
Abstract
Through paleoethnobotanical analysis, we illustrate mutualism between herding and cultivating crops among early Andean mobile pastoralists who lived in the Iroco region of the Central Altiplano of Bolivia during the Formative period (1500 BCE - 500 CE). These pastoralists, known as the Wankarani culture, cultivated quinoa, kañawa, and tubers even though they were mobile pastoralists. Cultivating these crops improved food security for the Wankarani and enhanced the landscape for pastoral production. In turn, a pastoral lifestyle enhanced agricultural productivity. Based on the absence of non-local plant remains, such as maize, or other exotic materials, there is no evidence that the Wankarani were involved in the early interregional llama caravan trade networks that underpinned the development of cities and states in the region. These findings demonstrate that herding and crop cultivation interact in unique ways that lead to farming strategies, crop varieties, and opportunities that would not otherwise exist. Above all, this article brings attention to the role of Andean pastoralists in the development of early agricultural programs.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Pastoralism is a subsistence strategy primarily based on animal husbandry and mobility (Capriles 2014:6), though most pastoralists complement their diet with cultivated crops and animal husbandry is often included as part of farming (Browman 1987; García-Granero et al. 2016; Harris 2007; Khazanov 1994; Spengler et al. 2014). For the most part, archeologists who study early food production focus on either plant or animal populations separately (Lezama-Núñez et al. 2018:2). Along these same lines, scholars have created an archetypal division between herders and farmers and few archeological case studies have looked for evidence of how human plant and animal domestication played out in tandem over time (Arbuckle and Hammer 2019:392; Harris 2007; Spengler et al. 2014). Nevertheless, as an ongoing co-evolutionary process (Harris 1989; Rindos 1984), it is important to consider mutualism between humans, plants, and animals in the development of socio-ecological systems through time (Lezama-Núñez et al. 2018).
Mutualism is an interspecific interaction that reciprocally benefits involved organisms (Boucher 1985). For farmers that practice both animal husbandry and crop cultivation, mutualism is central to their success. Growing crops ensures food security for humans and animals alike, and tending to herds provides farmers with a plethora of products, including dung fertilizer for their fields. In the Andes, archeologists have focused on identifying incipient domestication of llamas, alpacas, guinea pigs, quinoa, and potatoes, and refining the chronology and particular phenotypic and genotypic changes in each of these species (for example see Bruno 2006; Bruno and Whitehead 2003; Lord et al. 2020; Moore 2016; Mengoni-Goñalons and Yacobaccio 2006; Wheeler 1995). While each of these studies has focused on a particular species, this research has implicitly revealed mutualism between humans and plants, or animals and plants through their co-occurring co-evolution. Additionally, research focused on later time periods has demonstrated that these animals and plants played a fundamental role in the development of Andean societies including the rise of the Tiwanaku state (500–1100 CE) and the subsequent Inca Empire (1450–1532 CE). With few exceptions (Bruno and Hastorf 2016; Kuznar 1993; Pearsall 1989), little research in the Andes has directly assessed mutualism between plants, animals, and humans through time.
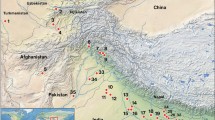
In this paper, we present the results of a paleoethnobotanical study that illustrates mutualism between herding and cultivating crops among early Andean mobile pastoralists. Specifically, we examine three facets of plant, animal, and human mutualism. First, we characterize plant use and agriculture by the Wankarani, an early mobile pastoralist cultural complex who lived in the central Altiplano of Bolivia during the Formative period (1500 BCE – CE 500) (Fig. 1). Second, we show how herding and crop cultivation interact in unique ways that lead to farming strategies, crop varieties, and opportunities that would not otherwise exist. Third, we use paleoethnobotanical analysis to identify the role pastoralists played in broader social developments in the south-central Andean highlands, particularly looking at whether they were involved in early llama caravanning, interregional staple goods exchange, or other long-distance trade networks involving plants.

Map of Oruro indicating the sites included in this study
Herding and agriculture
Llama (Lama glama L.) and alpaca (Vicguna pacos L.) domestication started around 6000 years ago (Mengoni-Goñalons and Yacobaccio 2006:239) while highland cultigens including potatoes (Solanum tuberosum L.), quinoa (Chenopodium quinoa), and kañawa (Chenopodium pallidicaule Aellen) were likely domesticated earlier than 3500 years ago (Bruno 2014a; Rumold and Aldenderfer 2016). Very limited research to date has been done on the Andean domesticates, in contrast to extensive research on domesticates from other parts of the world. With this in mind, the pace of domestication of livestock and crops in the Andes is still being unraveled by zooarchaeologists, paleoethnobotanists, and geneticists. In one of only a few models that factors in mutualism between plants and animals during the development of Andean agricultural systems, Pearsall (1989) and Kuznar (1993) point to the centrality of pastoralists in the process of quinoa domestication. Elaborating on the camp follower hypothesis in which particular small-seeded plants colonized human habitats drawing them into closer mutualistic relationships with humans (Anderson 1952; Sauer 1952), Pearsall (1989) and Kuznar (1993) give llamas a central role in facilitating quinoa propagation near human settlements. In this model, camelids ingest wild chenopods, defecate near human settlements, thereby sowing, fertilizing, and proliferating these undigested seeds in the human realm. Ethnographic work by Hastorf and Wright (1998) bolsters this hypothesis by providing evidence that wild chenopods are preferred camelid forage food and seeds often end up intact in llama dung. Early herding practices would have concentrated wild chenopods in human habitats through selective forage and reseeding. In other words, chenopod seeds ingested by llama herds pass intact through their guts and are deposited in dung heaps. Because llamas excrete in letrine mounds (Moore 2016), wild quinoa would have thrived in dump heaps rich in foraged chenopods and fertilized by the dung itself. If this model is correct, then we would expect pastoralists in the Andes to have incorporated agriculture into their herding lifestyles.
Nomadic pastoralists the world over move to exploit recurrent resources year-after-year (Cribb 1991:373) and they intentionally or unintentionally bring with them plant camp followers, progressively transforming the ecological makeup of their pastoralist niche (Frachetti et al. 2017). Herders who move back to the same location inherit a landscape transformed by previous herding activities. McClure (2015) terms this the ‘pastoral effect’ and elaborates that humans and their herd animals transform ecosystems from the moment they set foot in new zones through trampling, clearing vegetation, propagating weedy plants, and critically enriching the composition of soil by means of their dung and urine (see also Lezama-Núñez et al. 2018). Exemplifying this process, archeologists found that pastoralists left patches of nutrient rich soils in repeatedly utilized areas that encouraged the growth of a mosaic of plant species in African grasslands (Marshall et al. 2018). In this case study, soil enrichment resulting from the excrement of herd animals kept in pens was identified three millennia after occupation; anthropogenic grassy patches continue to be preferred grazing grounds for wild ungulates and grazing herds today. Extending this finding to other global pastoral ecologies, we agree with Marshall and colleagues (2018) that pastoralists elsewhere in the world would be drawn back to these enriched locations improved for herding by the actions of herders in the past.
In comparison to sedentary farming populations, pastoralists also gain access to a wider diversity of plant communities as well as ecosystems where those plants thrive by moving seasonally. For pastoralists engaged in cultivating crops, this would facilitate access to different ecological niches in which those crops and their varieties could grow. The agronomist Norman Borlaug termed this practice ‘shuttle breeding,’ or growing crops in two or more different ecosystems, when he carried out his crop experiments on wheat in Mexico (Borlaug 2007). Aside from being able to grow two crops a year, he found that shuttle breeding resulted in varieties adapted to thrive in various climates, different soil conditions, different photosensitivity requirements (day lengths), and had different tolerances to diseases and pests due to increased exposure and out-crossing with a wide range of wild relatives. The mobile pastoral lifestyle, while motivated by a desire to seek pasturage for herds, likely placed different selective pressures on crops, favoring those with shorter maturation periods, accommodated to the annual movement cycles and that could have needed to grow with little to no tending while pastoralists were away looking after their herds in distant pastures. For pastoralists, breeding crops in difference microenvironments as a result of seasonal transhumance would resulted in particularly hardy varieties that may have required less tending, weeding, and soil preparation than varieties sown by sedentary farmers. The abandonment of certain locations for periods of time also would have allowed enough fallow to replenish nutrients as well as deplete common pests such as nematodes (Bandy 2005). Finally, because of the reduced labor investment, these practices could have been incentivized in environmental settings that would otherwise, be too risky for agriculture.
Andean pastoralism and the Wankarani cultural complex
Situated above 3500 m above sea level, the Altiplano is a large high-elevation steppe covered by grasslands interspersed by wetlands and brushland situated at the heart of the south-central Andes. The northern Altiplano includes the region around Lake Titicaca, which not only receives more precipitation, but also experienced major demographic and sociopolitical transformations fueled by agricultural intensification and sedentism (Hastorf 2008; Stanish 2003). In contrast, the central and southern Altiplano are increasingly arid, and through history were mostly occupied by societies that primarily relied on herds of domesticated camelids.
Low and unpredictable rainfall, varying annual temperatures, and infertile soils that characterize the central Altiplano make it a rather harsh place to make a living for a typical farmer (Fig. 1). Here, between the Early Formative Period (1500–500 BCE) and Late Formative Period (500 BCE – 500 CE), the Wankarani cultural complex emerged and developed in the region. Preceded by Terminal Archaic Period (3000–1500 BCE) mobile hunter-gatherers, the Wankarani have been characterized as mobile pastoralists, in part due to their reverence for camelids, evidenced in stone sculptures of tenoned llama heads, as well similarities to the way people subsist in the region today (McAndrews 2005). Wankarani sites are composed of overlapping residential bases and dispersed logistical camps that facilitated seasonal mobility (Capriles 2014:8). Sites were dispersed in such a way as to provide optimal access to grazing pasture. Zooarchaeological analysis verifies the reliance on domesticated camelids but also the regular exploitation of wild resources including fish and aquatic birds (Capriles 2011, 2014, 2016). Researchers have also recovered lithic technologies such as stone hoes, adzes, and grinding stones as well as features that resemble ethnographic threshing bins suggesting that agricultural activity was also part of the Wankarani subsistence economy (Bermann and Estévez Castillo 1995; Capriles et al. 2011; Fox 2007; McAndrews 2005). Nevertheless, with the exception of one small and focused study (see Langlie et al. 2011), previous research has failed to offer conclusive evidence they were using cultivated plants. Notably, this study is the first systematic paleoethnobotanical publication focused on Wankarani plant use.
Materials and methods
In the summer of 2007, Capriles (2011) directed archeological survey and excavations of three sites in the Altiplano of Iroco, Oruro, with the goal of characterizing the productive system related to early camelid herders and the Wankarani cultural complex in particular (Fig. 1). Briefly, Irucirca (KCH21) is a 1-ha shallow mound initially excavated in 2003 by Albarracin-Jordan (2005). KCH21 includes a large (~ 11 m in diameter) circular corral feature surrounded by residential circular structures made of stone and compacted adobe foundations encircled by and containing hearths and pits of various depths and diameters. The 2007 excavations sampled the corral, a stratified midden north of the corral and residential features to the south and southeast of the mound totaling 39 m2. Korichaca (KCH11) is a taller mound of similar surface also initially excavated in 2003. The excavations in 2007 covered 16 m2 and focused on the center of the mound where a residential occupation was overlaid by three burials. Finally, Cochiraya (KC56) is a large site dispersed over the foothills of a rocky outcrop that included various circular wall foundations. In 2007, 16 m2 was excavated around one of these structures to the southeast of the site (Capriles 2016). A suite of 14 radiocarbon dates verifies all three occupations were occupied synchronously between 200 BCE and 100 CE (Capriles 2017).
Archeological research included a fine-grained recovery protocol that in addition to systematic screening involved the collection of 10-L bulk and scatter soil samples from every stratum and feature excavated (following procedures established by d’Alpoim Guedes and Spengler 2014; Lennstrom and Hastorf 1992). Flotation was carried out on a modified version of Watson’s (1976) SMAP machine. A sample of light fractions from formal features including hearths, floors, pits, middens, patios, and corral floors were selected for detailed macrobotanical analysis. During analysis, all samples were completely sorted using a stereoscopic light microscope with 10–40x magnification. Due to environmental conditions in the central Altiplano and the exposed nature of the sites, it is highly unlikely that uncarbonized plant remains would have preserved, so analysis was restricted to carbonized remains. Macrobotanical remains were sorted into different tissue categories (such as seeds, wood, wood and parenchyma [which is starchy plant storage tissue in the stem, root, leaf, and endosperm of a seed]) and identified to the most specific taxonomic level possible. To assess the relative importance of plants we use ubiquity, expressed as percentage presence of a taxon, proportion, as relative frequency of macrobotanical remains per flot sample, and density, expressed as specimens per liter of soil (Marston 2014; Miller 1988; Pearsall 2015). To illustrate the proportion of plant remains per flot sample, we graphed the results using the Rioja Package (Juggins 2019) in R (R Core Team 2020). Proportion and density sheds light on differences in plant use within Wankarani sites, whereas ubiquity is used to compare variation in plant use between Wankarani sites and Lake Titicaca Basin Formative sites because it reduces the impact of sample size, preservation, and recovery issues on quantification as compared to density (Miller 1988).
Macrobotanical remains were further grouped into categories based on known uses that reflect how the plant remains were likely included in the archeological assemblage. The first group includes plant remains that were crops and consumed by humans including cooking accidents and refuse from domestic consumption. This group includes seeds of domesticated species such as quinoa and kañawa as well as tuber parenchyma. To differentiate among domesticated and wild chenopod seeds, Langlie collected data on seed diameter, testa texture, and relative testa thickness of a sample of these seeds using an ocular micrometer in a light microscope under magnification of 10–40x.
The second group of plant remains includes those that were likely the by-product of llama grazing. Like other places in the world where trees are scarce, Altiplano people use livestock dung to stoke their fires for cooking, warming, and ceramic production (Hastorf and Wright 1998; Miller and Smart 1984). In this study, seeds from crop companion weeds, aquatic plants, small herbaceous plants, and succulents, as well as some of the crop and food plants likely derived from dung burned as fuel (Bruno and Hastorf 2016), though a few weedy species also have economic uses including construction and crafting material. As such, macrobotanical indicators reflect both human plant use, as well as camelid grazing patterns including carbonized dung (Bruno and Hastorf 2016; Langlie and Arkush 2016). We note that it is nearly impossible to identify precisely a particular use of a plant, without exceptional contexts, so we present all alternatives for which burned seeds may have been deposited in the archeological record. Although some of the plants identified in this study are often crop companions, sometimes referred to as weeds, they also grow everywhere in the region but are found in higher densities in disturbed environments like agricultural fields, gardens, and abandoned herd enclosures (Cuenca Sempertegui et al. 2005; Magne and Columba 2011). Grasses also grow just about everywhere in the Altiplano. Aquatic plants grow in the lacustrine and riverine ecosystems to the west of the study sites. Small herbaceous plants, as well as succulents, abound naturally on the undisturbed landscape as well as the hills to the east of the study sites.
Results
In total, 28 samples containing 4798 carbonized macrobotanical remains were identified in this study (Table 1, Figs. 2 and 3, and Supplementary Table 1).

Image of archeological seeds. a Chenopodium sp., b Malvaceae, c Fabaceae, d Cyperaceae, e Ruppia sp., f Poaceae, and g and h Cactatceae

Proportion of macrobotanical remains by site, context, and flot sample (rendered by Alejandra Domic)
Crops and food plants
Quinoa and kañawa. Chenopodium spp. seeds are the most abundant and ubiquitous macrobotanical remains in this study. Two chenopod species were domesticated and cultivated in the Andes, quinoa (Chenopodium quinoa) and kañawa (Chenopodium pallidicaule; sometimes spelled cañihua). The domestication processes of Andean Chenopodium spp. grains have been under increasing scrutiny over the past decade (Bruno 2006; Bruno and Whitehead 2003; Fritz et al. 2017; Langlie et al. 2011; Langlie 2018; Planella et al. 2015). Through increased management of wild stands, cultivation, and intensification of chenopod production, species exhibited marked changes as a product of domestication during the southern Andean Formative period (1500 BCE-CE 500) (Bruno 2006; Bruno and Whitehead 2003), though chenopods were probably first domesticated prior to this during the Terminal Archaic period (3000–1500 BCE) (Bruno 2014a). These studies have used a suite of up to six traits to identify human selection pressures on crops (for a description of traits and protocols used to measure them see Bruno 2006; Bruno et al. 2018; Fritz et al. 2017; Planella et al. 2015).
To further characterize the intra-specific variability of chenopods, Langlie measured the diameter, and testa texture, and determined the relative testa thickness of 50 seeds randomly selected from six samples in this study (Fig. 4 and Supplementary Table 2). To illustrate the results of testa thickness compared to seed diameter, and compare these results to modern crops, weedy, and wild chenopod types, we graphed the results in R (R Core Team 2020) using ggplot2 (Wickham 2016). The mean diameter is 1.03 mm and the range of the diameter is 0.6 to 1.8 mm. Diameter alone is not an indicator of domestication status of Andean chenopods (Bruno 2008:207), so testa thickness was categorized into four distinct relative categories: n = 10 thick, n = 5 medium, n = 5 thin, and n = 30 no testa. The thick testa chenopods are consistent with Quinoa negra the wild/weedy species that is thought to have coevolved alongside quinoa. The testa thickness of modern negra ranges from 22 to 51 μm (Bruno 2006:38). The medium thick testa chenopods identified are likely semi-domesticated or wild varieties. The thin testa chenopods appear to be domesticated quinoa (Bruno 2006). Additionally, 30 chenopods did not have a testa. It is possible that a portion of these chenopods represent a domesticated variety where testa was so thin that it charred and flaked off completely during firing, whereas the rest lost their testa to taphonomic processes. Four chenopods with medium to thick testas have canaliculate testa texture, particularly around the hilum. This testa texture is similar to kañawa and the weed illama (Bruno et al. 2018). Most of the thin testa seeds appeared smooth, which is similar to quinoa (Bruno 2008:41). These data indicate that the Iroco chenopod assemblage is composed of quinoa and kañawa. The presence of medium to thick testa chenopods might be attributed to variation among an early domesticated population of chenopods, or humans and animals seeking out wild chenopods for consumption. The identification of thick testa chenopods and possibly illama points to a crop/weed complex cultivated at the sites similar to that found at a Formative period site near Lake Titicaca (Bruno and Whitehead 2003).

Graph depicts attributes of Chenopodium sp. The x-axis depicts testa thickness in ascending order from thinnest to thickest of both archeological and modern chenopods. (Rendered by David Mixter)
Potatoes and other tubers
Parenchyma is found in dense concentrations and is ubiquitous in this assemblage. Andean paleoethnobotanists assume that parenchyma (or plant storage tissue) found consistently in archeological samples from the highland Andes derives from potatoes and tubers (for example see Bruno 2014:136 and Wright et al. 2003:388), though it is difficult to assign specific taxa to these remains at this time. No fewer than 17 species of roots and tubers belonging to at least nine plant families were domesticated in the Andes (Flores et al. 2003). These include the potato (Solanum tuberosum), oca (Oxalis tuberosum Molina), mashwa (Tropaeolum tuberosum Ruiz I Pav.), ulluco (Ullucus tuberosus Caldas), and maca (Lepidium meyenii Walp.) (Flores et al. 2003; Pearsall 2008). All of these tubers and roots are boiled or mashed in preparation for consumption. Given this preparation method, tubers rarely burned and preserved archeologically. Furthermore, the dense water content of tubers means that they also rarely carbonize wholly like grains (Pearsall 2000:157). However, in the high Andes, people freeze-dry potatoes for storage (referred to as chuño)(D’Altroy 2002). Without the water content, chuño burns and is preserved the same way as seeds. While it is not possible at this time to determine a taxonomic designation for the parenchyma, the large quantities and high ubiquity of parenchyma indicates tuber cultivation and inclusion in Wankarani diets.
Camelid grazing plant remains
Companion herbaceous seeds
In addition to grasses, two plant taxa that were identified in this study thrive in disturbed and rich soils: Malvaceae and Fabaceae. While these plants are found across the landscape, ethnobotanical research indicates that as crop companions, they would occur in high densities in agricultural fields, in soils increasingly impacted by herding intensification, and near pastoralist corrals and homesteads (Browman 1989:152; Bruno 2008:236–241; Domic et al. 2018; Sempertegui et al. 2005). Malvaceae has a high standardized density and ubiquity, similar to chenopod grains, indicating it was quite important in the assemblage. The Fabaceae seeds identified are Trifolium amabile Kunth, a wild legume in the Fabaceae family and found in lower quantities than other plant remains, but still have a high ubiquity of 63% indicating their importance. There are two potential scenarios to explain these seeds. The high ubiquity of Fabaceae and robust ubiquity and quantity of Malvaceae seeds potentially indicates the Wankarani grazed their camelids in fields, likely in fallow. Alternatively, grazing camelids may have preferentially foraged for these plants. At the moment, it is difficult to confirm which scenario is likely, but based on the identification of crop plants, the first hypothesis seems plausible.
Small herbaceous seeds
Lepidium sp., Plantago sp., Poaceae, Verbena sp., and nine small unknown seeds are small herbaceous wild types that indicate grazing in natural pastures. Lepidium sp. and Plantago sp. have comparatively low density and low ubiquity values. Bruno (2008:244) points out that the preferred habitat for Plantago sp. plants in particular is in undisturbed soils. Poaceae, or grass seeds were found in comparatively lower densities than other taxa, but with high ubiquities indicating these plants were regularly consumed by camelids, but in lower quantities than other plant types. The plethora of grass species in the region are currently used primarily for animal forage, as well as fodder, basketry, and construction material (Bruno 2008:246–247; Capriles 2011; Cuenca Sempertegui et al. 2005; Magne and Columba 2011; Whitehead 2007:224). As such, it is possible that grass was used as roof thatch or in other ways by humans, but these seeds also likely reflects dung burning. The presence of small herbaceous seeds, particularly the high ubiquity of grasses, indicates Wankarani herds were regularly grazed in natural pastures.
Succulents
Cactus seeds were found in over half the analyzed samples. Opuntoideae and Cactoideae seeds were both identified, though they were lumped into the Cactaceae family for the purposes of this study, as the two plants were likely used the same way. Cactus fruits are consumed by humans and animals in the Altiplano (Browman 1989:153; Hastorf and Wright 1999:218; Whitehead 2007:176). As a perennial succulent, the above ground growth of these plants does not die back every year and they take a comparatively long time to regenerate when they are disturbed.
Aquatic plant seeds
Cyperaceae and Ruppia sp. plants grow in lacustrine or riverine indicating camelids grazing in these habitats. Cyperaceae seeds have a high standardized density and ubiquity. While at this time, archaeobotanists working in the Altiplano have not been able to identify this seed type to species, Bruno (2008:233) believes that the Cyperaceae from archeological contexts on the Taraco peninsula is Schoenoplectus sp., Carex sp., or Scirpus sp., and these species all grow in moist soil conditions. Totora (Schoenoplectus californicus (C.A. Mey.) Soják) is a perennial aquatic sedge that is both collected from wild stands and intensively cultivated, is commonly consumed as food or fodder, and can also be used as thatching for roofing material, boats, mats, tools, cordage, and animal fodder (Banack et al. 2004; Browman 1989:150–151; Bruno 2008:234; Orlove 1991:6; Whitehead 2007:207). Totora may be been used by humans for construction or craft material, or it may reflect dung burning, though in very low quantities in either case. Ruppia sp., which commonly known as a ditch grass, has a comparatively lower standardized density and ubiquity, indicating it was less important than other taxa in the macrobotanical assemblage.
Dung remains and burned wood
Small charred pellets and pellet fragments of dung were found in 21% of the samples analyzed. Notably, no burned dung was found in samples analyzed from the corral context. We should not expect to find any dung in corrals since burning activity was not carried out where animals were kept. Indeed, the absence of burned materials provides proximal support these spaces were used as corrals. Future analysis targeted at identifying struvites and other geochemical signatures would surely identify strata rich in urine and dung, and compacted under the nails of camelids. Burned wood is also ubiquitous in the assemblage and because most charcoal fragments were relatively small, it is likely these were derived from small brushes. These findings confirm that the Wankarani were indeed using a combination of dung and brush to stoke their cooking, warming, and craft production fires.
Comparing Wankarani plant use to sedentary farming populations
We compare this macrobotanical assemblage to those from other highland Formative period sites including Chiripa and Kala Uyuni located on the shores of Lake Titicaca to elucidate similarities and differences in culinary practice as well as herding behavior among communities that were more typically engaged in sedentary farming. In contrast to the Wankarani’s transhumant lifestyle, during the Formative period, residents living in the northern Altiplano around Lake Titicaca built sedentary villages and relied on potato and quinoa farming as well as camelid pastoralism, fishing, and hunting (Browman 1989; Bruno 2008, 2014b; Bruno and Whitehead 2003; Capriles et al. 2014; Nordstrom 1990; Whitehead 2007). Notably, much paleoethnobotanical work has been carried out on Formative period sites near Lake Titicaca. Because populations near Lake Titicaca are believed to have been more sedentary, they lived in larger settlements, and in denser aggregations of populations, similarities in paleoethnobotanical assemblages might be revealing the habits of their herding animals than about differences in the agricultural strategies of contemporaneous sedentary and mobile populations.
Figure 5 illustrates crop use by the Wankarani, was generally similar to Formative period sites near Lake Titicaca. Both plant economies can be characterized generally as based on locally produced grains and tubers. Even though the Wankarani moved seasonally, crops were just as much a part of their economy as contemporaneous people who occupied more sedentary village sites including Chiripa and Kala Uyuni. Since the Wankarani spent a great deal of time seasonally relocating herds to greener pastures, they may have also spent less time disturbing and preparing the landscape, including removing succulent perennial competitors such as cactus. As a result, there may have been more cactus on the landscape in Oruro than along the the shores of Lake Titicaca.

Bar graph indicating ubiquity values depicting use of the most common plant remains in archeological sites in Formative period Altiplano archeological sites
Based on high ubiquity values, Wankarani people were grazing in wetlands and using Cyperaceae similarly to sedentary farmers (Table 2). In the otherwise harsh ecosystem of the Altiplano, lacustrine and riverine habitats provided humans and their herds important forage, water, and other resources. Figure 5 illustrates only minor differences in ubiquity values of crop companion seeds, and small herbaceous taxa. These findings reveal that, even though the Wankarani were more mobile than farmers living near Lake Titicaca during the Formative period, their camelids foraged on similar plants from similar habitats. Overall, the plants consumed by mobile Wankarani humans and their herds were quite similar to that of their more sedentary neighbors living in the northern Altiplano.
Discussion
Through an analysis of paleoethnobotanical remains, we elucidate the role of the Wankarani in the development of early highland Andean farming strategies. The specific types and varieties of crops grown by the Wankarani shed light on pastoralist’s breeding regimes. By looking at macrobotanical remains that reflect dung burning (substantiated by the identification of dung pellets) and thus grazing, these findings evince grazing strategies and hint at whether the camp follower hypothesis of quinoa domestication holds water. Furthermore, these findings bolster the hypothesis the Wankarani were not involved in early interregional exchange of staple foods.
Human subsistence
The high ubiquity of chenopods and parenchyma across all three sites in this study indicates cultigens were an important part of the Wankarani economy. The recovery of agricultural tools to turn the soil, such as stone hoes, indicates Wankarani peoples were the ones growing these crops; they were not imported from elsewhere (Bermann and Estévez Castillo 1995; Capriles et al. 2011; McAndrews 2005). The analysis of Chenopodium sp. seeds suggests that both kañawa and quinoa were cultivated. Although these seeds are smaller than modern varieties, pre-Columbian domesticated chenopods are also generally smaller than modern varieties (see Langlie 2019). Tubers and chenopod seeds complemented a protein-rich camelid diet.
Kañawa, as compared to quinoa, grows well at higher altitudes and requires less water to grow (Browman 1989:143). Based on this, it is unsurprising the Wankarani, living in a drier part of the Altiplano, were growing both quinoa and kañawa. At this time, there is not enough evidence to conclude that the Wankarani domesticated any particular crop, though these findings point to the Wankarani being involved in these co-evolutionary processes suggested in the camp follower hypothesis. Future research with a more robust temporal dimension may shed more light on this issue.
The only previous study on Wankarani plant use, also carried out by Langlie, focused on identifying a previously unknown domesticated chenopod variety at the Early Formative site of La Barca (Langlie et al. 2011). Morphologically dissimilar to quinoa or kañawa varieties grown today or in archeological contexts from Lake Titicaca, it was hypothesized that this unique chenopod may have been particular to Oruro and bred by the Wankarani. Notably, this variety with a distinct prominent beak was not identified in any of the archaeobotanical samples analyzed in this study even though La Barca is only 40 km to the west. Thus, we suggest different regions and individual mobile groups were experimenting with cultivating distinct cultigens that were probably adapted to particular habitats.
Today, farmers in the Altiplano cultivate hundreds of varieties of quinoa (Andrews 2017), each adapted to different environmental and climatological conditions (Winkel et al. 2015). Experimental research suggests local quinoa varieties from the southern Altiplano fail when planted in the moister soils and air around Lake Titicaca (Danielsen et al. 2003) and varieties hailing from Lake Titicaca struggle to grow in the drier and colder conditions in the southern Altiplano (Winkel et al. 2015). Therefore, it seems plausible that what we are detecting in identifying distinct varieties grown by different Wankarani groups is evidence of early cultivation of unique chenopod ecotypes, or in other words, crop selection based on local environmental factors. Transhumance meant that seed stock would have been exposed to a wide range of growing conditions, as well as the pollen of wild relatives naturally adapted to different habitats. As such, the mobility patterns of the Wankarani would have positioned them to access diverse gene stock and engage in plant breeding, which produced crop varieties adapted to different local habitats and regions.
Tuber planting requires significantly less land clearance than seed crops, they take very little work to tend; the part of the plant above ground does not need to be defended from other predators the way a seed crop does, and are storable for a period of time if left in the ground (Hildebrand 2007). Wankarani people could have even gathered some tubers during herding trips or even planted some, moved herds to seasonal pastureland, and came back to harvest their tubers months later. In this way, tubers uniquely fit a transhumant lifestyle, which may explain the ubiquity and abundance of parenchyma in samples.
This suite of crops is similar in composition to that identified among more sedentary farming settlements located near Lake Titicaca and occupied during the same time period. These findings highlight how a high degree of pastoral mobility is not necessarily correlated with less crop cultivation. These findings also underscore how camelid herds could also bias the paleoethnobotanical record towards their consumption patterns due to the practice of dung burning.
Grazing
The identification of seeds from crop companion taxa, aquatic plants, small herbaceous plants, and succulents suggests a range of ecosystems were exploited by grazing camelids. Llamas likely also grazed on chenopod plants in fields or in pastures evidenced in the high density of chenopods identified in this study. While Malvaceae and Fabacaeae are found throughout the region, as crop companions, they would be found in higher quantities in agricultural fields but also in intensively herded grazing areas. As such, camelid grazing provides an explanation for the high ubiquity of Fabacaeae seeds and high ubiquity and density of Malvaceae seeds. Indeed, ethnographic work among Aymara farmers living near Lake Titicaca indicates herders regularly graze camelids in fallowed fields today, thereby improving the legacy of soils through dung amendment (Bruno 2014b:133). Similarly, pollen data from the Altiplano in northern Chile verifies that Trifolium sp. became increasingly common as a result of overgrazing (Domic et al. 2018). Along these same lines, if livestock were consuming wild legumes in pastures or in fallowed fields, they would have contributed to propagating them in their dung, further improving soils for agricultural production and enhanced herding. Almost all Fabaceae plants play host to rhizobia in their root nodules which fix nitrogen in soils. Camelids grazing in fields would have aerated compacted soils with their small, but sharp nails. These actions would have, in turn, improved crop yields and the landscape for crop cultivation year-in and year-out, pulling camelid herders back to particular locations on the landscape.
All of these actions point to mutualism between herding and crop cultivation. When Wankarani herders seasonally returned to their residential base camps, they were not just returning to domestic architecture, corrals, and pastures; they were coming back to improved field systems that were cleared of perennials competitors like cacti and bushes, which created positive incentives for increasing agricultural productivity year-after-year. Similarly, they were returning to improved pasture lands. Further research needs to be done to bolster the finding that camelids were grazed in fields, perhaps in the form of geoarchaeological analysis of soils.
The high ubiquity and quantity of grass seeds, presence of other small herbaceous seeds, and high ubiquity of cactus seeds indicate the Wankarani were also regularly grazed in natural pastures. Notably, cactus take a long time to regenerate, unlike other small herbaceous annuals and perennials. As compared to the more sedentary farming populations living near Lake Titicaca, the higher ubiquity of cactus seeds points to less disturbance in the Oruro region. This might be due to lower population density living in Oruro as compared to Lake Titicaca during the Formative period, or perhaps, due to high degrees of mobility, the Wankarani disturbed the landscape less than sedentary farmers.
Two taxa were present that suggest camelids were regularly grazing in aquatic ecosystems (Ruppia sp. and Cyperaceae). The sites in this study are located in proximity to Lake Uru-Uru. Grazing in these ecotopes provided hydration for herds and humans alike, as well as important fodder. While grazing in these locations, it seems they also opportunistically hunted, fished, and collected wild fauna (see Capriles 2017).
Wankarani and caravans
In the Andes, herders have historically managed llama caravans that moved staple goods such as salt, maize, coca, and chili peppers between highland grasslands and more agriculturally productive inter-Andean valleys. The Oruro region is located between the Cochabamba eastern Andean valleys and the northern Altiplano, and thus, has long been part of interregional exchange networks such as those documented historically and ethnographically (Molina Rivero 2006). For example, Bonzani (2021) recently reported the macrobotanical analysis from a large sample of contexts from the Inca site Ce51 located near the Late Horizon period (1450–1550 CE) administrative center of Paria and approximately 23 km east of the Iroco. While her results also emphasize the importance of highland taxa including chenopods, she also reports a wide array of exotic taxa including maize, squash, and fruits that residents procured from the eastern lowlands, likely facilitated by increased interregional movement of llama caravans during the Late Horizon.
Although exotic materials including shells from the coast and other foreign objects in Wankarani sites suggest that some goods circulated across the region (Capriles 2017), the absence of exotic plant commodities in the archaeobotanical record such as maize, coca, and chile peppers indicates that staple good exchange might postdate the Late Formative Period. Around the same time the Oruro Wankarani sites were inhabited by mobile pastoralists, maize, likely consumed as chicha or maize beer, was starting to be imported into the Lake Titicaca Basin and increasingly so during the subsequent Middle Horizon period (500–1100 CE) (Berryman 2010; Hastorf et al. 2006; Logan et al. 2012; Wright et al. 2003). While llama caravans carrying goods from Cochabamba to the central Altiplano may have begun during the Formative period (Beaule 2002; Browman 1997), no evidence of exotic plant remains was found in this study and there is also an absence of exotic species in the faunal assemblage (Capriles 2014:51). Perhaps caravanners did not stop at Wankarani settlements. Perhaps Wankarani people did not consume exotic foodstuffs because they were too valuable or unfamiliar, or they consumed such low quantities that these plants were invisible in the archeological record until they started coming in higher quantities later in time. A few seeds in this study, categorized as unknowns, were not identifiable based on gross morphology. It is possible some of these seeds were exotics; however, they are not any well-known Andean or Amazonian staples. Future macrobotanical and microbotanical research in Oruro may shed more light on this subject.
Notably, the sample size in this study is small, possibly affecting the identification of rare or exotic species such as medicinal or hallucinogenic plants, which may have been overlooked. Nonetheless, we feel these findings provide important foundational data that inform interpretations on the role of the Wankarani in the development of agricultural economies in the Bolivian central Altiplano. We hope to build on this work going forward.
Conclusions
Global research on the role of mobile pastoralists in the development of agricultural strategies and social complexity has flourished over the past decade. Nonetheless, little attention has been paid to these topics in the Andes. The high mobility of Wankarani groups coupled with their reverence for camelids epitomized in their stone carvings lends to their designation as primarily pastoralists, but as the results of this study suggest, cultivating crops was an important part of their lifeways. Furthermore, these findings provide evidence of mutualism between herding and agriculture. The Wankarani were adept tuber, quinoa, and kañawa farmers. The identification of different chenopod varieties at geographically close Wankarani sites indicates pastoralists were cultivating unique varieties that were locally adapted or out-crossing with local wild populations. A mobile lifeway would have enhanced opportunities for a preponderance of chenopod varieties to flourish. Mutualism between camelid herding and agriculture is further demonstrated in these data by the finding that camelids may have been grazed in fields, based on the ubiquity of herbaceous companion seed types burnt in dung. Grazing in fields would have increased soil productivity through dung amendments and aeration by herd trampling. High-intensity archeological survey and excavations have revealed that the Wankarani recurrently returned to the same logistical and base camps (Capriles 2014). Although mobility seems to have shaped the settlement patterns associated with the Wankarani as a pastoralist society, it could have also generated incentives for increased engagement with agriculture. In other words, Wankarani herders returned not only to domestic infrastructure in the form of housing and corrals, but also improved grazing pasture and agricultural soils, creating additional incentives to return to the previously occupied residential locations.
While the primary goal of seasonal transhumance would have been to gain access to pasturage, mobility would have enhanced agricultural opportunities. Transhumance would have increased peoples’ access to diverse ecotopes for grazing, access to social networks, and would have facilitated seed exchange and transfer of farming know-how, and outcrossing opportunities with wild crop relatives. Though there is no evidence Wankarani mobility was associated with the development of early interregional exchange. As Borlaug (2007) found during his Green Revolution experiments on wheat, shuttle breeding facilitates the development of hearty crops adapted to a wide range of conditions, as compared to growing a crop in one environment generation after generation. As such, Andean pastoralists can be thought of as the original shuttle breeders, innovatively growing quinoa, kañawa, potatoes, and other tubers in multiple ecological conditions facilitating the development of the plethora of highly adaptable varieties we know today.
References
Anderson E (1952) Plants, man and life. University of California Press, Berkeley
Andrews D (2017) Race, status, and biodiversity: the social climbing of quinoa. Culture, Agriculture, Food and Environment 39(1):15–24
Arbuckle BS, Hammer EL (2019) The rise of pastoralism in the ancient near east. J Archaeol Res 27(3):391–449
Banack SA, Rondón XJ, Diaz-Huamnchuma W (2004) Indigenous cultivation and conservation of Totora (Schoenoplectus californicus, Cyperaceae) in Peru. Econ Bot 58:11–20
Bandy MS (2005) Energetic efficiency and political expediency in Titicaca Basin raised field agriculture. J Anthropol Archaeol 24(3):271–296
Beaule CD (2002) Late intermediate period political economy and household organization at Jachakala, Bolivia. PhD dissertation, University of Pittsburgh, Pittsburgh
Bermann M, Estévez Castillo J (1995) Domestic artifact assemblages and ritual activities in the Bolivian formative. J Field Archaeol 22(4):389–398
Berryman CA (2010) Food, feasts, and the construction of identity and power in Ancient Tiwanaku: a bioarchaeological perspective. PhD dissertation, Vanderbilt University, Nashville
Bonzani RM (2021) Ancient Paria, Bolivia: macrobotanical remains recovered from an administrative site on the royal Inca highway. In: Staller JE (ed) Andean foodways: pre-Columbian, colonial, and contemporary food and culture. Springer, pp 137–186
Borlaug NE (2007) Sixty-two years of fighting hunger: personal recollections. Euphytica 157(3):287–297
Boucher DH (1985) The idea of mutualism, past and future. In: Boucher DH (eds), The biology of mutualism: ecology and evolution. Oxford University Press, Oxford, pp 1–28
Browman DL (1987) Agro-pastoral risk management in the Central Andes. Res Econ Anthropol 8:171–200
Browman DL (1989) Chenopod cultivation, lacustrine resources, and fuel use at Chiripa, Boliva. The Missouri Archaeologist 47:137–142
Browman DL (1997) Political institutional factors contributing to the integration of the Tiwanaku state. In: Manzanilla L (ed) Emergence and change in early urban societies. Plenum Press, New York, pp 229–243
Bruno MC (2006) A morphological approach to documenting the domestication of Chenopodium in the Andes. In: Zeder MA, Bradley DG, Emshwiller E, Smith BD (eds) Documenting domestication: new genetic and archaeological paradigms. University of California Press, Berkeley, pp 32–45
Bruno MC (2008) Waranq Waranqa: ethnobotanical perspectives on agricultural intensification in the Lake Titicaca Basin (Taraco Peninsula, Bolivia). PhD dissertation, Washington University in Saint Louis, St. Louis
Bruno MC (2014a) Quinoa: origins and development. In: Smith C (eds) Encyclopedia of global archaeology, Springer, New York, pp 6215–6222
Bruno MC (2014b) Beyond raised fields: exploring farming practices and processes of agricultural change in the ancient Lake Titicaca Basin of the Andes. Am Anthropol 116(1):130–145
Bruno MC, Hastorf CA (2016) Gifts from the camelids: archaeobotanical insights into camelid pastoralism through the study of dung. In: Capriles JM, Tripcevic N (eds) The archaeology of Andean pastoralism. University of New Mexico Press, Albuquerque, pp 55–66
Bruno MC, Whitehead WT (2003) Chenopodium cultivation and formative period agriculture at Chiripa, Bolivia. Lat Am Antiq 14:339–355
Bruno MC, Pinto M, Rojas W (2018) Identifying domesticated and wild Kañawa (Chenopodium pallidicaule) in the archeobotanical record of the Lake Titicaca Basin of the Andes. Econ Bot 72(2):137–149
Capriles, JM (2011) The economic organization of early camelid pastoralism in the Andean highlands of Bolivia. PhD Dissertation, Washington University in St. Louis, St. Louis
Capriles JM (2014) Mobile communities and pastoralist landscapes during the formative period in the central Alitplano of Bolivia. Lat Am Antiq 25(1):3–26
Capriles JM (2016) The subsistence economy of early Andean pastoralists: zooarchaeological insights from a formative period Wankarani house. In: Capriles JM, Tripcevich N (eds) The archaeology of Andean pastoralism. University of New Mexico Press, Albuquerque, pp 39–54
Capriles JM (2017) Arqueología del pastoralismo temprano de camélidos en el Altiplano central de Bolivia. Instituto Francés de Estudios Andinos, Plural Editores, La Paz
Capriles JM, Moore KM, Domic AI, Hastorf CA (2014) Fishing and environmental change during the emergence of social complexity in the Lake Titicaca Basin. J Anthropol Archaeol 34:66–77
Capriles JM, Calla Maldonado S, Albarracin-Jordan J (2011) Tecnología lítica y estrategias de subsistencia durante los períodos Arcaico y Formativo en el Altiplano Central, Bolivia. Chungara Revista de Antropología Chilena 43:455–468
d’Alpoim Guedes J, Spengler RS (2014) Sampling strategies in paleoethnobotanical analysis. In: Marston JM, d’Alpoim Guedes J, Warriner C (eds) Method and theory in paleoethnobotany. University Press of Colorado, Boulder, pp 77–94
D’Altroy TN (2002) The Incas. Blackwell, Malden, Massachusetts
Danielsen S, Bonifacio A, Ames T (2003) Diseases of quinoa (Chenopodium quinoa). Food Rev Int 19:43–59
Domic AI, Capriles JM, Escobar-Torrez K, Santoro CM, Maldonado A (2018) Two thousand years of land-use and vegetation evolution in the Andean highlands of northern Chile inferred from pollen and charcoal analyses. Quaternary 1(3):32
Fox J (2007) Time and process in an early village settlement system on the Bolivian southern Altiplano. PhD dissertation, University of Pittsburgh, Pittsburgh
Frachetti MD, Smith EC, Traub CM, Williams T (2017) Nomadic ecology shaped the highland geography of Asia’s silk roads. Nature 543:193–198
Fritz GJ, Bruno MC, Langlie BS, Smith BD, Kistler L (2017) Cultigen chenopods in the Americas: a hemispherical perspective. In Social Perspectives on Ancient Lives from Paleothnobotanical Data, pp. 55–75. Springer
García-Granero JJ, Lancelotti C, Madella M, Ajithprasad P (2016) Millets and herders: the origins of plant cultivation in semiarid North Gujarat (India). Curr Anthropol 57(2):149–173
Harris DR (2007) Agriculture, cultivation and domestication: exploring the conceptual framework of early food production. In: Denham TP, Iriarte J, Vrydaghs L (eds) Rethinking agriculture: archaeological and ethnoarchaeological perspectives. Routledge, London, pp 16–35
Harris DR (1989) An evolutionary continuum of people-plant interaction. In: Harris DR, Hillman GC, Foraging and farming: the evolution of plant exploitation. Routledge, London, pp 11–26
Hastorf, CA (2008) The Formative Period in the Titicaca Basin. In: Silverman H, Isbell WH (eds) Handbook of South American archaeology. Springer, New York, pp 545–562
Hastorf CA, Wright MF (1998) Interpreting wild seeds from archaeological sites: a dung charring experiment from the Andes. J Ethnobiol 18:211–2227
Hastorf, Christine A, Whitehead WT, Bruno MC, Wright MF (2006) The movements of maize into middle horizon Tiwanaku, Bolivia. In: Staller JE, Tykot RW, Benz B (eds) Histories of maize: multidisciplinary approaches to the prehistory, linguistics, biogeography, domestication, and evolution of maize. Left Coast Press, Walnut Creek, pp 429–448
Hildebrand EA (2007) A tale of two tuber crops: how attributes of enset and yams may have shaped prehistoric human-plant interactions in Southwest Ethiopia. In: Denham T, Irarte J, Vrydaghs L (eds) Rethinking agriculture: archaeological and ethnoarchaeological perspectives. Routledge, London, pp 273–298
Juggins S (2019) Package Rioja: analysis of science data. https://www.rdocumentation.org/packages/rioja/versions/0.9-21
Khazanov AM (1994) Nomads and the Outside World. 2nd ed. Translated by Julia Crookenden. University of Wisonsin Press, Madison
Kuznar LA (1993) Mutualism between Chenopodium, herd animals, and herders in the south Central Andes. Mt Res Dev 13:257–265
Langlie BS (2018) Building ecological resistance: late intermediate period farming in the south-central highland Andes (CE 1100–1450). J Anthropol Archaeol 52:167–179
Langlie BS (2019) Morphological analysis of late pre-Hispanic Peruvian Chenopodium spp. Veg Hist Archaeobotany 28(1):51–63
Langlie BS, Arkush EN (2016) Managing mayhem: conflict, environment, and subsistence in the Andean Late Intermediate Period, Puno, Peru. In: VanDerwarker AM, Wilson GD (eds) The Archaeology of Food and Warfare. Springer, pp. 259–289
Langlie BS, Hastorf CA, Bruno MC, Bermann M, Bonzani RM, Condarco WC (2011) Diversity in Andean Chenopodium domestication: describing a new morphological type from La Barca, Bolivia 1300-1250 BC. J Ethnobiol 31(1):72–88
Lennstrom HA, Hastorf CA (1992) Testing old wives’ tales in paleoethnobotany: a comparison of bulk and scatter sampling schemes from Pancan, Peru. J Archaeol Sci 19(2):205–229
Lezama-Núñez PR, Santos-Fita D, Vallejo JR (2018) Herding ecologies and ongoing plant domestication processes in the Americas. Front Plant Sci 9:649
Logan AL, Hastorf CA, Pearsall DM (2012) “Let’s drink together”: early ceremonial use of maize in the Titicaca Basin. Lat Am Antiq 23(3):235–258
Lord E, Collins C, deFrance S, LeFebvre MJ, Pigière F, Eeckhout P, Erauw C, Fitzpatrick SM, Healy PF, Martínez-Polanco MF, Garcia JL, Roca ER, Delgado M, Urriago AS, Léon GAP, Toyne JM, Dahlstedt A, Moore KM, Diaz CL, Zori C, Matisoo-Smith E (2020) Ancient DNA of guinea pigs (Cavia spp.) indicates a probable new center of domestication and pathways of global distribution. Sci Rep 10(1):8901.
Magne J, Columba M (2011) Vegetación nativa en la región de Chuquiña y La Joya: Oruro - Bolivia. Arte y Papel, Oruro, Bolivia
Marshall F, Reid REB, Goldstein S, Storozum M, Wreschnig A, Hu L, Kiura P, Shahack-Gross R, Ambrose SH, (2018) Ancient herders enriched and restructured African grasslands. Nature 561(7723):387-390
Marston JM (2014) Ratios and simple statistics in paleoethnobotanical analysis: data exploration and hypothesis testing. In: Marston JM, d’Alpoim Guedes J, Warinner C (eds) Method and theory in paleoethnobotany. University Press of Colorado, Boulder, pp 163–179
McClure, SB (2015) The pastoral effect: niche construction, domestic animals, and the spread of farming in Europe. Curr Anthropol 56(6):901–910
McAndrews TL (2005) Wankarani Settlement Systems in Evolutionary Perspective: a study in early village-based society and long-term cultural evolution in the South-Central Andean Altiplano. University of Pittsburgh, Plural Editores, Pittsburgh
Mengoni-Goñalons GL, Yacobaccio HD (2006) The domestication of south American camelids: a view from the south-Central Andes. In: Zeder MA, Bradley D, Emshwiller E, Smith BD (eds) Documenting domestication: new genetic and archaeological paradigms. University of California Press, Berkeley, pp 228–244
Miller NF (1988) Ratios in paleoethnobotanical analysis. In: Hastorf CA, Popper VS (eds) Current Paleoethnobotany: analytical methods and cultural interpretations of archaeological plant remains. The University of Chicago Press, Chicago pp 72–85
Miller NF, Smart TL (1984) Intentional burning of dung as fuel: a mechanism for the incorporation of charred seeds into the archaeological record. J Ethnobiol 4(1):15–28
Molina Rivero R (2006) De memorias e identidades: los Aymaras y Urus del sur de Oruro. Instituto de Estudios Bolivianos, La Paz
Moore KM (2016) Early domesticated camelids in the Andes. In: Capriles JM, Tripcevich N (eds) The archaeology of Andean pastoralism. University of New Mexico Press, Albuquerque, pp 17–38
Nordstrom C (1990) Evidence for the Domestication of Chenopodium in the Andes. In Report to the National Science Foundation. University of California, Berkeley Paleoethnobotany Laboratory Reports #19, Berkeley
Orlove BS (1991) Mapping reeds and reading maps: the politics of representation in Lake Titicaca. Am Ethnol 18(1):3–38
Pearsall DM (1989) Adoption of prehistoric hunter-gatherers to the high Andes: the changing role of plant resources. In: Harris DR, Hillman GC (eds) Foraging and farming: the evolution of plant exploitation. Unwin Hyman, London, pp 318–332
Pearsall DM (2015) Paleoethnobotany: a handbook of procedures, 3rd edn. Academic Press, San Diego
Planella MT, López ML, Bruno MC (2015) Domestication and prehistoric distribution. In: Bazile D, Bertero D, Nieto C, State of the art report on quinoa around the world in 2013. FAO, Rome, pp 29–41
R Core Team (2020) R: A Language and Environment for Statistical Computing. R Foundation for Statistical Computing, Vienna, Austrica. URL https://www.R-project.org/
Rindos D (1984) The origins of agriculture: an evolutionary perspective. Academic Press, Orlando, Florida
Rumold CU, Aldenderfer MS (2016) Late archaic–early formative period microbotanical evidence for potato at Jiskairumoko in the Titicaca Basin of southern Peru. Proc Natl Acad Sci 113(48):13672–13677
Sauer CO (1952) Agricultural origins and dispersals. The American Geographical Society, New York
Sempertegui AC, Bahoz EG, Canelas EL, Cáceres IM (2005) Más allá de las pajas y espinas: Biodiversidad en el Municipio de Oruro (Communidades Cochirarya - Iroco - Chuzekery). Latinas Editorers, Oruro, Bolivia
Spengler R, Frachetti M, Doumani P, Rouse L, Cerasetti B, Bullion E, Mar’yashev A (2014) Early agriculture and crop transmission among bronze age mobile pastoralists of Central Eurasia. Proc R Soc B Biol Sci 281(1783):20133382
Stanish, CS (2003) Ancient Titicaca: The Evolution of Social Complexity in Southern Peru and Northern Bolivia. University of California Press, Los Angeles
Watson PJ (1976) In pursuit of prehistoric subsistence: a comparative account of some contemporary flotation techniques. Midcont J Archaeol 1(1):77–100
Wheeler JC (1995) Evolution and present situation of the south American Camelidae. Biol J Linn Soc 54(3):271–295
Whitehead WT (2007) Exploring the wild and domestic: paleoethnobotany at Chiripa, a formative site in Bolivia. PhD Dissertation, University of California, Berkeley
Wickham H (2016) ggplot2: elegant graphics for data analysis. Springer-Verlag, New York
Winkel T, Alvarez-Flores R, Bommel P, Bourliaud J, Chevarria-Lazo M, Cortes G, Cruz P, Del Castillo C, Gasselin P, Joffre R (2015) The southern altiplano of Bolivia. In: Bazile D, Bertero D, Nieto C (eds) State of the art report on quinoa around the world in 2013. FAO, Rome, pp 362–377
Wright MF, Hastorf CA, Lennstrom HA (2003) Pre-Hispanic agriculture and plant use at Tiwanaku: social and political implications. In: Kolata AL (ed) Tiwanaku and its hinterland: archaeology and paleoecology of an Andean civilization. Smithsonian Institution Press, Washington DC, pp 384–403
Acknowledgements
We would like to thank Maria Bruno who helped to coordinate the flotation of the samples as well as analysis of two of those included in this study. We also thank Sergio Calla Maldonado, Alejandro Barrientos Salinas, Juan Carlos Segurola, Javier Aguilar, Edwin Choque, Alex Flores, Israel Quispe, Vidal Colque, Mauricio Rocha, Jorge Ustarez, and Armando Vargas for their participation during fieldwork. Fieldwork and analysis in Bolivia were facilitated by the communities of Cochiraya, Iroco, and Chuzekery, Inti Raymi Mining Company, Universidad Técnica de Oruro, Universidad Mayor de San Andrés, and the Unidad Nacional de Arqueología of Bolivia of the Bolivian Ministerio de Culturas y Turismo. We would also like to thank Dave Browman and Gayle Fritz for the inspiration and feedback on this work.
Funding
Research in Bolivia was supported by the National Science Foundation DDRIG BCS# 0737793, the Lambda Alpha Graduate Overseas Research Charles R. Jenkins Award, and the Graduate School of Arts & Sciences at Washington University in St. Louis.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
The authors declare no competing interests.
Additional information
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
About this article

Cite this article
Langlie, B.S., Capriles, J.M. Paleoethnobotanical evidence points to agricultural mutualism among early camelid pastoralists of the Andean central Altiplano. Archaeol Anthropol Sci 13, 107 (2021). https://doi.org/10.1007/s12520-021-01343-y
Received:
Accepted:
Published:
DOI: https://doi.org/10.1007/s12520-021-01343-y