
Abstract
This study explored dietary behavior in the context of the developing medieval power centers of Bohemia (9th–11th Century AD, Czech Republic) with an emphasis on the dietary behavior of elites and the socio-economic stratification within the population of the central places. Carbon and nitrogen isotopic values were measured in collagen samples from 102 humans and 24 animals excavated from the sites of two castles representing the main power centers and their hinterlands. The Bayesian software package FRUITS was employed to estimate the caloric contribution of C3 and C4 plants, terrestrial animals, and freshwater fish. Statistically significant differences in the consumption of animal products and millet were observed between castle elites versus castle non-elite and hinterland samples. Among burial sites located outside the castle areas, substantial dietary variation in terms of both carbon and nitrogen was observed in males but not in females. These results suggest the deep socio-economic stratification within the population of centers. The notable consumption of millet was typical of the lower socio-economic groups buried outside the castle areas. The almost total absence of adult male skeletons at some sites was accompanied by the low contribution of animal products to the diets of the males present. The high dietary variation observed within the ducal family suggests either the rapidly changing position of the Přemyslids during the formation of the Bohemian state structure or the sharp increase in the influence of the Church in terms of elite dietary behavior.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Historical background
The 9th and 10th Centuries AD represent a period of the creation and consolidation of the state structure, which still represents a core of the current Czech state. Although archaeological findings as well as written evidence document the development of elite classes from as early as the 7th Century, the development of society at the beginning of the 9th Century strongly accelerated under the influence of the neighboring power centers of the Carolingian Empire, and the Great Moravian Empire, in particular. The 9th Century in central Bohemia was characterized by the gradual centralization of power. The cultural and political influence of Great Moravia culminated during the last quarter of the 9th Century, when the first, historically known, Duke of the Přemyslid dynasty, Bořivoj I, was baptized at the Great Moravian court (before 885 AD). Bohemia even became an integral part of Great Moravia for a short time (Berend et al. 2013; Bláhová et al. 2008).
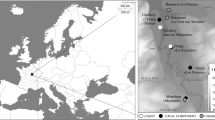
After the collapse of Great Moravia at the beginning of the 10th Century, the Přemyslid dynasty strengthened its power in central Bohemia, creating a web of fortified castles (Levý Hradec, Budeč, Prague etc.). From the start of the 10th Century onwards, Prague castle was the main center of the Přemyslid domain. At the same time, Přemyslids expanded into the rest of Bohemia. The consolidation of the Czech state was completed by the definitive annexation of Moravia at the beginning of the 11th Century (Berend et al. 2013; Bláhová et al. 2008; Charvát 2010) (Fig. 1).

Extension of the Early Czech State and neighboring European empires during the second half of the tenth century and the detailed view at the studied sites: Prague Castle and Levý Hradec. The positions of the studied burial sites are indicated by crosses. A—Third Courtyard; B—Second Courtyard and the Church of Virgin Mary; C—Střešovice–Triangl; D—Milady Horákové; E—Žalov–Cihelna; F—Žalov–Na Panenské. Modified from Sommer et al. (2009)
Climate and agricultural practices in early medieval Bohemia
The creation of the Czech state occurred during the so-called medieval warm period. For Central Europe, the beginning of this period began during the last quarter of the 9th Century, with climatic optimum culminating in the 11th century. Paleoclimate reconstructions are supported by historical data from this period, which highlight the relative scarcity of years of crop failure and famine. Such relatively favorable climatic conditions resulted in slow population growth. However, throughout this period, settlements remained concentrated in the most productive areas, generally under 250 m. a. s. l. (Beranová 2004; Bodri and Čermák 1995; Brázdil et al. 2005; Klápště 2005; Svoboda et al. 2003).
Based on archaeobotanical evidence, cereals were the most important staple food of plant origin. Wheat is considered to be the main bread cereal of this era. Generally, it is suggested that it was not before the high Middle Ages that wheat was gradually replaced by rye, although there are some indications of earlier changes in the distribution of crops during the 11th Century. Millet is always represented in the early medieval assemblages of plant macro-remains. It has been found in abundance in early medieval (9th–10th Century) contexts in Prague castle and its hinterland. Although millet could be over-represented in this type of analysis because of its processing method (in contrast to other grain, it was cleaned at home), it is still suggested to have an important role in the bohemian diet through the medieval period. In addition, legumes, such as peas and lentils, have been found at most early medieval sites (Kočár et al. 2010; Winklerová 2011).
During the study period, gradual differentiation of the composition of the zooarchaeological assemblages has been recorded, with the human population of power centers consuming more pork and fowl compared with those in rural areas, where there was a dominance of cattle and sheep and/or goats. This trend continued throughout the 9th–11th Centuries and culminated during the high Middle Ages. In addition, the proportion of wild fauna was generally higher in central places compared with rural regions, but still relatively low compared with the high medieval period. However, this comparison is based mainly on rural sites versus fortified centers. Information has been lacking on the immediate hinterland of these power centers. For pigs, extensive form of breeding occurred even in fortified centers, with the animals allowed to feed more or less freely on local vegetation (Kyselý 2000, 2005; Mlíkovský 2003; Winklerová 2011).
The consumption of fish remains an open issue. Osteological evidence is scarce and refers almost exclusively to freshwater fish. Remains of anadromous species (salmon or sea trout) are extremely rare (Kyselý 2003). However, this might be partly the result of the research methodologies used during ancient excavations (such as a lack of sieving, etc.) (Mlíkovský 2003; Winklerová 2011). Rare written sources relating exclusively to the beginning of the 11th Century mention a group of Anchorets from neighboring Poland, who literally ‘never ate fish’ (Hrdina et al. 2005). By contrast, the existence of ponds for fishing is documented at that time, usually as a part of donations to the church. At the end of the 11th Century, the first remarks appear related to the presence of imported salted sea fish (‘herynk’), which was given to the poor during fasting periods (Hrdina et al. 2005; Mlíkovský 2003). However, these rare written reports do not allow us to estimate the importance of fish in the diet of any population group. Moreover, it would be misleading to relate them to the situation during earlier Centuries because of the rapid development of religious as well as political and commercial spheres typical for the period under study.
Stable isotopes in the dietary reconstruction of past populations
Stable isotope analysis of bone collagen is, in terms of direct geochemical approaches, the most common approach to reconstructing past diet. It is based on the principal that the isotopic signal of the consumed diet and water is preserved in the tissues of consumer. Dietary reconstruction generally uses a combination of the isotopic values of carbon (δ13C) and nitrogen (δ15N). The carbon isotopic values enable us to define basic characteristics of the ecosystem that provided the dietary resources for the individuals under study: because of the distinct sources of carbon, δ13C allows us to distinguish between terrestrial and marine environments. Moreover, different discrimination against 13C during CO2 fixation in C3 vs C4 photosynthetic pathways, allows assessing the relative importance of C3 vs. C4 plants in the diet. Both carbon and nitrogen isotopic values increase significantly with trophic level, thus helping us to define the position of the organism within the food chain (Ambrose and Norr 1993; DeNiro and Epstein 1978, 1981; Lee-Thorp 2008; Minagawa and Wada 1984; Schoeninger and DeNiro 1984). The carbon isotopic values of herbivore collagen were shown to be about 5‰ higher than those of their forage (Ambrose et al. 1997), while the difference in collagen δ13C between animal species representing two adjacent trophic levels is on average about 1‰ (Bocherens and Drucker 2003; Lee-Thorp 2008). For δ15N, the magnitude of the stepwise trophic shift was estimated to vary between 3 and 5‰ based on controlled feeding experiments with different animal species (Hedges and Reynard 2007). Direct estimates for humans have recently become available (Fernandes et al., 2012a,b; O’Connell et al., 2012), suggesting higher values of ca.5.5–6‰. Although the isotopic values of freshwater ecosystems are highly variable, the combination of carbon and nitrogen isotopic values can also help estimate the importance of freshwater fish consumption (Dufour et al. 1999; Fuller et al. 2012b; Katzenberg and Weber 1999).
More recently, isotopic data analysis has been enriched beyond the simple interpretation of δ13C/δ15N biplots by attempting quantitative diet reconstruction. This employs mainly linear mixing models (Newsome et al. 2004; Phillips 2001) or, more recently, Bayesian approaches. The latter have the advantage of being able to incorporate different sources of uncertainty as well as coping with undetermined systems, in which the number of food sources is greater than the number of dietary proxies plus one (Fernandes et al. 2014; Parnell et al. 2010). These quantitative diet reconstructions enable us to account for a number of parameters, such as the isotopic baselines in the region and period studied, the composition of the various food groups included in the model, and offset values between dietary fractions and the dietary proxies used.
Aims of the study
The diet of the early medieval Bohemian population was characterized with a focus on the relation between diet and socio-economic status of each individual. The access to dietary resources, particularly to animal products, has been observed to be linked to socio-economic status in other medieval populations (Kaupová et al. 2016; Kjellström et al. 2009; Yoder 2012). Some observations suggest increases in socio-economically motivated dietary differences, along with growing social complexity, during the Middle Ages (Hakenbeck 2013). Accordingly, in the early medieval Bohemian population, the dietary proportion of animal products should be linked to socio-economic status, as reflected by the place of burial (within/outside the castle area, or in the hinterland) or by burial rite (e.g., presence and quality of grave goods). We also aimed to estimate the importance of millet consumption in the population subgroups and to compare the results with archaeobotanical and historical findings, which give contradictory evidence concerning the prestige and value of millet. According to historical resources, millet was consumed primarily by the poor and during famine periods (Adamson 2004); however, local archaeobotanical studies have found it in abundance even in Prague castle (Kočár et al. 2010; Winklerová 2011). In addition, we also explored the consumption of fish in relation to changing religious motivations (Barrett and Richards 2004; Rutgers et al. 2009). Increased pressure to abide by Christian rules is assumed to have resulted in the increased consumption of fish by elites and more recently dated groups from the 11th Century. Moreover, the increased intensity of trade after 1000 AD (Barrett et al. 2004) might have contributed to the increased consumption of fish (including those of marine origin) in these samples. Finally, special attention was paid to the dietary reconstruction of highest elites inhabiting Prague castle, especially members of the ruling Přemyslid dynasty. The samples under study come from the period of formation of the hereditary aristocracy. This process of the rise of individuals and families to the peak of the social hierarchy is thought to be reflected in dietary changes over time.
Material
This study focused on the dietary reconstruction of a population sample from Prague castle and its immediate hinterland (i.e. sites located less than 1 km from the castle) as well as a population sample from Levý Hradec castle as a comparative data set (Fig. 1). Both these strongholds were important hubs in the web of Přemyslid fortified castles, although their development followed distinct paths. The first signs of dominance of Prague castle within the Přemyslid domain are evident from the end of the 9th Century onwards. Gradually, its importance outgrew the local area, as demonstrated by the written testimony of a Judaeo-Arabic merchant, Ibrahim-Ibn-Yakub, who described Prague of the 960th as a ‘town built of stone and lime’ and important center of long-distance trade (Rapoport 1929). By contrast, the importance of Levý Hradec declined during the 10th Century along with the growing prestige of Prague. However, the castle continued to be one of the centers of the newly created state until the end of the 11th Century (Frolík and Smetánka 1997; Tomková 2013).
During the study period, burial sites were established both within and outside the castle areas. Under the growing influence of Christianity, the traditional means of demonstration of social status in the burial rite (e.g., by including rich grave goods) diminished, and the place of burial became the main way of declaring the power and importance of the buried person. Subsequently, burial sites within castle areas are supposed to be reserved for members of the nobility. Burial sites immediately surrounding the castle areas then belong to lower socio-economic groups of the castle population, but also to elites buried according to more traditional burial rites. Finally, in the relatively close vicinity of castles, burial sites are also found of population groups that lived in the hinterland and supplied in some way the needs of the castle. Thus, the skeletal material included in the current study was chosen to represent these three above mentioned contexts. The traditional use of grave goods continued to some extent in burial sites located outside the castle area, and these have been used to estimate the socio-economic stratification of the population group buried therein (Bláhová et al. 2008; Frolík 2016; Tomková 2012).
Given the unique historical contexts outlined above, the current study was limited by several problems relating to archaeological and anthropological praxis. Archaeological terrains are often largely disturbed by later development and have also long attracted the attention of both professional and nonprofessional archaeologists; thus, many archaeological sites, including those detailed here, had already been subject to excavations by the first half of the 20th Century (Frolík and Smetánka 1997; Tomková 2012). Thus, only relatively modest sample sizes were available for some of investigations reported here (Table 1).
Prague castle
The individuals investigated in this study came from three different contexts (Table 1). All were recovered from the castle area and, thus, were assumed to belong to members of the highest socio-economic rank. The first, small burial site at the so-called ‘3rd courtyard’ represents the oldest generations inhabiting Prague castle. This burial site, with 13 inhumations, but without a connection to any ecclesiastic building, was located at the most elevated part of the Prague promontory, in a highly prestigious and symbolic position (Frolík and Smetánka 1997; Profantová 2005).(Vlček 1997).
The second burial site was the cemetery at the so-called ‘2nd courtyard’ of Prague castle, located around one of the oldest known ecclesiastic buildings in Bohemia: The Church of the Virgin Mary. Thus, the skeletal collection of this cemetery included the first generations of Christians buried according to the new rite: that is, close to the church and without any grave goods (Frolík 2016).
The final set of samples from Prague castle came from graves attributed to members of the ruling Přemyslid dynasty. The sampled individuals came from the interiors of three churches (Table 1), established at Prague castle during the 9th and 10th Centuries (Frolík 2005; Vlček 1997). Most of the remains were secondarily exhumed and had been transferred (often repeatedly), which complicated the identification of the buried individuals. According to current understanding, the collection of dynastic graves studied here was assumed to include: (1) Duke Spytihněv I (died 915) and his wife (sample identities: PMC01 and PCM02); (2) Duke Vratislav I (PCG01; died 921); (3) an unknown duke and his wife (PCV01 and PCV02) [an alternative hypothesis suggests the male skeleton belong to Boleslav I (died 972) or his son Boleslav II (died 999)]; (4) and (5) two unknown dukes (PCG02 and PCG03); these skeletal remains were attributed by various researchers alternatively again to Boleslav I or to the members of the fifth generation of Přemyslids, who ruled at the start of the 11th Century: Oldřich (died 1034) or Jaromír (died 1035); and (6) a juvenile individual (PCG04; died between 16 and 20 years of age); there are no clues as to the identity of this individual, but because of the prestigious location and construction of the grave, he or she is thought to have been a member of the ducal family (Frolík 2005; Vlček 1997).
Hinterland of Prague castle
The hinterland of Prague castle is represented by two burial sites: Střešovice–Triangl and Milady Horákové Street (Table 1), both located less than 500 m from the castle. The character of the graves, as well as the quality of grave goods, suggests relatively poor character of these burial sites. The people who were buried in these sites were assumed to live outside the castle but to have some form of dependency on it (Frolíková-Kaliszová 2014; Tomková 2013). For the Střešovice sample, anthropological evidence suggested that the workload of this population group differed from the contemporaneous rural population groups, suggesting that they were perhaps craftsmen or servants (Frolíková-Kaliszová 2014; Stránská 2014). Interestingly, for the Milady Horákové collection, anthropological examination revealed some specific features of this population group. First, the majority of remains were those of subadult individuals (55%) and females, with an extremely low number of adult males. Moreover, a high prevalence of various pathological findings and dental anomalies was also found in this population sample (Stránská 2012a).
Levý Hradec burial sites
The population group inhabiting Levý Hradec castle buried their dead during the study period at three, at least partially temporary overlapping burial sites, located outside but close to the castle area. Unfortunately, the skeletal material from the burial site, which, based on findings of exclusive grave goods, belonged to the highest socio-economic classes, have not been preserved for analysis. From the two burial sites with preserved skeletal material (Fig. 1), Žalov–Cihelna B represents the lower socio-economic groups inhabiting Levý Hradec, whereas the slightly older (Table 1) data set from Žalov–Na Panenské, represents the first generations of Levý Hradec inhabitants, a more socio-economically stratified population with the potential presence of some elites buried according to traditional burial rites (Tomková 2012, 2013). Interestingly, Žalov–Na Panenské shares some untypical demographic features with the Milady Horákové burial site, with the majority of remains being female, although with a lower percentage of subadults (44%) (Stránská 2012b).
Data set
Only adults with estimated sex and age-at-death were included in the study with several exceptions resulting from the availability of skeletal material: because of the low number of individuals, several adults of undeterminable sex were included in the Prague castle data set; likewise, because of the low number of adult males, several juveniles (16–20 years) were measured as a part of the Milady Horákové sample.
For this study, it was impossible to gather animal bones from all the studied sites. Thus, the collection of animal bones examined (N = 24) came exclusively from the context of Prague castle and Levý Hradec castle (Table 1). The gathering of fish bones was difficult because ancient excavations had led to loss of the majority of tiny fish remains. Thus, these had to be collected from the context of slightly younger period spanning from the 11th to 13th Century.
Methods
Sex was estimated based on morphological and morphometric evaluations of the hip bone (Bruzek 2002; Murail et al. 2005) or, if the hip bone was absent, on skull morphology (Acsádi and Nemeskéri 1970; Ferembach et al. 1980). Age-at-death estimation was based on the assessment of skeletal maturation and morphological changes of the pubic symphysis and auricular surface (Ferembach et al. 1980; Lovejoy et al. 1985; McKern and Stewart 1957; Schmitt 2005, 2008). Given the sample size, individuals were divided in two groups: ‘younger’ (20–35 years old) and ‘older’ (> 35 years old) for further analysis.
For stable isotope analysis, 200 mg of cortical bone was sampled from each individual, preferentially from the ribs. In skeletal collections from ancient excavations (i.e., Prague castle and Žalov–Cihelna B), other bones had to be sampled: generally long bones of the upper and lower limbs, but in several cases also vertebrae (N = 2) or skulls (N = 2). In these cases, the skeletons were either only partially excavated (or at least stored) or we could not be sure that all the skeletal elements belonged to the same individual. Collagen extraction proceeded according to the Longin (1971) method, modified by Bocherens (1992). Elemental analyses were performed using a Europa Scientific EA elemental analyzer connected to a Europa Scientific 20–20 IRMS for carbon and nitrogen isotopes analysis at Iso-Analytical Limited, Crewe (UK). The uncertainty of isotopic measurements calculated on different standard replicates (IA-R042, IA-R045, IA-R046 and IA-R05, IAR06) was less than 0.1‰ (1SD) for both nitrogen and carbon.
In this paper, dietary compositions were evaluated using the Bayesian Mixing Model FRUITS (version 3.0; http://sourceforge.net/projects/fruits/ [last access: 12/21/2017]; Fernandes et al. 2014, 2015). FRUITS was chosen for its user-friendly interfaces, as well as its ability to handle different uncertainties, such as the isotopic signals of potential food groups, diet-to-tissue isotopic offsets and dietary routing. FRUITS also provides the possibility of introducing prior information into the model. This can include, for instance, physiological restrictions on the relative intake of macronutrients or information on dietary composition coming from e.g. archaeobotanical or zooarchaeological data.
On the basis of the archeological and historical sources, both C3 and C4 plants as well as terrestrial animals and freshwater fish must be considered as dietary sources in the model. Using δ13C and δ15N in bone collagen as dietary proxies, the weighted, offsets-dependent and concentration-dependent model was developed. Isotopic baselines and further modelling parameters are listed in Online Resource 1.
Given that the isotopic data and simulated foodstuff contributions were not normally distributed in most cases, nonparametric methods (Mann-Whitney test or Kruskal-Wallis test) were used to compare isotopic values as well as percentage calorie contributions among biologically and socio-economically defined subsamples. For individuals buried outside the castle area (Střešovice, Milady Horákové, and Levý Hradec), the character of the burial rite (i.e. single grave pits, the presence of grave goods, etc.) enabled us to evaluate the potential relationship between different markers of socio-economic status (presence and quality of grave goods, presence of stone and/or wood grave constructions) and the dietary characteristics of the individual. Statistical analyses were performed using R v12.1 software.
Results
All the samples met the criteria for good collagen preservation (Online Resources 2 and 3) (DeNiro 1985; Van Klinken 1999). C:N ratios of all the samples varied between 3.1 and 3.5, with a median of 3.2. Data sets were also examined for linear correlations between indicators of collagen preservation and isotopic values (Online Resource 4). Concerning human data, no significant correlation was found for any of the sites. For the animal samples, weak but significant correlations were found between nitrogen content and δ15N and between C:N ratio and δ13C. When restricting our sample only to animals of a similar habitat and subsistence (i.e. domesticated herbivores), these correlations disappeared, although a weak correlation between carbon content and δ15N occurred. However, because of the overall weak power of the correlation (p values were close to the level of significance), the narrow range of the collagen preservation criteria, and the absence of any outliers affecting the correlation significantly, we consider this to be probably an artifact of the small sample size. Subsequently, no samples were excluded from further analysis.
Animal data
For terrestrial animals (N = 19), the δ13C values ranged from −21.7‰ to −17.7‰ with a median of −20.9‰, and δ15N ranged from 4.3‰ to 9.1‰ with a median of 6.9‰. Excluding the two fowl samples showing distinct carbon isotopic values (Fig. 2), the maximal δ13C value of terrestrial animals was −19.6‰. Statistical analysis revealed no statistically significant difference (exact Mann-Whitney test; p = 0.998 for δ13C and 0.264 for δ15N) between pigs (N = 6; median is −21.0‰ for δ13C and 7.4‰ for δ15N) and domesticated herbivores (sheep/goats and cattle; N = 10; median is −21.0‰ for δ13C and 6.7‰ for δ15N). There was no isotopic difference (exact Mann-Whitney test; p = 0.441 for δ13C and 0.195 for δ15N) between data sets of the main domesticated species (pig, sheep/goat, cattle) from Prague castle (N = 8; median is −21.1‰ for δ13C and 5.9‰ for δ15N) and Levý Hradec (N = 8; median is −20.8‰ for δ13C and 7.1‰ for δ15N). The result was the same when only herbivores were considered (exact Mann-Whitney test; p = 0.537 for δ13C and 0.082 for δ15N). The small sample size precluded the more detailed statistical analysis of the animal sample. The fish data (N = 5) were highly variable, ranging from −29.0 to −16.2 with a median of −25.9‰ for δ13C and from 6.8‰ to 16.1‰ with a median of 7.8‰ for δ15N (Fig. 2).

Human (mean, percentile 2.5 and 97.5) and animal bone collagen isotope results, separated by species and site
Human isotopic data
When the dietary characteristics of the two burial sites from the area of Prague castle were compared, individuals from the 2nd and 3rd courtyards were undistinguishable isotopically, with carbon isotopic values ranging from −20.3 to −18.1‰ with a median of −19.2‰ and nitrogen isotopic values ranging from 9.3 to 11.4‰ with a median of 10.3‰ (for further details see Tables 2 and 3, Figs. 3 and 4). Thus, both samples were combined for further statistical analyses. In this combined sample, there was no relation between sex and isotopic values of the individual (Table 3). The average calorie contribution of each food group for the Prague castle sample was ca. 71% for C3 plants, 11% for C4 plants (i.e. millet), 15% for terrestrial animals and 2% for freshwater fish (Table 2, Fig. 5).

Human and animal (median, percentile 2.5 and 97.5) isotopic data for all sites. a PC = Prague castle. b without the outlier PCF15, with isotopic values suggesting catadromous/anadromous or marine origin

Human bone collagen isotope values from (a) Prague castle; (b) hinterland of Prague castle; (c) Levý Hradec castle; ND = undetermined sex. a sample identities from the Prague castle data set: PCM01 = Spytihněv I; PCM02 = Spytihněv’s wife; PCV01 = unknown duke from St. Vitus church ‘K1’; PCV02 = his wife ‘K2’; PCG01 = Vratislav I, PCG02 = unknown duke ‘G.92’, PCG03 = unknown duke ‘G.98’; PCG04 = unknown individual ‘G.102’

Average modeled contribution of four food classes (C3 plants, C4 plants, terrestrial animals, freshwater fish) to diet of various sample subgroups. PC = Prague castle; STR = Střešovice; MH = Milady Horákové street; LH = Levý Hradec castle
By contrast, the diet of supposed members of ducal family differed from the rest of the Prague castle population in terms of both δ13C (exact Mann-Whitney test; p = 0.035) and δ15N (exact Mann-Whitney test; p = 0.021) (Figs. 3 and 4). This group showed δ13C ranging from −19.4 to −18.1‰ with a median of −18.7‰ and δ15N ranging from 10.0‰ to 13.7‰ with a median of 11.3‰. However, these results lost their statistical significance when only males were considered (although the low number of samples should be taken into account) (Table 3). For this isotopically variable group, the group diet was not reconstructed. Individual estimations of the dietary contribution of each food group are shown in Fig. 6.

Modelled estimates of dietary composition of the members of Přemyslid family. The boxes represent the 16th and 84th percentiles (68% of the data) and the whiskers the 2.5 and the 97.5 percentiles (95% of the data) for the contribution of each food source. The continuous lines illustrate means and the discontinuous lines median values. Sample identities: PCM01 = Spytihněv I; PCM02 = Spytihněv’s wife; PCV01 = unknown duke from St. Vitus church ‘K1’; PCV02 = his wife ‘K2’; PCG01 = Vratislav I, PCG02 = unknown duke ‘G.92’, PCG03 = unknown duke ‘G.98’; PCG04 = unknown individual ‘G.102’
When looking at isotopic data from both sites representing the hinterland of Prague castle, the comparison between these two data sets revealed a statistically significant difference in δ15N (Mann-Whitney test; p = 0.042), with the Střešovice sample showing higher values. The result for δ13C was close to the level of statistical significance (Mann-Whitney test; p = 0.067), again with Střešovice sample showing higher values (Fig. 4). Considering each sex separately, differences occurred primarily between the male samples (exact Mann-Whitney test; p = 0.014 for δ13C and < 0.001 for δ15N), while females from both samples appeared undistinguishable isotopically (exact Mann-Whitney test; p = 0.881 for δ13C and 0.696 for δ15N). For this reason, each of the two samples was treated separately in all the following statistical analyses.
For the Střešovice sample, carbon isotopic data (Table 2) showed a narrow range, from −19.0‰ to −18.2‰ with a median of −18.6‰. Nitrogen isotopic data ranged from 7.3‰ to 10.1‰, with a median of 9.7‰. Neither sex nor age-at-death had any significant impact on isotopic values (Table 3). The average calorie contribution of each food group for the Střešovice sample was ca. 75% for C3 plants, 16% for C4 plants, 8% for terrestrial animals and 1% for freshwater fish (Table 2).
For the Milady Horákové sample, carbon isotopic data showed higher variability (Table 2), ranging from −20.4‰ to −17.0‰, with a median of −18.8‰. Nitrogen isotopic data ranged from 7.3‰ to 10.1‰ with a median of 9.3‰. Characteristic for this site was the isotopic difference between both sexes, with males showing both lower δ13C (exact Mann-Whitney test; p = 0.002) and δ15N (exact Mann-Whitney test; p = 0.041) values. There was also a significant effect of the age-at-death on δ13C (exact Mann-Whitney test; p = 0.010), with individuals younger than 35 years showing lower δ13C. However, this was likely to be a relict of the untypical demographic structure of the burial site, where most males (N = 6) were estimated to have died before 35 years of age, while there was only one female younger than 35 years in the whole sample.
To confirm that the inclusion of juveniles did not affected the representativeness of the Milady Horákové data set compared with the other skeletal samples, we compared the isotopic data of juvenile and adult males. Juveniles (N = 3) did not differ significantly from adults (N = 6) (exact Mann-Whitney test; p = 0.714 for δ13C and 0.998 for δ15N). Only one juvenile female was included into the sample and her isotopic values (δ13C = −19.3‰, δ15N = 10.1‰) were also located within the variability of the overall female sample (Table 2). Based on these results, there is no trace of the potential decrease in nitrogen isotopic values resulting from the negative nitrogen balance during growth (Waters-Rist and Katzenberg 2010). The average calorie contribution of each food group for the Milady Horákové sample was ca. 79% for C3 plants, 13% for C4 plants, 7% for terrestrial animals and 1% for freshwater fish. Substantial variation was however observed between the sexes, with the male diet containing only about 4% of terrestrial animals and 10‰ of C4 plants (Table 2).
The comparison of both data sets from Levý Hradec (Žalov–Cihelna B and Žalov–Na Panenské) (Table 3, Fig. 4) showed no significant isotopic differences between the samples (exact Mann-Whitney test; p = 0.640 for δ13C and 0.538 for δ15N). The result remained unchanged when only females were compared. Given the low number of males recovered at the Žalov–Na Panenské site (N = 2), it was not possible to compare the dietary characteristics between males from both sites. For the combined Žalov sample (Table 2), carbon isotopic data ranged from −19.4‰ to −17.3‰, with a median of −18.8‰, while nitrogen isotopic data ranged from 7.5‰ to 10.7‰, with a median of 9.4‰. Neither sex nor age at death appeared to have a significant impact on the carbon or nitrogen isotopic data from the Žalov sample (Table 3). The average calorie contribution of each food group for the Levý Hradec sample was ca. 77% for C3 plants, 15% for C4 plants, 7% for terrestrial animals and 1% for freshwater fish.
Neither the presence and quality of grave goods nor the presence of grave construction had any impact on the nitrogen isotopic values at any of the sites. In Levý Hradec females only, the quality of grave goods appeared to be linked to carbon isotopic data (exact Mann-Whitney test; p = 0.021), δ13C increased from graves without grave goods (group C: median = −19.2‰) to rich graves (group A: median = −18.6‰) (Table 3).
Intrapopulation variability in the diet of early medieval Czech populations
Given the dietary differences found among samples from the hinterland of Prague castle, we compared statistically the following four units: Prague castle, Střešovice, Milady Horákové, and Levý Hradec castle. The basic statistics and mean human–faunal offsets for these units are detailed in Table 2. Dynastic graves showed such specific dietary characteristics that they were excluded from all the following analyses.
The results of the statistical analysis revealed that the Prague castle sample showed significantly higher δ15N values (Table 4) than any of the other samples. In case of Střešovice, the result lost the statistical significance when each sex was considered separately, but the p values were still close to 0.05. The Prague castle sample showed lower δ13C values than the Střešovice and Levý Hradec samples as well as for females from Milady Horákové. Only males from the Milady Horákové sample showed similar δ13C values as those from Prague castle. For the Levý Hradec males, the result was close to the level of statistical significance (Table 4). When comparing the data from burial sites located outside the castle area, female isotopic values were comparable among all three units (Střešovice, Milady Horákové, and Levý Hradec). Male carbon isotopic values at Milady Horákové were lower than at Střešovice and Levý Hradec. Moreover, as stated previously, the nitrogen isotopic values of the male sample from Milady Horákové were significantly lower than from Střešovice. Levý Hradec males showed intermediate nitrogen isotopic values, but no statistically significant differences compared with the other two sites were found, although, compared with the Střešovice sample, the result was close to the level of statistical significance (Table 4).
Though large uncertainties are associated with the estimated mean percentage of resource contribution to individual as well as group diets (Table 2), comparisons of these values between sites are still informative. Statistical comparison of simulated foodstuff contributions confirms the results outlined above (Table 4).
The FRUITS simulation (Fig. 5 and Table 4) found a significant difference in the dietary contribution of terrestrial animals between Prague castle and all the other sites. According to the group diet reconstructions, the Prague castle population consumed roughly 7–8% more meat of terrestrial animals in comparison to other sites. In the cases of the Milady Horákové and Levý Hradec sites, p values were lower than 0.001 as recommended by Pestle et al. (2016) to account for the possibility of overfitting with the large sample sizes in the simulations (n = 10.000). Considering each sex separately, p values were generally higher but (with the exception of Střešovice) still significant at the 0.05 level. The higher p values are not surprising when taking into account the lower sample size. The Prague castle population also consumed ca. 4% less C4 millet than any other comparative dataset with the exception of Milady Horákové males (Table. 2 Fig. 5), The results of the statistical comparison were significant at the 0.05 level in all cases, with the exception of Levý Hradec vs Prague castle males. The results of the comparison between burial sites located outside the castle area also mirrors the trends outlined above. No significant differences were found in the composition of the female diet. The male sample shows higher variation, with the Střešovice vs Milady Horákové males differing significantly in the dietary contribution of terrestrial animal meat as well as millet, and the Levý Hradec sample showing intermediate values (Table 2, Fig. 5). Though the differences in the dietary contribution of fish were statistically significant, the actual dietary inputs from fish are so small (Table 2) that these could hardly be regarded as significant in terms of diet, and will not be discussed in the following section.
Discussion
Animal data: isotopic characteristics of the environment
With the exception of fowl, domesticated animals showed carbon and nitrogen isotopic values typical for the terrestrial C3-plant environment. One sample only (LHF05, pig) showed a δ13C higher than −20‰. Subsequently the minor use of millet as a supplementary fodder cannot be entirely excluded for this individual (Fuller et al. 2012a). As commonly observed previously (Fuller et al. 2012a; Kaupová et al. 2016; Lightfoot et al. 2012; Reitsema et al. 2013), two samples of fowl showed distinct isotopic values (Fig. 2), reflecting their omnivorous diet as well as their close coexistence with humans. Higher carbon isotopic values suggest a regular consumption of a certain amount of millet.
Although the sample size was small, the absence of statistically significant isotopic differences between pigs and domesticated herbivores confirmed the archaeological premise that, in 9th–11th-Century Bohemia, pigs were kept in a traditional extensive manner typical of less-urbanized areas. A similar pattern has been found at numerous European medieval sites leading up to the 13th Century (Hammond and O’Connor 2013; Kaupová et al. 2016; Knipper et al. 2013; Reitsema et al. 2013). Although with a distinct location as well as a slight difference in dating, both data sets from Prague and Levý Hradec showed comparable isotopic values, ensuring good comparability of human isotopic data between both sites.
From the highly variable fish data set, four of the samples fell within the range typical for European freshwater species. However, one sample (PHF04, Undetermined fish, δ13C = −16.2‰ and δ15N = 7.8‰) showed carbon isotopic values that might indicate catadromous, anadromous (such as sturgeon or eel), or even marine species (Fuller et al. 2012b; Kaupová et al. 2016; Reitsema et al. 2013; Vika and Theodoropoulou 2012). Given the slightly younger dating of the fish data set (Table 1), we could however not be sure if this fish could have been eaten by any member of the population group under study.
Diet in early medieval Bohemia
The isotopic results showed that the early medieval population of Bohemia had a terrestrial diet with varying proportions of C4 plants (Fig. 3). This was confirmed by the results of quantitative dietary analysis, which revealed the clear dominance of a C3 plant diet. The average dietary input of C4 plants varied between ca. 10–16% with regard to site and sex. The highest individual dietary contribution of C4 plants was 28% (Levý Hradec sample ZAP12). Despite the relatively large uncertainties of the estimates (Table 2), these results suggest that plants probably constituted more than half of the caloric intake, possibly much higher, with animals in second place. The average dietary contribution of terrestrial animals was relatively low, ranging from 4 to 15%. The highest individual dietary input from terrestrial animals, when dynastic graves were excluded, was observed in Prague castle individual PH203 (20%). At the other end of the spectra were several individuals from Milady Horákové and Levý Hradec with only marginal input of terrestrial animals (ca.3%).
Although the consumption of fish is probable in some individuals from dynastic contexts (see below), it appears that freshwater fish did not have an important role at the population level. The observed Δ15Nhuman-fauna (Table 2) in all the studied contexts were significantly lower than in populations that rely substantially on fish, and for which a Δ15Nhuman-fauna of 5 or more is typical (e.g. Katzenberg and Weber 1999; Piličiauskas et al. 2017; Rutgers et al. 2009). The FRUITS simulation confirmed that fish provided only a marginal caloric contribution, with an average input of only 1 to 2%. For fish, the consistently low estimates also have rather low uncertainties. This is in accordance with the general pattern observed across medieval Europe. Generally, the phenomenon of increased fish consumption as a reflection of the increased pressure to abide by the Christian fasting rules is dated after 1000 AD (e.g. Barrett et al. 2004; Kaupová et al. 2016; Müldner and Richards 2007b; Salamon et al. 2008). Although there are some exceptions, where the increased consumption of fish is evident in the recently Christianized populations of the first millennia (Barrett and Richards 2004; Rutgers et al. 2009), the 9th–10th Century population of Bohemia appears to have followed the general pattern of fish consumption.
Though a heavy reliance on plant diet has been observed in several studies reporting an average caloric input of plant food at about 75% or more (Maurer et al. 2017; Pestle et al. 2016; Sjögren 2017), it is however surprising, especially in the elite context of Prague castle. Being dependent on a substantial number of parameters, the results of the modelling may be sensitive to errors and inconsistencies in the suggested parameter values. Results may thus differ somewhat, when altering for example food or offset values. When testing the impact of changes in various model parameters, however, the overall dietary pattern of the studied samples appears relatively stable. The results of modelling always show a dominance of C3 plant derived food, as well as the marginal role of freshwater fish. The dietary contribution of C4 plants remains virtually unchanged. For a detailed discussion on the impact of various changes to baseline values and other model parameters, see Online Resource 1.
Dietary behavior as a reflection of socio-economic stratification: relation between isotopic data and burial site
According to our data, the dietary behavior of elites buried within the area of Prague castle differed from the rest of the studied sample (Figs. 4 and 5). When the group of dynastic graves was excluded, isotopic data from the Prague castle population as well as human–faunal isotopic offsets (Table 2) suggested an omnivorous, predominantly C3-based diet of terrestrial origin. Carbon isotopic values were at the border of distinguishable millet consumption (Fuller et al. 2012a). Mean carbon human–faunal offset slightly exceeded a maximum attributable to the higher trophic position (Δ13Chuman-fauna = 1.6) but similar isotopic offsets have also been observed in C3-based ecosystems of central Europe (Hakenbeck et al. 2010; Knipper et al. 2013; McGlynn 2007). The average calorie contribution of C4 plants estimated by FRUITS was ca.11%. This suggests millet was rather marginal in the diet of this population group.
The absence of any substantial difference in isotopic values between both studied units suggests that the diet of the Prague castle inhabitants did not change significantly during the study period and that those following the Christian burial rite (i.e. buried at the church cemetery in the 2nd courtyard) did not differ isotopically from individuals buried in a more traditional manner (in the 3rd courtyard).
Regarding the population groups from non-castle burial sites, inhabitants of Levý Hradec castle shared at least some dietary patterns with the hinterland population of Prague castle. Firstly, a measurable input of millet was typical of the diet of these population groups. The mean carbon isotopic offsets (Δ13Chuman-fauna) were higher than two in both the Střešovice and Levý Hradec samples (Table 2), as well as in females from Milady Horákové. The average calorie contribution of C4 plants was about 15% for these groups. This was significantly different from the population of Prague castle. The only exception was provided by males from Milady Horákové, who showed the lowest mean Δ13Chuman-fauna (1.4) and percentage contribution of C4 plants (10%) of all the studied samples, being comparable to the Prague castle population.
Second, compared with the population of Prague castle, isotopic data from both contexts suggested a relatively low consumption of animal protein. Although the consumption of eggs, poultry, or high-quality meat from young animals with a persistent breastfeeding signal can increase the isotopic values of castle elites and, thus, contribute to observed dietary differences (Hedges and Reynard 2007), the relatively low mean nitrogen isotopic offsets (Δ15Nhuman-fauna ranging from 2.5 to 3.0 at the non-castle burial sites) and the average calorie contribution of terrestrial animals about or below 10% suggested the substantially limited access of these population groups to animal protein.
Thus, both carbon and nitrogen isotopic values indicated that the diet of individuals from Levý Hradec had a ‘non-elite’ character, with a relatively low dietary input of terrestrial animals and higher proportion of millet. This corresponds with the prediction for the Žalov–Cihelna B samples, which were hypothesized to come from lower socio-economic groups of castle inhabitants. Although higher socio-economic stratification with the potential presence of some elite individuals was suggested for Žalov–Na Panenské, this was not reflected in the quality of their diet. If there were any individuals of higher socio-economic rank, they had the same dietary behavior as the rest of the population. This could correspond with the more ancient dating of the site (to the second half of the 9th Century) and, thus, the earlier phase of the formation of a social hierarchy. However, interestingly, at the same time at Prague castle, elites were clearly distinguishable by both place of burial and dietary behavior, as indicated by the isotopic data from the 3rd courtyard.
Elites excluded: dietary variation within and between non-castle burial sites
Though the above outlined common dietary patterns, the observed isotopic differences both among and within non-castle burial sites suggested further socio-economic stratification or specialization at early medieval power centers, which resulted in substantial differences in access to dietary resources. Despite the close geographic distance, there was a striking difference in dietary behavior between the two population groups from the hinterland of Prague castle. The Střešovice sample showed a narrow range of carbon (Min–Max = 0.8‰; SD = 0.3) isotopic values as well as the absence of any dietary differences between the sexes. Such a narrow range of δ13C with no outliers is uncommon, especially compared with other contexts of the documented input of C4 plants (Kaupová et al. 2016; Knipper et al. 2015; Lightfoot et al. 2012; Reitsema et al. 2017; Teschler-Nicola et al. 2015) and is comparable or even lower than in populations with a predominantly C3-based diet (Hakenbeck et al. 2010; Knipper et al. 2013; McGlynn 2007; Reitsema et al. 2017). This suggests highly homogenous dietary behavior within this group, who, based on anthropological findings, were suggested to be servants or craftsmen (Stránská 2014).
By contrast, the sample from the burial site at Milady Horákové showed higher isotopic variation in terms of nitrogen and especially carbon (Table 2) as well as a clear isotopic difference between the sexes, with the diet of males generally being lower in both millet and terrestrial animals. The mean Δ15Nhuman-fauna in Milady Horákové males (2.0) was the lowest from all the studied samples and suggested substantial limitation to their access to animal products. Despite the low number (N = 9) of male samples, this finding is interesting. Medieval populations are generally considered to have been male dominated (Bardsley 2007), which would be reflected by the preferential access of males to high-quality foodstuff (Müldner 2009). Although the isotopic results do not support this pattern unexceptionally, one of the two scenarios is generally observed: either in accordance with the above-mentioned premise, males consume a higher amount of animal (terrestrial or marine) products (Hakenbeck et al. 2010; Kaupová et al. 2016; Müldner and Richards 2007a; Reitsema et al. 2010; Richards et al. 2006) or both sexes show the same dietary behavior (Kaupová et al. 2016; Lightfoot et al. 2012; Müldner and Richards 2005; Reitsema et al. 2017). The exceptions, where males show a lower dietary proportion of animal products, are rare. Even in these cases, these differences are smaller (Reitsema and Vercellotti 2012) or are accompanied by higher isotopic variation in the male samples and, thus, might be linked to their higher mobility rate (Kjellström et al. 2009).
Interestingly, only one of the Milady Horákové males (MH20; δ13C = −18.8‰, δ15N = 9.2‰) showed isotopic values within the variation of the Střešovice sample. Although females showed average isotopic values comparable with the Střešovice sample, some shared the dietary pattern described above with males (MH22, MH29) or showed intermediate isotopic values (MH01, MH13).
Several hypotheses could be offered to explain this untypical dietary behavior. First, as some previous studies have suggested, male dietary behavior could be a more-sensitive indicator of the socio-economic status than that of females (Kaupová et al. 2016; Reitsema and Vercellotti 2012). However, this is in contrast to archaeological evidence that revealed no substantial differences in the burial rite between the Prague hinterland sites (Frolíková-Kaliszová 2014; Tomková 2013), suggesting a similar socio-economic structure of both population groups. Second, the explanation might be linked to the untypical demographic structure of the Milady Horákové burial site, which mainly comprised subadults and females. Moreover, most of the males were either juveniles or showed the presence of pathology, such as severe osteoarthritis (Stránská 2012a). Given that the burial site had almost been excavated in full, it appears that most of the adult male population were buried elsewhere. This might result from some specific function of adult males in relation to Prague castle, while only those who were too young, ill, or weak remained to be buried at Milady Horákové. Regrettably, the low number of male skeletons prevented a deep analysis of their health status, which might have found osteological signs of bad health or undernourishment on a statistical basis. Moreover, the observed difference in millet consumption is more difficult to explain. In general, millet is viewed as a crop of low socio-economic groups (Adamson 2004); thus, the relatively low δ13C values of the Milady Horákové sample suggest other possible explanations. It should also be noted that the average dietary contribution of terrestrial animals in the Milady Horákové males is so low (ca. 4%) that it forces us to consider the substantial input of dietary resources not included in the model (such as legumes). The presence of migrants coming from isotopically distinct areas also has to be taken into account. While several alternative scenarios may well explain the distinct isotopic values of the Milady Horákové males, the sex-based dietary difference in this population group appears unquestionable.
Data from the Levý Hradec sample showed isotopic values that were similar between both burial sites (Žalov–Cihelna B and Žalov–Na Panenské) and intermediate between both Prague hinterland sites. While the carbon isotopic values (mean Δ13Chuman-fauna = 2.2) and the average dietary proportion of millet (15%) were more similar to Střešovice, the nitrogen isotopic values (Δ15Nhuman-fauna = 2.5) and average dietary proportion of terrestrial animals (7%) were lower and more similar to the Milady Horákové sample (Table 2). When each sex was considered separately, the female diet appeared to be similar not only within the Levý Hradec area, but also compared with females living in the hinterland of Prague (Table 4). However, in males, the δ15N values were on average lower than in females (although the difference was not statistically significant). The Levý Hradec males also showed lower δ15N and lower average dietary input of terrestrial animals than the males from Střešovice, with the results varying around the 0.05 level of statistical significance (Table 4). Given the overall low number of male samples from Levý Hradec, this pattern could be influenced by the low δ15N values of the only two males recovered from Žalov–Na Panenské (ZAP02, ZAP13). The ZAP02 sample showed the lowest δ15N from the Levý Hradec sample (δ15N = 7.5), which was comparable with the lowest values from Milady Horákové. After the analogous demographic structure with the low percentage of males, the low dietary contribution of terrestrial animals in the present male individuals appears to a further analogy that also occurs in the Milady Horákové and Žalov–Na Panenské samples. Despite the incomplete excavation of skeletal material from Levý Hradec, as well as the overall low number of males recovered from all the non-castle contexts prompt caution when interpreting both the skeletal and isotopic data, the present results suggest that: (i) the diet of the male population was more stratified than that of the female population; and (ii) the population group of adult males with at least average input from terrestrial animals in the diet appears to be absent from some sites. Although, as mentioned above, alternative explanations such as the consumption of legumes must be taken into account, at least the first conclusion of higher stratification of the male diet remains valid even under such scenarios.
As stated above, the character of the burial rite (i.e., the presence of grave goods and constructions) enabled us to evaluate the potential relationship between dietary characteristics and these presumed indicators of socio-economic status at non-castle burial sites. However, the dietary proportion of animal products was not linked to any of the indicators of socio-economic status at any of the sites. The only statistically significant relationship was found between carbon isotopic values and the presence and quality of grave goods at Levý Hradec. This finding, suggesting the higher consumption of millet by individuals from well-equipped graves, does not fit the expected socio-economic gradient in millet consumption (Adamson 2004). Moreover, this result could be influenced by the two individuals from well-equipped graves, who showed the highest proportion of millet in their diet (ZAP12; δ13C = −17.3‰; ZAC09; δ13C = −17.6‰) in the whole data set. Thus, the observed difference is likely to be a relict of the small sample size.
Diet of the ruling house: dietary reconstruction of members of the Přemyslid family
Interestingly, the Přemyslid sample showed relatively high variability in terms of both carbon and nitrogen isotopic values (Table 2), which are clustered in two groups. For individuals buried in the church of the Virgin Mary (Spytihněv I and his wife) and St. Vitus rotunda (unknown duke PCV01 and his supposed wife PCV02), relatively high carbon isotopic values (> −18.5‰), together with relatively low nitrogen isotopic values (11.2‰ for Spytihněv I, as the highest) are typical. The calorie contribution of C4 plants varies between ca.17–19% for these individuals, while the proportion of terrestrial animals is typically below 20%. Freshwater fish were probably absent or marginal in the diet of these individuals. Their δ13C were located at the border of the Prague castle data range. Moreover, both carbon and nitrogen isotopic values were within the variation of the hinterland sample of Střešovice. Interestingly, both supposed couples shared the same pattern, with female δ13C being almost identical to that of the males, while δ15N being more than 0.5‰ less, suggesting most probably the lower importance of terrestrial animals in the female diet.
The group of burials from the St. George basilica showed slightly lower carbon isotopic values (clustering around −19‰) and the nitrogen isotopic values showed high variability. In the case of the unknown duke PCG03 and juvenile individual PCG04, δ15N values were only slightly higher than the maximum for the rest of the Prague castle sample, whereas Vratislav I (δ15N = 12.5‰; Δ15Nhuman-fauna = 5.9) and unknown duke PCG02 (δ15N = 13.7‰; Δ15Nhuman-fauna = 7.1) were clear outliers. The dietary proportion of millet was lower in these individuals, ranging between ca.11 to 15%. The dietary proportion of terrestrial animals was ca.20% in PCG03 and PCG04, and 30% in Vratislav I and PCG02. The Unknown Duke PCG02 is the only individual who consumed a substantial amount of fish (ca. 13% of caloric input).
Although the identity of members of the ducal family is disputable or completely unknown, the dating of the studied sample covers the relatively brief period of approximately 100 years, that is, the lifetimes of four to five generations of historically known Přemyslids (Vlček 1997). The above-mentioned results suggest that there was no typical diet for the ducal family during this period. Instead, dietary habits differed among individuals from a relatively plant-dominated diet with a traceable proportion of millet to a diet with a substantial proportion of terrestrial animals and freshwater products (Figs. 4 and 6) and lower importance of millet. This suggests that either there was no dietary norm and that the socio-economic position of these individuals enabled them to choose the diet according to personal taste, or that the dietary norm for the highest aristocracy changed over the studied period. The increasing influence of fasting prescriptions could be a potential explanation. The increased pressure to abide to the fasting rules is often suggested to explain the higher consumption of fish observed in medieval and early modern elites (Fornaciari 2008; Jiménez-Brobeil et al. 2016; Knipper et al. 2015; Lamb et al. 2014; Yoder 2012). However, the caloric input coming from fish was higher than 10% in only one case. Alternatively, the dietary change could reflect a challenge to the importance of the Přemyslid dynasty from one of the rival families and vassals in first generations to well established and independent ruling dynasty around the year 1000 (Berend et al. 2013; Bláhová et al. 2008).
Whatever the underlying causes, the results of our dietary analysis pose new questions concerning the identification of Přemyslid skeletal remains. Dietary dissimilarity between supposed brothers (Spytihnev I and Vratislav I) on the one hand and the similarity between Spytihnev I and the unknown duke from the St. Vitus church as well as between supposed Vratislav I and the unknown duke PCG02 on the other, offer new possibilities to explore. However, the influence of personal choice and taste in determining diets should not be underestimated.
Bohemian isotopic data in the context of medieval central Europe
A comparison of faunal isotopic data with other sites from central Europe (Table 5) showed slight but observable variation in carbon isotopic values (Hakenbeck et al. 2010; Halffman and Velemínský 2015; Kaupová et al. 2016; Knipper et al. 2013; Lightfoot et al. 2012; 'Olsen et al. 2016; Reitsema et al. 2013). This variation clearly shows that it would be misleading to compare the human isotopic values from central European contexts directly. On the other hand, it is not possible to set the estimates generated by FRUITS in a wider context. These studies are still scarce in the Central European context (Fernandes et al. 2015; Knipper et al. 2016), and completely omit the medieval period. Moreover, variations in the setting of model parameters (such as uncertainties or diet-collagen offsets) make the direct comparison of estimates problematic. Subsequently, despite substantial limitations (e.g. Fraser et al. 2013; Knipper et al. 2016), the comparison of human–faunal isotopic offsets is thus far the only way to set our data in a wider framework.
When comparing carbon human–faunal isotopic offsets among Central European populations (Table 6), it appears that the isotopic values of Levý Hradec castle and Prague hinterland samples (with the exception of males from Milady Horákové) are comparable with those from early medieval Moravian sites from the 9th–10th Centuries (Kaupová et al. 2016) and other population groups for which the notable consumption of millet is suggested (Lightfoot et al. 2012; Reitsema et al. 2017; Teschler-Nicola et al. 2015). By contrast, Prague castle data are comparable with populations with a predominantly C3-based diet (Hakenbeck et al. 2010; Knipper et al. 2013; McGlynn 2007). When focusing on a comparison with Moravian data, there is a different approach of the higher socio-economic classes to the consumption of millet. While, in Moravia, millet was consumed in an amount comparable across the rural population, the population of proto-urban centers and by members of highest socio-economic classes, in Bohemia a clear socio-economic gradient was observed, with the elite diet being predominantly C3 based. By contrast, human–faunal offsets from non-castle burial sites suggest a similar proportion of millet consumed as in the Great Moravian sample. The slight difference in the dating of both data sets must be taken into account when determining the underlying causes of the observed dichotomy. The core of the Moravian sample lies in the 9th Century with an overlap into the first third of the 10th Century, while the Bohemian data set is, on average, younger (Table 1). Moreover, the pattern outlined above in Bohemia has some exceptions, especially in the most ancient samples.
Nitrogen isotopic values and human–faunal offsets were comparable to other Central European medieval populations. The dietary proportion of animal products consumed by the Prague castle population appears to be comparable to those of Moravian centers (Kaupová et al. 2016). Interestingly, these values are not extraordinarily high in the central European context, even though the presence of the highest Bohemian elites of that time is supposed. Similar human-faunal offsets were for example observed at the German rural site of Dalheim (Olsen et al. 2016). The δ15N values of the Levý Hradec burial sites and Milady Horákové appear to be located in the lower part of this variation, comparable with the rural medieval sites of Josefov (Moravia) or Gruczno (Poland) but also with the German urban site of Magdeburg (Kaupová et al. 2016; Knipper et al. 2015; Reitsema et al. 2017), suggesting significant restriction to diet of animal origin. The Střešovice sample showed intermediate values comparable with population samples from the Polish medieval center of Kaldus (Reitsema et al. 2017).
Due to a number of factors, such as the small size of the animal data sets in most of the studies, as well as the untypical demography of some of the sites included in this study (see above), or the inevitable simplification of data interpretation caused by comparing human-faunal offsets, this comparison should not be over interpreted, However, the very fact that the human isotopic data explored in this study cover the full range observed in the early medieval populations of Central Europe (from the main centers of state formation to rural settlements) further confirms our finding of the profound socio-economic stratification of these centers.
Naturally, the integration of the isotopic data from the ducal family into the wider European context is even more difficult, given the relative scarcity of any comparative data. There are only a few studies of the isotopic data of the highest elites across the Middle Ages (ca. 600 to 1500 AD), spanning from Spain to England (Jiménez-Brobeil et al. 2016; Knipper et al. 2015; Lamb et al. 2014). However, several basic results occur repeatedly in these studies, suggesting the existence of some norm for elite dietary behavior. First, the nitrogen isotopic values of the concerned individuals are, as a rule higher, than the maximal value measured in comparative samples of non-elite (either urban or rural) populations. This is true for all the above-mentioned studies (although, for the Spain royal family, only when Muslim sites with distinct dietary behavior are excluded). However, the human–faunal nitrogen isotopic offsets as well as isotopic differences between ‘dynastic’ samples and the comparative non-elite data sets are higher in late medieval samples (king Richard III and Spain royal family) from the 14th to 15th Century (being in general higher than 5‰ for Δ15Ndynastic-fauna and higher than 2‰ for Δ15Ndynastic non-elite) than in samples dated before 1000 AD (Merovingian ‘Prince of Morken’ and the 10th-Century Queen Edith Table 7). The routinely accepted explanation of this trend is the significant influence of fasting prescriptions during the late Middle Ages (Jiménez-Brobeil et al. 2016; Lamb et al. 2014). Given that meat was prohibited by the Church for a substantial part of the year, fish are assumed to have become a highly priced foodstuff that was primarily available to people of the upper social classes (Knipper et al. 2015; Müldner and Richards 2007b; Salamon et al. 2008) (but see the Czech local historical sources mentioning the regular consumption of salted fish by the poor around 1100 AD; (Hrdina et al. 2005; Mlíkovský 2003). Thus, the isotopic values of the Přemyslids become especially interesting. Duke Spytihněv, his wife, as well as the unknown duke and duchess buried in St. Vitus rotunda (PCV01 and PCV02), showed similar nitrogen isotopic offsets as ‘Prince of Morken’ and Queen Edith: their nitrogen isotopic offsets to faunal samples varied from 3.3 to 4.6‰ and the nitrogen isotopic differences compared with hinterland population sample ranged from 0.6 to 1.9‰. Moreover, as stated previously, their nitrogen isotopic values fell, in contrast to other studies, within (in case of Spytihněv I, only slightly above) the isotopic variation of the hinterland sample. By contrast, the group of individuals buried in St. George basilica showed isotopic offsets that are comparable to late medieval elites (4.8–7.1 for human–faunal offsets; 2.1–4.4 compared with the hinterland sample). This is especially true of Duke Vratislav I and unknown duke PCG02. The last comparative datasets represented by two Dutch counts of the 12th century. These show isotopic values resembling Spytihnev and PCV01 (Lanting and van der Plicht 1995–1996). However, due to the absence of comparative data from either human or faunal samples, the value of this comparison is limited.
The carbon isotopic data are more difficult to compare in a wider European context, because the consumption of millet in some populations and of marine products in others can cause the comparatively increased isotopic offsets. However, special attention should be paid to the consumption of millet observed in the four individuals buried in St. Mary’s church and St. Vitus rotunda. As observed previously (Kaupová et al. 2016), millet was consumed by the elite classes of the neighboring Great Moravian Empire. During the 9th Century, Great Moravia is documented to have strongly influenced the annexed territory of Bohemia (Berend et al. 2013; Bláhová et al. 2008; Charvát 2010). Subsequently, the dietary behavior of the oldest generation of Přemyslids, who, as vassals of Moravians, established their court in Prague, might have been influenced by the Great Moravian nobility.
Although the influence of individual dietary preferences should not be underestimated, the comparison of the Přemyslid isotopic values in a wider context suggests that the high dietary variation observed within the relatively brief period (potentially even within a single generation in the case of brothers Spytihněv I and Vratislav I) is the evidence of the rapidly changing position of Přemyslids during the formation of the Bohemian state structure or, alternatively, the sharp increase in the influence of the Church on elite dietary behavior.
Conclusion
According to the present data, the early medieval populations of power centers in Bohemia had a terrestrial diet, without a substantial input of freshwater products and with a varying proportion of C4 plants. The comparison of dietary behavior of castle elites with lower socio-economic groups inhabiting both castles and their hinterlands clearly documents the unequal access of these population groups to high-quality foodstuff, namely animal products. Moreover, millet, which is generally viewed as a crop of lower socio-economic groups, was a more notable part of the diet of the lower socio-economic ranks of both castles and hinterland.
The substantial dietary variation observed between males, but not between females from different non-castle burial sites suggests, in accordance with previous studies, that male dietary behavior is a more sensitive indicator of socio-economic differences. Alternatively, this finding can be seen in the light of anthropological results showing that the almost complete absence of adult males at some of the sites was accompanied by a low proportion of animal products in the diet of the males present. The final place of a part of the male population has yet to be found but might be linked to their specific function in relation to close castles. Regardless of the underlying causes, the present data suggest the existence of specialization or socio-economic stratification both within the power centers and their immediate hinterland.
The substantial dietary variation observed within the small sample of dynastic graves appears to illustrate the rise of the Přemyslids to the very summit of the social hierarchy and opens new questions concerning the true identities of these oldest generations of this princely dynasty.
This study is, to the best of the authors’ knowledge, the first application of Bayesian mixing models to the early medieval European population. In future, it should be possible to further refine the estimates by (i) including more dietary proxies (such as the isotopic values of sulfur or individual amino acids as well as the carbon isotopic values of bone carbonate); (ii) setting-up baseline values for more isotopically distinct food groups of both plant and terrestrial origin; and (iii) enlarging our knowledge of further model parameters such as offsets between measured proxies and food fractions. Moreover, other analyses such as the isotopic analysis of residential mobility may shed light on some of the unexpected dietary patterns observed in this study.
References
Acsádi G, Nemeskéri J (1970) History of human life span and mortality. Akadémiai Kiadó, Budapest
Ambrose SH, Norr L (1993) Experimental evidence for the relationship of the carbon isotope ratios of whole diet and dietary protein to those of bone collagen and carbonate. In: Lambert JB, Grupe G (eds) Prehistoric human bone. Springer-Verlag, Berlin, pp 1–37
Ambrose SH, Butler BM, Hanson DB, Hunter-Anderson RL, Krueger HW (1997) Stable isotopic analysis of human diet in the Marianas archipelago, western Pacific. Am J Phys Anthropol 104:343–361
Adamson MW (2004) Food in medieval times. Greenwood Publishing Group, Westport
Bardsley S (2007) Women’s roles in the middle ages. Greenwood Publishing Group, Westport
Barrett JH, Richards MP (2004) Identity, gender, religion and economy: new isotope and radiocarbon evidence for marine resource intensification in early historic Orkney, Scotland, UK. Eur J Archaeol 7:2497–2271
Barrett JH, Locker AM, Roberts CM (2004) The origins of intensive marine fishing in medieval Europe: the English evidence. Proc R Soc Lond B Biol Sci 271:2417–2421
Beranová M (2004) Výživa a hladomory v Čechách na počátku 2. tisíciletí. Slovenská Archeológia 52:121–138
Berend N, Urbańczyk P, Wiszewski P (2013) Central Europe in the High Middle Ages: Bohemia, Hungary and Poland, c. 900–c. 1300. Cambridge University Press, Cambridge
Bláhová M, Frolík J, Profantová N (2008) Velké dějiny zemí Koruny české. Svazek I. do roku 1197. Paseka, Praha - Litomyšl
Bocherens H (1992) Biogéochimie isotopique ( 13C, 15N, 18O) et paléontologie des vertébrés: applications à l'étude des réseaux trophiques révolus et des paléoenvironnements. Dissertation, Université Paris VI
Bocherens H, Drucker D (2003) Trophic level isotopic enrichment of carbon and nitrogen in bone collagen: case studies from recent and ancient terrestrial ecosystems. Int J Osteoarchaeol 13:46–53
Bodri L, Čermák V (1995) Climate changes of the last millennium inferred from borehole temperatures: results from the Czech Republic—part I. Glob Planet Change 11:111–125
Brázdil R, Pfister C, Wanner H, Von Storch H, Luterbacher J (2005) Historical climatology in Europe–the state of the art. Clim Chang 70:363–430
Bruzek J (2002) A method for visual determination of sex, using the human hip bone. Am J Phys Anthropol 117:157–168
Charvát P (2010) The emergence of the bohemian state. Brill, Boston
DeNiro MJ (1985) Postmortem preservation and alteration of in vivo bone collagen isotope ratios in relation to palaeodietary reconstruction. Nature 317:806–809
DeNiro MJ, Epstein S (1978) Influence of diet on the distribution of carbon isotopes in animals. Geochim Cosmochim Acta 42:495–506
DeNiro MJ, Epstein S (1981) Influence of diet on the distribution of nitrogen isotopes in animals. Geochim Cosmochim Acta 45:341–351
Dufour E, Bocherens H, Mariotti A (1999) Palaeodietary implications of isotopic variability in Eurasian lacustrine fish. J Archaeol Sci 26:617–627
Ferembach D, Schwidezky I, Stoukal M (1980) Recommendation for age and sex diagnoses of skeletons. J Hum Evol 9:517–549
Fernandes R, Nadeau M-J, Grootes PM (2012a) Macronutrient-based model for dietary carbon routing in bone collagen and bioapatite. Archaeol Anthropol Sci 4:291–301
Fernandes R, Rinne C, Grootes PM, Nadeau M-J (2012b) Revisiting the chronology ofNorthern German monumentality sites: preliminary results. In: Hinz M, Müller J (eds) Frühe Monumentalität und soziale Differenzierung. Rudolf Habelt, Bonn, pp 87–103
Fernandes R, Millard AR, Brabec M, Nadeau MJ, Grootes P (2014) Food reconstruction using isotopic transferred signals (FRUITS): a Bayesian model for diet reconstruction. PLoS One 9:e87436. https://doi.org/10.1371/journal.pone.0087436
Fernandes R, Grootes P, Nadeau MJ, Nehlich O (2015) Quantitative diet reconstruction of a Neolithic population using a Bayesian mixing model (FRUITS): the case study of Ostorf (Germany). Am J Phys Anthropol 158:325–340
Fornaciari G (2008) Food and disease at the renaissance courts of Naples and Florence: a paleonutritional study. Appetite 51:10–14
Fraser R, Bogaard A, Schäfer M, Arbogast R, Heaton TH (2013) Integrating botanical, faunal and human stable carbon and nitrogen isotope values to reconstruct land use and palaeodiet at LBK Vaihingen an der Enz, Baden-Württemberg. World Archaeol 45:492–517
Frolík J (2005) Hroby přemyslovských knížat na Pražském hradě. In: Tomková K (ed) Castrum Pragense 7- Pohřbívání na Pražském hradě a jeho předpolích. Díl I.1. Archeologický ústav AV ČR, Prague, pp 25–46
Frolík J (2016) Pohřebiště u kostela Panny Marie a na II. nádvoří Pražského hradu. Archeologický ústav AV ČR, Prague
Frolík J, Smetánka Z (1997) Archeologie na Pražském hradě. Paseka, Praha – Litomyšl
Frolíková-Kaliszová D (2014) Pohřebiště z 10. století v Praze-Střešovicích – předběžná zpráva. Archaeologia historica 39:315–329
Fuller BT, De Cupere B, Marinova E, Van Neer W, Waelkens M, Richards MP (2012a) Isotopic reconstruction of human diet and animal husbandry practices during the Classical-Hellenistic, imperial, and Byzantine periods at Sagalassos, Turkey. Am J Phys Anthropol 149:157–171
Fuller BT, Müldner G, Van Neer W, Ervynck A, Richards MP (2012b) Carbon and nitrogen stable isotope ratio analysis of freshwater, brackish and marine fish from Belgian archaeological sites (1st and 2nd millennium AD). J Anal At Spectrom 27:807–820
Hakenbeck S (2013) Potential and limitations of isotope analysis in early medieval archaeology. Post-Class Archaeol 3:109–125
Hakenbeck S, McManus E, Geisler H, Grupe G, O'Connell T (2010) Diet and mobility in early medieval Bavaria: a study of carbon and nitrogen stable isotopes. Am J Phys Anthropol 143:235–249
Hamilton J, Hedges RE, Robinson M (2009) Rooting for pigfruit: pig feeding in Neolithic and Iron Age Britain compared. Antiquity 83:998–1011
Hammond C, O’Connor T (2013) Pig diet in medieval York: carbon and nitrogen stable isotopes. Archaeol Anthropol Sci 5:123–127
Hedges RE, Reynard LM (2007) Nitrogen isotopes and the trophic level of humans in archaeology. J Archaeol Sci 34:1240–1251
Halffman CM, Velemínský P (2015) Stable isotope evidence for diet in early medieval Great Moravia (Czech Republic). J Archaeol Sci Rep 2:1–8
Hrdina K, Bláhová M, Třeštík D, Kopal P (2005) Kosmova kronika česká (translation from Latin). Paseka, Prague
Jiménez-Brobeil S, Laffranchi Z, Maroto R, Sánchez FL, Huertas AD (2016) How royals feasted in the court of Pedro I of Castile: a contribution of stable isotope study to medieval history. J Archaeol Sci Rep 10:424–430
Katzenberg MA, Weber A (1999) Stable isotope ecology and palaeodiet in the Lake Baikal region of Siberia. J Archaeol Sci 26:651–659
Kaupová S, Velemínský P, Herrscher E, Sládek V, Macháček J, Poláček L, Brůžek J (In Press) Diet in transitory society: isotopic analysis of medieval population of Central Europe (ninth–eleventh century AD, Czech Republic). Archaeol Anthropol Sci:1–20 https://doi.org/10.1007/s12520-016-0427-8
Kjellström A, Storå J, Possnert G, Linderholm A (2009) Dietary patterns and social structures in medieval Sigtuna, Sweden, as reflected in stable isotope values in human skeletal remains. J Archaeol Sci 36:2689–2699
Knipper C, Peters D, Meyer C, Maurer A-F, Muhl A, Schöne BR, Alt KW (2013) Dietary reconstruction in migration period Central Germany: a carbon and nitrogen isotope study. Archaeol Anthropol Sci 5:17–35
Knipper C, Held P, Fecher M, Nicklisch N, Meyer C, Schreiber H, Zich B, Metzner-Nebelsick C, Hubensack V, Hansen L, Nieveler E, Alt KW (2015) Superior in life—superior in death: dietary distinction of central European prehistoric and medieval elites. Curr Anthropol 56:579–589
Knipper C, Pichler SL, Rissanen H, Stopp B, Kühn M, Spichtig N, Röder B, Schibler J, Lassau G, Alt KW (2016) What is on the menu in a Celtic town? Iron age diet reconstructed at Basel-Gasfabrik, Switzerland. Archaeol Anthropol Sci 9:1307–1326
Kočár P, Čech P, Kozáková R, Kočárová R (2010) Environment and economy of the early medieval settlement in Žatec. Interdiscip Archaeol 1:45–60
Klápště J (2005) Proměna českých zemí ve středověku. Nakladatelství Lidové noviny, Prague
Kyselý R (2000) Archeozoologický rozbor materiálu z lokality Rubín a celkový pohled na zvířata doby hradištní. Památky archeologické 91:155–200
Kyselý R (2003) Ryby (Pisces) a obojživelníci (Amphibia) z raně středověkého hradu Stará Boleslav (střední Čechy). In: Boháčová I (ed) Stará Boleslav. Přemyslovský hrad v raném středověku. Archeologický ústav AV ČR, Praha, pp 345–346
Kyselý R (2005) Archeologické doklady divokých savců na území ČR v období od neolitu po novověk. Lynx 36:55–101
Lamb AL, Evans JE, Buckley R, Appleby J (2014) Multi-isotope analysis demonstrates significant lifestyle changes in King Richard III. J Archaeol Sci 50:559–565
Lanting JN, van der Plicht J (1995-1996) Wat hebben Floris V, skelet Swifterbant S2 en visotters gemeen? Palaeohistoria 37(38):491–519
Le Huray JD, Schutkowski H (2005) Diet and social status during the La Tène period in Bohemia: carbon and nitrogen stable isotope analysis of bone collagen from Kutná Hora-Karlov and Radovesice. J Anthropol Archaeol 24:135–147
Lee-Thorp JA (2008) On isotopes and old bones. Archaeometry 50:925–950
Lightfoot E, Šlaus M, O'Connell T (2012) Changing cultures, changing cuisines: cultural transitions and dietary change in Iron Age, Roman, and Early Medieval Croatia. Am J Phys Anthropol 148:543–556
Longin R (1971) New method of collagen extraction for radiocarbon dating. Nature 230:241–242
Lovejoy CO, Meindl RS, Pryzbeck TR, Mensforth RP (1985) Chronological metamorphosis of the auricular surface of the ilium: a new method for the determination of adult skeletal age at death. Am J Phys Anthropol 68:15–28
Minagawa M, Wada E (1984) Stepwise enrichment of 15N along food chains: further evidence and the relation between δ15N and animal age. Geochim Cosmochim Acta 48:1135–1140
Maurer AF, Person A, Zazzo A, Sebilo M, Balter V, Le Cornec F, Zeitoun V, Dufour E, Schmidt A, de Rafélis M, Ségalen L, Bedaux R (2017) Geochemical identity of pre-Dogon and Dogon populations at Bandiagara (Mali, 11th–20th cent. AD). J Archaeol Sci Rep 14:289–301
McGlynn G (2007) Using 13C, 15N, and 18O stable isotope analysis of human bone tissue to identify transhumance, high altitude habitation and reconstruct palaeodiet for the early medieval alpine population at Volders, Austria. Dissertation, LMU München
McKern TW, Stewart TD (1957) Skeletal age changes in young American males analysed from the standpoint of age identification. In: Technical report EP 45. Quartermaster Research and Development Center, Environmental Protection Research Division. Natick, Massachusetts, pp 89–97
Mlíkovský J (2003) Zvířata a jejich role na raně středověkém hradě Stará Boleslav (střední Čechy). In: Boháčová I (ed) Stará Boleslav. Přemyslovský hrad v raném středověku. Archeologický ústav AV ČR, Praha, pp 347–365
Müldner G (2009) Investigating medieval diet and society by stable isotope analysis of human bone. In: Gilchrist R, Reynolds A (eds) Reflections: 50 years of medieval archaeology. Maney, Leeds, pp 327–346
Müldner G, Richards MP (2005) Fast or feast: reconstructing diet in later medieval England by stable isotope analysis. J Archaeol Sci 32:39–48
Müldner G, Richards MP (2007a) Diet and diversity at later medieval Fishergate: the isotopic evidence. Am J Phys Anthropol 134:162–174
Müldner G, Richards MP (2007b) Stable isotope evidence for 1500 years of human diet at the city of York, UK. Am J Phys Anthropol 133:682–697
Murail P, Bruzek J, Houët F, Cunha E (2005) DSP: a tool for probabilistic sex diagnosis using worldwide variability in hip-bone measurements. Bull Mem Soc Anthropol Paris 17:167–176
Newsome SD, Phillips DL, Culleton BJ, Guilderson TP, Koch PL (2004) Dietary reconstruction of an early to middle Holocene human population from the central California coast: insights from advanced stable isotope mixing models. J Archaeol Sci 31:1101–1115
O'Connell TC, Kneale CJ, Tasevska N, Kuhnle GG (2012) The diet-body offset in human nitrogen isotopic values: a controlled dietary study. Am J Phys Anthropol 149:426–434
Olsen KC, White CD, Longstaffe FJ, Rühli FJ, Warinner C, Salazar-García DC (In Press) Isotopic anthropology of rural German medieval diet: intra- and inter-population variability. Archaeol Anthropol Sci. https://doi.org/10.1007/s12520-016-0432-y
Parnell AC, Inger R, Bearhop S, Jackson AL (2010) Source partitioning using stable isotopes: coping with too much variation. PLoS One 5:e9672. https://doi.org/10.1371/journal.pone.0009672
Pestle WJ, Torres-Rouff C, Hubbe M (2016) Modeling diet in times of change: the case of Quitor, San Pedro de Atacama, Chile. J Archaeol Sci Rep 7:82–93
Phillips DL (2001) Mixing models in analyses of diet using multiple stable isotopes: a critique. Oecologia 127:166–170
Piličiauskas G, Jankauskas R, Piličiauskienė G, Dupras T (2017) Reconstructing Subneolithic and Neolithic diets of the inhabitants of the SE Baltic coast (3100–2500 cal BC) using stable isotope analysis. Archaeol Anthropol Sci 9:1421–1437
Profantová N (2005) Poznámky k meči z hrobu bojovníka na III. nádvoří Pražského hradu – Bemerkungen zum Schwert aus dem Kriegergrab auf dem III. Burghof der Prager Burg. In: Tomková K (ed) Castrum Pragense 7 – Pohřbívání na Pražském hradě a jeho předpolích. Díl I.1. Archeologický ústav AV ČR, Praha, pp 307–310
Rapoport S (1929) On the Early Slavs. The Narrative of Ibrahim-Ibn-Yakub. Slav East Eur Rev 8:331–341
Reitsema LJ, Vercellotti G (2012) Stable isotope evidence for sex-and status-based variations in diet and life history at medieval Trino Vercellese, Italy. Am J Phys Anthropol 148:589–600
Reitsema LJ, Crews DE, Polcyn M (2010) Preliminary evidence for medieval Polish diet from carbon and nitrogen stable isotopes. J Archaeol Sci 37:1413–1423
Reitsema LJ, Kozłowski T, Makowiecki D (2013) Human–environment interactions in medieval Poland: a perspective from the analysis of faunal stable isotope ratios. J Archaeol Sci 40:3636–3646
Reitsema LJ, Kozłowski T, Crews DE, Katzenberg MA, Chudziak W (2017) Resilience and local dietary adaptation in rural Poland, 1000–1400 CE. J Anthropol Archaeol 45:38–52
Richards MP, Fuller BT, Molleson T (2006) Stable isotope palaeodietary study of humans and fauna from the multi-period (Iron Age, Viking and Late Medieval) site of Newark Bay, Orkney. J Archaeol Sci 33:122–131
Rutgers L, Van Strydonck M, Boudin M, Van der Linde C (2009) Stable isotope data from the early Christian catacombs of ancient Rome: new insights into the dietary habits of Rome’s early Christians. J Archaeol Sci 36:1127–1134
Salamon M, Coppa A, McCormick M, Rubini M, Vargiu R, Tuross N (2008) The consilience of historical and isotopic approaches in reconstructing the medieval Mediterranean diet. J Archaeol Sci 35:1667–1672
Schmitt A (2005) Une nouvelle méthode pour estimer l’âge au décès des adultes à partir de la surface sacro-pelvienne iliaque. Bull Mem Soc Anthropol Paris 17:89–101
Schmitt A (2008) Une nouvelle méthode pour discriminer les individus décédés avant ou après 40 ans à partir de la symphyse pubienne. J Med Leg Droit Med 51:15–24
Schoeninger MJ, DeNiro MJ (1984) Nitrogen and carbon isotopic composition of bone collagen from marine and terrestrial animals. Geochim Cosmochim Acta 48:625–639
Sjögren K-G (2017) Modeling middle Neolithic funnel beaker diet on Falbygden, Sweden. J Archaeol Sci Rep 12:295–306
Sommer P, Třeštík D, Žemlička J (2009) Přemyslovci. Budování českého státu. Nakladatelství Lidové Noviny, Prague
Stránská P (2012a) Anthropological examination of the skeletal material from the Milady Horákové cemetery. Unpublished report, Archeologický ústav Akademie věd Praha
Stránská P (2012b) Pohřebiště na Levém Hradci a jeho předpolí z pohledu antropologie. In: Tomková K (ed) Levý Hradec v zrcadle archeologických výzkumů I. Archeologický ústav AV ČR, Prague, pp 354–380
Stránská P (2014) Antropologická analýza lidských kostrových pozůstatků z raně středověkého pohřebiště v Praze-Střešovicích, poloha Triangl. Archaeologia. For Hist 39:331–345
Svoboda J, Cílek V, Vašků Z (2003) Velká kniha o klimatu zemí koruny české. Regia, Prague
Teschler-Nicola M, Novotny F, Spannagl-Steiner M, Stadler P, Prohaska T, Irrgeher J, Däubl B, Haring E, Rumpelmayr K, Wild EM (2015) The early mediaeval manorial estate of Gars/Thunau, Lower Austria: an enclave of endemic tuberculosis? Tuberculosis 95:S51–S59
Tomková K (2012) Levý Hradec v zrcadle archeologických výzkumů I. Archeologický ústav AV ČR, Prague
Tomková K (2013) Urbs Praga und Levý Hradec - von Sklaven zu Fürsten. Beiträge zur Ur- und Frühgeschichte Mitteleuropas 70:43–55
Van Klinken GJ (1999) Bone collagen quality indicators for palaeodietary and radiocarbon measurements. J Archaeol Sci 26:687–695
Vika E, Theodoropoulou T (2012) Re-investigating fish consumption in Greek antiquity: results from δ13C and δ15N analysis from fish bone collagen. J Archaeol Sci 39:1618–1627
Vlček E (1997) Nejstarší Přemyslovci Fyzické osobnosti českých panovníků. Atlas kosterních pozůstatků prvních sedmi historicky známých generací Přemyslovců s podrobným komentářem a historickými poznámkami. Vesmír, Prague
Waters-Rist A, Katzenberg M (2010) The effect of growth on stable nitrogen isotope ratios in subadult bone collagen. Int J Osteoarchaeol 20:172–191
Winklerová D (2011) Zooarchaeological and archaeobotanical indicators for aspects of diet in medieval Kingdom of Bohemia. In: Klápště J, Sommer P (eds) Ruralia VIII.: processing, storage, distribution of food: food in the medieval rural environment. Brepols, Turnhout, pp 421–429
Yoder C (2012) Let them eat cake? Status-based differences in diet in medieval Denmark. J Archaeol Sci 39:1183–1193
Acknowledgements
We would like to thank the following institutions for financial support: Grant Agency of the Czech Republic (Grant number: 14-36938G), Ministry of Culture of the Czech Republic (Grant number: P15/01IG-KA and DKRVO 2017/18 and 2018/17, National Museum, 00023272). We thank International Science Editing for the English language editing. Also many thanks are due to Olga Trojánková for species determination of animal bones.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article

Cite this article
Kaupová, S., Velemínský, P., Stránská, P. et al. Dukes, elites, and commoners: dietary reconstruction of the early medieval population of Bohemia (9th–11th Century AD, Czech Republic). Archaeol Anthropol Sci 11, 1887–1909 (2019). https://doi.org/10.1007/s12520-018-0640-8
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s12520-018-0640-8