half of this fishery activity was undertaken in NWP (SERNAPESCA 2016), mainly north of the Guafo Entrance (Moreno et al. 2007; Molinet et al. 2016).
Loxechinus albus forms large beds (11,000–2,500,000 individuals), with densities of up to 10.7 individuals∙m−2 (Molinet et al. 2016), located between the intertidal zone and ~100 m depth (Moreno and Molinet 2013). It is a herbivorous species that feeds mainly on drifted brown algae, although it can also feed on green and red algae (González et al. 2008). Its ecological role in NWP is still unclear. Some authors have indicated that this species might play a significant role controlling the abundance of the kelp Macrocystis pyrifera, (Dayton 1974, 1985; Buschmann et al. 2004), which may affect the structure of the associated benthic communities. Other authors, however, have indicated that this species does not significantly affect the abundance of kelp species, which would imply lower structuring effects on local communities (Castilla and Moreno 1982; Vásquez et al. 1984; Castilla 1985; Vásquez and Buschmann 1997).
Despite the economic importance and potential impacts of L. albus fishery in NWP, little is known about the benthic fauna associated with this species. While several species of sea sponges, cnidarians, mollusks, crustaceans, and echinoderms (Molinet et al. 2014) occur in L. albus beds, no detailed descriptions of its associated fauna have been published. Even less information is available about the spatial distribution patterns exhibited by these communities across NWP, which may be a critical factor for management and/or conservation purposes. In the present study, we aim to reduce this gap in knowledge by providing a characterization and analysis of the composition and spatial distribution patterns of the main species present in macro-epibenthic communities associated with L. albus beds in NWP (from now on referred to as L. albus benthic communities).
In addition to fishery-related and/or ecological processes, L. albus benthic communities from NWP are subjected to highly heterogeneous environmental conditions. The region is characterized by a highly structured and extended coastline formed by two mountain ranges, enclosing a large number of channels, fjords, peninsulas, and small islands (Försterra 2009). Intense levels of rain, in addition to great tidal amplitudes (Bustamante 2009), lead to strong physico-chemical gradients and complex land-ocean interactions (Pantoja et al. 2011). These include north-south gradients of temperature, east-west gradients of salinity (Pickard 1971; Silva et al. 1995), and significant spatial and temporal variability in primary production (Lara et al. 2010). Considering these heterogeneity of environmental conditions, we provide a characterization of the study area, based on a novel statistical analysis (Dunstan et al. 2011; Leaper et al. 2014), which enables to predict the probability of presence of different species groups at a given location based on its environmental characteristics.
Material and methods
Study area
The study was carried out in NWP (Fig. 1), in an area located between 41° 40′ and 45° 44′ S and between 72° 32′ and 74° 52′ W. The NWP region is constituted by an insular (western) section and a continental-Andean (eastern) section, separated by multiple gulfs, sounds, fjords, and channels (Soto 2009). The insular section is constituted by low-height islands, archipelagos, and channels, where the main geographic milestone is the Chiloé Island (41° 47′ S–43° 28′ S), separated from the continent in its northern border by the Chacao Channel (~41° 47′ S). South of the Chiloé Island, separated by the Guafo Entrance (~43° 35′ S), is the Chonos Archipelago (43° 50′ S–45° 45′ S), constituted by a set of low-lying islands. The continental section of the NWP region is constituted by the Andes Mountain Range, characterized by the presence of highly stratified fjords and channels exhibiting low-salinity superficial waters (Pickard 1971; Sievers and Silva 2006). There are three main micro-basins between the insular and the continental sections, the Reloncaví Sound (~41° 40′ S), the Gulf of Ancud (~42° 10′ S), and the Gulf of Corcovado (~43° 30′ S), which constitute the Chiloé Inner Sea. The gulfs of Ancud and Corcovado are separated by the Desertores Sill Constriction (~42° 42′ S), which limits water exchange and circulation between these two micro-basins (Sievers and Silva 2006).

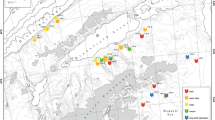
Northwest Patagonia (~41°–47° S) and spatial distribution of the 22 study sites, corresponding to natural Loxechinus albus beds. Colors represent different zones inside Northwest Patagonia. 1: Carelmapu, 2: Caulín, 3: Quenu Island, 4: Tautil Strait, 5: East Guar Island, 6: South Guar Island, 7: Point Pájaros, 8: Chincui Shoal, 9: Nayahue Island, 10: Point Paula, 11: Point Boigue, 12: Westhoff Island, 13: Leucayec Island, 14: Erizo Islet, 15: Midhurst Island, 16: North Skorpios Channel, 17: South Skorpios Channel, 18: Tahuenahuec Island, 19: Point Nicolás, 20: Stokes Island, 21: Rowlett Island, 22: Goñi Channel
A sampling was undertaken with the aim of identifying and studying L. albus beds representatives of the main fishing zones from NWP. The sampling was carried out at 22 L. albus beds in NWP (Fig. 1), all selected considering reports from local divers and historical fishery records and the experience of the team working in the study area (Molinet et al. 2014). Due to adverse meteorological conditions, southernmost sites (Stokes Island, Rowlett Island, and Goñi Channel) were sampled only on protected locations. No sites were selected at the continental section of the NWP region.
Video-transect recording and analysis
The sampling was based on video transects, primarily aimed at estimating L. albus density in NWP beds (Molinet et al. 2016). Three to four video-transects were recorded on most sites, however, due to technical limitations, just two video transects were recorded at Sites 6, 19, and 22 (numeration from Fig. 1), while one additional video was recorded at Sites 2, 7, and 15. A total of 76 videos, recorded between March and July, 2013, at a depth of 2–12 m, using a submarine camera (Seaviewer Sea Drop Model 960) mounted downwards on a sled. The sled was driven by a diver and towed from a vessel (RV Dr. Jürgen Winter) at a speed of ~10 cm s−1. Transect position was triangulated from vessel GPS records (Garmin GPS Map Sounder Model 420 s), while the camera had a visual field of ~28 × 25 cm and nominal transect length was 40 m. Additional information regarding sampling methodology may be found at Molinet et al. (2016).
Video transects were analyzed to identify benthic organisms at the lowest possible taxonomic level. Although this identification level actually ranged from phylum to species, we will make “taxa composition” equivalent to “species composition” hereafter, to facilitate text flow throughout the article. Similarly, we will make “taxa archetypes” equivalent to “species archetypes.” Taxonomic identification was mainly carried out using Häussermann and Försterra’s (2009) illustrated guide, followed by consultation with local experts when required. Individual counts were recorded for each species whenever possible. Otherwise, only presence was registered. Given camera resolution and overall methodological limitations, the study was restricted to epibenthic fauna with a body size ≥ 10 mm. The substrate was classified into six categories: sand, gravel, rock, and their combinations. Since different surface areas were observed for each substrate type, within each transect, all abundances observed by each transect and substrate-type were previously standardized to a nominal surface of 14 m2. To avoid spatial autocorrelation, observations median abundances were computed by site and substrate type, and analyses were performed at this level (i.e., considering these groups of observations as our sampling units). In view of the extensive exploitation of L. albus populations and the spatial and temporal variation that this exploitation may generate, they were not considered in statistical analyses and the study focused on L. albus benthic communities. Notwithstanding, the potential effects of L. albus density on its associated communities may be checked in Contreras et al. (2019).
Characterization of Loxechinusalbus benthic communities from NWP
To describe the spatial patterns of species composition in L. albus benthic communities, we defined a zoning scheme considering local oceanography and geographic proximity among sites. Key criteria to define these sampling zones were the relative positioning (north/south) of the study sites from the Desertores Sill Constriction (Sievers and Silva 2006) and their relative exposition to the open ocean (oceanic versus inner sea locations). The first zone included the two oceanic sites located at the Chacao Channel (Carelmapu and Caulín). The second zone included all inner sites located north or at the Desertores Sill Constriction (Quenu Island, Tautil Strait, East Guar Island, South Guar Island, Point Pájaros, Chincui Shoal, and Nayahue Island). The third zone was constituted by sites located at the Gulf of Corcovado, including two sites from the southern Chiloé Inner Sea (Point Paula and Point Boigue) and two sites from the northern Chonos Archipelago (Westhoff Island and Leucayec Island). The fourth and fifth zones included the sites located at the inner (North and South Skorpios Channel, Tahuenahuec Island, and Point Nicolás) and outer Chonos Archipelago (Erizo Islet, Midhurst Island, Stokes Island, Rowlett Island, and Goñi Channel), respectively (overall information about zones is provided in Table 1). The species composition for each of these zones, and also for the entire study area, was characterized considering the relative abundance of each epibenthic macrofauna taxa identified and quantified.
In order to evaluate the existence of species composition differences among zones, substrate types, and their interactions, a permutational analysis of variance (PERMANOVA) (Anderson and Walsh 2013) was performed (n = 999, Bray-Curtis dissimilarity), followed by a multiple comparison analysis using Benjamini and Yekutieli’s (2001) multiple test correction. PERMANOVA key assumption of homogeneity of within-groups dispersion (Anderson and Walsh 2013) was tested using the betadisper function of the R package vegan (Oksanen et al. 2016), finding no evidence against this assumption (F = 1.07, p > 0.3). In addition, a distance-based redundancy analysis (dbRDA, Legendre and Anderson 1999), also based on Bray-Curtis dissimilarities, was performed to represent sites in a 2-dimensional space, after accounting for zone and substrate-type effects. While observed richness was classified into 36 taxa, analyses only considered the 18 taxa being present in at least 3 of the 22 sites. All analyses were performed using the R package vegan (Oksanen et al. 2016).
Species archetypes models
A relatively novel statistical approach, suitable for analyzing the spatial patterns of groups of species based on their environmental response, is the use of species archetype models (SAMs; Dunstan et al. 2011; Leaper et al. 2014). These models group species (instead of sites) based on their presence/absence responses to environmental variables using finite mixtures of regression models (McLachlan and Peel 2004). According to this approach, species that respond similarly to the environment are grouped into the same species archetype (Leaper et al. 2014). The SAM approach enables the prediction of probability of presence of each species group (archetype) at a given (observed or unobserved) location, based on its environmental characteristics (Dunstan et al. 2011). Therefore, different sites or areas can be formed or compared using archetype probabilities instead of pre-defined zones.
SAMs were fitted to data following Leaper et al. (2014), and using the R package SpeciesMix (Dunstan et al. 2016). The first step to perform the analysis was the selection of the number of archetypes to be formed. To this end, a full model was fitted considering substrate type, geographic (latitude and longitude coordinates), and environmental (temperature, salinity, and chlorophyll-a concentration) covariates. The most informative number of archetypes was then selected by comparing models that considered up to 20 different archetypes using the Bayesian information criterion (BIC). To avoid selecting models with groups containing < 1 species, we followed Leaper et al. (2014), setting the minimum a priori archetype membership probability min(π) = 1/S, S being the number of species (min(π); Dunstan et al. 2011). Thus, the number of archetypes selected corresponded to the model with lower BIC value and with min(π) ≥ 1/S.
Once the most informative number of archetypes was defined, we proceeded to search for the most informative combination of geographic and environmental covariates, and their (first order) interactions, through a backward selection procedure based on BIC. We also compared models with covariates added as linear and/or quadratic terms using BIC. The selected SAM was used to predict the probability of presence of each species archetype across NWP from interpolated geographic and environmental data (see next section). This interpolation followed the environmental data interpolation grid, which had a resolution of 2 × 2 km2 (see below).
Environmental data
Temperature, salinity, and chlorophyll-a concentration data from NWP was collated considering surveys carried out between 2003 and 2013. CTD data on temperature and salinity at 2-m depth were obtained from reports of oceanographic cruises and expeditions carried out in NWP (Table 2; Fig. S1). Additionally, MODIS-Aqua L3 surface chlorophyll-a satellite data were obtained from the NASA website (https://giovanni.gsfc.nasa.gov). Chlorophyll-a data corresponded to monthly average concentrations, with a spatial resolution of 4 km.
Each environmental variable was interpolated by an inverse distance weighting method (Shepard 1968) with a resolution of 2 × 2 km2, using the R package ipdw (Stachelek 2017), and then rasterized using the R package rgdal (Bivand et al. 2014). Annual averages for CTD data were computed within each interpolated cell for the three environmental variables considered in the analysis.
Results
Overall findings
A total of 57,100 epibenthic macrofauna individuals were recorded, identified, and counted in all video transects. These individuals were classified into 46 taxa (Table 3), belonging to 5 phyla (Porifera, Cnidaria, Mollusca, Arthropoda, and Echinodermata), 12 classes, and 23 orders. About 20% of these individuals (11,389) corresponded to L. albus, which was the dominant species in 9 of the 22 study sites.
In the case of phylum Porifera, sea sponges were only identified up to class level. However, considering color, shape, and presence in NWP, some of the Demospongiae probably corresponded to either Cliona chilensis or Clionaopsis platei.
A number of species from phylum Cnidaria were classified into five taxa, which corresponded to classes Anthozoa (4) and Hydrozoa (1). Within Anthozoa, octocorals of the order Alcyonium, as well as the sea anemone Antholoba achates (lobed anemone) and the corallimorpharian (jewel anemones) of the genus Corynactis were identified. A generic Anthozoa group, labeled Actiniaria 1, included several species whose identity was not always certain. This group included Anthothoe chilensis, Acontiaria sp. 1 (yellow stinging anemone), Acontiaria sp. 2 (brown melting anemone), Isoparactis fabiani, Paranthus niveus, Parantheopsis ocellata, and Phellia exlex. No further identification was possible within class Hydrozoa.
Species of phylum Mollusca were classified into 13 taxa belonging to three classes: Polyplacophora (1), Bivalvia (1), and Gastropoda (11). Although Polyplacophora was not subdivided beyond class, some individuals within this group were identified as genus Chiton. All Bivalvia belonged to family Mytilidae. Gastropoda was subdivided into different taxonomic levels, which included the puff-ball snail Buchanania onchidioides, the Chilean abalone Concholepas concholepas, Crepidula, and the sea snails of genera Tegula and Diloma, which could not be differentiated from each other and, consequently, were grouped as Tegula/Diloma. The generic taxon Gastropoda 1 grouped several limpet-like organisms, some of them identified as belonging to the genera Fissurella and Nacella. The generic taxon Gastropoda 2 grouped several snail-like organisms, including those belonging to the genus Argobuccinum. The remaining Gastropoda belonged to the infraclass Opisthobranchia (sea slugs) and were classified as Tritonia, Aeolidia papilosa, Doris fontaini, and Phidiana lottini.
Six taxa were used to classify organisms from phylum Arthropoda, all corresponding to class Crustacea. Only one taxa from infraclass Cirripedia was identified, which corresponded to the giant barnacle Austromegabalanus psittacus. All remaining taxa corresponded to order Decapoda: the southern king crab Lithodes santolla, the squat lobster Munida gregaria, and hermit crabs of the superfamily Paguroidea. Two generic taxa, named Decapoda 1 and Decapoda 2, grouped stone-crab crabs and spider-like crabs, respectively. Decapoda 1 was probably dominated by genera Halicarcinus and/or Eurypodius.
Phylum Echinodermata was classified into 13 taxa, corresponding to classes Asteroidea (8), Ophiuroidea (2), Echinoidea (2), and Holothuroidea (1). Asteroidea (sea stars) included 6 species: Anasterias antarctica, Asterina fimbriata, Cosmasterias lurida, Diplodontias singularis, Meyenaster gelatinosus, and Stichaster striatus, and two taxa only identified up to genus Henricia and order Valvatida. Most Ophiuroidea could not be identified beyond class and were grouped as Ophiuroidea 1, except for the orange brittle star Ophiomyxa vivipara. Besides L. albus, two other Echinoidea (sea urchins) species were identified: Arbacia dufresnii and Pseudechinus magellanicus. Finally, Holothuroidea or sea cucumbers could not be identified beyond class.
Besides marine invertebrates, some species of algae were also observed in the L. albus beds studied. These algae corresponded to Callophyllis variegata, Macrocystis pyrifera, Gracilaria chilensis, Ulva sp., and other unidentified algae. Rhodoliths (nodules composed of coralline algae) were also observed in the study area, forming large aggregations (or rhodolith beds) at Point Pájaros, Chincui Shoal, Nayahue Island, Point Boigue, Point Nicolás, and Midhurst Island. Rhodoliths were present in 16 of the 22 study sites and in all zones. Polychaeta tubes, probably from family Spionidae, were observed forming large aggregations at Punta Pájaros.
Besides L. albus, which represented 20.3% of the total abundance, 50.6% of the quantifiable organisms belonged to phylum Cnidaria, 33.3% to Echinodermata (L. albus not included), 14.0% to Mollusca, and 2.1% to Arthropoda. The most abundant taxa (Fig. 2) were Corynactis, with a relative abundance (f) of 0.310, A. dufresnii (f = 0.203), A. achates (f = 0.125), P. magellanicus (f = 0.110), Tegula/Diloma (f = 0.069), and Actiniaria 1 (f = 0.067). Other relatively common, but less abundant, taxa (f = 0.004–0.029) were Crepidula, Mytilidae, Gastropoda 1, M. gregaria, Polyplacophora, Hydrozoa, C. lurida, Paguroidea, and A. antarctica. Demospongiae were also abundant in several locations of NWP, particularly in Carelmapu at the Chacao Channel, where they were ubiquitous on most rocky surfaces. Other taxa with extremely low frequencies (f ≤ 0.001) were O. vivipara, Henricia, D. fontainii, A. fimbriata, P. lottini, D. singularis, A. psittacus, L. santolla, and Alcyonium.

Total relative abundance of the most abundant epibenthic macrofauna taxa found associated with Loxechinus albus beds in the Northwest Patagonia region. Taxa correspond to Corynactis, Arbacia dufresnii, Antholoba achates, Pseudechinus magellanicus, Tegula/Diloma, Actiniaria 1, Crepidula, Mytilidae, Gastropoda 1, Munida gregaria, Polyplacophora, Hydrozoa, Cosmasterias lurida, Paguroidea, Anasterias antarctica, and others
Spatial patterns of species composition
All proposed zones were characterized by the presence of one or two highly abundant species, usually Corynactis, A. dufresnii, P. magellanicus, or A. achates (Fig. 3). In general, the more oceanic-influenced zones (Chacao Channel, Gulf of Corcovado, and Outer Chonos Archipelago) were characterized by the occurrence of large aggregations of Corynactis, while the more protected zones (Northern Chiloé Inner sea and Inner Chonos Archipelago) were characterized by high relative abundances of the sea urchins A. dufresnii and P. magellanicus. A more detailed description of each zone is provided below, and a complete list of taxa present in each zone is shown in Table S1 (supplementary material).

Relative abundance of the most abundant taxa found in Loxechinus albus benthic communities from the Chacao Channel (a), the northern Chiloé Inner Sea (b), the Gulf of Corcovado (c), the inner (d), and outer (e) Chonos Archipelago. Taxa correspond to Actiniaria 1, Antholoba achates, Corynactis, Hydrozoa, Anasterias antarctica, Arbacia dufresnii, Cosmasterias lurida, Pseudechinus magellanicus, Crepidula, Gastropoda 1, Mytilidae, Polyplacophora, Tegula/Diloma, Munida gregaria, Paguroidea, and others
Chacao Channel
The most abundant taxa here (Fig. 3a) were A. achates (f = 0.435), P. magellanicus (f = 0.264), Corynactis (f = 0.173), and A. dufresnii (f = 0.054). The two sites from the Chacao Channel presented communities dominated by different species: P. magellanicus in Caulín and A. achates and Demospongiae in Carelmapu. In addition, some unusually large aggregations of species were observed at these two sites, such as Corynactis observed on some rocky surfaces of Caulín, and a sea anemone belonging to Actiniaria 1, probably P. exlex, dominating a specific area along with A. achates, in Carelmapu.
Northern Chiloé Inner Sea
The most abundant taxa at the Northern Chiloé Inner Sea (Fig. 3b) were Arbacia dufresnii (f = 0.583), Crepidula (f = 0.121), Tegula/Diloma (f = 0.112), and Actiniaria 1 (f = 0.054). While an area from Quenu Island appeared dominated by a large aggregation of Ophiuroida, probably Ophiactis asperula, the three southernmost sites (Point Pájaros, Chincui Shoal, and Nayahue Island) were dominated by large aggregations of rhodoliths. Abundant patches of polychaeta tubes, probably from the Spionidae family, were observed at Punta Pájaros, but not at the remaining sites of this zone.
Gulf of Corcovado
The most abundant taxa in this zone (Fig. 3c) were Corynactis (f = 0.641), Actiniaria 1 (f = 0.120), A. dufresnii (f = 0.051), and M. gregaria (f = 0.050). High variability in species composition among sites was observed at the Gulf of Corcovado. At Point Paula, Actiniaria 1, M. gregaria, and Anthothoe chilensis were particularly abundant, while Corynactis was extremely abundant at Point Boigue, where it formed dense aggregations on rocky surfaces. The two sites located at Chonos Archipelago (Westhoff and Leucayec Islands) were dominated by A. dufresnii, followed by C. lurida at Westhoff Island and by Gastropoda 1 at Leucayec Island.
Inner Chonos archipelago
The most abundant taxa in this zone (Fig. 3d) were P. magellanicus (f = 0.343), A. dufresnii (f = 0.204), Mytilidae (f = 0.184), and Tegula/Diloma (f = 0.148). In general, the sites in this zone were dominated by P. magellanicus and/or A. dufresnii, except for Tahuenahuec Island, where the most abundant taxon was Tegula/Diloma (although it was followed by A. dufresnii). At Point Nicolás, large aggregations of rhodoliths dominated most of the substrate, along with large and dense aggregations of Mytilidae, with several individuals of P. magellanicus over them.
Outer Chonos archipelago
The most abundant taxa in the Outer Chonos Archipelago (Fig. 3e) were Corynactis (f = 0.559), A. dufresnii (f = 0.132), Tegula/Diloma (f = 0.107), and Actiniaria 1 (f = 0.070). High abundances of Corynactis were observed at all sites except the Goñi Channel, with particularly abundant aggregations at Stokes and Rowlett Islands. This zone also presented large aggregations of rhodoliths, which were present at all sites, reaching their highest abundance at Midhurst Island.
Variability in species composition among zones
Zone, substrate type, and their interaction were found to have significant effects, explaining 48% of the variance observed in the species composition of L. albus benthic communities (PERMANOVA, p < 0.01, Table 4). Posterior pairwise comparisons (Fig. 4) showed significant differences among most zones, except between the Gulf of Corcovado and the Outer Chonos Archipelago (p > 0.1), and between the Inner and Outer Chonos Archipelagos (p > 0.2, Fig. 4).

Heatmap showing results from pairwise comparisons of species compositions found in Loxechinus albus benthic communities from five zones of Northwest Patagonia: Chacao Channel (zone a), northern Chiloé Inner Sea (zone b), Gulf of Corcovado (zone c), inner (zone d), and outer (zone e) Chonos Archipelago. Comparisons were made through PERMANOVA and the probability values were corrected using Benjamini and Yekutielli’s method. Blue quadrants indicate zones with significantly different species composition, while light blue quadrants indicate zones with non-significantly different species composition
Similarly, dbRDA analysis (Fig. 5) showed a rather clear separation between the Chacao Channel and all remaining areas, contrasting with a great degree of overlapping between the Gulf of Corcovado and the Outer Chonos Archipelago, which tended to form a second ordination pole. A third ordination pole corresponded to the Inner Sea of Chiloé, which appeared segregated from the Chacao Channel and the Outer Chonos Archipelago. Samples from the Inner Chonos archipelago, on the other hand spread around the whole diagram, overlapping with most zones, especially with the Northern Chiloé Inner Sea. The taxa with the highest weights (> 0.65 on axis 1 or 2) were A. dufresnii, A. achates, Corynactis spp, P. magellanicus, and Crepidula spp.

Re-scaling of sampling sites representing Loxechinus albus benthic communities after a distance-based redundancy analysis of their species composition in Northwest Patagonia. Only taxa exhibiting weights > 0.4 are shown (Antholoba achates, Corynactis, Pseudechinus magellanicus, Crepidula, and Arbacia dufresnii). Symbols represent different zones within Northwest Patagonia
Species archetype analysis
The most informative SAM explained observed distribution patterns by grouping all identified taxa into three species archetypes (Fig. S2). The most informative model included substrate-type effects, linear effects of temperature, salinity, and chlorophyll-a and quadratic effects of temperature and chlorophyll-a (mean values and standard errors for all these covariates are provided in Table S2), with no interactions. Limited overlap in species composition was predicted among archetypes, as most taxa presented very high probabilities (> 0.99) of belonging to just one of them (Table 3).
The three species archetypes had noticeable differences in their interpolated probabilities of presence across NWP (Fig. 6). Although the first species archetype presented probabilities ≥ 0.25 across most NWP, its probability tended to increase towards the north, becoming particularly high (≥ 0.75) beyond the Desertores Sill Constriction (i.e., at the Northern Inner Sea of Chiloé). Archetype 2, presented, instead, a more complex distribution of probabilities, lacking a clear latitudinal trend, exhibiting very high probabilities (> 0.75) at the Gulf of Corcovado zone, the Outer Chonos Archipelago and the northern section of the Inner Chonos Archipelago, and very low probabilities (< 0.25) at the Northern Inner Sea of Chiloé and at the southernmost section of the Inner Chonos Archipelago. This probability distribution showed certain resemblance to salinity isolines 32 towards the north and 29 towards the south (Fig. S1). Archetype 3 presented very low probabilities (< 0.25) across the whole study area and grouped species of overall low abundance, such as Crepidula, C. concholepas, and A. psittacus.

Probability of presence of the species archetypes 1 (a), 2 (b), and 3 (c) across Northwest Patagonia for epibenthic macrofauna associated with Loxechinus albus beds, at 2–12 m depth and on gravel and rocky substrate
Each of the three species archetypes identified in the present study was formed by a combination of different taxonomic and trophic groups. For instance, archetype 1 was constituted by four phyla, and included deposit feeders (e.g., M. gregaria) and herbivore species (e.g., P. magellanicus). Archetype 2 was constituted by three phyla, including carnivore (e.g., C. lurida) and herbivore species (e.g., Tegula/Diloma). Archetype 3 was constituted by three phyla, including carnivores (e.g., C. concholepas and P. lottini) and filter feeders (e.g., A. psittacus). However, since the biology and trophic ecology of many of the species we identified are still unknown, the trophic diversity of these species archetypes would be expected to be even greater than reported here.
Discussion
The identification of spatial biodiversity patterns is a critical task for the development of conservation and management plans (Whittaker et al. 2005). Thus, information on spatial patterns is particularly important for communities susceptible or exposed to anthropogenic impacts. In the present study, we analyzed the spatial patterns of L. albus benthic communities using a traditional distance-based approach (Gotelli and Ellison 2004), which allowed us to identify zones with different species composition. However, the processes generating this zonation remain unclear. Therefore, we complemented these analyses by modeling the distribution of groups of species considering their environmental response, using Dunstan et al.’s (2011) species archetype modeling approach. This approach has been considered to be particularly useful to model species distributions in regions with strong environmental gradients (Woolley et al. 2013), such as NWP.
The present study is a first attempt to characterize L. albus benthic communities from NWP, finding > 45 taxa of epibenthic macrofauna, which, given methodological restrictions discussed below, can be considered a lower species richness bound. Thus, our results suggest that the NWP L. albus beds host a very diverse epibenthic macrofauna community. Most of the taxa we found to be abundant here have also been reported as “abundant” (Corynactis, Tegula/Diloma, and P. magellanicus) or “common” (the sea urchin A. dufresnii and the sea anemone A. achates) across western Patagonia (Häussermann and Försterra 2009). Thus, further studies are required to determine whether L. albus benthic communities are different from epibenthic macrofauna communities from outside L. albus beds, in the same region.
Zones corresponding to the Gulf of Corcovado and the Outer Chonos Archipelago failed to present significant differences from each other regarding species composition. Besides the physical proximity of these zones, their similarity with regard to species composition might be explained by their high exposition to oceanic influence, which would be consistent with the preliminary zonation scheme proposed for this region by Häusserman and Försterra (2009), who grouped together all zones subjected to strong oceanic influence. Nonetheless, both the Gulf of Corcovado and the Outer Chonos Archipelago presented significant differences with the third oceanic zone considered in our study, the Chacao Channel. Since this channel is located further north and demarks the northern border of the Magellan Province (Spalding et al. 2007), observed differences could reflect true dissimilarities. Nonetheless they could be an artifact related to the fact that the Chacao Channel was represented by only two, very dissimilar, sampling sites.
The species archetypes found in the present study, included species from different taxonomic and trophic groups, resembling results obtained by Leaper et al. (2014), who found that archetypes in southern Australia and northern Tasmania were constituted by different and complementary functional groups rather than by similar species. While our SAMs were fitted considering presence-absence data, following Dunstan et al. (2011) and Leaper et al. (2014), more recent developments allow consideration of high-dimensional counts and biomass data (Dunstan et al. 2013), which might provide further insights into relationships existing between distribution patterns and environmental variability within the study area. It is also important to consider that our species archetypes analysis was restricted to L. albus beds, between 2 and 12 m depth. Therefore, our conclusions should not be extrapolated freely to other benthic habitats within the NWP region. Finally, our SAMs were fit with taxa identified at heterogeneous taxonomic resolutions; however, it is likely that single species have narrower environmental preferences than highly grouped taxa.
Our results showed a reasonable degree of consistency and complementarity between the more classical multivariate approach, based upon a priori zones, and the more novel modeling approach leading to a posteriori species archetypes. This consistency between approaches was particularly clear for the Chiloé Inner Sea, where zones located at the north and south of the Desertores Sill Constriction exhibited important differences in species archetypes probabilities, especially for archetype 2, while multivariate analysis indicated significant species composition differences between the Northern Chiloé Inner Sea and southern zones. Notwithstanding, it is important to emphasize that, in SAMs, each species present at a single site is considered a realization of its corresponding species archetype as a whole (Leaper et al. 2014). Thus, study sites with similar species archetypes probabilities could potentially have very different species composition.
After integrating all evidence provided by multivariate and archetype analyses, it is evident that the tentative zoning scheme we applied needs to be reviewed and improved in order to provide an accurate representation of observed diversity patterns. First of all, our results show the Inner Sea of Chiloe, north from the Desertores Sill Constriction, represent a unique area, of strong estuarine influence, whose species composition departs from all other zones. Second, the Gulf of Corcovado and the Outer Chonos Archipelago tended to present similar species composition, which might justify grouping them together. Third, the biological relevance of species composition differences found between the Inner Chonos Archipelago and both the Outer Chonos Archipelago and the Gulf of Corcovado remains unclear, although it seemed to increase as salinity decreases southward. Finally, the Chacao Channel, represented by two, rather heterogeneous sites, showed a large separation from other zones, following PERMANOVA and dbRDA results, but similar archetype probabilities than the Inner Sea of Chiloé.
Hence, we propose dividing the whole area into three main zones by separating oceanic from estuarine areas: (1) a northern estuarine zone corresponding to the Inner Sea of Chiloé north from the Desertores Sill Constriction and (2) an oceanic zone grouping the Outer Chonos Archipelago, the Gulf of Corcovado, and the northern section of the Inner Chonos Archipelago. A southern estuarine zone is corresponding to the Southern section of the Inner Chonos Archipelago, whose geographic or oceanographic limit remains to be defined. Further research would be required to delineate each of these limits more precisely, as well as to clarify further the relative position of the Chacao Channel within this regional picture.
It is important to consider that our results were based on a heterogeneous combination of taxonomic resolution levels, which ranged from order up to the species level. The main constraint to achieving higher levels of taxonomic resolution was insufficient video resolution and/or the need for physical samples required to achieve higher resolution in the taxonomic identification of some taxa. Nevertheless, although the use of video transects can involve these kinds of limitations, it has the outstanding advantage of covering a large sea bottom surface area in a nondestructive way. Another limitation to achieving higher levels of taxonomic resolution was the lack of taxonomic information available for some taxa from NWP. Conversely, some taxa were relatively easy to identify by video recordings and with the taxonomic information available. For instance, several species of nudibranchs from NWP may be identified by their external morphology and coloration (Schrödl 2009). A consequence of this heterogeneity of taxonomic resolutions is that visually distinct taxa will have more weight in the analysis in comparison to visually similar taxa, which would be grouped together.
The present study contributed to the characterization of species composition and distribution patterns of a poorly known subset of NWP epibenthic macrofauna communities, currently affected by intense L. albus fishery activities (Molinet et al. 2014; Barahona and Molinet 2015; Molinet et al. 2016). To achieve these goals, we found that combining distance-based zone comparisons with archetype modeling was an effective strategy to enhance our understanding of observed spatial distribution patterns. This approach could also be extended to incorporate anthropogenic factors, such as spatially explicit fishing effort data, which, if available, could easily be accommodated into the SAMs. Predictive models and new zoning schemes resulting from these combined approaches could be useful to guide management decisions in data poor scenarios such as this, as well as to produce more informative hypotheses, testable through experimental manipulation and/or improved field observation programs.
References
Anderson MJ, Walsh DCI (2013) PERMANOVA, ANOSIM, and the mantel test in the face of heterogeneous dispersions: what null hypothesis are you testing? Ecol Monogr 83:557–574
Barahona N, Molinet C (2015) Explotación de la pesquería de erizo destino fresco en la X–XI Región. Comité Científico Bentónico
Benjamini Y, Yekutieli D (2001) The control of the false discovery rate in multiple testing under dependency. Ann Stat:1165–1188
Bivand R, Keitt T, Rowlingson B (2014) Rgdal: bindings for the geospatial data abstraction library. R Package
Buschmann AH, García C, Espinoza R, et al (2004) Sea urchin (Loxechinus albus) and kelp (Macrocystis pyrifera) in protected areas in southern Chile. In: Lawrence J, Guzmán O (eds) Sea urchin biology. DEStech Publications, Pennsylvania, USA, pp 120–130
Bustamante MS (2009) The southern Chilean Fjord Region: oceanographic aspects. In: Häussermann V, Försterra G (eds) Marine benthic fauna of Chilean Patagonia. Nature in focus, Santiago, Chile, pp 53–60
Cairns SD, Häussermann V, Försterra G (2005) A review of the Scleractinia (Cnidaria: Anthozoa) of Chile, with the description of two new species. Zootaxa 1018:15–46
Castilla JC (1985) Food webs and functional aspects of the kelp, Macrocystis pyrifera, community in the Beagle Channel, Chile. In: Siegfried W, Condy P, Laws R (eds) Antarctic nutrient cycles and food webs. Springer-Verlag, pp 407–414
Castilla JC, Moreno CA (1982) Sea urchins and Macrocystis pyrifera: experimental test of their ecological relations in southern Chile. In: Lawrence JM (ed) Proceedings of the International Echinoderm Conference. AA Balkema, Rotterdam, The Netherlands, pp 257–263
Contreras C, Niklitschek E, Molinet C, Díaz P, Díaz M (2019) Fishery-induced reductions in density and size truncation of sea urchin Loxechinus albus affects diversity and species composition in benthic communities. Estuarine, Coastal and Shelf Science 219:409–419
Dayton PK (1974) Kelp communities of southern South America. Antarct J US 9:22–23
Dayton PK (1985) The structure and regulation of some south American kelp communities. Ecol Monogr 55:447–468
Dunstan PK, Foster SD, Darnell R (2011) Model based grouping of species across environmental gradients. Ecol Model 222:955–963
Dunstan PK, Foster SD, Darnell R (2016) SpeciesMix: fit mixtures of archetype species. R Package
Dunstan PK, Foster SD, Hui FK, Warton DI (2013) Finite mixture of regression modeling for high-dimensional count and biomass data in ecology. J Agric Biol Environ Stat 18:357–375
FAO (2014) Fisheries and aquaculture software, FishStatJ–software for fishery statistical time series. Food and Agriculture Organization of the United Nations
Försterra G (2009) Ecological and biogeographical aspects of the Chilean Fjord Region. In: Häussermann V, Försterra G (eds) Marine benthic fauna of Chilean Patagonia. Nature In Focus, Santiago, Chile, pp 61–76
Försterra G, Häussermann V (2003) First report on large scleractinian (Cnidaria: Anthozoa) accumulations in cold-temperate shallow water of south Chilean fjords. Zool Verh, pp 117–128
González SJ, Caceres CW, Ojeda FP (2008) Feeding and nutritional ecology of the edible sea urchin Loxechinus albus in the northern Chilean coast. Rev Chil Hist Nat 81:575–584
Gotelli NJ, Ellison GN (2004) A primer of ecological statistics, Sinauer associates. In: Sunderland. USA, Massachusetts
Häussermann V, Försterra G (2009) Marine benthic fauna of Chilean Patagonia. Nature In Focus, Santiago, Chile
Häussermann V, Försterra G (2007) Large assemblages of cold-water corals in Chile: a summary of recent findings and potential impacts. Bull Mar Sci 81:195–207
Lara C, Miranda M, Montecino V, Iriarte JL (2010) Chlorophyll-a MODIS mesoscale variability in the Inner Sea of Chiloé, Patagonia, Chile (41-43 S): patches and gradients. Rev Biol Mar Ocean 45:217–225
Leaper R, Dunstan PK, Foster SD et al (2014) Do communities exist? Complex patterns of overlapping marine species distributions. Ecology 95:2016–2025
Legendre P, Anderson MJ (1999) Distance-based redundancy analysis: testing multispecies responses in multifactorial ecological experiments. Ecol Monogr 69:1–24
McLachlan G, Peel D (2004) Finite mixture models. John Wiley & Sons, New York
Molinet C, Barahona N, Díaz M et al (2016) Using drift video transects and maximum likelihood geostatistics for quantifying and monitoring exploited subpopulations of Loxechinus albus at a mesoscale. Mar Coast Fish 8:70–80
Molinet C, Diaz M, Diaz P, et al (2014) Diseño de una red de estaciones fijas de monitoreo para la pesquería del recurso erizo en la X y XI Regiones. Informe final, Proyecto FIP 2000–18
Molinet C, Solari M, Díaz M, Marticorena F, Díaz PA, Navarro M, Niklitschek E (2018) Fragmentos de la historia ambiental del Sistema de Fiordos y Canales Nor-Patagónicos, Sur de Chile: dos siglos de explotación. Magallania 46: 107–128
Moreno CA, Barahona N, Molinet C, Orensanz JM, Parma A, Zuleta A (2007) From crisis to institutional sustainability in the Chilean sea urchin fishery. In: McClanahan TR, Castilla JC (eds) Fisheries management: progress towards sustainability. John Wiley & Sons, pp 43–67
Moreno CA, Molinet C (2013) El paradigma de la distribución batimétrica de Loxechinus albus (Molina) en Chile. Rev Chil Hist Nat 86:225–227
Niklitschek EJ, Soto D, Lafon A et al (2013) Southward expansion of the Chilean salmon industry in the Patagonian fjords: main environmental challenges. Rev Aquac 5:172–195
Oksanen J, Blanchet G, Friendly M, et al (2016) Vegan: community ecology package. R Package
Orensanz JM, Parma AM, Jerez G et al (2005) What are the key elements for the sustainability of. Bull Mar Sci 76:527–556
Pantoja S, Iriarte JL, Daneri G (2011) Oceanography of the Chilean Patagonia. Cont Shelf Res 31:149–153
Pickard GL (1971) Some physical oceanographic features of inlets of Chile. Fish Res Board Can 28:1077–1106
Schrödl M (2009) Opisthobranchia – Sea slugs. In: Häussermann V, Försterra G (eds) Marine benthic fauna of Chilean Patagonia. Nature in focus, Santiago, Chile, pp 53–60
SERNAPESCA (2016) Anuario Desembarque Artesanal Regional www.sernapesca.cl (accessed: 04 Dec 2017). http://www.sernapesca.cl/index.php?option=com_remository&Itemid=246&func=select&id=892. Accessed 4 Dec 2017
Shepard D (1968) A two-dimensional interpolation function for irregularly-spaced data. In: Blue R, Rosenberg A (eds) Proceedings of the 1968 23rd ACM national conference. ACM Press, New York, USA, pp 517–524
Sievers HA, Silva N (2006) Masas de agua y circulación en los canales y fiordos australes. In: Silva N, Palma S (eds) Avances en el conocimiento oceanográfico de las aguas interiores chilenas, Puerto Montt a cabo de Hornos. Comité Oceanográfico Nacional-Pontificia Universidad Católica de Valparaíso, pp 53–58
Silva N, Sievers HA, Prado R (1995) Características oceanográficas y una proposición de circulación para algunos canales Australes de Chile entre 41o20′S y 46o40′S. Rev Biol Mar Oceanogr 30:207–254
Soto MV (2009) Geography of the Chilean Fjord Region. In: Häussermann V, Försterra G (eds) Marine benthic fauna of Chilean Patagonia. Nature in focus, Santiago, Chile, pp 53–60
Spalding MD, Fox HE, Allen GR et al (2007) Marine ecoregions of the world: a bioregionalization of coastal and shelf areas. BioScience 57:573–583
Stachelek J (2017) ipdw: spatial interpolation via Inverse path distance weighting. West Palm Beach 237–240
Vásquez JA, Buschmann AH (1997) Herbivore-kelp interactions in Chilean subtidal. Rev Chil Hist Nat 70:52
Vásquez JA, Castilla JC, Santelices B (1984) Distributional patterns and diets of four species of sea urchins in giant kelp forest (Macrocystis pyrifera) of Puerto Toro, Navarino Island, Chile. Mar Ecol Prog Ser 19:55–63
Viviani CA (1979) Ecogeografía del litoral chileno. Stud Neotropical Fauna Environ 14:65–123
Whittaker RJ, Araújo MB, Jepson P et al (2005) Conservation biogeography: assessment and prospect. Divers Distrib 11:3–23
Woolley SN, McCallum AW, Wilson R et al (2013) Fathom out: biogeographical subdivision across the Western Australian continental margin–a multispecies modelling approach. Divers Distrib 19:1506–1517
Acknowledgments
Ours thanks to Verena Häussermann for her valuable help with the identification of several taxa of marine benthic fauna. We also thank the Subsecretaría de Pesca y Acuicultura de Chile (SUBPESCA) for issuing scientific fish collection permits.
Funding
This study was funded by the Fondo de Investigación Pesquera FIP Project (2012-14) and the Comisión Nacional de Investigación Científica y Tecnológica CONICYT (Beca de Magíster Nacional No. 22161612).
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
All authors declare that they have not conflicts of interest regarding this publication.
Ethical approval
All applicable international, national, and institutional guidelines for the care and use of animals were followed.
Sampling and field studies
Authors declare that all necessary permits for sampling and observational field studies were obtained from the competent authorities and are mentioned in the acknowledgements.
Data availability
The datasets generated during and/or analysed during the current study are available from the corresponding author on reasonable request
Additional information
Communicated by P. Lana
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Electronic supplementary material
ESM 1
(PDF 523 kb)
Rights and permissions
About this article

Cite this article
Contreras, C., Niklitschek, E., Molinet, C. et al. Benthic fauna associated with Loxechinus albus beds in Northwest Patagonia: spatial patterns and species archetypes. Mar. Biodivers. 49, 2535–2548 (2019). https://doi.org/10.1007/s12526-019-00977-5
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s12526-019-00977-5