
Abstract
Despite the widespread application of high-resolution quantitative methods in bone taphonomy, very few studies have focused on projectile impact marks. Therefore, in a previous work, we explored the potential of 3D microscopy in distinguishing bone hunting injuries from other taphonomic marks, developing a widely applicable diagnostic framework based on experimental data and focused on Late Epigravettian projectiles. This paper aims to continue that research by applying 3D morphometrical analysis to zooarcheological bone surfaces, in order to verify the validity and feasibility of this method and evaluate the reliability of the experimental record. Here, we present the detailed analysis of a projectile impact mark, found on a rib of Ursus arctos from the Late Epigravettian site of Cornafessa rock shelter. The injury, located on the rib’s external surface, consists of a drag with several flint fragments embedded. X-ray μCT volume rendering and SEM imaging allowed us to analyze bone microstructure and drag’s qualitative features, while 3D measurements, processed through statistic, confirmed the interpretation of this mark as a hunting injury. The drag’s morphometric features are consistent with the experimental ones, connecting this mark to Late Epigravettian composite projectiles and declaring this evidence as the first direct proof of a bear hunted by using bow and arrow.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Studies of projectile impact marks (PIMs; after O’Driscoll and Thompson 2014) on bones still play a minor role in zooarcheological analysis. Recent research has identified punctures and the occasional presence of embedded lithic fragments as relevant features for recognizing archeological hunting injuries (Bachechi et al. 1997; Boëda et al. 1999; Bratlund 1991, 1996; Churchill et al. 2009; Fernández-Crespo 2016, 2017; Fischer 1985; Gaudzinski-Windheuser 2016; Leduc 2012; Milo 1998; Morel 1998; Münzel and Conard 2004; Nikolskiy and Pitulko 2013; Noe-Nygaard 1974; O’Driscoll and Thompson 2018; Péquart et al. 1937; Pitulko et al. 2016; Pöllath et al. 2017; Rust 1943), but the design of a systematic protocol for the study of this rare archeological material is just beginning. Besides, the commonly accepted assumption that PIMs would be barely recognized (Castel 2008; Letourneux and Pétillon 2008; Noe-Nygaard 1974; Smith et al. 2007), along with the limitation of experimental tests and the lack of uniformity in terminology and methodology, has hindered the identification of PIMs from becoming a standard in zooarcheological analysis. New impulse in this direction has recently come from several experimental studies focused on projectile traumas (Castel 2008; Letourneux and Pétillon 2008; O’Driscoll and Thompson 2014; Pétillon and Letourneux 2003, 2008; Pétillon et al. 2011; Smith et al. 2007; Wood and Fitzhugh 2018; Yeshurun and Yaroshevich 2014) and from the application of quantitative methods in bone surface modification analyses (Arriaza et al. 2017; Bello and Soligo 2008; Bello et al. 2009; Boschin and Crezzini 2012; Courtenay et al. 2017; Maté-González et al., 2015, 2017; Moretti et al. 2015; Pante et al. 2017; Val et al. 2017; Wallduck and Bello 2018; Yravedra et al. 2017). Nevertheless, only few studies have been specifically designed to determine differences between marks produced by projectile impacts and those generated by taphonomic effects (O’Driscoll and Thompson 2014, 2018), while most of them focused on cut mark characterization through the application of high-resolution 3D quantitative methods (Bello and Soligo 2008; Boschin and Crezzini 2012; Courtenay et al. 2017; Maté-González et al. 2015, 2017; Oxilia et al. 2017; Pante et al. 2017; Wallduck and Bello 2018; Yravedra et al. 2017). These methods were also recently applied to different bone lesions as percussion damages related to the extraction of bone marrow and knapping marks due to shaping and retouching of lithic artifacts (Abrams et al. 2014; Bello et al. 2013; Van Kolfschoten et al. 2015).
In a previous study, we explored the potential of 3D microscopy in distinguishing PIMs from other taphonomic marks, developing a diagnostic framework based on experimental data (Duches et al. 2016). This widely applicable method is based on the collection of reproducible morphometrical parameters that can be processed through statistical analysis, allowing for a reliable distinction of PIMs from cut marks and carnivore tooth marks. This work provided not only specific evidence for the identification of hunting traumas caused by Late Epigravettian projectile implements but also comparative data for taphonomic analyses of bones from other Late Glacial contexts (Duches et al. 2016). The present article aims to continue this research applying 3D morphometric analysis to zooarcheological bone surfaces, in order to test the following: (a) confirm the validity and feasibility of the method; (b) evaluate the reliability of the experimental record; (c) verify the bearing of post-depositional alterations in PIMs identification; and (d) estimate the preservation of PIMs qualitative features on archeological remains.
Material and methods
Late Pleistocene adaptations in the Alps
The peopling of northeastern Italy after the Last Glacial Maximum was a gradual process marked by the progressive colonization of new territories that had been previously abandoned, following changes of vegetation and animal distribution (Naudinot et al. 2014; Ravazzi et al. 2007). Late Epigravettian penetration, firstly limited to valley floors and high plateaus around 500 m asl (Aimar et al. 1992; Peresani et al. 2011), reaches mid-altitude territories during the second part of the Late Glacial interstadial with the full development of a logistical occupation network at the limit between coniferous woods and alpine prairies (Bertola et al. 2007). This organization corresponds to a seasonal mobility strategy that sometimes included sites that are functionally complementary to each other and located at different altitudes (Bertola et al. 2007). Technological analysis of the lithic industries from the most relevant sites has highlighted a progressive simplification of the lithic production system throughout the Late Glacial interstadial (Montoya 2008; Montoya and Peresani 2005) associated with the persistence of standardized lithic backed tools used as projectile implements (Duches et al. 2017). The existence of specific retouched forms, characterized by distinct metric and morphological parameters, reveals as many mental templates developed by the Epigravettian craftsmen in response to specific functional requirements and constraints (Duches et al. 2017). These constraints fit within the conception of the bow-arrow delivery system, whose generalized use in the Epipaleolithic complexes of Europe has been hypothesized by many scholars (Caspar and De Bie 1996; Cattelain 1997; Pelegrin 2000; Plisson 2005; Valentin 2008). Functional analyses carried out on Late Epigravettian industries of northeastern Italy (Lemorini and Rossetti 2004; Lemorini et al. 2006; Peresani et al. 2011; Ziggiotti 2006, 2008) confirmed the exclusive use of lithic backed tools as elements in hunting projectiles.
Cornafessa rock shelter
The Cornafessa rock shelter, located at about 1240 m asl on the Monti Lessini Plateau (Trentino), is a Late Epigravettian site of recent discovery (Fig. 1). It opens in a wide overhanging Late Jurassic limestone wall (Oolite di San Vigilio; Avanzini and Masetti 2007), about 28 m long and projecting up to 7 m. The Paleolithic settlements at Cornafessa rock shelter were discovered in 2014 after a test-pit; an archeological excavation was carried out in 2015 by MUSE-Science Museum of Trento. This first excavation, involving a total area of 6 m2, has established the existence of a layered filling deposit characterized by at least one anthropic unit, placed at a depth of about 2 m from ground level. The layer, dated to the Younger Dryas (charcoal-LTL15976A: 10,080 ± 70 BP, 10,050–9350 cal BC at 95.4%; charcoal-LTL15977A: 10,171 ± 70 BP, 10,200–9450 cal BC at 95.4%; dates modeled in OxCal v.3.10 using IntCal13 calibration curve; Reimer et al. 2013), has yielded a Late Epigravettian lithic industry and 827 faunal remains in a good state of preservation. The lithic assemblage consists of 130 artifacts ≥ 1 cm, belonging to different phases of the reduction sequence. At present, there are no retouched tools, except for few undetermined fragments of backed points and bladelets. The archeological investigation are still in progress.

Location and picture of Cornafessa rock shelter
Analytical methods
Taxonomic and skeletal identifications were based on the complete alpine fauna reference collection of MUSE-Science Museum of Trento. Taphonomic analyses were carried out using a stereomicroscope (Leica M 165C with magnification from 0.75× to 125×), a scanning electron microscope (ZEISS EVO 40 XVP) and a coupled energy dispersive X-ray detector (EDX) at MUSE-Science Museum of Trento. The SEM was operated in variable pressure mode (chamber pressure ~ 14 Pa), enabling backscattered electron images to be obtained at magnifications between 70× and 500× without the application of a conducting layer on the specimen. EDX microanalysis was carried out to determine the elemental composition of surface inclusions, using an Oxford Instrument x-act PentaFET precision detector and Aztec 3.1 SP1 software.
In order to identify the nature of surface alterations and to distinguish different taphonomic agents, we referred to well-established literature (Binford 1981; Blumenschine and Selvaggio 1988; Capaldo and Blumenschine 1994; Domínguez-Rodrigo et al. 2009; Domínguez-Rodrigo and Piqueras 2003; Fernandez-Jalvo and Andrews 2016; Fisher 1995; Outram 2002; Pickering and Egeland 2006; Potts and Shipman 1981; Selvaggio and Wilder 2001; Shipman 1981; Shipman and Rose 1984; Villa and Mahieu 1991). The descriptive criteria and the terminology used for PIMs, follows O’Driscoll and Thompson (2014) as already discussed in Duches et al. (2016).
The 3D analysis of bone surfaces was carried out using a Hirox Digital Microscope KH-7700, with MXG-10C body, OL-140II lens and AD-10S Directional Lighting Adapter (Arrighi et al. 2016; Boschin and Crezzini 2012; Crezzini et al. 2014; Moretti et al. 2015) at the University of Siena. Metrical parameters of drags were recorded according to criteria already published (Duches et al. 2016, Fig. 4), using one cross-section per mark in its median part (Fig. 2). In addition, the ratio between breadth at the top (BT) and depth of cut (DC) was calculated (RTD). Measures and profiles taken on archeological specimens were compared to drags produced experimentally by Late Epigravettian projectiles on mouflon carcasses (Duches et al. 2016). Of the total 35 drags available, only 7 could be analyzed through 3D microscopy due to the statistical need of recording all dimensional parameters. The marks were indeed frequently incomplete because of either the total removal of a bone edge or the absence of a continuous floor in the case of spongy or trabecular bones.

Measurements taken on drag marks and cut marks profiles. DC, depth of cut; BT, breadth at the top of the cut; BF, breadth at the floor of the cut; GD/SD, greatest and smallest distances from the middle of the floor to the edges; OA, opening angle
Besides, the values were compared to experimental cut marks (n = 93), of which 27 were inflicted on cattle autopodials (22 using an unretouched flake, 5 using a retouched implement) (Boschin and Crezzini 2012) and 66 were produced with unretouched flakes during skinning experiments on cat carcasses (Crezzini et al. 2014). All bone modifications considered on this paper (including the samples used for comparisons) have been directly scanned by two of the authors (FB and JC), who also collected all the measurements processed in the various statistical tests (Table S1). Measurements were collected on the cross-sections using the software directly installed in the digital microscope and then processed using the PAST software (Hammer et al. 2001).
A rib of Ursus arctos from Cornafessa rock shelter and four modern U. arctos ribs belonging to different age classes were analyzed by microfocus X-ray computed tomography (μCT) at the Multidisciplinary Laboratory of the “Abdus Salam” International Centre of Theoretical Physics (Trieste, Italy), using a system specifically designed for the study of paleontological and archeological materials (Tuniz et al. 2013). The μCT acquisitions of the specimens were carried out by using a sealed X-ray source (Hamamatsu L8121-03) at a voltage of 110 kV, a current of 90 μA, and with a focal spot size of 5 μm. The X-ray beam was filtered by a 1-mm-thick aluminum absorber. A set of 1440 projections of the sample were recorded over a total scan angle of 360° by a flat panel detector (Hamamatsu C7942SK-25) with an exposure time/projection of 1.5 s. The resulting μCT slices were reconstructed using the commercial software DigiXCT (DIGISENS) in 32-bit format and an isotropic voxel size of 18 μm. Using the Avizo v.8 software (Visualization Sciences Group Inc.), a semi-automatic threshold-based segmentation was carried out in order to separate the bone component from the interstitial air between the trabeculae (Bouxsein et al. 2010; Coleman and Colbert 2007; Fajardo et al. 2002).
Experimental data and 3D analysis in PIM identification
As already mentioned, a previous ballistic experimentation provided comparative data for hunting lesions identification, with specific clues for the distinction of trauma caused by Late Epigravettian projectiles on medium-sized ungulates (Duches et al. 2016). In particular, 160 arrows armed with lithic backed points and backed (bi)truncated bladelets have been shot against five complete carcasses in a realistic simulation of game hunting, generating a total of 70 hunting injuries on bones. Each lesion has been classified by following O’Driscoll and Thompson (2014) and analyzed, when possible, through 3D digital microscopy with the acquisition of multiple morphometrical parameters useful for statistical processing and comparison (Duches et al. 2016).
As a main methodological outcome, drag and puncture marks have been recognized as the only diagnostic categories in PIMs identification. Crucial for their interpretation are the following elements: (a) the location of the mark; (b) the presence of flaking and/or cracking as secondary features of both categories; (c) the occurrence of specific morphometrical values, different for drags and punctures (detailed below); and (d) the presence of stone embedded characterized by diagnostic impact fractures.
Drag marks were the most frequent experimental trauma (50%) and, consequently, the most likely preserved on zooarcheological material. This category was recorded mainly on ribs (40%), vertebrae (34.3%), and on long bones such as the radius and ulna (14.3%). On the most resistant bones the drag profile is extremely sharp and clean, with frequent unilateral flaking (48.6%) and cracking (34.3%) (Fig. 3). In the case of a total removal of the bone edge, drags are characterized by a polygonal shape (Duches et al. 2016, Fig. 7). The 3D analysis compared drags to carnivore tooth marks and cut marks (produced both by unmodified flakes and retouched implements) using the parameters listed in the analytic method section (see also Fig. 2). Drags turned out to be significantly different from other taphonomic marks, especially considering the breadth at the cut floor (considerably wider than cut marks while comparable to that of tooth marks), the depth of cut (considerably higher than other taphonomic marks), the RTD index (significantly different suggesting a more U-shaped profile than other taphonomic marks), and the opening angle (narrower and more standardized in drags, ranging from 65.8° to 94.5°) (raw data and statistical analysis in Duches et al. 2016).

Experimental drag marks on mouflon bones made by Late Epigravettian lithic projectiles. a and b: stereomicroscope image, 3D rendering and cross-section of drags on two specimen of radius; c: stereomicroscope image, 3D rendering and cross-section of a drag on rib
Puncture marks were the second most frequent experimental bone injury (24.3%) and were recorded on the entire skeleton. In the case of the complete perforation of the bone, the projectile left an imprint characterized by a more or less regular polygonal outline. Cracking occurs in the 48.1% of the punctures. The 3D analysis compared punctures to carnivore pits and corrosion cavities considering the following parameters: opening area (A), volume (V), depth of cavity (D), and the ratio between the opening area and volume of pits (RAV) (Duches et al. 2016). Puncture marks resulted significantly deeper than carnivore tooth pits and characterized by a statistically lower RAV (Duches et al. 2016, Fig. 14). Morphological features such the contour of the opening area and the shapes of slopes and floor led to easily distinguish impact punctures from corrosion cavities.
Besides, it is noteworthy that some peculiar features of both drags (drag with sharp edges and polygonal shape) and punctures (polygonal outline) turned out to be strictly related to Late Epigravettian projectiles, confirming the hypothesis of a significant relationship between projectile design and PIMs morphometrical features (Duches et al. 2016).
Thus, the outcomes of 3D digital analysis offer new clues in PIM identification and distinction from other taphonomic marks, qualifying drags and punctures as reliable archeological indicators of prey capture modalities.
Results
Cornafessa rock shelter bone assemblage
The faunal assemblage analyzed includes 827 bone fragments, with a total weight of 108.7 g (Table 1). Only 18 fragments could be taxonomically classified (Table 2). The low NISP value is due to the high rates of bone breakage (more than 96.5% are smaller than 2 cm) and thermal alteration (25%). These conditions are consistent with a set of syn- and post-depositional processes observed in bone assemblages from other Late Epigravettian contexts of northeastern Italy (for a synthesis, see Bertola et al. 2007; Fiore and Tagliacozzo 2005). The remains altered by mechanisms of mobilizations, like trampling, are few (0.2%). The most common cause of degradation is root etchings (2.9%), with a variety of patterns and dimensions. Some fragments show manganese staining (0.7%) and concretions (0.4%). No traces of carnivore and rodent tooth marks are recorded on bone surfaces. At present, the small sample examined does not allow for relevant paleo-ecological considerations, except for the coexistence in the same site of faunas typical of rocky habitats as ibex and species featuring lower elevations (grasslands, wetlands, and forests) as red deer and bear. The scarcity of the sample and the fair preservation of bone surfaces, corresponding to weathering stage 1 or 2 according to Behrensmeyer (1978), allowed us to recognize few stages of carcass exploitation. Defleshing and periosteum removal are well represented by scraping marks along the surface of unidentified medium and large mammals’ diaphysis altered by fire (n = 6; 0.7%; Table 2); conversely, no butchering cut marks or percussion marks have been identified on this small zooarcheological assemblage.
Analysis of the projectile impact mark
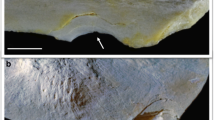
Except for the scrapings, the only anthropic mark identified is a notch located on a rib of U. arctos, which exhibits some morphological traits consistent with a PIM (Fig. 4). The bone is fragmented and corresponds to a rib probably in the range between the 4th–8th: the body (corpus costæ) with the external costal groove (sulcus costæ), the cranial and the caudal edges have been mostly preserved, as has the neck (collum costæ) and the tuberosity of the muscoli longissimi; conversely, the tubercle (tuberculum costæ) and the head (caput costæ) are very deteriorated.

Ursus arctos rib from Cornafessa rock shelter with taphonomic features highlighted. Green lines: roots marks; red lines: scraping marks; blue lines: trampling marks; black arrow: PIM. a: PIM and root etchings; b: scraping marks
Morphometric features are consistent with both a juvenile bear of regular dimensions (1–5 years old; 80–100 kg) and an unusually small adult bear (about 80 kg), as suggested by a reference bear skeleton from Trentino stored in the historical zoological collections at the MUSE-Science Museum of Trento (record code: MSN 105533). The μCT analysis of the Cornafessa U. arctos rib has allowed to detail its age-class. As already reported by Boschin et al. (2015), this method can indeed provide bone micro-structural information useful to estimate the age at death of animal remains. The microstructure of juvenile and young U. arctos ribs, compared to that of adult samples, shows a more porous cortical portion and thicker trabeculae (Fig. 5). Such condition is probably related to the increase in bone volume fraction in growing bones, before the architectural adaptation to external loading during life (Tanck et al. 2001). According to its bone microstructure (Fig. 5e), the Cornafessa specimen is probably a juvenile individual.

Longitudinal and transversal cross-sections of five bear ribs. a: 2 months old individual; b: young individual not fully grown; c: adult individual; d: adult individual of small body size; E: Cornafessa specimen
The surface of the specimen was altered by root etching, which produced easily recognizable grooves (Fig. 4a) (Lyman 1994). Rare manganese staining and trampling marks with the characteristic U-shape are also present (Domínguez-Rodrigo et al. 2009). Longitudinal scraping marks in lateral and medial portions of the rib (Fig. 4b) suggest that the bone was partially worked on and then abandoned. The scrapings are in fact fully consistent with the traces found on other Late Epigravettian bone tools (Cristiani 2008; Gurioli 2008). Moreover, the only bone point yielded from Cornafessa rock shelter shows very similar scrapings in all the worked surfaces (Fig. 14).
The PIM, located in the middle of the rib’s external surface, consists of a drag with two small flint fragments embedded. The mark is about 6 mm long and presents a wider breadth in its initial part. Although unilateral flaking is visible around the projectile’s entrance point, the drag profile is extremely sharp and clean for its entire length (Fig. 6a, b). Cracking is only observable around the stone embedded at the top of the mark (Fig. 6c). Though this one was visible at low magnification, the existence of a second fragment fixed within the drag’s floor and covered by sediment was revealed only by μCT (Fig. 7). These two fragments are perfectly aligned to the drag’s orientation and parallel to each other, whereas they show different depths. Unfortunately, it was not possible to classify any of them either in terms of tip portion or fracture typology.

Stereomicroscope images of Cornafessa projectile impact mark. a Evidence of straight trajectory of the lesion. b Wider breadth of the drag in its initial part and flaking of one side of the mark. c Evidence of directional microstriations on the drag’s floor

μCT virtual renderings of Cornafessa rib showing the stone fragments still embedded in the bone (a, b) and metallographic microscope images of the lithic fragments using bright field illumination (c, d)
SEM analysis showed several other small lithic fragments embedded in the drag’s floor and let us evaluate the conservation of PIM qualitative features (Fig. 8). As clearly visible in Fig. 8, the projectile’s impact dug a trapezoidal groove, causing flaking on one side and leaving directional microstriations on the opposite wall. EDX spectra from four selected areas allow to characterize the lithic inclusions and distinguish them clearly from the embedding bone tissues (Fig. 9). The mark has been documented from different orientations because of the uneven topography of the drag and the consequent problem in the collection of elemental data from shielded areas. The large silicon and oxygen peaks in Fig. 9, spectra 1 to 3 (S1–S3), indicate the presence of silicon dioxide (SiO2), confirming the visual attribution of the fragments to the Lower Cretaceous Maiolica flint lithotype (Bertola 2016; Bosellini et al. 1978; Weissert 1981). Analyses of the surrounding matrix (Fig. 9f) showed the presence of calcium and phosphorus peaks typical of bone. Smaller peaks representing carbon and aluminum are probably associated with environmental contamination from soil and handling. Furthermore, EDX maps of elemental distribution distinctly localize stone embedded in areas marked by high silicon levels and isolate them from the surrounding calcium-rich matrix (bone) (Fig. 9c, d, g, h).

SEM secondary electron images of Cornafessa projectile impact mark

a, e SEM secondary electron images of Cornafessa projectile impact mark with the location of the four areas analyzed by energy dispersive X-ray spectroscopy (EDX; S1–4). The mark was documented from different orientations because of the uneven topography of the drag and the consequent problem in the collection of elemental data from shielded areas. b, f EDX spectra corresponding to the four areas analyzed; the horizontal axis of the spectrum is the X-ray energy and the vertical axis is the total number of X-rays counted. The peak at zero keV is created by the instrument electronics as a reference. c, g BSE images and combined elemental maps shown as false color: phosphorus in light blue, silicon in blue, oxygen in green and calcium in orange. d, h Discrete EDX maps of elemental distribution
The measurements taken on the median cross-section of the drag are: depth of cut (DC) = 478 μm, breadth at the floor (BF) = 112 μm, breadth of the top (BT) = 556 μm, opening angle (OA) = 34.6°, greatest and smallest distances from the middle of the floor to the edges (respectively GD and SD) = 309 and 247 μm. The morphology of this drag resembles that of experimental PIMs (Duches et al. 2016): it is deeper than all observed cut marks and has a narrower opening angle (Figs. 10 and 11). Moreover, the floor of the drag is wide. The value of the RTD index (ratio between BT and DC) lies within the variability of experimental drags and outside that of cut marks. A principal components analysis was performed on measurements from the whole sample (raw data in Electronic Supplementary Material) considering the specimen from Cornafessa separately from experimental PIMs (Fig. 12). All the measurements are considered in this analysis: DC, BT, BF, GD, SD, and OA. The resulting outcome counts for 89.1% of sample variability. PC1, which counts for the 71.2% of variability, is mainly related to the size of marks: the greater the values are, the wider and deeper the marks, but the narrower the opening angle. Regarding PC1, drags are significantly different from cut marks (Mann-Whitney U test: U = 2; p = 1.35E−05). PC2, which counts for the 17.9% of variability, is related to the shape of marks: the greater the values are, the more V-shaped and the more shallower the cross-sections and the wider the opening angle. The Cornafessa drag is clearly far from the variability of cut marks while it is close to (although still outside) the convex hull of experimental PIMs. The position of Cornafessa specimen in the PCA scatter plot is mainly influenced by its depth and narrow opening angle, confirming data presented in the boxplots (Fig. 12).

3D rendering and median cross-section of the Cornafessa drag

Distribution of values of three parameters measured on the analyzed sample. RTD, ratio between the breadth at the top and the depth; DC, depth of cut; OA, opening angle; PIMs: n = 7; experimental cut marks made by unretouched flakes: n = 88; experimental cut marks made by retouched implements: n = 5; CF, Cornafessa; PIMs, projectile impact marks; UF, unretouched flakes; RI, retouched implements

PCA performed on the sample. Components 1 and 2 count respectively for 86.1 and 9.8% of the sample variability. Blues squares: cut marks made by unretouched flakes; red plus: cut marks made by retouched implements; green crosses: experimental PIMs; blue asterisk: Cornafessa drag
Discussion
Methodological outcomes and ballistic considerations
The morphometric features of the archeological drag are consistent—even if not fully correspondent—to the experimental ones (Duches et al. 2016), confirming its interpretation as a PIM. This result also proves the usefulness of 3D in distinguishing PIMs from other taphonomic marks and the reliability of the experimental data assumed as reference in this work. With regard to cut marks, a clear distinction is defined by statistic: drags depth is significantly higher than that of cut marks (Mann-Whitney U test: U = 9; p = 1.963E−05); RTD index of drags is lower than that of cut marks (Mann-Whitney U test: U = 128; p = 0.007); lastly, the opening angle is narrower in drags (Mann-Whitney U test: U = 112; p = 0.004). In comparison to indentations found on bone or antler hammers used as knapping tools (Bello et al. 2013; Van Kolfschoten et al. 2015), drags generally look different in shape, being deeper (and sometimes wider) than scores measured by Bello et al. (2013) and Van Kolfschoten et al. (2015, supplementary data). The Cornafessa drag, in particular, is deeper (0.58 mm) than any other score described by these authors. Since only length, depth, and width were measured on knapping marks, a statistical comparison with PIMs is difficult. In particular, it would be necessary to know the RTF/RTD indexes of knapping scores.
Post-depositional alterations of bone surfaces—in the form of roots etching, corrosion, manganese staining, and trampling marks—do not compromise the feasibility of this method on the Cornafessa specimen, while the effects of weathering and fluvial abrasion in PIMs identification remain difficult to evaluate. Nevertheless, even considering the possibility of an alteration of the original profile in archeological drags, some morphometric parameters, such as depth, opening angle, and RTD index, remain crucial in PIMs identification. With regard to the conservation of qualitative features, SEM analyses show that some microstriations on the walls of the drag are preserved.
Concerning the small lithic fragments embedded, this new archeological evidence contributes to the debate around the diagnostic significance of this feature in drags identification. Although some authors suggested that small lithic fragments could be trapped in bones either during butchering actions or projectile impacts (Milo 1998; Parsons and Badenhorst 2004), there are hitherto no evidence of experimental or archeological stone embedded in cut marks. We can thus support the hypothesis that only a considerable amount of force could plunge lithic fragments into bones (O’Driscoll and Thompson 2014), making this feature a relevant indicator for drags identification. It is however necessary to consider that stone embedded were also documented in knapping marks left on bones during shaping and retouching of lithic artifacts, as the result of high-energy impacts (Abrams et al. 2014; Bello et al. 2013; Bordes 1974; Mallye et al. 2012; Olsen 1989; Van Kolfschoten et al. 2015). In these cases though, the association of lithic fragments with a series of characteristic features of knapping damages (i.e., gouges, pits, scores, scratches, and internal striations; Van Kolfschoten et al. 2015) allows for an easy recognition of taphonomic causality both on experimental and archeological material (Abrams et al. 2014; Bello et al. 2013; Bordes 1974; Mallye et al. 2012; Olsen 1989; Van Kolfschoten et al. 2015). Even though the fragments embedded in Cornafessa drag are small and difficult to classify, they offer some elements of interpretation. We can assume, in fact, that the deeper one results from the initial impact of the lithic point that dug a wider and deeper groove before the breakage, while it is not clear if the second, more superficial, belongs to the point itself or to a different implement assembled on the same shaft. Given the belonging of the fragments to the same lithotype (i.e., Maiolica; Weissert 1981), EDX spectra are inconclusive in distinguishing different implements. Based on their alignment and different depth, however, it seems difficult to ascribe the second piece to the same backed point, already fragmented. It is more likely that another implement arranged behind the point hit in turn against the bone surface, leaving a slice of the cutting edge exposed from the hafting. Such evidence thus supports the hypothesis of a Late Epigravettian composite projectile, with a backed point hafted as piercing element and backed (bi)truncated bladelets functioning as lateral cutting elements. This arrangement has already been suggested by many authors for the Late Epigravettian of northeastern Italy, on the basis of backed tool morphological features and use-wear analysis (Agogué and Dalmeri 2005; Duches et al. 2017; Ziggiotti 2008). Considering drags produced experimentally by Late Epigravettian backed implements, we verified that composite projectiles could create drags with variable width and depth along the profile, due to the actions of multiple cutting elements against the bone (Fig. 13).

Experimental drag marks on bones made by Late Epigravettian composite projectiles. a: stereomicroscope image of a drag on radius; b: stereomicroscope image of a drag on rib
Regarding the weapon system used to launch this projectile, further discussion is required. The hypothesis of a general widespread in Europe of the bow-arrow delivery system during the Late Upper Paleolithic has been assumed by many scholars for different chrono-cultural contexts (Caspar and De Bie 1996; Cattelain 1997; Duches et al. 2017; Pelegrin 2000; Plisson 2005; Serwatka 2018; Valentin 2008). Although Stelmoor findings (12,600–11,500 cal BP; Beckhoff 1968; Litt and Stebich 1999; Rust 1943) represent the oldest archeological evidence, considerations supporting a previous introduction of this technology in Europe deal with the progressive miniaturization of lithic projectile elements, the disappearance of spear-throwers from archeological contexts, the transformation of hunting strategies, and the evolution of climatic and environmental conditions (Ammann et al. 2013; Bignon 2008; Caspar and De Bie 1996; Cattelain 1997; Duches et al. 2017; Naudinot et al. 2017; Nielsen 2013; Pelegrin 2000; Plisson 2005; Valentin 2008). With regard to the Late Epigravettian of northeastern Italy, technological analyses testify a general simplification of the lithic production systems between 17,000 and 11,600 years cal BP, with a shift in technical investment from shaping on the core to a subsequent shaping on the derived flake blank (Duches et al. 2017; Montoya 2008; Montoya and Peresani 2005). In this framework, the persistence of standardized lithic backed tools used as projectile implements suggest the need for easily repairable projectiles through the substitution of functional elements (Duches et al. 2017). Since the interface between shaft and point represents the fundamental fulcrum of the projectile (Bergman et al. 1988; Carrère 1990; Friis-Hansen 1990; Kooi and Bergman 1997; Pétillon et al. 2011), thus a correct fit of the armature will profoundly condition its capability to perforate and wound: a discrepancy between shaft diameter and implements’ width could cause indeed the rebound of the projectile (hilt effect sensu Guthrie 1983; Pétillon et al. 2011). Shape and dimensions of Late Epigravettian backed pieces (Duches et al. 2017) suggest a shaft diameter not wider than 0.8 cm, excluding it from the range documented in archeology and ethnography for javelins or spear-thrower darts (Cattelain 1997; Churchill 1993; Cundy 1989; Davidson 1934; Ellis 1997; Hughes 1998; Shott 1997; Thomas 1978). Consistent data are also provided by Late Epigravettian antler and bone projectile points: complete items from Dalmeri and Tagliente rock shelter show a maximum width rarely beyond 1 cm in front of a high variable length (Bertola et al. 2007; Cristiani 2008; Gurioli 2008). Bone point from Cornafessa rock shelter (Fig. 14) confirms this tendency, showing a width corresponding to the expected range for arrow tips on the basis of ethnographic (Cattelain 1997, 2004; Churchill 1993; Ellis 1997; Shott 1997; Thomas 1978; Witthoft 1968), experimental (Caspar and De Bie 1996; Cattelain and Perpére 1993; Geneste and Plisson 1986, 1990; O’Farrell 2004; Soriano 1998), and paleo-ballistic data (Bergman et al. 1988; Carrère 1990; Friis-Hansen 1990; Kooi and Bergman 1997). Using as reference 150 complete backed points from Dalmeri rock shelter (raw data and statistical values in Duches et al. 2017), TCSA (tip cross-sectional area, after Hughes 1998) values are largely lower than the expected ones for arrowheads on the base of ethnographic data (Fig. 15); considering the weights, ranging between 0.05 and 1.50 g (Duches et al. 2017), Late Epigravettian projectiles fall into low-power/high-velocity projectiles which are consistent with the bow and arrow delivery system (Hughes 1998; Pétillon et al. 2011; Serwatka 2018). Lastly, spear-throwers are completely absent from Late Epigravettian archeological record.

Bone point from Cornafessa rock shelter. a: intentional breakage of the base and hafting traces on the inner surface; b: longitudinal striations and hafting traces on the mesial part; c: ochre residues and root etchings on the base; d: close-up of scraping marks related to fabrication; e: faceting of the profile of the distal part; f: longitudinal striations and rounding of the tip

Boxplot showing the comparison between TCSA values of Late Epigravettian backed points, ethnographic arrowheads, and ethnographic dart tips. BP: Late Epigravettian backed points from Dalmeri rock shelter (n = 150); A: ethnographic arrowheads (n = 118 after Thomas 1978); D: ethnographic dart tips (n = 40 after Thomas 1978 and Shott 1997)
As suggested by the location of the Cornafessa injury and the projectile’s entrance point, the arrow probably hit the bear from behind with an oblique trajectory headed toward its vital organs. Just like butchery marks, bone injuries might be unintentional and undesired. Hitting osseous tissues may increasingly damage the arrow and compromise its penetration ability. As such, we can suppose that this shot erroneously hit the rib but could anyway cause the bleeding to death of the animal. Some authors even suggest that a more “dorsal” or “lateral” angle of the shot could help the projectile’s penetration of the ribcage in the case of medium or large animals (Frison 1998; Pöllath et al. 2017).
Archeological evidence of bear hunting
It is nowadays generally recognized that the frequent association of bear and cultural remains in Paleolithic sites does not necessarily demonstrate an interaction between the two species (Auguste 2003; Bona et al. 2007; Cvetković and Dimitrijević 2014; Fourvel et al. 2014; Gargett 1996; Stiner 1998; Torres et al. 2007). The sharing of ecological and ethological characteristics led in fact bears and human to live in the same habitats and shelters, cohabiting in a good part of Europe all through the Pleistocene (Arribas and Palmqvist 1999; Brugal and Fosse 2004; Pacher 2003; Quilès 2004; Stiner 1999; Wolverton 2001). The taphonomic history of bear faunal assemblages is therefore the most significant aspect in investigating the interaction between ursids and Paleolithic human groups. If the presence of cut marks and percussion marks confirms the anthropic exploitation of this animal in multiple periods and cultural contexts (Abrams et al. 2014; Arilla et al. 2014; Auguste 2003; Binford 2002; Brugal and Fosse 2004; Enloe 2012; Fosse et al. 2002; Germonpré and Hämäläinen 2007; McLaren et al. 2005; Miracle 2007; Pitulko and Kasparov 1996; Romandini and Nannini 2012; Romandini et al. 2018; Stiner 1994; Tillet and Binford 2002; Turk 2014; Wojtal et al. 2015), the information concerning the modality of bear procurement is limited.
Evidence from different Mousterian sites suggests intensive exploitation of both cave bear and brown bear through targeted visits at watersides or in caves used for lair. The finding of consistent bear bone accumulations associated with a high rate of anthropic marks and a prime adult dominated spectrum supports in fact the hypothesis of active and directed hunting at Biache-Saint-Vaast in France (Auguste 1992, 1995, 2003), Taubach in Germany (Bratlund 1999; Gaudzinski-Windheuser and Roebroeks 2011), Krapina in Croatia (Miracle 2007), and Rio Secco Cave in Italy (Romandini et al. 2018).
Regarding the Upper Paleolithic, there is scarce information from Aurignacian contexts with the notable exception of the Slovenian site of Potočka zijalka. This cave, frequented by humans during the Early Upper Paleolithic, has yielded several osseous projectile points mostly fabricated from ursid bones (Hofreiter and Pacher 2004; Jéquier 2016), associated to a small Protoaurignacian lithic assemblage (Moreau et al. 2015). The zooarcheological record was mainly represented by ursids (in particular Ursus spelaeus, sensu lato), but its accumulation was in most cases natural (Pacher 2001; Pacher and Marinelli 2004; Rabeder et al. 2004). Nevertheless, the identification of a cave bear shoulder blade with a pathology indicating an artificial traumatic origin is noteworthy (Withalm 2004). Wound size and shape led the researcher to infer that the injury was caused by a bone peak, very similar to those found in the Aurignacian layers. Moreover, the position and direction of the channel suggests a perforation from behind with a down-to-upside inclination. Finally, the clear evidence of healing attests to hunting failure (Withalm 2004).
Human exploitation of both cave bear (U. spelaeus, sensu lato) and brown bear is well documented in Gravettian sites of Central Europe (Münzel and Conard 2004; Wojtal et al. 2015). Even though in the Gravettian layers of Hohle Fels Cave (Swabian Jura) and Deszczowa Cave (Krakowsko-Częstochowska Upland), the majority of the accumulated bear remains is the result of natural deaths during hibernation, butchering marks are present on all skeletal elements and document all the phases of the carcasses’ dismemberment (Münzel et al. 2001; Münzel and Conard 2004). Thanks to the presence of fetal horse bones and cut marks on juvenile (about 1 year old) cave bear remains, Münzel (2004) could demonstrate winter occupation of Hohle Fels by Paleolithic hunters and thus confirmed vulnerability of the bears during hibernation. Some exceptional hunting evidence is represented by a projectile impact mark recorded on the lateral process of a cave bear thoracic vertebra dated to 27,830 ± 150–140 uncal BP (Münzel 2004; Münzel and Conard 2004; Münzel et al. 2001). The puncture, characterized by an embedded flint, confirms the active predation of cave bears and suggests a winter hunting during hibernation on the basis of the lesion’s location and shooting angle (Münzel and Conard 2004). The authors also proposed a spear or lance with flint tip as hunting weapon. Finally, the success of the hunt is demonstrated by several cut marks recorded on the spinal process of the same vertebra (Münzel and Conard 2004).
Conspicuous bear bone assemblages also come from Early Epigravettian sites located on the Berici Hills in northeastern Italy, which testify to both bear hibernation and human occupations. Cave bears frequented this area during the Last Glacial Maximum and Epigravettian hunters repeatedly visited the Covolo Fortificato di Trene and the Buso Doppio del Broion Cave for the exploitation of this hunting resource. Taphonomic analysis revealed in fact several anthropic traces on cave bear remains (Romandini and Nannini 2012) stratigraphically associated with shouldered and backed points, some of which characterized by diagnostic impact traces (Broglio et al. 2009).
The Late Epigravettian evidence from the southern slope of the Alps are on the contrary very scanty, testifying the occasional exploitation of U. arctos in faunal assemblages generally dominated by medium-sized ungulates (Bertola et al. 2007; Fiore and Tagliacozzo 2005). In Tagliente (Rocci Ris et al. 2005), Biarzo (Bertolini et al. 2016) and Dalmeri (Fiore e Tagliacozzo 2008) rock shelters, all located in northeastern Italy, taphonomic evidence is mostly related to skinning actions and, rarely, to defleshing and disarticulation. From Dalmeri rock shelter, multiple cut marks on third phalanges also attest to the deliberate removal of bear claws (Fiore and Tagliacozzo 2008).
The only direct information about Late Glacial bear hunting comes from the alpine site of Grotte du Bichon (La Chaux-de-Fonds, Swisse). In this cave, a unique evidence of hunting accident is preserved: a female brown bear skeleton injured by a projectile impact mark was found covering the almost complete skeleton of a 20-year-old man. The archeological interpretation supposes that a female bear was driven out from her den while hibernating with two cubs, wounded, and successively chased in the deepest part of the cave by a hunter who found in turn his death, being injured by the animal (Morel 1998). The abundance of charcoals suggests that the bear was smoked out of the cave and then attacked at the entrance by one or more hunters. The animal displays a flint fragment embedded in a cervical vertebra but it is not clear whether bow and arrow or a spear-thrower were used to inflict the hunting lesion. Since the location of the injury supports the hypothesis of a shot against the animal when it charged standing upon its haunches (Morel 1998), both weapon systems could have been used.
Ethnographical comparison on bear hunting strategies
Ethnographic data show that bear hunting was a pan-boreal practice across the northern hemisphere (McLaren et al. 2005). The synthesis provided by Hallowell (1926), focusing on bear hunting strategies traditionally used from northern Eurasia to North America, offers relevant clues for archeological interpretations. The first noteworthy aspect is the evidence of different annual favorable moments in bear approachability, which considerably affects weapons and hunting techniques employed. Certain predictable aspects of seasonal bear behavior are in fact strategical for the procurement of this prey and hunting success.
Hunting during the hibernating season, between winter and early spring, is well documented in ethnography and connected to bear vulnerability. The animal’s habit of using the same lair for several consecutive years (Kurten 1976) and the hunters’ custom of tracking bear during late fall in order to find its den guarantee a potential mid-winter source of food, fur, grease, and bone (Hallowell 1926). The animal is never killed in the den but the hunters generally lure it out using smoke or dogs and then dispatch it through spear or axe (Hallowell 1926). Spears could be either wielded by the hunter or set in a stationary position at which the bear was provoked or tricked into charging (McLaren et al. 2005). The ethnographical documentation of the use of bow and arrow in such hunting situation is on the contrary very rare (Hallowell 1926). This kind of weapon is more frequently employed in the open during warmer seasons, as documented for some Paleo-Siberian peoples (Ainu: Batchelor 1901; Koryak: Jochelson 1905-1908; Gilyak of Sagyhalin: Labbé 1903) and natives of the North Pacific Coast (Algonkian: Denys 1908; Tlingit: Krause 1885). It is also interesting that during the bear festival among the Ainu, the animal is firstly shot at with blunt arrows before being ceremonially slaughtered (Batchelor 1901). In closer hand-to-hand combats, bow and arrow are frequently replaced by spear or lance engaging a very dangerous venture (de Laguna 1972; Hallowell 1926).
The use of traps and snares is frequently documented among ethnographic people and can occur all year round. The construction of traps is strictly related to the bears’ habit of “step-marking,” i.e., their tendency to step over their own footprints while walking their habitual trails (Holzworth 1930; Hummel and Pettigrew 1991). Such behavior facilitates bear trapping and considerably decreases hunting risk. Bear deadfall traps were used among numerous peoples of the Northwest Coast of North America as well as along the Siberian Coast (Emmons 1991; Hallowell 1926; Harrison 1925; Swanton 1905). Butchering activities always occurred in the vicinity of trapping localities that are, in turn, always far away from the residential camps (McLaren et al. 2005).
Regardless of the modality of predation, ethnographic data testify carcass-selective processes and differential transport from the initial kill site: when the frequencies of recovered skeletal remains in sites other than butchering places are compared, indeed, bear trunk elements are always underrepresented. This is interpreted as the habit of leaving the spinal column and the ribcage in the dispatching area (Binford 1983, 2002). Archeological data from two early Holocene residential sites with abundance of bear remains strengthen this assumption: the Kilgii Gwaay site on the American Northwest Coast (McLaren et al. 2005) and the Zhokhov Island site in Siberia (Pitulko and Kasparov 1996) show a similar pattern of bear skeletal element abundance, marked by the strong underrepresentation of vertebral elements, ribs, and hind limbs.
Conclusion
This work represents the first archeological application of a diagnostic method for PIMs identification, based on statistic comparison of morphometrical parameters acquired through 3D digital microscopy. The recognition of the Cornafessa drag mark is particularly relevant because it confirms the method’s validity on the most challenging PIM category. If indeed puncture marks are frequently recognized on zooarcheological material because of the presence of stone embedded (Gaudzinski-Windheuser 2016; O’Driscoll and Thompson 2018; Pöllath et al. 2017), the distinction of drags from cut marks and carnivore tooth scores is much more complicated. Moreover, we proved that different projectiles leave different PIMs and that only a reliable ballistic experimentation could provide the diagnostic elements useful in drag identification (Duches et al. 2016). The presence of small lithic fragments embedded in the Cornafessa specimen enable us to add some methodological considerations concerning the relevance of this feature in drag identification and the potential significance of profile width and depth variations for the reconstruction of projectile design. In this regard, the morphometrical features of Cornafessa specimen compared to the experimental ones suggest that the lesion was caused by a lithic composite projectile, probably characterized by a backed point associated with at least one backed (bi)truncated bladelet.
The identification of this drag on a U. arctos rib enriches the current debate about Pleistocene bear hunting, offering new clues about the strategies employed by Epigravettian groups during the Late Glacial. Several remains from Late Epigravettian sites in northeastern Italy testify to bear processing for the acquisition of fur, meat, and bones (Bertolini et al. 2016; Fiore and Tagliacozzo 2008; Rocci Ris et al. 2005) but none provides clear evidence of bear hunting. If evidence of skin removal suggests a primary access to the carcass for these sites, the reconstruction of procurement season and hunting strategy is more challenging. Seasonal information is not available for the Cornafessa rock shelter yet, but the PIM analysis confirms a direct predation on this bear.
Going beyond the limit of the Late Glacial period, direct information about bear hunting strategies remains sporadic. The drag from Cornafessa rock shelter represents the fourth hunting lesion recognized on bear bones for all the Pleistocene (Table 3). The other evidence from Potočka zijalka, Hohle Fels, and Grotte du Bichon (Morel 1998; Münzel and Conard 2004; Withalm 2004) consist of three impact punctures, interpreted as hunting predation during winter taking advantage of bears vulnerability in hibernation. Given the different location of the lesions, the authors hypothesize a predation toward a bear lying in hibernation for Hohle Fels (Münzel and Conard 2004) and an assault against a bear charging in a standing position for both Potočka zijalka and Grotte du Bichon (Morel 1998; Withalm 2004). Regarding the weapon employed, a spear or lance with osseous or lithic tips are believed for Potočka zijalka and Hohle Fels, while both bow and arrow or spear-thrower are considered for Grotte du Bichon. Nevertheless, ethnographic data suggest that spear or lance are widely preferred by hunters when attacking a bear at the cave entrance after the interruption of its hibernation period.
Since Cornafessa rock shelter is not suitable as hibernating den, this new evidence represents a dissimilar attestation of bear hunting strategies, testifying to the predation of a bear in a place different from lair caves. Besides, the PIM location on a rib and the presence of two vertebrae within the identified bear bones possibly suggest the transportation into the site of the whole carcass, an unusual behavior for such a big prey. In this regard, ethnographic and archeological data frequently attest to bear carcass differential transport from the initial kill site to residential site, on the basis of comparative frequencies of recovered skeletal remains marked by a strong underrepresentation of bear trunk elements (Binford 1983, 2002; McLaren et al. 2005; Pitulko and Kasparov 1996).
Finally, since the other few direct evidence of bear hunting suggests a different scenario, i.e., den hunting through spears, this rib represents the first archeological proof of bear predation using bow and arrow.
References
Abrams G, Bello SM, Di Modica K, Pirson S, Bonjean D (2014) When Neanderthals used cave bear (Ursus spelaeus) remains: bone retouchers from unit 5 of Scladina Cave (Belgium). Quat Int 326:274–287
Agogué O, Dalmeri G (2005) Lamelles à dos tronquées et pointes à dos: deux modèles d’armatures dans l’Epigravettien récent du Riparo Dalmeri (Grigno, Trentin, Italie). PreistAlp 41:231–243
Aimar A, Alciati G, Broglio A, Castelletti L, Cattani L, D’Amico C, Giacobini G, Maspero A, Peresani M (1992) Les Abris Villabruna dans la Vallée du Cismon. PreistAlp 28:227–254
Ammann B, van Raden UJ, Schwander J, Eicher U, Gilli A, Bernasconi SM, van Leeuwen JFN, Lischke H, Brooks SJ, Heiri O, Novàkovà K, van Hardenbroek M, von Grafenstein U, Belmecheri S, van der Knaap WO, Magny M, Eugster W, Colombaroli D, Nielsen E, Tinner W, Wright HE (2013) Responses to rapid warming at termination 1a at Gerzensee (Central Europe): primary succession, albedo, soils, lake development, and ecological interactions. Palaeogeogr Palaeoclimatol Palaeoecol 391(Part B):111–131
Arilla M, Rosell J, Blasco R, Dominguez-Rodrigo M, Pickering TR (2014) The “bear” essential: actualistic research on Ursus arctos arctos in the Spanish pyrenees and its implications for paleontology and archaeology. PLoS One 9:1–25
Arriaza MC, Yravedra J, Domínguez-Rodrigo M, Mate-González MA, García Vargas E, Palomeque-González JF, Aramendi J, González-Aguilera D, Baquedano E (2017) On applications of micro-photogrammetry and geometric morphometrics to studies of tooth mark morphology: the modern Olduvai Carnivore Site (Tanzania). Palaeogeogr Palaeoclimatol Palaeoecol 488:103–112
Arribas A, Palmqvist P (1999) On the ecological connection between sabre-tooths and hominids: faunal dispersal events in the lower Pleistocene and a review of the evidence for the first human arrival in Europe. J Archaeol Sci 26(5):571–585
Arrighi S, Bazzanella M, Boschin F, Wierer U (2016) How to make and use a bone “spatula”. An experimental program based on the Mesolithic osseous assemblage of Galgenbühel/Dos de la Forca (Salurn/Salorno, BZ, Italy). Quat Int 423:143–165
Auguste P (1992) Ėtude archéozoologique des grands mammifères du site pléistocène moyen de Biache-Saint-Vaast (Pas-de-Calais, France): apports biostratigraphiques et palethnographiques. Anthropologie 96:49–70
Auguste P (1995) Chasse et charognage au Paléolithique moyen: L’apport du gisement de Biache-Saint-Vaast (Pas-de-Calais). Bull Soc Préhist Fr 92:155–167
Auguste P (2003) La chasse à l’ours au Paléolithique moyen: mythes, réalités et état de la question. In: Patou Mathis M, Bocherens H (eds) Le rôle de l’Environnement dans les Comportements des Chasseurs Cueilleurs Préhistoriques, vol. 1105, BAR, pp 135–142
Avanzini M, Masetti D (2007) Oolite di San Vigilio. In: Cita Sironi MB, Abbate E, Balini M, Conti MA, Falorni P, Germani D, Groppelli G, Manetti P, Petti FM (eds) Carta Geologica d’Italia – 1:50.000, Catalogo delle Formazioni, Unità tradizionali, APAT, Dipartimento Difesa del Suolo, Servizio Geologico d’Italia, volume V, fascicolo VI, pp 89–97
Bachechi L, Fabbri P-F, Mallegni F (1997) An arrow-caused lesion in a Late Upper Palaeolithic human pelvis. Curr Anthropol 38(I):135–140
Batchelor J (1901) The Ainu and their folk-lore. London
Beckhoff K (1968) Eignung und Verwendung einheimischer Holzarten für prähistorische Pfeilschäfte. Die Kunde 19:85–101
Behrensmeyer AK (1978) Taphonomic and ecologic information from bone weathering. Paleobiology 4(2):150–162
Bello SM, Soligo C (2008) A new method for the quantitative analysis of cut mark micromorphology. J Archaeol Sci 35:1542–1552
Bello SM, Parfitt SA, Stringer C (2009) Quantitative micromorphological analyses of cut marks produced by ancient and modern handaxes. J Archaeol Sci 36:1869–1880
Bello SM, Parfitt SA, De Groote I, Kennaway G (2013) Investigating experimental knapping damage on an antler hammer: a pilot-study using high-resolution imaging and analytical techniques. J Archaeol Sci 40(12):4528–4537
Bergman CA, McEwen E, Miller R (1988) Experimental archery: determination of projectile velocities and comparison of bow performances. Antiquity 62:658–670
Bertola S (2016) Southern Alpine (Trento Plateau) and Northern Apennine flints: ages, distribution and petrography. In: Tomasso A, Binder D, Martino G, Porraz G, Simon P, Naudinot N (eds) Ressources lithiques, productions et transferts entre Alpes et Méditerranée. Actes de la Séance de la Société Préhistorique Française 5, Paris, pp 55–75
Bertola S, Broglio A, Cassoli P, Cilli C, Cusinato A, Dalmeri G, De Stefani M, Fiore I, Fontana F, Giacobini G, Guerreschi A, Gurioli F, Lemorini C, Liagre J, Malerba G, Montoya C, Peresani M, Rocci Ris A, Rossetti P, Tagliacozzo A, Ziggiotti S (2007) L’Epigravettiano recente nell’area prealpina e alpina orientale. In: Martini F (ed) L’Italia tra 15.000 e 10.000 anni fa, Cosmopolitismo e regionalità nel Tardoglaciale. Studi di Archeologia Preistorica 5, Millenni, Museo Fiorentino di Preistoria “Paolo Graziosi”, pp 39–94
Bertolini M, Cristiani E, Modolo M, Visentini P, Romandini M (2016) Late Epigravettian and Mesolithic foragers of the eastern Alpine region: animal exploitation and ornamental strategies at Riparo Biarzo (Northern Italy). Quat Int 423:73–91
Bignon O (2008) Chasser les chevaux à la fin du Paléolitique dans le Bassinparisien: stratégie cynégétique et mode de vie au Magdalénien et à l'Azilienancien, Brit Archaeol Rep 1747, Oxford
Binford LR (1981) Bones, ancient men and modern myths. Academic Press, New York
Binford LR (1983) In pursuit of the past. Thames and Hudson, London
Binford LR (2002) L’interaction ethnograpique homme-ours et le gisements européens d’ours des cavernes. In Tillet T, Binford LR (eds) L’ours et l'Homme. Symposium d'Auberives-en-Royans-Isère- France E.R.A.U.L. 100, pp 141–155
Blumenschine RJ, Selvaggio MM (1988) Percussion marks on bone surfaces as a new diagnostic of hominid behavior. Nature 333:763–765
Boëda E, Geneste JM, Griggo C (1999) A Levallois point embedded in the vertebra of a wild ass (Equus africanus): hafting, projectiles and Mousterian hunting weapons. Antiquity 73:394–402
Bona F, Peresani M, Tintori A (2007) Indices de fréquentation humaine dans les Grottes à ours au Paléolithique moyen final. L’exemple de la Caverna Generosa dans les Pealps lombardes, Italie. Anthropologie 111:290–320
Bordes F (1974) Percuteur en bois de renne du Solutréen supérieur de Laugerie-Haute Ouest. In: Camps Fabrer H (ed) Premier Colloque International sur l'Industrie de l'Os dans la Préhistoire Abbaye de Sénanque, avril 1974. Editions de l'Université de Provence, Aix-en-Provence, pp 97–100
Boschin F, Crezzini J (2012) Morphometrical analysis on cut marks using a 3D digital microscope. Int J Osteoarchaeol 22:549–562
Boschin F, Bernardini F, Zanolli C, Tuniz C (2015) MicroCT imaging of red fox talus: a non-invasive approach to evaluate age at death. Archaeometry 57(S1):194–211
Bosellini A, Broglio A, Loriga C, Busetto C (1978) I bacini cretacei del Trentino. Riv Ital Paleontol Stratigr 84(4):897–946
Bouxsein M, Boyd SK, Christiansen BA, Guldberg RE, Jepsen KJ, Müller R (2010) Guidelines for assessment of bone microstructure in rodents using micro-computed tomography. J Bone Miner Res 25:1468–1486
Bratlund B (1991) A study of hunting lesions containing flint fragments on reindeer bones at Stellmoor, Schleswig-Holstein, Germany. In: Barton N, Roberts AJ, Roe DA (eds) The Late Glacial in North-west Europe. CBA Research Report 77, pp 193–207
Bratlund B (1996) Hunting strategies in the Late Glacial of northern Europe: a survey of the faunal evidence. J World Prehist 10:1–48
Bratlund B (1999) Taubach revisited. Jahrbuch des Römisch-Germanischen Zentralmuseums Mainz 46:67–174
Broglio A, Bertola S, De Stefani M, Gurioli F (2009) The shouldered points of the Early Epigravettian of the Berici Hills (Venetian Region – North of Italy). Materials, blanks, typology, exploitation. In: Burdukiewicz JM, Cyrek K, Dyczek P, Szymczak K (eds) Understanding the past. Papers offered to Stefan K. Kozlowski. Centre for Research on the Antiquity of Southeastern Europe, University of Warsaw, Warszawa, pp 59–68
Brugal JP, Fosse P (2004) Carnivores et Hommes au Quaternaire en Europe de l'Ouest. Rev Paléobiol 23:575–595
Capaldo SD, Blumenschine RJ (1994) A quantitative diagnosis of notches made by hammerstone percussion and carnivore gnawing on bovid long bones. Am Antiq 59:724–748
Carrère E (1990) Contribution de la balistique au perfectionnement des études techno-fonctionnelles des pointes de projectiles préhistoriques. Paléo 2:167–176
Caspar JP, De Bie M (1996) Preparing for the hunt in the Late Paleolithic camp at Rekem, Belgium. J Field Archaeol 23:437–460
Castel J-C (2008) Identification des impacts de projectiles sur le squelette des grands ongulés. Ann Paléontol 94:103–118
Cattelain P, Perpère M (1993) Tir expérimental des sagaies et de flèches emmanchées de pointes de la Gravette. Archéo-Situla 17-20:5–28
Cattelain P (1997) Hunting during the Upper Paleolithic: bow, spearthrower, or both. In: Knecht H (ed) Projectile technology. Plenum Press, New York, pp 213–240
Churchill SE (1993) Weapon technology, prey size selection and hunting methods in modern huntergatherers: implications for hunting in the Palaeolithic and Mesolithic. In: Peterkin GL, Bricker HM, Mellars E (ed) Hunting and animal exploitation in the Later Palaeolithic and Mesolithic of Eurasia, Archeological Papers of the American Anthropological Association 4, Washington D.C., pp 11–24
Churchill SE, Franciscus RG, McKean-Peraza HA, Daniel JA, Warren BR (2009) Shanidar 3 Neandertal rib puncture wound and Paleolithic weaponry. J Hum Evol 57:163–178
Coleman MN, Colbert MW (2007) Technical note: CT thresholding protocols for taking measurements on three-dimensional models. Am J Phys Anthropol 133:723–725
Courtenay LA, Yravedra J, Mate-González MÁ, Aramendi J, González-Aguilera D (2017) 3D analysis of cut marks using a new geometric morphometric methodological approach. Archaeol Anthropol Sci. https://doi.org/10.1007/s12520-017-0554-x
Crezzini J, Boschin F, Boscato P, Wierer U (2014) Wild cats and cut marks: exploitation of Felis silvestris in the Mesolithic of Galgenbühel/Dos de la Forca (South Tyrol, Italy). Quat Int 330:52–60
Cristiani E (2008) Analisi funzionale dei manufatti in materia dura animale del Riparo Dalmeri (Altopiano della Marcesina, Trento). PreistAlp 43:259–287
Cundy BJ (1989) Formal variation in Australian spear and spearthrower technology. BAR International Series 546 (Oxford, England)
Cvetković NJ, Dimitrijević VN (2014) Cave bears (Carnivora, Ursidae) from the Middle and Late Pleistocene of Serbia: a revision. Quat Int 339-340:197–208
Davidson DS (1934) Australian spear traits and their derivations. J Polyn Soc 43(41–72):143–162
Denys N (1908) The description and natural history of the coasts of North America (Acadia). Edition of the Champlain Society, Toronto
Domínguez-Rodrigo M, Piqueras A (2003) The use of tooth pits to identify carnivore taxa in tooth-marked archeofaunas and their relevance to reconstruct hominid carcass processing behaviours. J Archaeol Sci 30:1385–1391
Domínguez-Rodrigo M, de Juana S, Galán AB, Rodríguez M (2009) A new protocol to differentiate trampling marks from butchery cut marks. J Archaeol Sci 36:2643–2654
Duches R, Nannini N, Romandini M, Boschin F, Crezzini J, Peresani M (2016) Identification of Late Epigravettian hunting injuries: descriptive and 3D analysis of experimental projectile impact marks on bone. J Archaeol Sci 66:88–102
Duches R, Peresani M, Pasetti P (2017) Success of a flexible behavior. Considerations on the manufacture of Late Epigravettian lithic projectile implements according to experimental tests, Archaeological and Anthropological Sciences (March 2017). doi:https://doi.org/10.1007/s12520-017-0473-x
Ellis CJ (1997) Factors influencing the use of stone projectile tips. In: Knecht H (ed) Projectile technology. Plenum Press, New York, pp 37–75
Emmons GT (1991) The Tlingit Indians. University of Washington Press, Seattle
Enloe JG (2012) Neanderthals, bears and hyenas, oh my! Competition for exclusive use of space. Journal of Taphonomy 10(3–4):185–195
Fajardo RJ, Ryan TM, Kappelman J (2002) Assessing the accuracy of high resolution X-ray computed tomography of primate trabecular bone by comparisons with histological sections. Am J Phys Anthropol 118:1–10
Fernàndez-Crespo T (2016) An arrowhead injury in a Late Neolithic/Early Chalcolithic human cuneiform from the Rockshelter of La Peña de Marañón (Navarre, Spain). Int J Osteoarchaeol 26:1024–1033
Fernàndez-Crespo T (2017) New evidence of early chalcolithic interpersonal violence in the Middle Ebro Valley (Spain): two arrowhead injuries from the Swallet of Las Yurdinas II. Int J Osteoarchaeol 27:76–85
Fernandez-Jalvo Y, Andrews P (2016) Atlas of taphonomic identifications. 1001 + images of fossil and recent mammal bone modification. Vertebrate Paleobiology and Paleoanthropology Series. Springer, Berlin
Fiore I, Tagliacozzo A (2005) Lo sfruttamento delle risorse animali nei siti di altura e di fondovalle nel Tardiglaciale dell’Italia nord-orientale. In: Malerba G, Visentini P (eds) Atti del 4° Convegno Nazionale di Archeozoologia, Quaderni del Museo Archeologico del Friuli Occidentale 6, pp 97–109
Fiore I, Tagliacozzo A (2008) Oltre lo stambecco: gli altri mammiferi della struttura abitativa dell’US 26c a Riparo Dalmeri (Trento). PreistAlp 43:209–236
Fischer A (1985) Hunting with flint-tipped arrows: results and experiences from practical experiments. In: Bonsall C (ed) The Mesolithic in Europe. Colloque UISPP Edinburgh 1985. John Donald Publishers Ltd, Edinburgh
Fisher WJ (1995) Bone surface modifications in zooarchaeology. J Archaeol Method Theory 2:7–68
Fosse P, Morel P, Brugal J-P (2002) Taphonomie et éthologie des ursidés pléistocènes. In: Tillet T, Binford LR (eds) L’ours et l'Homme. Symposium d'Auberives-en-Royans-Isère- France E.R.A.U.L. 100, pp 79–100
Fourvel JB, Fosse P, Brugal JP, Cregut-Bonnoure E, Slimak L, Tournepiche JF (2014) Characterization of bear remains consumption by Pleistocene large carnivores (Felidae, Hyaenidae, Canidae). Quat Int 339-340:232–244
Friis-Hansen J (1990) Mesolithic cutting arrows used in the hunting used in the hunting of large game. Antiquity 64:494–504
Frison GC (1998) Paleoindian large mammal hunters on the plains of North America. Proc Natl Acad Sci 95(24):14576–14583
Gargett RH (1996) Cave bears and modern human origins: the spatial taphonomy of Pod Hradem Cave, Czech Republic. University Press of America, Lanham
Gaudzinski-Windheuser S (2016) Hunting lesions in Pleistocene and Early Holocene European bone assemblages and their implications for our knowledge on the use and timing of lithic projecyile technology. In: Iovita R, Sano K (eds) Multidisciplinary approaches to the study of Stone Age weaponry. Springer Science+Business Media, Dordrech, pp 77–100
Gaudzinski-Windheuser S, Roebroeks W (2011) On Neanderthal subsistence in last interglacial forested environments in Northern Europe. In: Conard NJ, Richter J (eds) Neanderthal lifeways, subsistence and technology: one hundred fifty years of Neanderthal study. Springer, Dordrecht, pp 61–71
Geneste JM, Plisson H (1986) Le Solutréen de la Grotte Combe-Saunière, première approche palethnologique. Gallia Préhistoire 29(1):9–27
Geneste JM, Plisson H (1990) Technologie fonctionnelle des pointes à cran solutréennes: l'apport des nouvelles données de la grotte de Combe Saunière (Dordogne). In: Kozlowski JK (ed) Feuilles de pierre. Les industries à pointes foliacées du Paléolithique supérieur européen, Etudes et Recherches archéologiques de l'Université de Liège 42, pp 293–320
Germonpré M, Hämäläinen R (2007) Fossil bear bones in the Belgian Upper Paleolithic: the possibility of a proto bear-ceremonialism. Arct Anthropol 44:1–30
Gurioli F (2008) Analisi tecnologica dei manufatti in materia dura animale dell’Epigravettiano recente di Riparo Dalmeri (Altopiano della Marcesina, Trento). PreistAlp 43:237–258
Guthrie RD (1983) Osseous projectile points: biological considerations affecting raw material selection and design among palaeolithic and paleoindian people. In: Clutton-Brock J, Grigson C (eds) Animals and archaeology I: hunters and their prey, BAR International Series 165, British Archaeological Reports, Oxford, pp 273–294
Hallowell AI (1926) Bear ceremonialism in the Northern Hemisphere. Am Anthropol 28:1–175
Hammer Ø, Harper DAT, Ryan PD (2001) PAST: Paleontological Statistics Software Package for Education and Data Analysis. Paleontologia Electronica 4(1):1-9Harrison C (1925) Ancient Warriors of the North Pacific: The Haidas, their Laws, Customs and Legends, with some Historical Account of the Queen Charlotte Islands. H. F. & G. Witherby, London
Harrison C (1925) Ancient warriors of the North Pacific: the Haidas, their laws, customs and legends, with some historical account of the Queen Charlotte Islands. H. F. & G. Witherby, London
Hofreiter M, Pacher M (2004) Using ancient DNA to Elucidate raw material origin of bone points from Potoćka zijalka (Slovenia): preliminary results. In: Pacher M, Pohar V, Rabeder G (eds) Potoćka zijalka, palaeontological and archaeological results of the campaigns 1997–2000. Oesterrichische Akademie der Wissenschaften, Wien, pp 201–210
Holzworth JM (1930) The wild grizzlies of Alaska. G. P. Putnam’s Sons, New York
Hughes S (1998) Getting to the point: evolutionary change in prehistoric weaponry. J Archaeol Method Theory 5:345–408
Hummel M, Pettigrew S (1991) Wild hunters: predators in peril. Key Porter Books Limited, Toronto
Jéquier C (2016) The incised bone points from the Early Aurignacian of Potoćka zijalka (Slovenia), hafting system or ornament? Quat Int 403:51–56
Jochelson W (1905-1908) The Koryak. Jesup, vol. 6, New York
Kooi BW, Bergman CA (1997) Archery using mathematical modeling. Antiquity 71:124–134
Krause A (1885) Die Tlinkit Indianer. H. Costenoble, Jena
Kurten B (1976) The cave bear story. Columbia University Press, New York
Labbé P (1903) Un Bagne Russe, l'Isle de Sakhaline. Paris
de Laguna F (1972) Under Mount Saint Elias: the history and culture of the Yakitat Tlingit. Smithsonian Contributions to Anthropology, Volume 7. Smithsonian Institution, Washington D.C.
Leduc C (2012) New Mesolithic hunting evidence from bone injuries at Danish Maglemosian sites: Lundby Mose and Mullerup (Sjælland). Int J Osteoarchaeol 22:1–16
Lemorini C, Rossetti P (2004) Lo studio delle tracce d’uso dei manufatti litici. In: Peresani M (ed) 12.000 anni fa al Bus de la Lum. Un accampamento paleolitico sull’Altopiano del Cansiglio, Società Naturalisti “Silvia Zenari”, Pordenone, pp 165–172
Lemorini C, Rossetti P, Cusinato A, Dalmeri G, Hrozny Kompatscher NM, Kompastcher K (2006) L’analisi delle tracce d’uso e l’elaborazione spaziale: il riconoscimento di un’area specializzata nel sito epigravettiano di Riparo Dalmeri, livelli 26b e 26c (Trento). PreistAlp 41:171–197
Letourneux C, Pétillon J-M (2008) Hunting lesions caused by osseous projectile points: experimental results and archaeological implications. J Archaeol Sci 35:2849–2862
Litt T, Stebich M (1999) Bio- and chronostratigraphy of the Late Glacial in the Eifel region, Germany. Quat Int 6:5–16
Lyman RL (1994) Vertebrate taphonomy. Cambridge University Press, Cambridge, p 524
Mallye J-P, Thiébaut C, Mourre V, Costamagno S, Claud E, Weisbecker P (2012) The Mousterian bone retouchers of Noisetier Cave: experimentation and identification of marks. J Archaeol Sci 39(4):1131–1142
Maté-González MÁ, Yravedra J, González-Aguilera D, Palomeque-González JF, Domínguez-Rodrigo M (2015) Micro-photogrammetric characterization of cut marks on bones. J Archaeol Sci 62:128–142
Maté-González MÁ, Aramendi J, González-Aguilera D, Yravedra J (2017) Statistical comparison between low-cost methods for 3D characterization of cut-marks on bones. Remote Sens 9(9):873. https://doi.org/10.3390/rs9090873
McLaren D, Wigen RJ, Mackie Q, Fedje DW (2005) Bear hunting at the Pleistocene/Holocene transition on the northern northwest coast of North America. Canadian Zooarchaeology 22:3–29
Milo R (1998) Evidence for hominid predation at Klasies River Mouth, South Africa, and its implications for the behaviour of early modern humans. J Archaeol Sci 25:99–133
Miracle PT (2007) The Krapina Paleolithic site: zooarchaeology, taphonomy and catalog of the faunal remains. Croatian Natural History Museum, Zagreb
Montoya C (2008) Evolution des concepts de productions lithiques et artistiques à l’Epigravettien récent: analyses de collections des Préalpes de la Vénétie et des Préalpes du sud françaises. In: Mussi M (ed) Il Tardiglaciale in Italia. Lavori in corso. Brit Archaeol Rep 1859, pp 43–55
Montoya C, Peresani M (2005) Premiers éléments de diachronie dans l’Epigravettien récent des Préalpes de la Vénétie. In: Bracco JP, Montoya C (eds) D’un monde à l’autre. Les systèmes lithiques pendant le Tardiglaciaire autour de la Méditerranée nord-occidentale, Actes de la Table Ronde, Mémoire de la Societé Préhistorique Française XL, pp 123–138
Moreau L, Odar B, Higham T, Horvat A, Pirkmajer D, Turk P (2015) Reassessing the Aurignacian of Slovenia: techno-economic behaviour and direct dating of osseous projectile points. J Hum Evol 78:158–180
Morel P (1998) La grotte du Bichon (La Chaux-de-Fonds, canton de Neuchâtel, Suisse): un site archéologique singulier, ou lʼhistoire dʼune chasse à lʼours brun il y a 12 ans dans le Jura suisse. In: Cupillard C, Richard A (eds) Les derniers chasseurs-cueilleurs du Massif jurassien et de ses marges. Centre jurassien du Patrimoine, Lons-le-Saunier, pp 88–93
Moretti E, Arrighi S, Boschin F, Crezzini J, Aureli D, Ronchitelli A (2015) Using 3D microscopy to analyze experimental cut marks on animal bones produced with different stone tools. Ethnobiology Letters 6(2):267–275
Münzel SC (2004) Subsistence patterns in the Gravettian of the Ach Valley, a former tributary of the Danube in the Swabian Jura. In: Svoboda J, Sedlàĉkovà L (eds), The Gravettian along the Danube, the Dolni Vestonice studies, vol. 11, pp 71–85
Münzel SC, Conard NJ (2004) Cave bear hunting in the Hohle Fels, a cave site in the Ach Valley, Swabian Jura. Rev Paléobiol 23:877–885
Münzel SC, Langguth K, Conard N, Uerpmann HP (2001) Höhlenbärenjagd auf der Schwäbischen Alb vor 30.000 Jahren. Archäologisches Korresspondenzblatt 31(3):317–328
Naudinot N, Tomasso A, Tozzi C, Peresani M (2014) Changes in mobility patterns as a factor of 14C date density variation in the Late Epigravettian of Northern Italy and southeastern France. J Archaeol Sci 52:578–590
Naudinot N, Tomasso A, Messager E, Finsinger W, Ruffaldi P, Langlais M (2017) Between Atlantic and Mediterranean: changes in technology during the Late Glacial in Western Europe and the climate hypothesis. Quat Int 428(Part B):33–49
Nielsen E (2013) Response of the Late Glacial fauna to climatic change. Palaeogeogr Palaeoclimatol Palaeoecol 391(Part B):99–110
Nikolskiy P, Pitulko V (2013) Evidence from the Yana Paleolithic site, Artic Siberia, yields clues to the riddle of mammoth hunting. J Archaeol Sci 40:4189–4197
Noe-Nygaard N (1974) Mesolithic hunting in Denmark illustrated by bone injuries caused by human weapons. J Archaeol Sci 1:217–248
O’Driscoll CA, Thompson JC (2014) Experimental projectile impact marks on bone: implication for identifying the origins of projectile technology. J Archaeol Sci 49:398–413
O’Driscoll CA, Thompson JC (2018) The origins and early elaboration of projectile technology. Evol Anthropol 27:30–45
O’Farrell M (2004) Les pointes de La Gravette de Corbiac (Dordogne) et considérations sur la chasse au Paléolithique supérieur ancien. In: Bodu P, Costantin C (eds) Approches fonctionnelles en en Préhistoire, (actes du XXVe Congrès de la Sociètè préhistorique française, Nanterre, 24–26 Novembre, 2000). Editions de la Société préhistorique française, Paris, pp 121–138
Olsen SL (1989) On distinguishing natural from cultural damage on archeological antler. J Archaeol Sci 16:125–135
Outram AK (2002) Bone fracture and within-bone nutrients: an experimentally based method for investigating levels of marrow extraction. In: Miracle P, Milner N (eds) Consuming passions and patterns of consumption. McDonald Institute for Archaeological Research, Cambridge, pp 51–64
Oxilia G, Fiorillo F, Boschin F, Boaretto E, Apicella SA, Matteucci C, Panetta D, Pistocchi R, Guerrini F, Margherita C, Andretta M, Sorrentino R, Boschian G, Arrighi S, Dori I, Mancuso G, Crezzini J, Riga A, Serrangeli MC, Vazzana A, Salvadori PA, Vandini M, Tozzi C, Moroni A, Feeney RNM, Willman JC, Moggi-Cecchi J, Benazzi S (2017) The dawn of dentistry in the late upper Paleolithic: an early case of pathological intervention at Riparo Fredian. Am J Phys Anthropol 163:446–461
Pacher M (2001) New excavation campaigns in the upper Pleistocene bear site Potoćka zijalka, Slovenia—state of investigation. Cadernos do Laboratorio Xeolòxico de Laxe 26:301–310
Pacher M (2003) Upper Pleistocene cave assemblages at alpine sites in Austria and adjacent regions. PreistAlp 39:115–127
Pacher M, Marinelli M (2004) Taphonomic analyses of cave bear remains from Potoćka zijalka (Slovenia): spatial distribution analysis. In: Pacher M, Pohar V, Rabeder G (eds) Potoćka zijalka, palaeontological and archaeological results of the campaigns 1997–2000. Oesterrichische Akademie der Wissenschaften, Wien, pp 89–96
Pante MC, Muttart MV, Keevil TL, Blumenschine RJ, Njau JK, Merritt SR (2017) A new high-resolution 3-D quantitative method for identifying bone surface modifications with implications for the Early Stone Age archaeological record. J Hum Evol 102:1–11
Parsons I, Badenhorst S (2004) Analysis of lesions generated by replicated Middle Stone Age lithic points on selected skeletal elements. S Afr J Sci 100:384–387
Pelegrin J (2000) Les techniques de débitage laminaire au Tardiglaciaire: critères de diagnose et quelques réflexions. In: Valentin B, Bodu P, Christensen M (eds) L’Europe centrale et septentrionale au Tardiglaciaire, Mémoires du Musée de Prèhistoire d’Ile-de-France 7, pp 73–86
Péquart M, Péquart S-J, Boule M, Vallois H (1937) Téviec. Station nécropole Mésolithique du Morbihan. Archives de l’Institut de Paléontologie Humaine, Mémoire 18. Masson et Cie. Editeurs, Paris
Peresani M, Duches R, Miolo R, Romandini M, Ziggiotti S (2011) Small specialized hunting sites and their role in Epigravettian subsistence strategies. A case study in Northern Italy. In: Bon F, Costamagno S, Valdeyron N (eds) Hunting camps in prehistory. Current Archaeological Approaches, Proceedings of the International Symposium, May 13-15 2009, University Toulouse II - Le Mirail, P@lethnology, 3, pp 251–266
Pétillon J-M, Letourneux C (2003) Au retour de la chasse. Observations expérimentales concernant les impacts sur le gibier, la récupération et la maintenance des projectiles dans le Magdalénien supérieur d’Isturitz. Préhistoire Anthropologie Méditerranéenne 12:173–188
Pétillon J-M, Letourneux C (2008) Traces d'impacts de projectiles sur le gibier: résultats comparés des tirs expérimentaux à l'arc et au propulseur effectués au Cedarc (Treignes, Belgique) en 2003 et 2004. Ann Paléontol 94:209–225
Pétillon J-M, Bignon O, Bodu P, Cattelain P, Debaut G, Langlais M, Laroulandiae V, Plisson H, Valentin B (2011) Hard core and cutting edge: experimental manufacture and use of Magdalenian composite projectile tip. J Archaeol Sci 38:1266–1283
Pickering TR, Egeland CP (2006) Experimental patterns of hammerstone percussion damage on bones: implications for inferences of carcass processing by humans. J Archaeol Sci 33:459–469
Pitulko V, Kasparov AK (1996) Ancient Arctic hunters: material culture and survival strategy. Arct Anthropol 33:1–36
Pitulko V, Tikhonov AN, Pavlova EY, Nikolskiy PA, Kuper KE, Polozov RN (2016) Early human presence in the Arctic: evidence from 45,000-year-old mammoth remains. Science 351(6270):260–263
Plisson H (2005) Examen tracéologique des pointes aziliennes du Bois-Ragot. In Chollet A, Dujardin V (eds) La grotte du Bois-Ragot à Gouex (Vienne). Magdalénien et Azilien. Essais sur les hommes et leur environnement, Paris: Société Préhistorique Française (Mémoire XXXVIII de la Société Préhistorique Française), pp 183–189
Pöllath N, Dietrich O, Notroff J, Clare L, Dietrich L, Koksal-Schmidt Ç, Schmidt K, Peters J (2017) Almost a chest hit: an aurochs humerus with hunting lesion from Göbekli Tepe, south-eastern Turkey, and its implications. Quat Int. https://doi.org/10.1016/j.quaint.2017.12.003
Potts R, Shipman P (1981) Cutmarks made by stone tools on bones from Olduvai Gorge, Tanzania. Nature 291:577–580
Quilès J (2004) Tanières d’ours des cavernes (Carnivora, Ursidae) du pourtour Méditerranéen: étude taphonomique et paléobiologique de huit assemblages du Pléistocène supérieur. Paléo 16:171–192
Rabeder G, Döppes D, Krofel M, Pacher M, Pohar V, Rauscher KL, Withalm G (2004) List of fossil faunal remains from Potoćka zijalka. In: Pacher M, Pohar V, Rabeder G (eds) Potoćka zijalka, palaeontological and archaeological results of the campaigns 1997–2000. Oesterrichische Akademie der Wissenschaften, Wien, pp 47–48
Ravazzi C, Peresani M, Pini R, Vescovi E (2007) Il Tardoglaciale nelle Alpi italiane e in Pianura Padana: evoluzione stratigrafica, storia della vegetazione e del popolamento antropico, Il Quaternario. Ital J Quat Sci 20(2):163–184
Reimer PJ, Bard E, Bayliss A, Warren Beck J, Blackwell PG, Bronk Ramsey C, Buck CE, Cheng H, Edwards RL, Friedrich M, Grootes PM, Guilderson TP, Haflidason H, Hajdas I, Hatté C, Heaton TJ, Hoffmann DL, Hogg AG, Hughen KA, Kaiser KF, Kromer B, Manning SW, Niu M, Reimer RW, Richards DA, Scott EM, Southon JR, Staff RA, Turney CSM, van der Plicht J (2013) Intcal13 and marine13 radiocarbon age calibration curves 0-50,000 years Cal BP. Radiocarbon 55(4):1869–1887
Rocci Ris A, Cilli C, Malerba G, Giacobini G, Guerreschi A (2005) Archeozoologia e tafonomia dei reperti provenienti da un complesso epigravettiano di Riparo Tagliente (Vr). In: Malerba G, Visentini P (eds) Atti del IV Convegno Nazionale di Archeozoologia, Quaderni del Museo Archeologico del Friuli Occidentale 6: pp 111–123
Romandini M, Nannini N (2012) Chasseurs épigravettiens dans le territoire de l’ours des cavernes: le cas du Covolo Fortificato di Trene (Vicenza, Italie). L’Anthropologie 116(1):39–56
Romandini M, Terlato G, Nannini N, Tagliacozzo A, Benazzi S, Peresani M (2018) Bears and humans, a Neanderthal tale. Reconstructing uncommon behaviors from zooarchaeological evidence in southern Europe. J Archaeol Sci 90:71–91
Rust A (1943) Die alt- und mittelsteinzeitlichen Funde von Stellmoor. Karl-Wachholtz Verlag, Neumünster
Selvaggio MM, Wilder J (2001) Identifying the involvement of multiple carnivore taxa with archaeological bone assemblages. J Archaeol Sci 28:465–470
Serwatka K (2018) What’s your point? Flexible projectile weapon system in the Central European Final Palaeolithic. The case of Swiderian points. J Archaeol Sci Rep 17:263–278
Shipman P (1981) Life history a fossil. An introduction to taphonomy and paleoecology. Harvard University Press, Harvard
Shipman P, Rose J (1984) Cutmark mimics on modern fossil bovid bones. Curr Anthropol 25:116–177
Shott MJ (1997) Stones and shafts redux: the metric discrimination of chipped stone dart and arrow points. Am Antiq 62:86–102
Smith MJ, Brickley MB, Leach SL (2007) Experimental evidence for lithic projectile injuries: improving identification of an under-recognized phenomenon. J Archaeol Sci 34:540–553
Soriano S (1998) Les microgravettes du Périgordien de Rabier à Lanquais (Dordogne). Analyse technologique fonctionnelle. Gallia Préhistoire 40:75–94
Stiner MC (1994) Honor among thieves: a zooarchaeological study of Neanderthal ecology. Princeton University Press, Princeton
Stiner MC (1998) Mortality analysis of Pleistocene bears and its paleoanthropological relevance. J Hum Evol 34:303–326
Stiner MC (1999) Cave bear ecology and interactions with Pleistocene humans. Ursus 11:41–58
Swanton JR (1905) Contributions to the ethnology of the Haida. Publications of the Jesup North Pacific Expedition 10(2), Memoirs of the American Museum of Natural History 8, pp 1–300
Tanck E, Homminga J, van Lenthe GH, Huiskes R (2001) Increase in bone volume fraction precedes architectural adaptation in growing bone. Bone 28:650–654
Thomas DH (1978) Arrowheads and atlatl darts: how the stones got the shaft. Am Antiq 43:461–472
Tillet T, Binford LR (2002) L'Ours et L'Homme. Études et Recherches Archéologiques de l'Université de Liège, Liège
Torres T, Ortiz JE, Cobo R, de Hoz P, García-Redondo A, Grün R (2007) Hominid exploitation of the environment and cave bear populations. The case of Ursus spelaeus Rosenmüller-Heinroth in Amutxate cave (Aralar, Navarra-Spain). J Hum Evol 52:1–15
Tuniz C, Bernardini F, Cicuttin A, Crespo ML, Dreossi D, Gianoncelli A, Mancini L, Mendoza Cuevas A, Sodini N, Tromba G, Zanini F, Zanolli C (2013) The ICTP-Elettra X-ray laboratory for cultural heritage and archaeology. Nucl Instrum Methods Res A 711:106–110
Turk I (ed) (2014) Cave Bear: prey of Neanderthal hunters or animal of special importance?, vol. 2. Opera Instituti Archaeologici Sloveniae, Ljubljana
Val A, Costamagno S, Discamps E, Chong S, Claud E, Deschamps M, Mourre V, Soulier M-C, Thiébaut C (2017) Testing the influence of stone tool type on microscopic morphology of cut-marks: experimental approach and application to the archaeological record with a case study from the Middle Palaeolithic site of Noisetier Cave (Fréchet-Aure, Hautes-Pyrénées, France). J Archaeol Sci Rep 11:17–28
Valentin B (2008) Jalons pour une paléohistoire des derniers chasseurs (XIVe-VIe millénaire avant J.-C.), Publications de la Sorbonne, Cahiers Archéologiques de Paris 1, Paris
Van Kolfschoten T, Parfitt SA, Serangeli J, Bello SM (2015) Lower Paleolithic bone tools from the ‘Spear Horizon’ at Schöningen (Germany). J Hum Evol 89:226–263
Villa P, Mahieu È (1991) Breakage patterns of human long bones. J Hum Evol 21:27–48
Wallduck R, Bello SM (2018) Cut mark micro-morphometrics associated with the stage of carcass decay: a pilot study using three-dimensional microscopy. J Archaeol Sci Rep 18:174–185
Weissert H (1981) Depositional processes in an ancient pelagic environment: the Lower Cretaceous Maiolica of the Southern Alps. Eclogae Geol Helv 74(2):339–352
Withalm G (2004) New evidence for cave bear hunting from Potočka zijalka (Slovenia). Mitt Komm Quartärforsch Österr Akad Wiss 13:219–234
Witthoft J (1968) Flint arrow points from the Eskimo of Northwestern Alaska. Expedition, Winter 1968
Wojtal P, Wilczyński J, Nadachowski A, Münzel SC (2015) Gravettian hunting and exploitation of bears in Central Europe. Quat Int 359-360:58–71
Wolverton S (2001) Caves, ursids, and artifacts: a natural-trap hypothesis. J Ethnobiol 21:55–72
Wood J, Fitzhugh B (2018) Wound ballistics: the prey specific implications of penetrating trauma injuries from osseous, flaked stone, and composite inset microblade projectiles during the Pleistocene/Holocene transition, Alaska U.S.A. J Archaeol Sci 91:104–117
Yeshurun R, Yaroshevich A (2014) Bone projectile injuries and Epipaleolithic hunting: new experimental and archaeological results. J Archaeol Sci 44:61–68
Yravedra J, Maté-Gonzàlez MÁ, Palomeque-Gonzàlez JF, Aramendi J, Estaca-Gòmez V, San Juan Blazquez M, Garcìa Vargas E, Organista E, Gonzàlez-Aguilera D, Arriaza MC, Cobo-Sànchez L, Gidna A, Uribelarrea Del Val D, Baquedano E, Mabulla A, Domìnguez-Rodrigo M (2017) A new approach to raw material use in the exploitation of animal carcasses at BK (Upper Bed II, Olduvai Gorge, Tanzania): a microphotogrammetric and geometric morphometric analysis of fossil cut marks. Boreas 46(4):860–873
Ziggiotti S (2006) Studio funzionale delle armature microlitiche dei siti del Piancavallo (Pordenone). Bollettino della Società Naturalisti “Silvia Zenari” 30:37–51
Ziggiotti S (2008) Strategie di caccia degli ultimi epigravettiani. Lo studio funzionale delle armature litiche di Riparo La Cogola, livello 19. PreistAlp 43:13–24
Acknowledgements
The fieldwork in Cornafessa rock shelter has been supported by MUSE-Science Museum of Trento; the authors are grateful to M. Lanzinger, director of MUSE, for endorsing this research project. The authors also thank the Soprintendenza per i Beni Culturali della Provincia Autonoma di Trento for granting permission to excavation activities; F. Cavulli, D.E. Angelucci (University of Trento) and S. Neri (MUSE) for collaborating in excavation and research activities; N. Angeli and P. Ferretti (MUSE) for the assistance in SEM and EDX analysis; and P. Cocco (MUSE) for the support in language editing. Finally, our thanks are also due to the two anonymous reviewers for their useful comments and helpful suggestions.
Author information
Authors and Affiliations
Contributions
R.D. and N.N. designed research; R.D. and G.D. supervised archeological excavations; N.N., A.F., and R.D. performed zooarcheological and taphonomical analysis; F.Bo. and J.C. carried out 3D documentation and analysis; and F.Be. and C.T. carried out μCT analysis; all the authors wrote and edited the manuscript.
Corresponding author
Electronic supplementary material
Table S1
(DOCX 59 kb)
Rights and permissions
About this article

Cite this article
Duches, R., Nannini, N., Fontana, A. et al. Archeological bone injuries by lithic backed projectiles: new evidence on bear hunting from the Late Epigravettian site of Cornafessa rock shelter (Italy). Archaeol Anthropol Sci 11, 2249–2270 (2019). https://doi.org/10.1007/s12520-018-0674-y
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s12520-018-0674-y