
Abstract
Six geological studied sections from the Ouled Maallah region (Dahra) allow the identification of lacustrine deposits, sandwiched between Messinian gypsum and Mio-Pliocene blue marls. The gypsum followed by white micritic limestones deposits is correlated with the “Terminal Carbonated Complex” subsequent to the coral platforms (Oran, Melilla, Sorbas). Correlated to the Lago Mare 1 episode, which is slightly older than 5.60 Ma, the lacustrine deposits tend to cover a large extension area, both on the southern margin of the Dahra and on the Beni Chougrane northern slopes. Subsequently, the early Messinian-Zanclean deposits fossilize the Messinian Erosional Surface and indicate the post-Messinian Crisis marine reflooding. They are characterized by two microfossiliferous successions: (i) the presence of Sphaeroidinellopsis subdehiscens, Globorotalia margaritae (foraminifers), Ceratolithus acutus, Reticulofenestra zancleana (calcareous nannofossils) indicating the latest Messinian to earliest Zanclean, followed by an association with (ii) G. puncticulata, G. bononiensis, G. aff. crassaformis and Helicosphaera sellii, Discoaster asymmetricus, and D. tamalis, indicating a late Zanclean age up to an early Piacenzian one. The regional correlations reveal large subsiding sedimentary depocentres built along the Chelif Valley. Likewise, the Mio-Pliocene sedimentation seals folded tectonics active during the latest Messinian, individualized by the tilted gypsum and Lago Mare deposits of Ouled Maallah. This deformation has resulted from gravity tectonics responsible for the “olistolite” made or reworked materials within the geological series located at the Chelif Valley and the Beni Chougrane Mounts. This testifies to a large-scale, compressive deformation that would correspond to a restructuring of paleogeographic spaces associated with the Tellian Massif uplifts.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
For the past 20 years, stratigraphic research based on the micropaleontology of Neogene deposits in the Lower Chelif Basin has experienced a certain decline, due to the lack of thematic programs or a change of perspective, targeting specific aspects depending on fossil organisms, more or less suitable, among other items, for the paleoenvironment reconstructions. These aspects have made much progress, for example, our knowledge of the history of the Messinian Salinity Crisis in this region and contributed effectively to the wide understanding context of this crisis in the Mediterranean (Arambourg 1927; Rouchy 1982a, b; Moissette 1988; Saint Martin 1990; Poignant and Moissette 1992; Chikhi 1992; Cornée et al. 1994a; Mansour 2004; Haddadi-Hamdane 1996; Hebib 2014).
The purpose of this study is to take up this theme in order to diversify the dating methods having as object the precision and calibration of the biostratigraphic scales of the geological series of the northwestern basins of Algeria. This will provide different dating tools, necessary in particular for synthesis work or for editing the geological map at different scales or for their revision.
The planktonic foraminifera biozonation has advanced our knowledge of the Miocene in this region, making it possible to design a stratigraphic scheme that is at least acceptable but not sufficient (Perrodon 1957; Mazzola 1971; Belkebir 1986; Belkebir et al. 1996; Bessedik et al. 2002; Belhadji et al. 2008; Belkebir et al. 2008). Other microfossils such as calcareous nannofossils are under study (Mansouri 2001; Mansouri et al. 2008; doctorate in progress); they consolidate and/or clarify this biozonation.
The systematic and constantly renewed study of sectors and sections in a vast territory, such as these intramontane Tellian basins, allows a better understanding of their specificity and their morphological diversity. Within this framework and from the exploration of new sectors, we target two objectives: (i) lithostratigraphic recording in the Messinian (post-gypsum) and the Pliocene, (ii) the search for bioevents (planktonic foraminifera and calcareous nannofossils) for calibration and correlation. These calibrations and correlations were carried out on the Mediterranean scale and in particular with those based on the dating of radiometric and/or astronomical events (Gautier et al. 1994; Berggren et al. 1995; Cunningham et al. 1997; Cornée et al. 2004; Lourens et al. 2005; Backman et al. 2012; Clauzon et al. 2015a; Benzina et al. 2019; Lirer et al. 2019).
In this context, the biozonation scale can be constantly improved, especially when it comes to integrating the calibration tools between bioevents of species from the same organism group and/or between different microfossil scales. This method allows one to refine biostratigraphy in general and to appreciate paleobiodiversity and regional correlations.
Geologic context
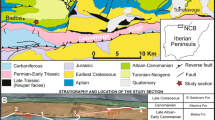
Intramontane, the Lower Chelif Basin, shares borders to the south by the Southern Tell Mountains (Tessala, Ouled Ali, Beni Chougrane, Ouarsenis). In the North are the Tellian coastal massifs of Murdjadjo, Orousses, and Dahra, which underline its Northern and Northeastern margin. This basin extends over a length of approximately 250 km and 35 km wide, as an ENE-WSW oriented lozengic form, made up of plains, the M’leta in the West, the Habra in the center, and the Chelif plain in the East (Fig. 1: A and B). It is the place of a significant Neogene and Quaternary sedimentation, which can reach 4800 or even 5000 m in thickness (Perrodon 1957; Meghraoui et al. 1988; Harbi et al. 1999; Arab et al. 2015).

Location map of the Dahra Massif and Lower Chelif Basin. A: Global West-Mediterranean location; B: North and South Tellian massifs bordering the Intramontane Lower Chelif Basin (Map extracted from “Global Mapper v18.2,” Projection: SRTM Worldwide Elevation Data (3-arc-second Resolution); White square: studied zone; C: Position of studied sections: 1: Moul Settara; 2a: Azaizia (a) and addition, 2b: Azaizia (b); 3: Aïn Yacoub; 4: Oued Tarhia; 5: Garbous Ben Saada
Through its history, this region, located on the north and northeast margin of the western Chelif Plain, often echoes during the Neogene an active tectonics (Anderson 1936; Perrodon 1957; Delteil 1974; Guardia 1975; Fenet 1975; Meghraoui 1982; Meghraoui et al. 1986; Leprêtre et al. 2018). It witnessed the Alpine orogenic activity whose neotectonic impact is still felt today (Meghraoui et al. 1988; Harbi et al. 1999; Derder et al. 2011, 2013; Benbakhti et al. 2018; Abbouda et al. 2019). It manifests itself by a structure in “tilted blocks,” known from the end of the Cretaceous (Brive 1897; Anderson 1936; Badji et al. 2015; Leprêtre et al. 2018: Fig. 2a). This Tellian morphology is constantly linked to and called upon by the global dynamics of the African plate and its displacement towards the northwest (2 cm/year), resulting in the closure to the north of the Neogene sedimentary basins (Meghraoui et al. 1988; Derder et al. 2011, 2013; Badji et al. 2015; Leprêtre et al. 2018). Such a dynamics is also deep, in view of data obtained from magmatism (Lucas 1942; Sadran 1952, 1958; Mattauer 1958; Polvèche 1959). The magmatic activity dates from the Middle Miocene to Quaternary (Megartsi 1985; El Azzouzi et al. 2003). It manifests itself intermittently first calco-alkaline then alkaline, as ash or lava forms, in the Neogene geological series including the Lower Chelif Basin and its margins (Brive 1897; Gourinard 1958; Delfaud et al. 1973; Megartsi 1985; Guardia 1975; Bellon et al. 1984; Thomas 1985; Neurdin-Trescartes 1992; Ameur-Chehbeur 1988; Louni-Hacini et al. 1995).

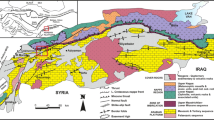
Geology and structure of the Dahra Massif. A. NNW-SSE structural section in “tilted blocks,” going from the passive margin to the southern Tell, passing through the Dahra and the Chelif basins (Leprêtre et al. 2018). B. Geological and structural map of Dahra (modified from: Perrodon 1957; Meghraoui et al. 1988; Belkebir et al. 2002; Bessedik et al. 2002)
The stratigraphy of the geological series of the Dahra Massif and the surrounding areas began very early with the mollusks, having made it possible to carry out the first stratigraphic divisions of the Neogene (Pomel 1892; Ficheur 1896; Brive 1897, 1920; Dalloni 1915). Subsequent works (Anderson 1936; Gourinard 1952, 1958; Perrodon 1957) contributed substantially to the knowledge of the stratigraphy of this region.
Miocene deposits are organized into two major sequences (Delfaud et al. 1973; Thomas 1985; Neurdin-Trescartes 1992): the first of the Lower Miocene age with detritic (conglomerates, sandstones) to fine sedimentation (gray to variegated clay), contemporary with a compressive tectonics, having controlled the establishment of the Tellian thrusts or "first post-thrust cycle" (Perrodon 1957; Mattauer 1958). This sedimentation bypasses, in a “scarf,” the Dahra Massif. The revision of the age of these deposits made it possible to recognize, at the oldest, the terminal Burdigalian followed by the Langhian and the Serravallian stages (Bessedik et al. 2002). Recent biostratigraphic and tectonic data (Meghraoui 1982; Belkebir and Anglada 1985; Meghraoui et al. 1988; Belkebir et al. 1996, 2002, 2008; Bessedik et al. 1997, 2002; Belhadji et al. 2008; Mansouri et al. 2008) are plotted on the map of geological surveys (Anderson et al. in: Perrodon 1957), allowing some improvements (Fig. 2B).
At the same time, continental sedimentation took place from the upper Serravallian with Bouhanifia and Tafna formations (Guardia 1975, 1976; Ouda and Ameur 1978; Ameur-Chehbeur 1988; Bessedik et al. 2002; Belkebir et al. 1996).
Generally controlled by a transtensive tectonics, the second major sequence belongs to the second post-thrust cycle (Meghraoui 1982, 1988; Perrodon 1957), which is characterized by marly deposits of Tortonian and Messinian ages (Perrodon 1957; Thomas 1985; Neurdin-Trescartes 1992, 1995; Belkebir et al. 1996). Upstream of the southern and eastern edges of the Lower Chelif Basin, major contemporary continental spreadings (Vallesian, Turolian) are organized at the same time (Guardia 1975; Jaeger 1977; Ameur-Chehbeur 1988, 1992; Bessedik et al. 1997; Mahboubi 2014; Mahboubi et al. 2015).
The Messinian deposits overlie this marine and continental sedimentation: marls and diatomaceous marls, evaporites, post-evaporitic marine or lagoonal (Anderson 1936; Perrodon 1957; Rouchy 1982a, b; Thomas 1985; Saint Martin 1990; Neurdin-Trescartes 1992, 1995; Mansour and Saint Martin 1999; Mansour 2004; Cornée et al. 2004, 2006; Rouchy et al. 2007).
Evaporites (gypsum), witnesses of the Messinian Salinity Crisis, reach in some places 40 m (two gypsum bars) or even 250 to 300 m in thickness. Their distribution is well-known on the northern margin of the Lower Chelif Basin and more particularly on the Dahra southern slope (Fig. 3). Indeed, the Ouled Maallah region (studied area) has recorded great thicknes.

Isopach map of Messinian areas (gypsum: red color; Lago Mare: asterisk) and Pliocene blue marls in the Lower Chelif Basin (Perrodon 1957)
In the center of the Chelif Basin, the transition between the Messinian and the Pliocene seems, according to Perrodon (1957: p. 204), marked by marine marls evolving to lacustrine environments, with fluvial currents in some places.
The transgressive Pliocene cycle on the previous deposits is well-represented (Fig. 3) in the Chelif Basin (Brive 1897; Anderson 1936; Perrodon 1957; Mazzola 1971; Fenet and Irr 1973; Belkebir 1986; Belkebir and Anglada 1985; Thomas 1985; Neurdin-Trescartes 1992; Belhadji 2005; Rouchy et al. 2007; Atif et al. 2008; Abbouda et al. 2019). It displays large masses of blue marls up to 750 to 800 m in thickness (Perrodon 1957; Rouchy et al. 1982; Rouchy et al. 2007: Abbouda et al. 2019), which evolve vertically to sands and sandstones (example: Djebel Meni).
The Pliocene cycle is divided into two phases (PI and PII: Thomas 1985; Neurdin-Trescartes 1992): the first marine one is marked by a transgression (detrital at the base evolving towards deep facies) then regressive at the end (detritic). The second phase represents a generally continental sedimentation preparing the Pleistocene cycle in the region (Perrodon 1957; Thomas 1985; Neurdin-Trescartes 1992). The Quaternary continental red formations clearly underline the southern slope of the Dahra Massif (Brive 1897; Anderson 1936; Perrodon 1957). The Pliocene period is affected by a compressive tectonics, some faults being still active nowadays (Perrodon 1957; Thomas 1985; Meghraoui 1982; Meghraoui et al. 1988; Derder et al. 2011, 2013; Arab et al. 2015; Maouche et al. 2019; Abbouda et al. 2018, 2019).
In this context, the study carried out here relates to the geological series of Ouled Maallah whose deposits cape the Messinian selenite gypsum or post-gypsum, followed by those of the Lower and Middle Pliocene. This region is located at 38 km, east of Mostaganem, near the Ouled Mallah agglomeration (Fig. 1C), on road CW8A and west of CW n° 52 that connects the axis Tazgait-Beni Zenthis road (RN 90).
Material and methods
The Dahra Massif sector has excellent Miocene to Pliocene outcrops whose lithology and stratigraphy are well-known (Brive 1897; Anderson 1936; Perrodon 1957). In fact, in-depth micropaleontological studies have been carried out on this Massif (Belkebir 1986; Belkebir and Anglada 1985) and the northern edge of the Lower Chelif plain (Mazzola 1971; Belhadji 2005; Belhadji et al. 2008; Mansouri 2001; Mansouri et al. 2008).
This study is to particularly establish the chronology of post-gypsum deposits located in the Ouled Maallah sector, an agglomeration located on the meridional edge of the Dahra Massif. The outcrops allowed us to describe in detail their lithology and paleontological contents. In order to do so, several samples were taken there (75) and processed in the laboratory.
For the dating of these deposits, this investigation has necessitated the use of foraminifera and calcareous nannofossils, which permitted the establishment of a biostratigraphic calibration or correlation using biostratigraphic scales. Comparable methods were also been carried out in marine deposits (Berggren et al. 1995; Lourens et al. 2005), transitional marine and continental ones (Bessedik 1984; Bessedik et al. 1997).
Six geological sections aging between the Uppermost Miocene and the Lower-Middle Pliocene, and located at Ouled Maallah, were under study: Moul Settara, Ain Yacoub, Azaizia (a), Azaizia (b), Oued Tarhia, and Garbous Ben Saada (Fig. 1C). The lithological description, sampling, and treatment of the sediments revealed the presence of gastropods, bivalves, scaphopods, solitary polyps, charophytes, ostracods, foraminifera and calcareous nannofossils, and some unidentified bones of proboscidians (S. Sen in litteris).
A systematic sampling was carried out there, at 7- or even 10-m intervals; their microfossil content (foraminifera and calcareous nannofossils) was analyzed, where only the qualitative presence of stratigraphic taxa or biomarkers was taken into account, as bioevents.
This paleontological study involved more than 70 samples processed and analyzed, and in effect, has demonstrated the presence of microfossils; whereas additional (ad hoc) sampling was dispatched when necessary (stratigraphic precision). The extraction of planktonic foraminifera and calcareous nannofossils was performed on the same samples.
A total of 200 to 300 grams of sediment was soaked in lukewarm water for deflocculation and then was washed under a stream of water in a sieve (diameter mesh: 100 μm). Taxonomic analysis and identification of taxa were carried out using a binocular magnifier (magnification × 250 and × 500). These particular samples were subjected to the treatment (smear-slides) of calcareous nannofossils. Afterward, a very small fragment of sediment stored on a slide was diluted in a drop of distilled water, then stirred and well spread. Subsequently, it was put on a hot plate, at moderate temperature for 30 s to 1 min so as to dry it before finally covering it with a bonded coverslip, using 4 to 5 drops of Eukitt resin. Some pressure could be applied on the slide in order to thin down the glue under the coverslip and expel air bubbles. The slide has ultimately dried up for at least 24 h before labeling the sample code and cleaning it.
After observing the slides using a polarizing optical microscope (magnification × 500), analysis was carried out by a systematic scanning of each slide (or several slides belonging to the same sample). The determination was carried out with a magnification of × 500 to × 1000 μm.
This study reports on two stratigraphic situations revealed, following the demonstration of the Messinian-Zanclean boundary. The first concerns the chronology of the Pliocene marine deposits located above this boundary, which are dated and calibrated using foraminifera and calcareous nannofossils. The second situation concerns the deposits located below the Messinian-Zanclean boundary and post-gypsum, and which have provided microfossils indicating a lacustrine environment to a lagoon one (charophytes, ostracods, benthic, and some planktonic foraminifera). Their stratigraphic position and chronological relationship with the gypsum are discussed and correlated on a local and regional scale.
Biostratigraphy: planktonic foraminifera and calcareous nannofossils
Planktonic microorganisms with a wide geographical distribution and, for some of them, with a short chronological extension are generally used in biozonation. With this in mind, we began to study the Mio-Pliocene lands of the Dahra Massif. We compared the associations of planktonic foraminifera and calcareous nannofossils recorded in the Ouled Maallah sections with the different biostratigraphic subdivisions applied in the Mediterranean Basin and calibrated with the standard biozonation of Blow (l969). The adopted biozones are generally based on the criterion of appearance, disappearance, or association of the different taxa used.
Although they efficiently contributed to the knowledge of Neogene strata, the first studies of planktonic foraminifera carried out in the northern Algerian Neogene basins are fragmentary (Magné in: Perrodon 1957; Bizon in: Thomas 1985; Bizon in: Neurdin-Trescartes 1992). For the marine Pliocene, Mazzola (1971) was the first to recognize several successive biozones in the Chelif Basin (from the oldest to the most recent): Sphaeroidinellopsis subdehiscens paenedehiscens, Globorotalia hirsuta praehirsuta-G. margaritae, G. puncticulata and G. crotonensis. Yassini (1975) followed for the Pliocene of the Sahel of Algiers with in particular Sphaeroidinellopsis subdehiscens subdehiscens, Globorotalia margaritae, G. puncticulata and G. crassaformis. Subsequently, the G. inflata biozone was recognized there (Saoudi 1982). The study of the Hachacha Plateau revealed a comparable biozonation (Belkebir and Anglada 1985; Belkebir 1986): G. margaritae, G. puncticulata and G. bononiensis, G. crassaformis / G. crassula viola and G. inflata, completed by that of Belhadji (2005) and Belhadji et al. (2008) with Sphaeroidinellopsis seminulina, G. margaritae, G. puncticulata-G. margaritae, G. puncticulata and G. aemiliana.
For the calcareous nannofossils, the work of Mansouri (2001) and Mansouri et al. (2008) mainly concerned the Upper Miocene. Pliocene in age, the Telegraph of Sidi Brahim section revealed the occurrence of calcareous nannofossils: Discoaster quinqueramus, Amaurolithus delicatus, A. primus, Discoaster surculus, D. brouweri, Reticulofenestra pseudoumbilicus, D. asymmetricus, D. tamalis (Mansouri Med El Habib in: Tchouar 2013).
The Miocene to Pliocene associations of planktonic foraminifera and calcareous nannofossils recorded in the Ouled Maallah sector confirm previous biostratigraphic successions whose marker taxa were collected in the same samples, which gives the latter a clear advantage in terms of calibrating foraminifera with calcareous nannofossils.
In this sector, the Azaizia (a) and Azaizia (b) sections appear stratigraphically more complete, revealing the presence of about ten microfossil taxa (planktonic foraminifera and calcareous nannofossils).
The taxa recorded in the Ouled Maallah sector and those recorded in the Lower Chelif Basin (Mazzola 1971; Belkebir and Anglada 1985), mentioned above, are taken into consideration (Fig. 4). The resulting subdivision is correlated with the subdivisions established in other parts of the Mediterranean Basin (Bizon and Bizon 1972; Bizon and Müller 1976; Zachariasse 1975; Cita 1975; Thunell 1979; Langereis and Hilgen 1991). The results are referred to the Blow standard scale (Blow 1969). For the calcareous nannofossils, the subdivision is based on Martini (1971) and Backman et al. (2012)

Chronologically calibrated bioevents highlighted in the Ouled Maallah succession (Dahra Massif, Lower Chelif Basin)
The biostratigraphic datums of the taxa and their ages are calibrated on the basis of the work of Martini (1971), Bukry (1973, 1975), Okada and Bukry (1980), Berggren et al. (1995), Lourens et al. (2005), Raffi et al. (2006), Iaccarino et al. (2007), Hilgen et al. (2012), Backman et al. (2012), Gradstein et al. (2012), and Lirer et al. (2019).
Note that these authors agree with the Zanclean base (GSSP) defined at 5.332 Ma (Van Couvering et al. 2000). In relation to this boundary, the age of the datums is organized. Indeed, among the planktonic foraminifera (see Fig. 4), the first common occurrence of the Globorotalia margaritae species is considered at 5.08 Ma (Lourens et al. 2005; Lirer et al. 2019), its last common occurrence (LCO) at 3.98 Ma (Lourens et al. 2005; Lirer et al. 2019). We must recall that the top acme for Sphaerodinellopsis subdehiscens is reported at around 5.21 Ma and 5.30 for its base (Lourens et al. 2005; Lirer et al. 2019). In addition, G. margaritae and Ceratolithus acutus may together also date latest Messinian layers; these data have enabled some authors (Bache et al. 2012, 2015; Do Couto et al. 2014; Clauzon et al. 2015a; Suc et al. 2015, among others) to place the marine reflooding of the Mediterranean Basin in the latest Messinian and not at the Messinian-Zanclean boundary (Fig. 4).
The first appearance (FAD) of Globorotalia puncticulata is reported around 4.52 Ma (Lourens et al. 2005; Lirer et al. 2019). The last occurrence of this species (LOD) is dated at around 3.56 Ma (Lourens et al. 2004; Sprovieri et al. 2006), more precisely at 3.57 Ma (Lirer et al. 2019).
The first appearance (FAD) of Globorotalia crassaformis is indicated around 3.6 Ma (Lourens et al. 2004; Sprovieri 1993; Lirer et al. 2019). Note that the first occurrence (FOD) of G. bononiensis is indicated at around 3.59 Ma (Channell et al. 1988, 1990, 1992; Sprovieri et al. 2006; Lirer et al. 2019) that of G. aemiliana is recorded around 3.36 Ma (Lourens et al. 2005; Lirer et al. 2019).
Concerning calcareous nannofossils, the datum of the first occurrence of Ceratolithus acutus is sometimes dated at 5.345 Ma (Raffi et al. 2006) and sometimes at 5.36 (Backman et al. 2012); the age considered by Gradstein et al. (2012) for this bioevent is 5.35 Ma (Fig. 4). So, the species datum’s extinction is indicated at 5.04 Ma (Backman et al. 2012).
The first appearance datums (Fig. 4) of Reticulofenestra cisnerosii (Footnote 1) (5.332 Ma; Di Stefano and Sturiale 2010), Helicosphaera sellii (4.58 Ma; Backman et al. 2012), and Discoaster asymmetricus (4.04 Ma; Backman et al. 2012) are also specified, respectively. Finally, the considered last appearance datum of the species Reticulofenestra pseudoumbilicus (3.79 Ma) and Discoaster tamalis (2.76 Ma) are those proposed by Backman et al. (2012).
These foraminifera and calcareous nannofossil biozones are calibrated with the biozonations of Blow (1969) and Martini (1971) (Fig. 4).
Results: lithology of the studied sections and their bioevents
Six sections from the sector of Ouled Maallah, located on the southern margin of the Dahra Massif and near the beginning of the syneclise of the Lower Chelif are described in this work, dealing with their facies, their paleontological content, and the arrangement of their sedimentary bodies.
Azaizia (a) section
Geological description
This section is located at Oued Sikh, a tributary of Oued Er Razzaz. Five geological formations are listed there. The series begin with Gray Marls, about 13 m in thickness (Fig. 5: D). Rich in gypsum debris, the first level of this formation rests, unconformably, on the massive gypsum. These deposits, with abundant marine macrofauna (gastropods, bivalves, scaphopods, solitary polyps, etc.), have produced calcareous nannofossils and planktonic foraminifera (Figs. 5 and 6; sample A2). Note that the sample A1 mainly provided gypsum debris and did not contain any microorganism. The second alternation of fossiliferous Gray Marls and Calcareous Marls (11 m in thickness) overcomes the previous formation. Like the previous deposits, this formation is rich in malacofauna (Natica sp., Neopycnodonte cochlear, ostreidae, vermets, etc.); malacofauna generally abounds in calcareous marl banks, which gives them a certain resistance to erosion. Sample No. A3 was found to be rich in planktonic foraminifera and calcareous nannofossils (Fig. 6). This geological formation evolves towards Blue Marls (20 m in thickness), which contain bivalves, oyster (Neopycnodonte cochlear), and isolated polyps.

Geological series of Azaizia sections with focus on its part (a): A succession (from the base): Gypsum selenite, Gray Marls, Alternation of fossiliferous gray marls and Calcareous Marls, Blue Marls, Marls, and Calcareous Marls Alternation, Biodetritic calcareous sandstone. A: Azaizia geological section (a); B: Panoramic view and location of the Lower to Upper Pliocene of Azaizia [(a), (b)]; this photography shows the “filling” of the Pliocene marly series within the gypsum; C: Upper part of the “Alternation of marls and calcareous marls”; C1: internal cast of vermets; C2: internal cast of Veneridae; C3: Internal cast of Gastropoda. D: Lower part of “Alternation of marls and calcareous marls”; D1: detail of marls and calcareous marls alternation; D2: Condensation level of bivalves and gullying; D3: Ferruginous coquina sandy marl level. E: The gypsum basement and the lowermost Pliocene Azaizia (a) series. Legend: 1 = Gypsum; 2 = Gray marls with calcareous marl intercalations; 3 = Marly limestone; 4 = Blue marls with marly limestone intercalations; 5 = Biodetritic calcareous sandstone. The hiatus is linked to a lack of visibility

The Lower Pliocene planktonic foraminifera from of the Azaizia (a) section (Dahra Massif). A, B, C: Sphaeroidinellopsis subdehiscens Blow (sample A2); D, E, F: Globorotalia margaritae Bolli & Bermudez (sample A3); G, H, I: G. puncticulata (Deshayes) (sample A6); J, K, L: G. bononiensis Dondi (sample A9); M, N, O: G. aff. crassaformis Galloway & Wissler (sample A13)
This series continues with an Alternation of marls and calcareous marls (more than 145 m in thickness), overlying the previous formation. This alternation is characterized by prevailing marls. Towards the half of this formation, a level of 60 cm appears between samples A9 and A10, rich in bivalve debris. This level is marked at its base by ferruginous deposits, probable witness of emersion (?). The last 5 to 6 m, located at the top of the section, are beige or even yellowish marls to slightly sands, and by cracks filled with whitish material. This formation is characterized by an abundant macrofauna (Neopycnodonte cochlear, vermets, Dentales, Turritella sp., Natica sp., Pectenidae, Cardiidae …). Microfauna (planktonic foraminifera and calcareous nannofossils) is also present there in samples A6 to A17. The section ends with a Biodetritic calcareous sandstone, which outcrops locally with a minimum of thickness (50 cm) in the additional Azaizia (a) section. This deposit goes through a significant spatial and vertical extension near Sidi Abdelkader, which overlooks north of the Ain Yacoub village. These deposits overlay (unconformably) the previous formation, marked by conglomerates at the base, containing Venerid mollusks, evolving to slightly sandy yellow marls with burrows, and becoming limestone at the top.
Bioevents
The washing residues and the treatment of the 12 samples (A2 to A13 are proved to be rich in planktonic foraminifera and calcareous nannofossils; accordingly, this allows us to identify taxa with stratigraphic interest (or biomarkers).
-
Planktonic foraminifera: several markers are highlighted in this section (Fig. 5), in particular the chronological succession of bioevents (from the base to the top: see Figs. 6 and 9): Sphaerodinellopsis/Globorotalia margaritae (sample A2), G. puncticulata (sample A4), G. bononiensis (sample A7) - G. aff. crassaformis (sample A9). This succession is described by Mazzola (1971) in the Lower Chelif Basin, except for the species Globorotalia crassaformis whose position is occupied by that of Globorotalia crotonensis. On the other hand, the succession of biomarkers described in the Azaizia (a) section (Ouled Maallah) is recognized as complete in the Pliocene series of the Hachacha Plateau (northern Dahra Massif) by Belkebir and Anglada (1985) and Belkebir (1986). The same events are recognized in the Miocene-Pliocene series of the Oued Chelif valley mouth where the Globorotalia aemiliana species succeeds to G. bononiensis, G. crassaformis species being absent there (Belhadji et al. 2008). In the Algiers Sahel, Yassini (1973, 1975) describes the same biochronological succession, except the Globorotalia bononiensis biozone.
-
Calcareous nannofossils: The section reveals, from bottom to top, the succession of the following bio-events: Ceratolithus acutus (sample A2)–C. rugosus (sample A2)–Reticulofenestra cf. cisnerosii (sample A3)–Helicosphaera sellii (sample A4)–Discoaster asymmetricus (sample A10). This succession is pointed out for the first time in the Lower Chelif Basin and the Dahra Massif (Figs. 7 and 9).
Fig. 7 
The Lower Pliocene calcareous nannofossils from of the Azaizia (a, b) sections (Dahra Massif). A, B, C, D: Ceratolithus acutus (Gartner and Bukry 1974) (section a, sample A2); E, F, G, H: C. acutus (Gartner and Bukry 1974) (section a, sample A3); I, J: C. armatus/acutus (Müller 1974/Gartner & Bukry 1974) (section a, sample A3); K, L: Reticulofenestra cisnerosii/zancleana (Lancis and Flores 2006/2007; Di Stefano and Sturiale 2010; Lancis et al. 2015) (section a, sample A3); M, N: Sphenolithus neoabies (Bukry & Bramlette 1968) (section a, sample A3); O, P: Helicosphaera sellii (Bukry & Bramlette 1969) (section a, sample A5); Q: Discoaster asymmetricus (Gartner 1969) (section a, sample A11); R: Reticulofenestra pseudoumbilicus (Gartner 1967) (section a, sample A14); S: Sphenolithus abies (Deflandre and Fert 1954) (section b, sample A11); T: Discoaster tamalis (Kamptner 1967) (section b, sample A12). Scale bar = 5 μm

Furthermore, comparable biostratigraphic successions are described by Martini (1971), Raffi and Flores (1995), Berggren et al. (1995), Okada and Bukry (1980), and Backman et al. (2012). This succession can be summarized as follows: Ceratolithus acutus, C. armatus (Footnote 2), Reticulofenestra cf. zancleana, Helicosphaera sellii, Discoaster asymmetricus, Reticulofenestra pseudoumbilicus. Backman et al. (2012) relate the same succession that we describe in the Azaizia (a) section. It is also described elsewhere (Martini 1971; Okada and Bukry 1980). It should be noted that Reticulofenestra cisnerosii is considered to be a bioevent marker for the beginning of the Pliocene (Di Stefano and Sturiale 2010; Lancis et al. 2015).
Azaizia (b) section
A geological description
Unconformable onto selenite gypsum, this section is located on the side of the municipal road connecting Ouled Mallah to the RN 90 (Sidi M’hamed Benali -Tazgait), crossing the Ain Yacoub agglomeration. Overlooking Oued Sikh, this section is topographically higher in altitude than that of Azaizia (a). It begins with white marls and calcareous marls (Fig. 8) generally made up alternating marly limestones with calcareous marls; called white limestones, these deposits are characterized by Ostrea (Neopycnodonte) cochlear (Brive 1897; Perrodon 1957). Macrofaunas are abundant there (Ostreidae including Neopycnodonte cochlear, Natica sp., isolated polyps, scaphopods, Veneridae, etc.). Six samples were taken from these deposits. Above, the Gray Marls Formation is rich in macrofauna, in particular scaphopods (Dentalium), Gasteropods (Natica sp.) and isolated polyps. The samples taken from these Gray Marls are rich in planktonic foraminifera and calcareous nannofossils (Figs. 6, 7, and 9). Indeed, the following succession is reported below.

Azaizia (b) lithologic section. A: Lithologic succession, sampling, and fossil emplacement; B: Post Pliocene faulted (F) Marly Limestone bars and Marls; C: Panoramic view of the White Marls and Marly Limestone alternation, Gypsum; C1: Details in Marly and Calcareous Marls

Stratigraphic distribution of planktonic foraminifera and calcareous nannofossils from the six sections close to Ouled Maallah
Bioevents
This section is located upstream of the first one, near the CW 52 road. It is about 50 m thick and leans against gypsum. By its geometry, it seems to complete, upwards, the Azaizia (a) section. Sixteen samples were taken there, delivering planktonic foraminifera and calcareous nannofossils.
-
Planktonic foraminifera (Fig. 6): three species are identified in this section corresponding to three “bioevents.” From the base, the first sample revealed the presence of the Globorotalia puncticulata and throughout the section. Globorotalia bononiensis appears in sample A11 followed by Globorotalia aff. crassaformis (sample A12).
This succession corroborates the upper position of this section confirmed by the foraminifera content of the Azaizia (a) section, corresponding to its upper part. It is comparable with the succession on the Hachacha Plateau with Globorotalia puncticulata, G. bononiensis and G. crassaformis (Belkebir 1986).
-
Calcareous nannofossils (Fig. 7): two species are identified in these calcareous marls followed by blue to gray marls. This is Discoaster asymmetricus (sample A5) followed by D. tamalis (from sample A12). This succession is described elsewhere (Martini 1971; Okada and Bukry 1980; Backman et al. 2012).
For all the sections of Azaizia (a) and Azaizia (b), it is necessary to note the complementarity of the Pliocene marly series. This constitutes an important sedimentary body, ranging in age from the Mio-Pliocene to the Middle Pliocene, resting in unconformity on previously folded and eroded gypsum; the latter reappears on the margins of the marly series to the north and south, which describes a series interlocking. The impact of erosion appears to be significant at the center, but significantly less at the edges of the structure; the example of the Ain Yacoub section, 800 m from Azaizia (a) and located on its edges at an altitude of 315 m is demonstrative in the fact that the white micritic limestones and white marl with charophytes (lagoon facies) succeeding the gypsum would represent the witness of the material remaining from erosion.
Moul Settara section
Geological description
Consisting of marly, limestone, and fossiliferous sandstone deposits (Fig. 10), this series is faulted; it is evident in particular in the mass of selenite gypsum that constitutes the basement of the succession. There are five distinct geological formations. The first formation begins with Beige marls with sandy intercalations (A), the contact of which with the underlying gypsum is not reached here. The beige color, sometimes only relating to their surface, can change to gray. They contain macrofauna like Ostreids including Neopycnodonte cochlear, Pectenidae, and Gastropods. Three samples (Ms1 to Ms3) were taken there, delivering foraminifera and calcareous nannofossils. This formation is followed by Gray marls (B & D) with an intercalation of Fossiliferous calcareous sandstone (C) whose thickness can reach 4 to 5 m. The latter is very rich in malacofauna (Pectenidae, Ostreidae, Gastropods, etc.). Three samples (Ms4 to Ms6) from the gray marls yielded foraminifera and calcareous nannofossils. This section ends with White calcareous marls, which can reach 10 m in thickness in some places. They are very rich in broken and whole oyster shells (including Neopycnodonte cochlear), Gastropods, dentals, solitary polyps. By their facies, their color, and their paleontological content, they become identical to the alternation of Marls and white Calcareous Marls, located at the base of the Azaizia (b) section, which also yields foraminifera and calcareous nannofossils.

Moul Settara section. A: Lithologic succession, sampling, and fossil emplacements; B. Panoramic views of the formations with their location; C: B, C, and D formations; D: A, B and C formations; E: Gypsum selenite Formation. Legend: 1: Gypsum; 2: Beige sandy marls; 3: Gray marls; 4: Sandy limestones; 5: Calcareous marls; 6 F: Fault
Bioevents
Opposite to Azaizia (a), this section is located on the left bank of Oued Sikh. Six (6) samples are taken in the gray to beige marls, delivering three biomarkers.
-
Planktonic foraminifera (Figs. 6 and 9): only the species Globorotalia puncticulata is notified there (samples 1–6), already reported in the Dahra Massif (Belkebir 1986) and in the Lower Chelif Basin (Mazzola 1971; Belhadji et al. 2008).
-
Calcareous nannofossils (Figs. 7 and 9): Two events are revealed by the presence of Helicosphaera sellii (samples 1–6) and Discoaster asymmetricus (samples 1, 2, 6).
Ain Yacoub section
Geological description
Five geological formations are identified in this section (Fig. 11):

Ain Yacoub geological section. A: Lithologic succession, sampling, and fossil emplacements; B: Panoramic views of the five geological formations. Note the geometry of the gypsum (tilted) and the (subhorizontal) overlying Pliocene deposits. C: Limestone biodetritic sandstone. D: The white Messinian micritic limestone formations, laminated, bechified, and fractured micritic white limestone formations (B) and beige Pliocene marls (D); Ay1, Ay2,…: samples; E: The formation of white micritic limestones (B) followed by gypsum lenses in spearhead and the formation of white marls with charophytes and ostracods (C). F: details in the formation of the micritic white limestones (B) showing contoured and slumped limestone banks (Ga); the laminite arrangement covered (Gb) by gypsum in spearhead (Gc) in vertical position. The whole shows a dip conforming to that of massive gypsum. Legend: 1: Gypsum selenite; 2: White waved calcareous marls; 3: Laminated calcareous marls with gypsum; 4: White calcareous marls; 5: White marls; 6: Beige gray marls; 7: Calcareous biodetritic sandstone
Resting (unconformably) on the massive selenite gypsum (A), the laminated micritic white limestones (B) are 6 m thick. They are disturbed, fractured, and brecciated with sometimes gypsum balls and slumps of micritic limestones. A sample taken from these levels showed, in a thin section, filaments, fairly comparable with stromatolites (?). These limestones contain gypsum lenses in spearhead whose crystals, in the shape of sheaves, are planted vertically on limestone platelets. Evoking a gypsum substratum instability, this sedimentation testifies to an already existing slope or in the course of construction. Above are White Marls with Characeae and Ostracods (C), around 5 to 6 m thick. They follow with the same dip of the previous formation. Washing of the marls revealed the presence of characeous oogonias and agglutinated or separated shells of generally smooth ostracods. These organisms indicate a lake environment.
These deposits are followed by Beige marls (D), with a thickness exceeding 12 m. They are almost horizontal and unconformably capping the previous deposits (Micritic limestones and White marls with charophytes). They are rich in Ostreidae including the species Neopycnodonte cochlear. Two samples were taken there, revealing the presence of foraminifera and calcareous nannofossils (Figs. 6, 7, and 9). These beige marls are capped by Biodetritic calcareous sandstones (E). With yellow color, these sandstones have massive bars, containing fragments of bivalve shells (2 m in thickness locally) they can reach more than 50 m at Sidi Abdelkader locality (Fig. 9: photos 1 and 2).
Bioevents
The micritic white limestones in platelets or finely lined and bypassed rest unconformably on the Messinian selenite gypsum by means of pockets of brecciated white limestones or gypsum blocks (Fig. 11). These limestones are covered by white marls that provided Ostracods and Characeae, indicating a lake or even lagoonal environment. These deposits did not provide a stratigraphic bio-marker, on contrary, the unconformably overlying beige marls yielded the foraminifera Globorotalia puncticulata and the calcareous nannofossils Discoaster asymmetricus and Helicosphaera sellii (Fig. 9).
Oued Tarhia section
Geological description
This section is carried out on a north-south transect, along the Oued Tarhia, with a thickness exceeding 350 m.
This series overlies the massive selenite gypsum (Fig. 12); the latter is stretched and fractured in particular in the center of the structure where there are forms of extrusion and tightening in the normal fault direction (Fig. 12, J, K1, and K2). In addition, the transect of the Oued Tarhia Valley is a major NNE/SSW fault axis, causing the Hamri sector to move north (translation), while giving it a slight eastward rotation.

Oued Tarhia section. A: Lithological succession, sampling, and fossil presence; B: Beige marls and yellow biodetritic sandy Limestone Formation; B1: Topographic slope revealed by onlap system; B2: Tectonic and subsidence movements; C: Onlap of sedimentary bodies’ image (by Google earth) showing north vergence; D, D1, D2: Faunal content on lower bench surface; E: position of the samples in the gray marls, along the Tarhia Valley; E1: Position of the samples in the Blue marls; F: Laminated sandstone benches; G: Conglomerate and crossbedded sandstones; H: Lacustrine gray marls, unconformity, Messinian Gypsum; I: To the left: Pliocene blue marls, to the right: Gypsum, Lacustrine gray marls, and sandstone alternation; J: in the center: the massive faulted gypsum; K1, K2: details in the gypsum and anhydrite fault movements. Legend: 1: Gypsum; 2: gray and blue marls; 3: conglomerates; 4: sandstones and sandy Marls; 5: Limestone; 6: sandstones; 7: Marls and sandstones; 8: cross-bedded sandstones; 9: Calcareous marls; 10: sandy limestones.
This series consists of five geological formations:
Unconformably resting on selenite gypsum, the Gray to variegated marls (B) begin this section with a thickness of around 6 m. Four samples were taken there which are poor in planktonic microfauna, except for the presence of some smooth forms of internal mold of ostracods, attesting to a palustrine or even lacustrine environment. About 10 m away, an Alternation of Conglomerate, marls, and sandstones (C) starts at the base with an unconformity observed on the Gray to variegated marls, represented by conglomerates and sandstones. The latter evolves towards marls and calcareous sandstones, sandy marls, sandstones that ends with crossbedded sandstones. Above, succeed Gray Marls (C), 70 m thick, having delivered foraminifera and calcareous nannofossils (Ot7 to Ot9). They are followed by Blue marls (D), comparable with the previous gray marls but they differ in color. Their thickness exceeds 200 m, much of which disappears under crops (hiatus). Their upper part is enriched with calcareous marl benches with a thickness of 3 to 4 m each time. Two samples were taken from the lower part of the Blue Marls which delivered foraminifera and calcareous nannofossils (samples Ot10, Ot11, Ot12, Ot13, and Ot14).
On the previous deposits, rest (because of erosion) Beige marls and yellow biodetritic calcareous sandstones (F). This alternation exceeds 150 m of thickness. The biodetritic calcareous sandstones form bundles of benches, each with a thickness of 3 to 4 m. They are rich in bivalves such as Ostreidae (Neopycnodonte cochlear, among others) and Pectenidae observable on the lower base of the beds (Fig. 12, A, D1 to D3); the benches are arranged in “onlapping” sedimentary bodies, forming structures indicating the presence of subsidence phenomena associated with a syn-sedimentary tectonics (Fig. 12: E1 and E2). The samples taken in this formation have delivered foraminifera (samples Ot15, Ot17) and calcareous nannofossils (Ot16).
Bioevents
Eleven samples were taken in this section: eight in gray marls and alternating marl with marly limestone and three in alternating yellow marls and marly limestones.
-
Planktonic foraminifera (Figs. 6 and 9): four biomarkers are evidenced in the gray marl formation: Globorotalia margaritae (samples Ot1-Ot3), G. puncticulata (samples Ot1-Ot11), G. bononiensis (samples Ot9-Ot11 in yellow marls) and G. aff. crassaformis (Ot9-Ot11 samples). This succession is known in the northern part of the Dahra Massif (Belkebir 1986; Belkebir and Anglada 1985), in the Lower Chelif Basin (Mazzola 1971; Belhadji et al. 2008) and in other sectors located eastward of this basin (Yassini 1973, 1975).
-
Calcareous nannofossils (Figs. 7 and 9): two taxa are recorded in the analysis of this geological series: Discoaster asymmetricus (samples Ot1–Ot4 and 8) and D. tamalis (samples Ot6–Ot8 and Ot10).
Garbous Ben Saada section
Geological description
The geological surveys of this section (Fig. 13) were carried out on the southern slope of the Dahra Massif, along a tributary of the Oued Chelif that runs along the municipal road located at the eastern exit of Ouled Maallah at Douar Mellalha, connecting it to the agglomeration of Tazgaït, via the RN 90.

Geological series of Garbous Ben Saada section. A: Lithologic, sampling and fossil emplacements; B: Quaternary red continental formation; C: Formation of biodetritic and sandy marls; C1: The location of the Proboscidian fossil remains; C2: Fossil bones of indeterminate Proboscidians and details; D: Turitella sp.; D1: Natica sp.; D1: Conidae; E: Fossiliferous marly limestone: Cardiidae, Dentalia sp.; F: Panoramic view of Garbous Ben Saada geological series (The southern slope of the Dahra Massif) with the principal formations (A, B, C, D, E and F: see description) and sampled levels (G1, G2, ..., G9). Legend: 1. Blue marls; 2. Fossiliferous marly limestones; 3. Biodetritic sandy marls; 4. Sanstones; 5. Sandstones and red silt alternation; 6. Red conglomerates and microconglomerates
The section was sampled in the last 12 m, completing, in the sector of Ouled Maallah, the large areas of blue marls which initiate the of Lower Chelif Plain. It begins with Marine blue marls (A and C) from which four samples have been taken; they delivered planktonic foraminifera and calcareous nannofossils (Fig. 13). An intercalation of fossiliferous calcareous Marls (B) is observed within the preceding Marine blue marls. It is a bar of 2-3 m of thickness; it is rich in macrofauna with generally whitish tests, notably of Bivalves, Gastropods, Scaphopoda, among others. The microfauna is dominated by benthic foraminifera.
The blue marls evolve progressively to yellow biodetritic sandy marls (D) with a thickness exceeding 140 m. Marked by sandstone intercalations, they are rich in Ostreidae and Gastropods; they also delivered fossil remains of indeterminable Proboscidians (Sevket Sen, in litteris). Above, comes an Alternation of Sandstone and red clays (E), clearly unconformable on the previous formation, containing levels with root remains; emphasize the margin of the southern slope of the Dahra Massif, this formation is attributed to Quaternary (Brive 1897; Perrodon 1957). The section ends with large spreading of Conglomerates (F) and microconglomerates
Bioevents
This section, located on the southern margin of the Dahra Massif, was the subject of four samples from the gray marls, which revealed the presence of planktonic foraminifera and calcareous nannofossils. The yellow sandy marls totaling a hundred meters thick did not deliver microfossil. Four biomarkers were recorded in this part of the section (Fig. 6):
-
Planktonic foraminifera (Figs. 6 and 9): two species are identified from the base of this geological series. These are Globorotalia margaritae (samples G1-G2) and G. punticulata (samples G1–G4).
-
Calcareous nannofossils (Figs. 7 and 9): two biomarkers are recorded in these gray marls (samples G1–G4). These are Discoaster asymmetricus and D. tamalis.
Discussion
The age of the selenite gypsum is here discussed based on its local and regional correlations, environment, and stratigraphic position in the context of the Messinian Salinity Crisis in the Mediterranean region. The lacustrine deposits overlying it are correlated, dated, and integrated into this chronostratigraphic context. Moreover, the identified bioevents in the Pliocene deposits are discussed and correlated with the planktonic foraminifer and calcareous nannofossil datums. Finally, local and regional correlations are proposed, including some tectonic deformations and gravity movements highlighted in the Lower Chelif Basin and the Dahra Massif.
The age of the selenite gypsum
Evaporite deposits are well-known in the Chelif Basin and the Dahra Massif (Brive 1897, 1920; Anderson 1936; Busson and Magné 1955; Perrodon 1957; Gourinard 1952, 1958; Delfaud et al. 1973; Rouchy 1982; Thomas 1985; Saint Martin 1990; Neurdin-Trescartes 1992). These deposits are generally attributed to the Sahelian (Pomel 1889) or to the latest Miocene (Anderson 1936; Perrodon 1957). The gypsum deposit or "Mellah" of Anderson (1936) is linked, in Oran region to a trilogy of facies whose vertical evolution (diatomites or tripolis - algal limestone / coral-gypsum) even lateral passage (p.p.), seems to be increasingly well-established (Rouchy 1980, Rouchy 1982a, b; Saint Martin 1990; Saint Martin and Rouchy 1990; Rouchy and Saint Martin 1992; Saint Martin et al. 1992; Rouchy and Caruso 2006, among others).
Basically, the dating of the gypsum is often provided by planktonic foraminifera, extracted from the underlying deposits (marls, diatomites, and diatomitic marls). In fact, the picked samples, at the level of Djebel Sikh (Ouled Maallah), in an alternation of marls and diatomitic marls, capped by gypsum, reveal the presence of Globorotalia mediterranea and Amaurolithus primus associated with A. delicatus. The first occurrence of this species marks the beginning of the NN11b Subzone, the base of which is still located in the Tortonian; the first occurrence of A. delicatus evidences the Tortonian-Messinian boundary (Colalongo et al. 1979; Theodoridis 1984; Rakic El Bied and Benson 1996; Negri and Villa 2000; Raffi et al. 2003; Morigi et al. 2007; Mansouri et al. 2008). The Subzone NN11c, of Messinian age, is marked by the first occurrence of Nicklithus amplificus.
This age is generally confirmed everywhere on the southern margin of the Dahra Massif, from the Chelif Mouth in the West to Bouzghaia locality in the East, based on the occurrence of Globorotalia mediterranea (Rouchy 1982a, b, Belkebir and Anglada 1985; Belkebir 1986; Belkebir et al. 2002; Aifa et al. 2003; Mansour 2004; Belkebir et al. 1996, 2008; Belhadji et al. 2008; Hebib 2014); The presence of Amaurolithus delicatus and Reticulofenestra rotaria is followed by the species Nicklithus amplificus which marks the NN11c Subzone (Mansouri 2001; Mansouri et al. 2008). This attribution is also confirmed in the sediments of the Beni Chougrane northern slopes (Rouchy 1982a, b; Saint Martin et al. 1983; Poignant and Moissette 1992; Chikhi 1992; Mansour et al. 1995; Mansour 2004; Mansour et al. 2008).
Some dating elements could be sought in the sedimentary bodies constituting the carbonate platform of the Murdjadjo (Arambourg 1927; Delfaud et al. 1973; Rouchy 1982a, b; Saint Martin 1990; Cornée et al. 1994a; Belkebir et al. 1994; Belkebir et al. 1996); this provides a post-reef sedimentation, commonly called “Terminal Carbonated Complex (TCC)” where massive lenticular selenite gypsum is integrated therein (Rouchy 1982a, b; Saint Martin 1990; Cornée et al. 1994b). The coral reef platform of Murdjadjo is correlated, by its anatomy, its stratigraphic position, and biostratigraphy, with those well-known from Melilla in Morocco and Sorbas in Spain (Cunnigham et al. 1997; Cunningham and Collins 2002; Cornée et al. 2004; Clauzon et al. 2015a; Manzi et al. 2013: Roveri et al. 2014b, 2016).
The radiometric dating of the TCC (including gypsum), estimated at Melilla between 5.82 and 5.79 Ma (Cunningham et al. 1997), has been extended stratigraphically, between 5.95 and 5.60 Ma (Cornée et al. 2004). The Sorbas gypsum is dated between 5.94 and 5.96 Ma at the base and around 5.60 - 5.54 Ma at the top (Gautier et al. 1994; Krijgsman et al. 1999). Recently, the section of Sorbas (Martin and Braga 1994; Riding et al. 1998; Fortuin et al. 2000) was reviewed, which allowed the gypsum to be dated at its base of 5.971 Ma (Manzi et al. 2013) and at its top at 5.60 Ma (Krijgsman et al. 2001; Clauzon et al. 2015a). This landmark (5.60 Ma) would correspond to the top of the lagoon deposits of Vera and San Miguel de Salinas (Fortuin et al. 1995; Clauzon et al. 2005; Popescu et al. 2009, 2015; Caruso et al. 2020). At Sorbas, TCC corresponds, for Clauzon et al. (2015a), to the upper (more marine) part of the gypsum while for Roveri et al. (2009, 2020) it corresponds to the whole gypsum.
In the paleogeographic context of the Messinian Salinity Crisis, the evaporites (sulfates) are generally deposited in shallow peripheral basins, compared with those (chlorides: K, Na, Mg) lying in deeper (marginal or central) basins. This relative stratigraphic position is still debated (Clauzon et al. 1996; Riding et al. 1998; Butler et al. 1995, 1999; Krijgsman et al. 1999; Rouchy and Caruso 2006; Manzi et al. 2013; Roveri et al. 2014b; Clauzon et al. 2015a; Roveri et al. 2016), though it currently tends towards consensus (CIESM 2008).
Indeed, the chronological succession of evaporites in the Mediterranean Basin is established as follows: in a first step (5.97–5.60 Ma), evaporites are deposited in peripheral basins followed, in a second step (5.60–5.46 Ma), by evaporites setting in central basins; this last stage corresponds to a huge drop of the Mediterranean Sea level and a strong subaerial erosion of its margins (Clauzon et al. 1996, 2015a; CIESM 2008; Bache et al. 2012, 2015).
The Age of lacustrine deposits (or Lago Mare) and their regional correlation and stratigraphic position
In the Ain Yacoub section, the white micritic limestones in platelets or those finely lined and contoured lay unconformably on the Messinian selenite gypsum by means of pockets of brecciated white limestones or blocks of gypsum (Fig. 11-4). These limestones are capped by white marls providing ostracods (details: see below) and Characeae.
These white marls indicate a lacustrine or even lagoonal environment. The beige marls unconformably overlying them provided the presence of Globorotalia puncticulata, Discoaster asymmetricus, and Helicosphaera sellii, thus allowing us to attribute them to the Lower Pliocene (Zanclean). Consequently, the lacustrine deposits are trapped between the Messinian gypsum and the Lower Pliocene deposits. This stratigraphic position is not different from that of Oued Tarhia (section) gray marl to variegated deposits. Resting on the Messinian gypsum through an irregular surface (Fig. 11-H), these provided ostracods, indicate a lacustrine environment.
All these data demonstrate that the white micritic limestone could have a coastal marine character as it has been considered elsewhere (Belhadji et al. 2008). However, the sedimentation with a lacustrine to lagoonal character, overcoming gypsum in the Aïn Yacoub and Oued Tarhia sectors, would correspond to a Lago Mare episode, compared with that described (Rouchy 1982a, b; Rouchy and Saint Martin 1992; Rouchy et al. 2007) in the Dahra (Oued Aicha, Oued Ben Zar) and Beni Chougrane (Ghomri, Sahaouria, Sig) massifs (Fig. 14-B).

Stratigraphic correlations between the Lago Mare layers: A: Oued Tarhia, Ain Yacoub, Murdjajdo corallian Platform. The ostracods collected in the Aïn Yacoub and Oued Tarhia sections are comparable with those determined at Ouled Slama-Djebel Rokba (Loxochochissa, Cyprideis, Limnocythere, etc.). B: Regional Lago Mare correlations (Dahra, Beni Chougrane). The ostracods collected in the Touaka, El Ghomri, Oued Aicha, and Oued Ben Zar sections are listed in Rouchy et al. (2007): C. agrigentina, C. anlavauxensis associated with Amnicythere accicularia, Amnicythere sp., Candona sp., Camptocypria sp., and Loxoconcha sp.
The Lago Mare episode described in the Lower Chelif Basin shows some peculiarity, notably a runoff of continental freshwaters marked by charophytes. Indeed, the Lago Mare concept is sometimes linked to desalination episodes experienced by the Mediterranean Basin during the Messinian Salinity Crisis when Paratethyan waters entered the Mediterranean Basin under more humid climatic conditions (Orszag-Sperber 2006).
Because of (1) the identity of the Lago Mare fauna (mollusks: dreissenids and lymnocardiids; ostracods: Cyprideis pannonica group) and microflora (dinoflagellate cysts) in the Mediterranean Basin with those endemic of the Eastern Paratethys, and (2) the coeval occurrence of Mediterranean marine species (foraminifera, calcareous nannofossils, and dinoflagellate cysts) in the Eastern Paratethys, crossed exchanges of surface waters have been considered between the two basins during episodes of high sea level (Clauzon et al. 2005; Popescu et al. 2009, 2015).
The presence of lacustrine deposits in the Ouled Maallah sectors and their extension towards the East (Oued Aicha, Oued Ben Zar), on the one hand on the edge of the Dahra Massif and South of the Lower Chelif Basin, on the other hand in the Beni Chougrane sectors (Ghomri, Sahaouria, Sig), shows a vast extension of these lacustrine to lagoonal facies (Fig. 3; Fig. 13, B). The study of other sectors is in progress.
The first studies and conditions of Lago Mare deposits in the Lower Chelif Basin were carried out by Rouchy et al. (1982) and Rouchy et al. (2007) who attributed it to the late Messinian. Their microfossil content is marked, according to Rouchy et al. (2007), by ostracods including the Cyprideis group (C. agrigentina and C. anlavauxensis) associated with Amnicythere accicularia, Amnicythere sp., Candona sp., Camptocypria sp. and Loxoconcha sp., indicating hyposaline water conditions, but also Characeae and benthic foraminifera typical of oligohaline environments.
Near those of Aïn Yacoub and Oued Tarhia, the Ouled Slama-Djebel Rokba section (A. Atik, study in progress), located to the east of Ouled Maallah, reveals the presence in the Lago Mare deposits of foraminifera, charophyta and an ostracod association dominated by Loxochochissa, Cypereides, Limnocythere, of which we can consider the Paratethian origin (Faranda et al. 2007).
Yet, the Ouled Maallah sections (Ain Yacoub, Oued Tarhia) complete this correlation and allow a better understanding of this episode as well as the reflooding modalities in the region.
Moreover, the sedimentary succession of gypsum and white micritic limestones registred at Ouled Maallah may constitute a lateral facies grading to those (p.p.) of the Murdjadjo reef complex (Fig. 14, A). Note that the micritic limestones and the Murdjadjo stromatolites are correlated with the Terminal Carbonated Complex of the Melilla platform and that of Sorbas (Cornée et al. 2004). If the dates of these deposits (TCC) are estimated between 5.8 and 5.6 Ma (Clauzon et al. 2015a) or 5.97-5.6 (Roveri et al. 2020), the radiometric dates obtained on the Melilla platform and correlated to that of the Murdjadjo allow us to locate the white micritic limestones of Murdjadjo between 5.95 and 5.60 Ma (Cornée et al. 2004). Consequently, the white marls (Ain Yacoub section) or gray to variegated marls (Tarhia section) with lacustrine to a lagoonal character could be correlated to a platform marine episode (limestones with stromatoliths and oolites). The post-Lago Mare conglomerates intersecting unconformably the gray marls variegated with Ostracods (Tarhia section: Fig. 12-H) seem to correspond to a tectono-eustatic activity of Messinian age, because of the existence of overlying Pliocene deposits with Globorotalia puncticulata. This sedimentary unconformity can be correlated with that evidenced in the Sahaouria section (Atif 2001; Rouchy et al. 2007), achieved before the olistotith set up.
Furthermore, the Messinian lagoonal deposits are well-known in the Mediterranean regions (Ruggieri and Sprovieri 1974, 1976; Ciaranfi et al. 1978; Colalongo et al. 1978; Cita and Colombo 1979; Rouchy et al. 1980; Geerlings et al. 1980; Cita and Vismara-Schilling 1980; De Deckker et al. 1988; Orszag-Sperber et al. 1989; Fortuin et al. 1995; Orszag-Sperber et al. 2000; Rouchy et al. 2003; Bassetti et al. 2006; Cosentino et al. 2006; Orszag-Sperber 2006; Pierre et al. 2006; Rouchy and Caruso 2006; Rouchy et al. 2007; Popescu et al. 2009, 2015; Do Couto et al. 2014; Popescu et al. 2007), representing, like gypsum, important environmental successions in the Mediterranean (MSC) history. This ended with generalized Lago Mare episodes which were fully marine, marked by Paratethyan influxes and interrupted by the Pliocene marine reflooding (Do Couto et al. 2014; Popescu et al. 2015).
Marked by the presence of brackish or marine organisms (mollusks, ostracods, dinoflagellate cysts, charophytes), this sedimentation is of an earlier (or contemporary) age than the reflooding of the Mediterranean Basin (Gignoux 1936; Ruggieri 1962; Cita and Colombo 1979; Clauzon et al. 2005; Orszag-Sperber 2006; Rouchy et al. 2007; Popescu et al. 2009, 2015), or even slightly later than the Messinian evaporites (Hsü et al. 1973). This reflooding of almost completely dried-out basins (Cita et al. 1978a, b) resulted from a rapid supply of water with low salinity from the Paratethys. Since then, recent studies concerning the history of the Mediterranean MSC have made it possible to recognize several high sea-level exchange “episodes” of water masses between the Mediterranean and the Paratethys (Clauzon et al. 2005; Snel et al. 2006; Popescu et al. 2009, 2015; Suc et al. 2011; Do Couto 2014, and references herein). This led to distinguish three chronologically distinct Lago Mare (LM) events (Clauzon et al. 2005; Popescu et al. 2015). The LM1 is located between 5.64 and 5.60 Ma, coating the peripheral evaporites and affected by the Messinian Erosional Surface (MES) (Clauzon et al. 2005; Popescu et al. 2009; Clauzon et al. 2015a). The LM2 is reported in a deep drilling of the central basins (ca. 5,500 and 5,460 Ma), The LM3, following the reflooding in the Mediterranean, is estimated at 5,460 and 5,332 Ma (Clauzon et al. 2005; Popescu et al. 2007, 2009, 2015; Bache et al. 2012, 2015, Do Couto et al. 2014, and references herein). Manzi et al. (2013) and Roveri et al. (2014a, b, 2016) consider that the Lago Mare phases (LM1 and LM3) represent a single phase, located between 5,42 and 5, 332 Ma in deep basins.
In Spain, the Vera and San Miguel de Salinas sections are considered to represent the first Lago Mare (LM1), estimated slightly older than 5.60 Ma (Bache et al. 2012, 2015; Popescu et al. 2015; Caruso et al. 2020); it is also the age of the youngest selenite gypsum. With an age slightly earlier than 5.46 Ma, the second (LM2) is identified “offshore” in the Alboran and Balearic basins (Bache et al. 2015; Do Couto et al. 2014; Popescu et al. 2015). The last one (LM3) is recognized “offshore and onshore,” in deep holes and in the marginal (Malaga section) and semi-deep basins (Sicily), of an age between 5.46 and 5.3 Ma. While recalling that some other authors only recognize a single Lago Mare event located before the Pliocene transgression (Manzi et al. 2013; Roveri et al. 2014b; Roveri et al. 2016).
Consequently, the Lago Mare highlighted in Ouled Maallah (Fig. 14) and those known in the Lower Chelif Basin, could correspond to LM1 (Fig. 15) with local lacustrine conditions (Charopytes) if we take into account field observations made in the Ouled Mallah (Aïn Yacoub, Oued Tahria) and Beni Chougrane sectors (Sahaouria: Atif 2001; Rouchy et al. 2007).

Stratigraphic position of Messinian evaporites and Lago Mare events in the Mediterranean (modified after Bache et al. 2012, 2015; Popescu et al. 2015). Algeria: Dahra Massif tectonic events, Lago Mare 1; position of the Azaizia (a) section bottom based on the Globorotalia margaritae and Ceratolithus acutus presence
Mio-Pliocene bioevents, ages, and correlations
More than ten bioevents have been recorded (foraminifera and calcareous nanofossils) in the Azaizia (a) section considered the local reference section. The succession of foraminifera species is marked by Sphaeroidinellopsis subdehiscens, Globorotalia margaritae, G. puncticulata, G. bononiensis, and G. aff. crassaformis. It indicates the biozones N18 p.p., N19 and N20 p.p. (Mazzola 1971; Belkebir 1986) corresponding to the Lower and Upper Pliocene (Zanclean and Piacenzian). The Sphaerodinellopsis subdehiscens biozone can be integrated into the G. margaritae biozone (Belkebir, op. cit.) or separated (Mazzola 1971; Yassini 1975; Belhadji et al. 2008). The upper part of the Globorotalia aff. crassaformis would indicate a Piacenzian age. The succession of bioevents of calcareous nannofossil species (Ceratolithus acutus, Reticulofenestra cf. zancleana, Helicosphaera sellii, Discoaster asymmetricus) supports this attribution (Fig. 16).

Chronostratigraphy (foraminifera, calcareous nannofossils) of the Ouled Maallah section (Dahra Massif). Horizontal discontinuous lines: datum correspondence; squares = proposed chronostratigraphic extension
The chronologically calibrated two scales show that Sphaerodinellopsis subdehiscens, Globorotalia margaritae, and Ceratolithus acutus indicate the lowermost Zanclean, but can also be from the latest Messinian (Fig. 4), as demonstrated at Sorbas (Clauzon et al. 2015a) and in many other locations (Popescu et al. 2017, and references herein). These elements have enabled some authors (Bache et al. 2012, 2015; Do Couto et al. 2014; Clauzon et al. 2015a; Suc et al. 2015, among others) to date the reflooding of the Mediterranean basin of the latest Messinian and not at the Messinian-Zanclean boundary (see Figs. 4 and 15).
The occurrence of Ceratolithus rugosus and Reticulofenestra cf. cisnerosii, Helicosphaera selli species confirms the chronostratigraphic continuity of the deposits. The top of the section is attributed to the Zanclean on the basis of the first occurrence of Discoaster asymmetricus, which marks the base of biozone NN14. The beginning of the Piacenzian seems to have not been recorded, probably because of a not enough resolution in sampling. This possibility is, yet, supported (see below) in the Azaizia (b) section, which delivers G. puncticulata and Discoaster tamalis as the tenth bioevent.
The recorded succession of biomarkers in the Azaizia section (b) leads to three remarks:
-
The first concerns the possible stratigraphic discontinuity between the Azaizia (a) and Azaizia (b) sections, some evidence of which is provided by their respective topographic location, the latter being higher in altitude;
-
The second relates to the basal part of the Azaizia (b) section belonging to the G. puncticulata biozone (N19), the topmost part of which corresponds fairly well to the biozone NN13 marked by the first occurrence of C. rugosus; the NN14 biozone is indicated by Discoaster asymmetricus (Martini 1971) / (CN11a of Okada and Bukry 1980);
-
The third concerns the top of the section, which would belong to the biozone with G. aff. crassaformis (N20: Belkebir 1986) which is attributed to the Piacenzian. So, the record of this stage is supported by the Discoaster tamalis bioevent, which occurs in the biozones NN15b / CN11b (Perch-Nielsen 1985); its last occurrence is located at the end of the NN16a / CN12a biozones (Martini 1971; Okada and Bukry 1980).
The Globorotalia puncticulata bioevent recorded in the Aïn Yacoub section corresponds to the biozone N19 of Blow (Belkebir and Anglada 1985), equivalent to the common extension interval of the biozones N19 and NN14 + NN15 (late Zanclean). This stratigraphic extension is supported by the presence of Helicosphaera sellii (NN13), Discoaster asymmetricus (NN14), and D. tamalis (NN15b) according to Martini (1971), evidencing the CN11b biozone of Okada and Bukry (1980). Correlated with the Azaizia (a) section, the Azaizia (b) section seems to be incomplete at its base and truncated at its top (Fig. 16). Consequently, the base and the top of this section could be limited by discontinuities. The first discontinuity (bottom) would be related to the continuing Pliocene sea-level rise after the reflooding, and the second discontinuity to the erosion of the upper part of the Pliocene deposits, resulting in the carbonate sandstone platform (yellow bio-detrital).
The first appearance of Globorotalia puncticulata, Helicosphaera selli, and Discoaster asymmetricus in the Moul Settara section implies its attribution (Fig. 16) to the biozones N19, NN13, and NN14 + NN15 (CN10c, CN11a). The calibration of these events confirms and specifies the age usually considered for this section, i.e., the biozone N19 (upper part excluded) and the upper part of the biozone NN13 (CN11a) and NN14 + NN15 (middle to late Zanclean).
The occurrence of Globorotalia margaritae associated with G. puncticulata, Discoaster asymmetricus and D. tamalis results in proposing a younger age of the gray marls of Garbous Ben Saada and to attribute (Fig. 16) them to the latest Zanclean or even the earliest Piacenzian (N19 equivalent to NN4 + NN15 and NN16). The absence of the Globorotalia aff. crassaformis could exclude the Piacenzian, which could in fact correspond to the yellow sandy marls overlying the gray marls.
The calibration based on planktonic foraminifera and calcareous nannofossils suggests an extension (Fig. 16) of the Oued Tarhia section going from the N19 biozone (upper part) to the N20 biozone, which corresponds to the NN14 + NN15 and NN16 biozones of Martini (1971). This time-interval encompasses the Zanclean-Piacenzian boundary, to be placed within the transition from gray to yellow marls in the sector.
Stratigraphic implications and paleogeography
Sections of the Ouled Maallah sector
The biostratigraphic landmarks (i.e., Globorotalia aff. crassaformis and/or Discoaster tamalis) provided by some sections [Oued Tarhia, Garbous Ben Saada, Azaizia (a), Azaizia (b)] are considered, by calibration, to be contemporary because they are attributed to N20/NN16 biozones. From this biostratigraphic attribution and correlation of the sections in the geological series of Ouled Maallah (Fig. 17), three remarks can be asserted:
-
The presence of an irregular pre-Pliocene topography probably linked to the sea-level drop and tectonic context, generated by the Messinian Salinity Crisis;
-
The presence (for the first time) of the calcareous nannofossil Ceratolithus acutus associated with Sphaerodinellopsis subdehiscens and G. margaritae demonstrates that the base of the marly series of the Azaizia section (a), would belong to the base of Zanclean but it can also be dated as of the end of the Messinian (Figs. 4 and 16). This last hypothesis suggests a latest Messinian reflooding over the gypsum and Lago Mare sediments, the base of which can be assimilated to the Messinian Erosional Surface (Fig. 19). This data is in agreement with the presence of C. acutus, located not far from the base, in the white marls “Trubi facies” of the Telegraph Sidi Brahim section (Mansouri in: Popescu et al. 2017);
-
The Zanclean/Piacenzian boundary is recorded in the Azaizia section (a), above a ferruginous level (identified between samples A9 and A10) followed by a bio-organic concentration (bivalves). Indeed, level A10 reveals the presence of Globorotalia aff. crassaformis;

Correlations of the Ouled Maallah sections
-
A significant subsiding depocentre, expressed by significant thicknesses of blue marls in the Oued Tarhia series;
-
The coeval sedimentation of blue marls, gray marls, and marly-limestones with Neopyctodonte cochlear, suggests a vertical and lateral evolution of the sedimentary facies towards a more or less rich in carbonate supply. This sedimentological context can probably be linked to an environment warm enough to allow the development in this region of warm-temperate faunas towards the end of the Zanclean and the Piacenzian (Belkebir 1986; Satour et al. 2013/2014).
The Dahra Massif and Lower Chelif Basin cross sections
The biostratigraphic results obtained from planktonic foraminifera led us to establish robust correlations (Fig. 18) between the exposed sections or boreholes carried out in other sectors of the Lower Chelif Basin and the Dahra Massif. Indeed, six sections are correlated along a west-east oriented transect, going from the mouth of the Chelif River to Oued Tarhia (Hamri) and materialized by the Djebel Djezzar borehole and sections of Djebel Aizeub (Belhadji et al. 2008), Sidi Brahim Telegraph (Mazzola 1971), Azaizia (a) and Oued Tarhia (the present work). The Oued El Kramis-Kef El Asfar section (Belkebir and Anglada 1985; Belkebir 1986), located on the Hachacha Plateau (north of the Dahra Massif) is also taken into account. Several remarks and observations are made (Fig. 18):

Location and correlation of the Ouled Mallah Pliocene marine successions with those of the Hachacha Plateau (Dahra Massif) and Lower Chelif Basin
-
Although Sphaerodinellopsis subdehiscens is present in the sector of Ouled Maallah, it does not allow the definition of a biozone, due to the lack of its acme. This is associated with Globorotalia margaritae, which suggests the G. margaritae biozone; this suggestion is correlated with that defined at the bottom of the blue marls of the Kef El Asfar section (Belkebir 1986);
-
The base of the marly series is characterized by the presence of Sphaerodinellopsis subdehiscens, downstream of the Chelif Valley, in particular in the Telegraph section of Sidi Brahim, the Djebel Djezzar borehole (Mazzola 1971) and the Djebel Aizeub section (Belhadji et al. 2008), preceding the G. margaritae biozone. On the other hand, these two species are associated in the Azaizia section. This base was not reached (visibility gap) at the Oued Tarhia section and that of Oued Kramis / Kef El Asfar one.
-
The biomarkers defining the Zanclean-Piacenzian boundary are different in this region according to the authors. The latter is marked by Globorotalia crassaformis in the Achacha Plateau (Belkebir and Anglada 1985; Belkebir 1986) or G. aff. crassaformis (present work), G. crotonensis (Mazzola 1971) and G. aemiliana (Belhadji 2005; Belhadji et al. 2008) in the Lower Chelif Basin. It should also be noted that this difference can be explained: Globorotalia crotonensis is considered to be a variant (or synonym) of G. crassula (Hornibrook 1982); it is included in the definition of the G. crassaformis biozone (Belkebir and Anglada 1985). The G. aemiliana biozone is equivalent (or partially) to that of G. crassaformis (Cita et al. 1968; Cita 1975);
-
The transition of the Zanclean to Piacenzian is noticed above a ferruginous level (emersion?) capped by a bio-organic concentration in the Azaizia (a) section or following gypsy marls in the Sidi Brahim Telegraph section;
-
An evidence of two important areas of Zanclean sedimentary subsidence (depocentres), one located at the Sidi Brahim Telegraph and the other at Oued Tarhia. These depocenters are also reported on isopaque maps produced in the Lower Chelif Basin (more than 500 m thick: Perrodon 1957). According to Perrodon (1957), the Pliocene subsidence remains particularly important in the eastern part of the basin. Indeed, thicknesses of blue marls of the order of 500 m are recorded at Aïn Zeft and Belhacel localities, 600 m at Aïn Merane or even 700 m in the region of Ouled Farès, which shows a certain displacement (translation) towards the North of the area of the Pliocene cycle compared with that of the Miocene one (Perrodon 1957);
-
Diverse sedimentary facies were deposited at the same time: white marls or "trubi" facies (Djebel Aizeub, Sidi Brahim Telegraph), blue marls and/or gray marls (Sidi Brahim Telegraph, Azaizia, Oued Tarhia, Oued Kramis-Kef El Asfar), white marly limestone/calcareous marls with Neopycnodonte cochlear (Azaizia, Oued Tarhia sections). These facies reflect a remarkable environmental and paleobiological diversity. In addition, their sedimentary evolution (vertical and spatial) towards carbonate enrichment (platform) characterizes essentially the sector of Ouled Maalah and Oued Tarhia, which in fact concerns the entire southern margin of the Dahra Massif, from Ouled Maallah to the West and to Mazouna and Oued Ouarizane to the East (Perrodon 1957).
Tectonic implications
The Lower Chelif Basin has been affected, since the Middle Miocene, by tectonics allowing the readjustment of the basement, the orographic restructuring, and the filling of the basin with the “onset” of the thrusts (Perrodon 1957; Thomas 1985; Meghraoui 1982; Bessedik et al. 2002; Belkebir et al. 1996). This context continued during the Messinian, characterized by compressive tectonic activity (Brive 1897; Meghraoui 1982; Meghraoui et al. 1986; Thomas 1985; Arab et al. 2015) and/or extension (Thomas 1985). In some localities, the Messinian gypsum and the overlying Lower Pliocene deposits sometimes hide clues allowing the deciphering of this type of deformation.
Near the Cretaceous substratum and the Tortono-Messinian lands of the Dahra Massif, the paleogeographic extent (West–East) of gypsum shows in the Ouled Maallah sector, dips of 20 to 30° to the south (Fig. 19).

Folded Messinian structures of Azaizia (a, b) and Aïn Yacoub sections with Late Messinian-Early Pliocene sediments showing the brutal reflooding and the progressive sedimentary filling during the Late Pliocene as a result of the continuing sea-level rise
The geometry of the gypsum recorded in the sectors of Ouled Mallah and Aïn Zeft reveals a structure showing a dip of around 20–25° towards the south. While the dip of the northern edge of the transgressive marl series over the gypsum is barely 5 to 7 ° to the south; this changes in the center to become subhorizontal (1/50 000 map: Brive 1897). This difference in dip suggests a “cut-and-filled” marly series and a post-gypsum tectonics, sealed by the base of the Mio-Pliocene sediments which is reflective as it has just been precisely illustrated by its morphology (see Fig. 5-B; Fig. 19-B and C) and dated (see Fig. 16).
This difference in dip has long been pointed out by Brive (1897) when speaking of a folded tectonics of gypsum from an age prior to the Pliocene reflooding. These latest Messinian tectonic movements were also highlighted by Reyre (in: Perrodon 1957) in the Aïn Zeft sector, not far from the Ouled Maallah agglomeration.
In this case, the anticline and syncline structures of Azaizia are demonstrative (Fig. 19-A, B, and C). The Pliocene deposits of these folded structures show a post-gypsum compressive tectonics, during the latest Messinian. The sedimentary filling of this structure was carried out during the late Messinian and entire Zanclean: if its bottom (Azaizia a) includes latest Messinian to earliest Zanclean sediments with Globorotalia margaritae and Ceratolitus acutus up to around 5.08 to 5.04 Ma, its margins are reached at around 4.04 Ma according to the presence of Discoaster asymmetricus, i.e., a duration of a million years for the Zanclean sea-level rise after the reflooding (see Fig. 19-A).
In addition, the post-gypsum Lago Mare (Messinian) facies of Ain Yacoub is marked by micritic white limestones, brecciation, and the presence of slumped limestone banks. These facts support the syn-sedimentary tectonic activity and therefore the existence of an initial slope, the source of some gravitational dynamics in sedimentation.
In the Oued Tarhia sector, the outcrops show a dip of about fifteen degrees, or even more in places where the flexure of the reliefs tends to accentuate, when coming in the Lower Chelif Basin. The marly-limestones, forming the cornices, show, in their upper part, figures of overlapping or progradation of the benches, with north vergency, which expresses the existence of a fairly marked topography and therefore a sufficient slope of probably tectonic origin (Messinian?), allowing the establishment of these sedimentary bodies.
In the Djebel Aizeub sector, Belhadji (2005) and Belhadji et al. (2008) described a chaotic gypsum-limestone material consisting of marls, diatomites, and gypsum they attribute to the Messinian on the basis of the Globorotalia mediterranea presence. Being “wedged” between two abnormal contacts, this material is limited at the base by blue Tortonian marls (with Neogloboquadrina acostaensis) and overlain by Pliocene marine marls reported to the Sphaerodinellopsis subdehiscens biozone. The N. humerosa and N. dutertrei (Upper Tortonian) Biozone seems to be absent, a phenomenon probably linked to erosion. This material is interpreted (Belhadji et al. 2008) as a post-gypsum olistolith and correlated with those described in the Beni Chougrane (Rouchy 1982a, b; Rouchy et al. 2007).
So, the El Ghomri section (Beni Chougrane), located in the southern margin of the Lower Chelif Basin (Rouchy 1982a, b; Rouchy et al. 2007) shows the existence of an olistolith made of diatomitic marls bypassed in a post-gypsum Lago Mare deposit. It is sealed by sandy marls attributed to the Lower Pliocene (with Globorotalia margaritae: Rouchy et al. 2007). This is also the case in the Sahaouria section where Rouchy et al. (2007) indicate a comparable phenomenon (olistolith), occupying the same stratigraphic position.
In addition, other tectonic events of the Messinian age are described in the salt lakes of Arzew and Telamine sediments (tectonics in distension: Benbakhti et al. 2018) or in the Oran Bay (submarine discordance linked to the Messinian Erosional Surface (MES): Just et al. 2010). Note that the MES is considered to have affected the gypsum, the Terminal Carbonated Complex (TCC: Esteban 1979), and Lago Mare 1 (i.e., of age 5.60 Ma: Clauzon et al. 2005; Popescu et al. 2015; Bache et al. 2012, 2015).
These data make it possible to envisage a tectonic activity during the latest Messinian, post-gypsum or even post-Lago Mare 1 in the Dahra Massif and the Lower Chelif Basin. This hypothesis was also suggested by Rouchy et al. (1982) and Rouchy et al. (2007) for the Beni Chougrane olistoliths, corresponding to a period of readjustment and reorganization of the basin at the end of the Messinian. Chronologically, this phenomenon corresponds to the isostasic rebound as described in Roussillon by Clauzon et al. (2015b), succeeding to the MSC.
These events achieved the uplift of the coastal massifs (Brive 1897; Perrodon 1957; Rouchy 1982a, b; Rouchy et al. 2007) before the Mio-Pliocene marine reflooding (Clauzon et al. 1996). Indeed, the offshore drilled sections, located in the mouth of the Chelif River (Arab et al. 2015: page 366, Fig. 3) as well as a reflection seismic profile (Sonatrach and Total companies) which provides additional evidence (Fig. 20; Rubino et al. 2010) of the MES, sealing an important latest Messinian compressive deformation.

Seismic profile produced in the Chelif Basin (location: see Fig. 3). The limits of the Lower and Upper Miocene strata (black and green lines) show the existence of a clear deformation sealed by the MES (red line) overlain by the latest Messinian-earliest Zanclean (reflooding) sediments
Conclusion
The biostratigraphic study of six sections (Azaïzia (a), Azaïzia (b), Ain Yacoub, Moul Settara, Oued Tarhia, and Garbous Ben Saada) located in the Ouled Maallah region (Dahra, Algeria) allows the identification of a geological succession starting with Messinian peripheral gypsum (1st step of the MSC), followed by Messinian lacustrine deposits (Lago Mare 1 event) and blue marls of the latest Messinian-Zanclean age, from which they are separated by the MES.
The deposits underlying the gypsum have revealed the presence of Globorotalia mediterranea and Amaurolithus primus associated with A. delicatus and Nicklithus amplificus which mark the Messinian stage (N17/NN11c). This former age is confirmed everywhere on the southern margin of the Dahra Massif, from the Chelif Valley to the West up to Bouzghaia to the East. In addition, the primary gypsum overlain by white micritic limestones is correlated with the “Terminal Carbonated Complex” following the coral reef platforms of the Mudjadjo Massif (Oran), Melilla (Morocco), and that of Sorbas (Spain). These correlations suggest an estimated age of the gypsum ranging from 5.971 (at its base) to 5.60 Ma (at its top), the latter being a chronological landmark which would correspond to the top of the lagoonal deposits of Vera and San Miguel de Salinas (Clauzon et al. 2005; Popescu et al. 2009, 2015; Corbí et al. 2016).
According to the CIES (2008), the gypsum of the Oran area, known as “upper gypsum” (Rouchy and Caruso 2006; Rouchy and Saint Martin 1992; Saint Martin and Rouchy 1990; Cornée et al. 2004), was deposited in a peripheral basin (primary gypsum) before the desiccation (erosional) phase (2nd step, paroxysmal, of the MSC) and the subsequent marine reflooding.
As a consequence, the lacustrine deposits overlying the gypsum can be correlated with the Lago Mare 1, slightly earlier than 5.60 Ma. These lacustrine deposits display a large extent, both on the southern margin of the Dahra and on the northern slopes of the Beni Chougrane whose first studies and correlations were carried out by Rouchy et al. (1982) and Rouchy et al. (2007). The Ouled Maallah sections (Ain Yacoub, Oued Tarhia) complete the information, resulting in a better understanding of this Lago Mare episode as well as the marine reflooding conditions in the region.
The arrival of coarse detrital sediments in the basin (unconformable conglomerates) described in the Oued Tarhia (this study) and the Sahouria sections (Atif 2001; Rouchy et al. 2007; Atif et al. 2008 indicates the instability of the basin margin (tectonic activity or isostatic rebound) during the Lago Mare episode.
The Azaizia (a and b) sections illustrate an important sedimentary body, developed from the Mio-Pliocene to the Late Pliocene and resting unconformably on previously folded and eroded gypsum, describing a cut-and-filled series. The course of erosion is geometrically irregular and appears to be deep and significant in the center, but markedly less expressed at the edges of the Azaizia structure (Fig. 5-B).
The blue marls are characterized by several taxa, notably Sphaerodinellopsis subdehiscens, Globorotalia margaritae, Ceratolithus acutus, Reticulofenestra. This last succession is followed by the occurrence of G. puncticulata, G. bononiensis, G. aff. crassaformis and Helicosphaera sellii, Discoaster asymmetricus, and D. tamalis.
These biomarkers indicate an age ranging from the latest Messinian – earliest Zanclean to the early Piacenzian, corresponding to the N18 p.p., N19, and N20 p.p. biozones (Mazzola 1971; Belkebir 1986). The successive occurrence of the nannoplankton (Ceratolithus acutus, Reticulofenestra cf. cisnerosii, Helicosphaera sellii, Discoaster asymmetricus and D. tamalis) confirms this attribution (NN12 p.p., NN13, NN14/NN15, and NN16).
The calibration of these successions allows us not only to specify or qualify some biostratigraphic attributions, but also to reinforce the association of Sphaerodinellopsis subdehiscens, Globorotalia margaritae, Ceratolithus acutus, and R. cf. cisnerosii as indicators of the topmost Messinian or the base of the Zanclean. However, the R. cisnerosii (= R. zancleana) was found in the basin of San Miguel de Salinas, simultaneously with Ceratolithus acutus, located above the Messinian Erosional Surface (Corbí et al. 2016). Consequently, we retain a terminal Messinian age (5.46 Ma) can be retained since the marine reflooding took place before the beginning of the Zanclean.
The Zanclean-Piacenzian boundary is marked by a gully and covered by a ferruginous level associated with a shelly concentration, preceding the first occurrence of Globorotalia aff. crassaformis. These data reveal the existence in the Lower Chelif Basin of two large subsiding depocentres (Telegraph of Sidi Brahim, Oued Tarhia).
The pre-Pliocene paleotopography appears to be irregular, which can be linked to the paleogeographic and tectonic context, generated by the Messinian Salinity Crisis, i.e., the Messinian Erosional Surface and the impact of isostasic readjustments. Consequently, the Pliocene sedimentation, diversified and varied, seems to be contemporaneous: blue marls, gray marls, and calcareous marls with Neopyctodonte cochlear, which suggest a vertical and lateral evolution of the sedimentary facies towards a more or less rich in carbonate facies, located generally on the central Dahra Massif (Ouled Mallah-Mazouna axis). This contribution can be linked to shallow reliefs and an environment that becomes warm enough to allow the development of the rich fauna and flora recorded in the sector (Belkebir 1986; Satour et al. 2013/2014).
The setting of Mio-Pliocene deposits seals the Messinian Erosional Surface after a folding tectonic phase. This phase affected gypsum and Lago Mare deposits of Oued Maallah. It has also resulted in gravitational tectonics, responsible for the “olistolith” setting within the geological series at the north and south of the Lower Chelif Basin (Dahra and Beni Chougrane massifs). This compressive deformation seems to be far-reaching, having marked the Oran region. It would correspond to a restructuring of paleogeographical spaces associated with the uplift of the Tellian Chain, preceding the post-Messinian Crisis marine reflooding in the region.
References
Abbouda M, Bouhadad Y, Benfedda A, Slimani A (2018) Seismotectonic and seismological aspects of the Mostaganem (Western Algeria) May 22, 2014 (Mw 4.9) seismic event. Arab J Geosci 11(57):1–9. https://doi.org/10.1007/s12517-018-3404-y
Abbouda M, Maouche S, Bouhadada Y, Belhai D (2019) Neotectonic and active tectonics of the Dahra- Lower Cheliff Basin (Tell Atlas, Algeria): seismotectonic implication. Journal of African Earth Sciences 153:250–267. https://doi.org/10.1016/j.jafrearsci.2019.02.023
Haddadi-Hamdane A (1996) Bryozoaires du Pliocène du Sahel d’Alger. Docum Lab Géol Fac Sci Lyon 14:189
Aïfa T, Feinberg H, Derder MEM, Merabet NE (2003) Contraintes magnéto-stratigraphique concernant la durée de l’interruption des communications marines en Méditerrannée occidentale pendant le Messinien supérieur. Geodiversitas 25(4):617–631 http://sciencepress.mnhn.fr/fr/periodiques/geodiversitas/25/4/contraintes-magnetostratigraphiques-concernant-la-duree-de-l-interruption-des-communications-marines-en-mediterrannee-occidentale-pendant-le-messinien-superieur
Ameur-Chehbeur A (1988) Biochronologie des formations continentales du Néogène et du Quaternaire de l’Oranie: contribution des micromammifères. Thèse Doctorat d’Etat, Université d’Oran (Algérie)
Ameur-Chehbeur A (1992) Age accuracy of some Hipparion fossiliferous sites in Algeria. In: Spitz FG, Janeau G, Gonzalez S, Aulagniern, (Eds.). Ongulés/Ungulates, 91, S.F.E.P.M.-I.R.G.M., Paris Toulouse: 27-30
Anderson RVV (1936) Geology in the coastal Atlas of western Algeria. Geol Soc Am Memoir 4, 450 p. https://doi.org/10.1130/MEM4-p1
Arab M, Bracene R, Roure F, Zazoun RS, Mahjoub Y, Badji (2015) Source rocks and related petroleum systems of the Chelif Basin, (western Tellian domain, north Algeria). Marine and Petroleum Geology 64:363–385. https://doi.org/10.1016/j.marpetgeo.2015.03.017
Arambourg C (1927) Les poissons fossiles d’Oran. Matériaux carte géol d’Algérie. Sér 1, Palaeontol 6, 291 p
Atif KFT (2001) Modalités du passage mio-pliocène dans le bassin du Bas Chélif. Mém. Magister, Oran Univesity
Atif KFT, Bessedik M, Belkebir L, Mansour B, Saint Martin JP (2008) Le passage mio-pliocène dans le bassin du Bas Chélif (Algérie). Biostratigraphie et paléoenvironnements. Geodiversitas 30(1):97–116 http://sciencepress.mnhn.fr/fr/periodiques/geodiversitas/30/1/le-passage-mio-pliocene-dans-le-bassin-du-bas-chelif-algerie-biostratigraphie-et-paleoenvironnements
Bache F, Popescu SM, Rabineau M, Gorini C, Suc JP, Clauzon G, Olivet JL, Rubino JL, Dobrinescu MCM, Estrada F, Londeix L, Armijo R, Meyer B, Jolivet L, Jouannic G, Leroux E, Aslanian D, Dos Reis AT, Mocochain L, Dumurdžanov N, Zagorchev I, Lesič V, Tomič D, Çačatay MN, Brun JP, Sokoutis D, Csato I, Ucarkus G, Çakir Z (2012) A two-step process for the reflooding of the Mediterranean after the Messinian Salinity Crisis. Basin Research 24:125–153. https://doi.org/10.1111/j.1365-2117.2011.00521.x
Bache F, Gargani J, Suc JP, Gorini C, Rabineau M, Popescu SM, Leroux E, Do Couto D, Jouannic G, Rubino JL, Olivet JL, Clauzon G, Dos Reis AT, Aslanian D (2015) Messinian evaporite deposition during sea level rise in the Gulf of Lions (Western Mediterranean). Marine and Petroleum Geology 66:262–277. https://doi.org/10.1016/j.marpetgeo.2014.12.013
Backman J, Raffi I, Rio D, Fornaciari E, Pälike H (2012) Biozonation and biochronology of Miocenethrough Pleistocene calcareous nannofossils from low and middle latitudes. Newsletters on Stratigraphy 47(2):221–244. https://doi.org/10.1127/0078-0421/2012/0022
Badji R, Charvis P, Bracene R, Galve A, Badsi M, Ribodetti A, Benaissa Z, Klingelhoefer F, Medaouri M, Beslier MO (2015) Geophysical evidence for a transform margin offshore Western Algeria: a witness of a subduction-transform edge propagator? Geophys J Int 200:1027–1043. https://doi.org/10.1093/gji/ggu454
Bassetti MA, Miculan P, Sierro FJ (2006) Evolution of depositional environments after the end of Messinian Salinity Crisis in Nijar basin (SE Betic Cordillera). Sedimentary Geology 188–189:279–295. https://doi.org/10.1016/j.sedgeo.2006.03.009
Belhadji A (2005) La série miocène supérieur-pliocène du versant oriental de Djebel Diss (Dahra occidentale): attribution biostratigraphique. Mém. Magister Oran University
Belhadji A, Belkebir L, Saint Martin JP, Mansour B, Bessedik M, Conesa G (2008) Apports des foraminifères planctoniques à la biostratigraphie du Miocène supérieur et du Pliocène de Djebel Diss (bassin du Chélif, Algérie). Geodiversitas 30(1):79–96 http://sciencepress.mnhn.fr/fr/periodiques/geodiversitas/30/1/apports-des-foraminiferes-planctoniques-la-biostratigraphie-du-miocene-superieur-et-du-pliocene-de-djebel-diss-bassin-du-chelif-algerie
Belkebir L (1986) Le Néogène de la bordure nord occidentale du massif de Dahra (Algérie). Biostratigraphie, paléoécologie, paléogéographie. Thèse Doctorat, Provence University
Belkebir L, Anglada R, (1985) Le Néogène de la bordure nord-occidentale du massif du Dahra. 110econgr Natl Soc Savantes Sciences Montpellier, 6: 279-290; https://cths.fr/ed/edition.php?id=156
Belkebir L, Mansour B, Bessedik M, Saint Martin JP, Belarbi M, Chaix C (1994) Présence d’une construction récifale corallienne à Djebel Chott (Dahra occidental, Algérie): témoin du maximum transgressif du Miocène moyen en Méditerranée. Géologie Méditerranéenne, Tome 21(1-2):1–7
Belkebir L, Bessedik M, Ameur-Chehbeur A, Anglada R (1996) Le Miocène des bassins nord-occidentaux d’Algérie: biostratigraphie et eustatisme. Elf Aquitaine éditions, Pau 16:553–561
Belkebir L, Bessedik M, Mansour B (2002) Le Miocène supérieur du bassin du Bas Chélif: attribution biostratigraphique à partir des foraminifères planctoniques. Mém Serv Géol Algérie 11:187–194
Belkebir L, Labdi A, Mansour B, Bessedik M, Saint Martin JP (2008) Biostratigraphie et lithologie des séries serravallo-tortoniennes du massif du Dahra et du bassin du Chélif (Algérie). Implications sur la position de la limite serravallo-tortonienne. Geodiversitas 30(1):9–19 http://sciencepress.mnhn.fr/fr/periodiques/geodiversitas/30/1/biostratigraphie-et-lithologie-des-series-serravallo-tortoniennes-du-massif-du-dahra-et-du-bassin-du-chelif-algerie-implications-sur-la-position-de-la-limite-serravallo-tortonienne
Bellon H, Guardia P, Magné J (1984) Les associations volcaniques du Miocène spérieur de la région oranaise (Algérie occidentale). Conséquences géodynamiques. Géol Méditerr. Tome XI 3:255–264
Benbakhti IM, Maouche S, Belhai D, Harbi A, Ritz JF, Rabai G, Rezouk A, Doumaz F (2018) Characterizing the active tectonics in the Oran region (Algeria) and recasting the 1790 earthquake. J Seismol 22:1549–1561. https://doi.org/10.1007/s10950-018-9784-3
Benzina M, Hebib H, Bensalah M (2019) New insights in late Miocene lower Chelif basin biostratigraphy based on planktonic foraminifera (Algeria). Rev de Micropal 62:9–24
Berggren WA, Hilger FJ, Langereis CG, Kent DV, Obradovich JD, Raffi I, Raymo M, Shackleton NJ (1995) Late Neogene chronology: New perspective in high-resolution stratigraphy. GSA Bulletin 107(11):1272–1287. https://doi.org/10.1130/0016-7606(1995)107<1272:LNCNPI>2.3.CO;2
Bessedik M (1984) The Early Aquitanian and Upper Burdigalian-Lower Serravallian environments in the Northwestern Mediterranean region. Paléobiol contin, Montpellier 14(2):153–179 https://www.semanticscholar.org/paper/The-Early-Aquitanian-and-Upper-Langhian-Lower-in-Bessedik/b7ccd9ede687f21cd2a863b30f237daa8049d1f7#related-papers
Bessedik M, Benammi M, Jaeger J J, Ameur-Chehbeur R, Belkebir L, Mansour B (1997) Gisement à rongeurs d’âge tortonien dans des dépôts lagunaires et marins de transition en oranie: corrélation marin continental. Actes du congrès BiochroM’97, Aguilar J P, Legendre S, Michaux J (Eds.), Mém Trav E P H E, Inst. Montpellier, 21:293-300
Bessedik M, Belkebir L, Mansour B (2002) Révision de l’âge miocène inférieur (au sens des anciens auteurs) des dépôts du Bassin du Bas Chélif (Oran, Algérie): conséquences biostratigraphique et géodynamique. Mém Serv Géol Algérie 11:167–186
Bizon G, Bizon JJ (1972) Atlas des principaux foraminifères planctoniques du Bassin méditerranéen Oligocène à Quaternaire. Technip (edit.), Paris, 316 p
Bizon G, Müller C (1976) Flemarks on some biostratigrahic problems in the Mediterranean Neogene. lnt Sympos on the structural history of the Mediterranean bassins, Split (Yugoslavia). Technip (ed.): 381-390
Blair SA, Bergen JA, de Kaenel E, Browning E, Boesiger TM (2017) Upper Miocene-Lower Pliocene taxonomy and stratigraphy in the circum North Atlantic Basin: radiation and extinction of Amauroliths, Ceratoliths and the D. quinqueramus lineage. Journal of Nannoplankton Research. 37(2-3):113–144
Blow WH (1969) Late middle Eocene to recent planctonic foraminiferal biostratigraphy. Proc lst lnt Conf Plankt Microfossils, Genève 1:199–422
Bown PR, Young JR (1998) Techniques. In: Bown PR (ed) Calcareous nannofossil biostratigraphy (British Micropalaeontological Society Publications Series). Chapman and Kluwer Academic, London, pp 16–28
Brive A (1897) La carte géologique de Renault au 1/50 000è, Feuille n° 104 et notice explicative. Edition du Service géographique de l’Armée
Brive A (1920) Sur l’âge des gypses du Dahra. C R somm Soc Géol Fr: 54-55
Bukry D, Bramlette MN (1968) Stratigraphic significance of two genera of Tertiary calcareous nannofossils. Tulane Stud Geol Paleontol 6:149–155
Bukry D, Bramlette MN (1969) Some new and stratigraphically useful calcareous nannofossils of the Cenozoic. Tulane Stud Geol Paleontol 7:131–142
Bukry D (1973) Coccolith and silicoflagellate stratigraphy, Deep Sea Drilling Project Leg 18, eastern North Pacific. In: Kulm LD et al. (eds.): Initial Reports of the DSDP, Washington (U.S. Government Printing Office), 18: 817-831
Bukry D (1975) Silicoflagellate and coccolith stratigraphy, Deep Sea Drilling Project Leg 29. In: Kennett JP et al. (eds.): Initial Reports of the DSDP, Washington (U.S. Government Printing Office), 29: 845-872
Busson G, Magné J (1955) Extension du Miocène Supérieur du Nord du bassin du Chélif. C R somm Soc Géol Ff, Paris: 143-145.
Butler RWH, Lickorish HW, Grasso M, Pedley MH, Ramberti L (1995) Tectonics and sequence stratigraphy in Messinian basins, Sicily: constraints on the initiation and termination of the Mediterranean salinity crisis. Geol Soc Am Bull 107:425–439. https://doi.org/10.1130/0016-7606(1995)107<0425:TASSIM>2.3.CO;2
Butler RWH, McClelland E, Jones RE (1999) Calibrating the duration and timing of the Messinian salinity crisis in the Mediterranean: linked tectonoclimatic signals in thrust-top basins of Sicily. Journal of the Geological Society, London 156:827–835. https://doi.org/10.1144/gsjgs.156.4.0827
Caruso A, Blanc-Valleron MM, Da Prato S, Pierre C, Rouchy JM (2020) The late Messinian “lago-Mare” event and the Zanclean reflooding in the Mediterranean Sea: insights from the Cuevas del Almanzora section (Vera Basin, South-Eastern Spain). Earth-Sci Rev 200(102993):1–20. https://doi.org/10.1016/j.earscirev.2019.102993
Channell JET, Rio D, Thunell RC (1988) Miocene/Pliocene magnetostratigraphy at Capo Spartivento, Calabria, Italy. Geology 16:1096–1099. https://doi.org/10.1130/0091-7613(1988)016<1096:MPBMAC>2.3.CO;2
Channell JET, Rio D, Sprovieri R, Glacon G (1990) Biomagnetostratigraphic correlations from Leg 107 in the Tyrrhenian Sea. In: Kastens KA, Mascle J et al., Proc. ODP, Sci. Results 107:669–682. https://doi.org/10.2973/odp.proc.sr.107.180.1990
Channell J, Di Stefano E, Sprovieri R (1992) Calcareous plankton biostratigraphy, and paleoclimatic history of the Plio-Pleistocene Monte San Nicola Section (Southern Sicily). Bollettino Società Paleontologica Italiana 31:351–382
Chikhi H (1992) Une palynoflore méditerranéenne à subtropicale au Messinien pré évaporitique en Algérie. Géol Méditer, Marseille 19(1):19–30
Ciaranfi N, Dazzaro L, Pieri P, Rapisardi L, Sardella A (1978) Preliminary description of some Messinian evaporitic facies along the Abruzzi–Molise boundary. In: Catalano R, Ruggieri G, Sprovieri R (Eds.): Messinian evaporites in the Mediterranean. Mem Soc Geol Italia, vol 16: 251–260
CIESM (2008) Executive summary. In: The Messinian salinity crisis from mega-deposits to microbiology a consensus report (Ed F. Briand). CIESM Workshop Monographs 33: 7–28. http://www.ciesm.org/online/monographs/Almeria.html
Cita MB (1975) Studi sul Pliocene e egli strati di passagio dal Miocene al Pliocene. VII. Planktonic foraminiferal biozonation of the Mediterranean Pliocene deep-sea record, a revision. Rivista Italiana di Paleontologia e Stratigrafia 81:527–544
Cita MB, Colombo L (1979) Sedimentation in the latest Messinian at Capo Rossello (Sicily). Sedimentology 26:497–522. https://doi.org/10.1111/j.1365-3091.1979.tb00926.x
Cita MB, Vismara-Schilling A (1980) Planktonic-foraminifer assemblages across the Miocene/ Pliocene boundary at Deep Sea Drilling Project sites 415 and 416, and correlations with other North- Atlantic successions. Deep sea Drilling Project initial Reports 50:497–505. https://doi.org/10.2973/dsdp.proc.50.113.1980
Cita MB, Mazzola G, Premoli Silva I (1968) Sur l’âge de la mise en place des nappes telliennes dans la région de Beni Chougrane (Algérie). Instituto Lombardo di Scienze e Lettere 102:16–170
Cita MB, Wright RC, Ryan WBF, Longinelli A (1978a) Messinian paleoenvironments. In: Hsü KJ, Montadert L, et al. (Eds.), Initial Reports Deep Sea Drilling Project, Government Printing Office 42: 1003–1035
Cita MB, Colalongo ML, d’Onofrio S, Laccarino S, Salvatorini G (1978b) Biostratigraphy of Miocene deep sea sediments (Sites 372 and 375) with special reference to the Messinian/pre-Messinian interval. In: Hsü K J, Montadert L, et al. (Eds.), Initial Reports of the Deep Sea Drilling Project, vol. 42. U.S. Govt. Printing Office, Washington: 671–685. https://pdfs.semanticscholar.org/0f49/ca9e4938e4bc2ca0950e785a60ddfbb47ef9.pdf
Clauzon G, Suc JP, Gautier F, Berger A, Loutre MF (1996) Alternate interpretation of the Messinian salinity crisis, controversy resolved? Geology 24:363–366. https://doi.org/10.1130/0091-7613(1996)024<0363:AIOTMS>2.3.CO;2
Clauzon G, Suc JP, Popescu SM, Marunteanu M, Rubino JL, Marinescu F, Malinte MC (2005) Influence of the Mediterranean sea-level changes over the Dacic Basin (Eastern Paratethys) in the Late Neogene. The Mediterranean Lago Mare facies deciphered. Basin Research 17:437–462. https://doi.org/10.1111/j.1365-2117.2005.00269.x
Clauzon G, Suc JP, Do Couto D, Jouannic G, Melinte-Dobrinescu MC, Jolivet L, Quillévéré F, Lebret N, Mocochain L, Popescu SM, Martinell J, Doménech R, Rubino JL, Gumiaux C, Warny S, Bellas SM, Gorini C, Bache F, Estrada F (2015a) New insights on the Sorbas Basin (SE Spain): the onshore reference of the Messinian Salinity Crisis. Mar Petrol Geol 66:71–100. https://doi.org/10.1016/j.marpetgeo.2015.02.016
Clauzon G, Le Strat P, Duvail C, Do Couto D, Suc JP, Molliex S, Bache F, Besson D, Lindsay EH, Opdyke ND, Rubino JL, Popescu SM, U. Haq B, Gorini C (2015b) The Roussillon Basin (S. France): a case-study to distinguish local and regional events between 6 and 3 Ma. Mar and Petroleum Geology 66:18–40. https://doi.org/10.1016/j.marpetgeo.2015.03.012
Colalongo ML, Cremonini G, Farabegoli E, Sartori R, Tampieri R, Tomadin L (1978) Paleoenvironmental study of the “Colombacci” formation in Romagna (Italy): the Cella section. In: Catalano R, Ruggieri G, Sprovieri R (Eds.), Messinian evaporites in the Mediterranean. Mem. Soc. Geol Italia, vol. 16: 197–216. 1976.
Colalongo ML, Di Grande A, D’onofrio S, Giannelli L, Iaccarino S, Mazzei R, Poppi Brigatti MF, Romeo M, Rossi A, Salvatorini G (1979) A proposal for the Tortonian/Messinian boundary. Annale de Géologie des Pays Hellèniques. Tome Hors Série, Fasc. 1: 285-294
Corbí H, Soria JM, Lancis C, Giannetti A, Tent-Manclús JE, Dinarès-Turell J (2016) Sedimentological and paleoenvironmental scenario before, during, and after the Messinian Salinity Crisis: The San Miguel de Salinas composite section (western Mediterranean). Marine Geology 379:246–266. https://doi.org/10.1016/j.margeo.2016.05.017
Cornée JJ, Guieu G, Muller J, Saint-Martin JP (1994a) Mediterranean Messinian carbonate platforms: some controlling factors., Géol. Méditerr XXI, 3-4: 45-48. https://doi.org/10.3406/geolm.1994.1523
Cornée JJ, Saint-Martin JP, Conesa G, Muller J (1994b) Geometry, palaeoenvironments and relative sea-level (accommodation space) changes in the Messinian Murdjadjo carbonate platform (Oran, western Algeria): consequences. Sed. Geol. 89(1-2):143–158. https://doi.org/10.1016/0037-0738(94)90087-6
Cornée JJ, Saint-Martin JP, Conesa G, Münch P, André JP, Saint Martin S, Roger S (2004) Correlations and sequence stratigraphic model for Messinian carbonate platforms of the western and central Mediterranean. Int. J. Earth Sci (Geol Rundsch) 93:621–633. https://doi.org/10.1007/s00531-004-0400-0
Cornée JJ, Ferrandini M, Saint Martin JP, Münch P, Moullade M, Ribaud-Laurenti A, Roger S, Saint Martin S, Ferrandini J (2006) The late Messinian erosional surface and the subsequent reflooding in the Mediterranean: new insights from the Melilla–Nador basin (Morocco). Palaeogeography, Palaeoclimatology, Palaeoecology 230:129–154. https://doi.org/10.1016/j.palaeo.2005.07.011
Cosentino D, Federici I, Cipollari P, Gliozzi E (2006) Environments and tectonic instability in central Italy (Garigliano Basin) during the late Messinian Lago–Mare episode: new data from the onshore Mondragone 1 well. Sedimentary Geology 188–189:297–317. https://doi.org/10.1016/j.sedgeo.2006.03.010
Cunningham KJ, Collins LS (2002) Controls on facies and sequence stratigraphy of an upper Miocene carbonate ramp and platform, Melilla basin, NE Morocco. Sedimentary Geology 146(3-4):285–304. https://doi.org/10.1016/S0037-0738(01)00131-2
Cunningham KJ, Benson RH, Rakic-El Bied K, McKenna LW (1997) Eustatic implications of Late Miocene depositional sequences in the Melilla Basin, northeastern Morocco. Sedimentary Geology 107:147–165. https://doi.org/10.1016/S0037-0738(96)00037-1
Dalloni M (1915) Recherches sur la période néogène dans l’Algérie occidentale. Bull Soc Géol Fr, Paris, 4e s., t. XV: 428-457.
De Deckker P, Chivas AR, Shelley JMG (1988) Palaeoenvironment of the Messinian Mediterranean “Lago Mare” from strontium and magnesium in ostracod shells. Palaios: 352–358.
Deflandre G, Fert C (1954) Observations sur les Coccolithophoridés actuels et fossiles en microscope ordinaire et electronique. Ann Paleontol 40:117–176
Delfaud J, Michaux J, Neurdin J, Revert J (1973) Un modèle paléogéographique de la bordure méditerranéenne: évolution de la région oranaise (Algérie) au Miocène supérieur. Conséquences stratigraphiques. Bull Soc Hist Nat Afr Nord, Alger 64:219–241
Delteil J (1974) Tectonique de la chaine alpine en Algérie d’après l’étude du Tell oranais (Monts de la Mina, Beni Chougrane, Dahra). Thèse Doctorat Sci, Nice University
Derder MEM, Henry B, Amenna M, Bayou B, Maouche S, Besse J, Abtout A, Boukerbout H, Bessedik M, Bourouis S, Ayache M (2011) Tectonic evolution of the active “Chelif” Basin (Northern Algeria) from paleomagnetic and magnetic fabric investigations. In: New frontiers in tectonic research at the midst of plate convergence Intech Publisher Book, pp. 3–26. Intech (2011). ISBN 978-953-307-594-5
Derder MEM, Henry B, Maouche S, Bayou B, Amenna M, Besse J, Bessedik M, Belhai D, Ayache M (2013) Transpressive tectonics along a major E–W crustal structure on the Algerian continental margin: Blocks rotations revealed by a paleomagnetic analysis. Tectonophysics 593:183–192. https://doi.org/10.1016/j.tecto.2013.03.007
Di Stefano A, Sturiale G (2010) Refinements of calcareous nannofossil biostratigraphy at the Miocene/Pliocene boundary in the Mediterranean region. Geobios 43:5–20
Do Couto D, Popescu SM, Suc JP, Melinte-Dobrinescu MC, Barhoun N, Gorini C, Jolivet L, Poort J, Jouannic G, Auxietre JL (2014) Lago Mare and the Messinian salinity crisis: evidence from the Alboran Sea (S. Spain). Marine and Petroleum Geology 52:57–76. https://doi.org/10.1016/j.marpetgeo.2014.01.018
El Azzouzi M, Maury RC, Fourcade S, Coulon C (2003) Evolution spatiale et temporelle du magmatisme néogène de la marge septentrionale du Maghreb: manifestation d’un détachement lithosphérique. Notes et Mém. Serv Géol Maroc 447:107–116
Esteban M (1979) Significance of the Upper Miocene coral reefs of the western Mediterranean. Palaeogeography, Palaeoclimatology, Palaeoecology 29:169–188. https://doi.org/10.1016/0031-0182(79)90080-4
Faranda C, Gliozzi E, Ligios S (2007) Late Miocene brackish Loxoconchidae (Crustacea, Ostracoda) from Italy. Geobios 40:303–324. https://doi.org/10.1016/j.geobios.2006.11.001
Fenet B (1975) Recherche sur l’alplinisation de la bordure septentrionale du bouclier africain à partir de l’étude d’un élément de l’orogène nord-maghrébin: les monts du Djebel Tessala et les massifs du littoral oranais. Thèse Doctrat ès sciences, Nice, Insti. Polytechnique méditerranéen: 31 p
Fenet B, Irr F (1973) Observations sur le Pliocène inférieur et moyen de la région des Andalouses (littoral oranais, Algérie). C R Acad Sci, Paris 276:2761–2764
Ficheur E (1896) Réunion extraordinaire de la Société géologique en Algérie. Note sur le bassin tertiaire de Médéa. Bull Soc Géol Fr, Paris, 3 sér XXIV:1042–1047
Fortuin AR, Kelling JMD, Roep TB (1995) The enigmatic Messinian–Pliocene section of Cuevas del Almanzora (Vera Basin, SE Spain) revisited, erosional features and strontium isotope ages. Sedimentary Geol 97:177–201
Fortuin AR, Krijgsman W, Hilgen FJ, Sierro FJ (2000) Late Miocene Mediterranean desiccation: topography and significance of the “Salinity Crisis” erosion surface on-land in southeast Spain: comment. Sedimentary Geology 133:167–174. https://doi.org/10.1016/S0037-0738(00)00040-3
Gartner S Jr (1967) Calcareous nannofossils from Neogene of Trinidad, Jamaica, and Gulf of Mexico. Kansas Univ Paleontol Contr 29:1–7
Gartner S Jr (1969) Correlation of Neogene planktonic foraminifer and calcareous nannofossil zones. Gulf Coast Assoc Geol Soc Trans 19:585–599
Gartner S, Bukry D (1974) Ceratolithus acutus Gartner and Bukry n. sp. and Ceratolithus amplificus Bukry and Percival — nomenclatural clarification. Tulane Stud Geol Paleontol 11:115–118
Gautier F, Clauzon G, Suc JP, Cravatte J (1994) Age et durée de la crise de salinité messinienne. C R Acad Sci, Paris 318(2):1103–1109
Geerlings LPA, Dronkert H, Van de Poel HM, Van Hinte JE (1980) Chara sp. in Mio-Pliocene marls at Cuevas del Almanzora, Vera Basin, Spain. Proc. K. Ned. Akad. Wet B 83:29–37
Gignoux M (1936) Géologie stratigraphique, 2°édition. Masson, Paris
Gourinard Y (1952) Chaines telliennes oranaises. Un livret-guide de l’excursion croisière organisée dans la région littorale d’Algérie et de Tunisie. Publi XIXe Congr Géol Intern, Alger,: 109-115
Gourinard Y (1958) Recherches sur la géologie du littoral oranais. Publ. Serv. Carte géol. Algérie, nouvelle Sér, 6, 111 p
Gradstein FM, Ogg JG, Schmitz MD, Ogg GM (2012) The Geologic Time Scale. Vol 1, 1st edit, Elsevier. Publisher: 413 p.
Guardia P (1975) Géodynamique de la marge alpine du continent africain d’après l’étude de l’Oranie nord-occidentale. Thèse Doctorat d’Etat, Nice University
Guardia P (1976) Carte géologique de l’Oranie nord occidentale au 1/100 000è. Nice University, CRGM
Harbi H, Maouche S, Ayadi A (1999) Neotectonics and associate seismicity in the Eastern Tellian Atlas of Algeria. J Seismol 3(1):95–104. https://doi.org/10.1023/A:1009743404491
Hebib H (2014) La limite tortono-messinienne dans la marge nord du bassin du bas Chélif. Précisions biostratigraphiques et évolution des assemblages de foraminifères benthiques. Thèse Doctorat Sci, Oran 2 University.
Hilgen FJ, Lourens LJ, Van Dam JA (2012) The Neogene period. In: Gradstein et al edit. The geological Time scale 2012. 1st edit Elsevier BV. Publisher: 923-977.
Hornibrook N d B (1982) Late Miocene to Pleistocene Globorotalia (Foraminiferida) from DSDP Leg 29, Site 284, Southwest Pacific. New Zealand Journal of Geology and Geophysics 25(1):83–99. https://doi.org/10.1080/00288306.1982.10422507
Hsü KJ, Ryan BW, Cita MB (1973) Late Messinian desiccation of the Mediterranean. Nature 242:240–244
Iaccarino SM, Premoli Silva I, Biolzi M, Foresi LM, Lirer F, Turco E, Petrizzo MR (2007) Practical manual of Neogene planktonic foraminifera. International School on Planktonic Foraminifera, 6th course, Ferugia 19-23: 141 p.
Jaeger JJ (1977) Les rongeurs du Miocène moyen et supérieur du Maghreb. Palaevertebrata, Montpellier, 8 (1): l-166.
Just J, Hübscher C, Betzler C, Lüdmann T, Reicherter K (2010) Erosion of continental margins in the Western Mediterranean due to sea-level stagnancy during the Messinian Salinity Crisis. Geol Mar Lett 31:51–64. https://doi.org/10.1007/s00367-010-0213-z
Kamptner E (1967) Kalkfalgellaten-Skelettreste aus Tiefseeschlamm des Sudatlantik. Ann Naturhist Mus Wien 71:117–198
Krijgsman W, Hilgen FJ, Raffi I, Sierro FJ, Wilson DS (1999) Chronology, causes and progression of the Messinian salinity crisis. Nature 400:652–655
Krijgsman W, Fortuin AR, Hilgen FJ, Sierro FJ (2001) Astrochronology for the Messinian Sorbas basin (SE Spain) and orbital (precessional) forcing for evaporite cyclicity. Sedim Geol 140:43–60
Lancis C, Flores JA (2006/2007) A new biostratigraphically significant calcareous nannofossil species in the Early Pliocene of the Mediterranean. Micropaleontology 52(5):477–481. https://doi.org/10.2113/gsmicropal.52.5.477
Lancis C, Tent-Manclús JE, Flores JA, Soria JM (2015) The Pliocene Mediterranean infilling of the Messinian Erosional Surface: New biostratigraphic data based on calcareous nannofossils (Bajo Segura Basin, SE Spain). Geol Acta 13(3):211–228. https://doi.org/10.1344/GeologicaActa2015.13.3.4
Langereis CG, Hilgen FJ (1991) The Capo Rossello composite: a Mediterranean and global reference section for the early to early late Pliocene. Earth Planet Sci Lett 104(2-4):211–225. https://doi.org/10.1016/0012-821X(91)90205-V
Leprêtre R, de Lamotte DF, Combier V, Gimeno-Vives O, Mohn G, Eschard R (2018) The Tell-Rif orogenic system (Morocco, Algeria, Tunisia) and the structural heritage of the southern Tethys margin. BSGF Earth Sciences Bulletin 189(10):1–35. https://doi.org/10.1051/bsgf/2018009
Lirer F, Foresi LM, Iaccarino SM, Salvatorini G, Turco E, Cosentino C, Sierro FJ, Caruso A (2019) Mediterranean Neogene planktonic foraminifer biozonation and biochronology. Earth-Science Reviews 196(102869):1–36. https://doi.org/10.1016/j.earscirev.2019.05.013
Louni-Hacini A, Bellon H, Maury RCh, Megartsi M, Coulon Ch, Semroud B, Cotten J, Coutelle A (1995) Datation 40K-40Ar de la transition du volcanisme calco-alcalin au volcanisme alcalin en Oranie au Miocène supérieur. C R Acad Aci Paris, 321, sér IIa: 975-982.
Lourens LJ, Hilgen FJ, Shackleton NJ, Laskar J, Wilson D (2004) The Neogene Period. In: Gradstein FM, Ogg JG, Smith AG (Eds.), A geologic time scale 200, 21. Cambridge University Press: 409–440.
Lourens LJ, Sluijs A, Kroon D, Zachos JC, Thomas E, Rohl U, Bowles J, Raffi I (2005) Astronomical pacing of late Palaeocene to early Eocene global warming events. Nature 435(23):1083–1087. https://doi.org/10.1038/nature03814
Lucas G (1942) Description géologique et pétrographique des Monts de Ghar Rouban et du Sidi el Abed (Frontière algéro-marocaine). Bull Serv Carte géol Algérie, Sér 2, Stratigraphie, n°16: 538 p.
Mahboubi S (2014) Les rongeurs du Miocène supérieur et terminal d’Afrique du Nord-occidentale: biochronologie, manétostratigraphie, biogéographie et paléoenvironnements. Thèse Doctorat, Poitiers University.
Mahboubi S, Bennami, M, Jaeger JJ (2015) New datation of the Tafna Basin (Algeria): a combination between biochronological and magnetostratigraphical data. Palaevertebrata, Montpellier, 39(1-e1): 1-11.
Mansour B (2004) Diatomées messiniennes du bassin du bas Chélif (Algérie Nord-occidentale). Thèse doctorat Sci, Oran University.
Mansour B, Saint Martin JP (1999) Conditions de dépôt des diatomites messiniennes en contexte de plateforme carbonatée d'après l'étude des assemblages de diatomées: Exemple du Djebel Murdjadjo (Algérie). Geobios 32(3):395–408. https://doi.org/10.1016/S0016-6995(99)80016-3
Mansour B, Moissette P, Noel D, Rouchy JM (1995) L'enregistrement par les associations de diatomées des environnements messiniens: L'exemple de la coupe de Sig (bassin du Chélif-Algérie). Geobios 28(3):261–279. https://doi.org/10.1016/S0016-6995(95)80001-8
Mansour B, Bessedik M, Saint Martin JP, Belkebir L (2008) Signification paléoécologique des assemblages de diatomées du Messinien du Dahra sud occidental (bassin du Chélif, Algérie nord-occidentale). Geodiversitas 30(1):117–139
Mansouri MEH (2001) Les nannofossiles calcaires du Miocène supérieur du Bassin du Chélif. Intérêt biostratigraphique. Mém Magister, Oran University.
Mansouri MEH, Bessedik M, Aubbry MP, Belkebir L, Mansour B, Beaufort L (2008) Contribution biostratigraphiques et paléo environnementales de l’étude des nannofossiles calcaires des dépôts tortono-messiniens du bassin du Chélif (Algérie). Geodiversitas 30(1):59–77 http://sciencepress.mnhn.fr/fr/periodiques/geodiversitas/30/1/contributions-biostratigraphiques-et-paleoenvironnementales-de-l-etude-des-nannofossiles-calcaires-des-depots-tortono-messiniens-du-bassin-du-chelif-algerie
Manzi V, Gennar R, Hilgen F, Krijgsman W, Lugli S, Roveri M, Roveri FJ (2013) Age refinement of the Messinian salinity crisis onset in the Mediterranean. Terra Nova 25(4):315–322. https://doi.org/10.1111/ter.12038
Maouche S, Bouhadad Y, Harbi A, Rouchiche Y, Ousadou F, Ayadi A (2019) Active tectonics and seismic hazard in the Tell Atlas (Northern Algeria): a review. A. Bendaoud et al. (eds.), in: The geology of the Arab world-an overview. Springer Geology: 381-400. https://doi.org/10.1007/978-3-319-96794-3_10
Martin JM, Braga JC (1994) Messinian events in the Sorbas basin in southeastern Spain and their implications in the recent history of the Mediterranean. Sedimentary Geology 90(3/4):257–268. https://doi.org/10.1016/0037-0738(94)90042-6
Martini E (1971) Standard Tertiary and Quaternary calcareous nannoplankton zonation. Proceedings of the 2nd Planktonic Conference Roma, 2: 739–785.
Mattauer M (1958) Etude géologique del’Ouarsenis oriental (Algérie). Bulletin du Service de la Carte géologique de l’Algérie, Alger. Nouvelle série n° 17: 534 p.
Mazzola C (1971) Les foraminifères planctoniques du Mio-Pliocène de l’Algérie nord-occidentale. Proceeding on the Second International Conference on the Planktonic Microfossils, Roma, 2: 787-818.
Megartsi M (1985) Le volcanisme mio-plio-quaternaire de I’Oranie. Thèse doctorat Sci, Algiers University.
Meghraoui M (1982) Etude néotectonique de la région nord-est d’El-Asnam: relation avec le séisme du 10 octobre 1980. Thèse doctorat 3° cycle, Paris XII University.
Meghraoui M, Cisternas A, Philip H (1986) Seismotectonic of the Lower Cheliff Basin: Structural background of the El Asnam (Algeria) Earthquake. Tectonics 5(6):809–836. https://doi.org/10.1029/TC005i006p00809
Meghraoui M, Philip H, Albarède F, Cisternas A (1988) Trenches investigations through the trace of the 1980 El-Asnam thrust fault: evidence for paleoseismicity. Bull Seismol Soc Amer 78(2):979–999 https://www.researchgate.net/publication/281263571
Moissette P (1988) Faunes de Bryozoaires du Messinien d’Algérie occidentale. Docum. Lab. Géol. Lyon, 102: 351 p.
Morigi C, Negri A, Giunta S, Kouwenhoven T, Krijgsman W, Blanc-Valleron MM, Orszag-Sperber F, Rouchy JM (2007) Integrated quantitative biostratigraphy of the latest Tortonian-early Messinian Pissouri section (Cyprus): An evaluation of calcareous plankton bioevents. Geobios 40:267–279
Müller C (1974) Calcareous nannoplankton, Leg 25 (Western Indian Ocean). In: Initial Reports of the Deep Sea Drilling Project, vol 25. U.S. Government Printing Office, Washington, pp 579–633
Negri A, Villa G (2000) Calcareous nannofossil biostratigraphy, biochronology and paleoecology at the Tortonian/Messinian boundary of the Faneromeni section (Crete). Palaeogeography, Palaeoclimatology, Palaeoecology 156:195–209
Neurdin-Trescartes J (1992) Le remplissage sédimentaire du bassin néogène du chélif , modèle de référence de bassins intramontagneux. Thèse doctorat ès-Sci, Pau & Pays de l’Adour University, 1: 332 p.
Neurdin-Trescartes J (1995) Paléogéographie du Bassin du Chélif (Algérie) au Miocène. Causes et conséquences. Géol Méditer 22(2):61–71
Okada H, Bukry D (1980) Supplementary modification and introduction of code numbers to the low-latitude coccolith biostratigraphic zonation (Bukry, 1973; 1975). Marine Micropaleontology 5:321–325. https://doi.org/10.1016/0377-8398(80)90016-X
Orszag-Sperber F (2006) Changing perspectives in the concept of “Lago-Mare” in Mediterranean Late Miocene evolution. Sedimentary Geology 188(189):259–277. https://doi.org/10.1016/j.sedgeo.2006.03.008
Orszag-Sperber F, Rouchy JM, Elion P (1989) The sedimentary expression of regional tectonic events during the Miocene-Pliocene transition in the Southern Cyprus basins. Geological Magazine 126(3):291–299. https://doi.org/10.1017/S001675680002238X
Orszag-Sperber F, Rouchy JM, Blanc-Valleron MM (2000) La transition Messinien-Pliocène en Méditerranée orientale (Chypre): la période du Lago-Mare et sa signification. C R Acad Sci Paris, Sci Terre et Planètes 331:483–490. https://doi.org/10.1016/S1251-8050(00)01433-6
Ouda K, Ameur A (1978) Contribution of the biostratigraphy of the Miocene sediments associated with primitive Hipparion fauna of Bouhanifia, North West Africa. Revista Española Micropaleontol 10(3):407–420
Perch-Nielson K (1985) Cenozoic calcareous nannofossils. In: Plankton stratigraphy. Boli HM, Saunders JB, Perch-Nielsen K Edits. Cambridge University Press, Cambridge: 427-554.
Perrodon A (1957) Etude géologique des bassins néogènes sublittoraux de l’Algérie occidentale. Bulletin du Service de la Carte Géologique de l’Algérie, Alger.12:1-382.
Pierre C, Caruso A, Blanc-Valleron MM, Rouchy JM, Orzsag-Sperber F (2006) Reconstruction of the paleo environmental changes around the Miocene–Pliocene boundary along a West–East transect across the Mediterranean. Sedimentary Geology 188(189):319–340. https://doi.org/10.1016/j.sedgeo.2006.03.011
Poignant A, Moissette P (1992) Foraminifères benthiques des intercalations marneuses d’une série diatomitique du Messinien d’Algérie. N.Jb. Geel. Paliiont. Mh., H. 11, Stutgart: 657-674.
Polvèche J (1959) Contribution à l'étude géologique de l’Ouarsenis oranais. Thèse Doctorat ès-Sci, Lille 1 University: 707 p.
Pomel A (1889) Description stratigraphique générale de l’Algérie. Fontana P édit, Alger: 211p.
Pomel A (1892) Sur la classification des terrains miocènes de l’Algérie et réponses aux critiques de M. Peron. Bull Soc géol Fr, 96, sér 3, 20: 166-174.
Popescu SM, Melinte-Dobrinescu MC, Suc JP, Clauzon G, Quillévére F, Suto-Szentai M (2007) Earliest Zanclean age for the Colombacci and uppermost Di Tetto formations of the “latest Messinian” northern Apennines: new palaeoenvironmental data from the Maccarone section (Marche Province, Italy). Geobios 40:359–373. https://doi.org/10.1016/j.geobios.2006.11.005
Popescu SM, Melinte-Dobrinescu MC, Dalesme F, Sütö-Szenta M, Jouannic G, Bakra K, Escarguel G, Clauzon G, Head M, Suc JP (2009) Galeacysta etrusca complex, dinoflagellate cyst marker of Paratethyan influxes into the Mediterranean Sea before and after the peak of the Messinian Salinity Crisis. Palynology 33(2):105–134. https://doi.org/10.1080/01916122.2009.9989688
Popescu SM, Dalibard M, Suc JP, Barhoun N, Melinte-Dobrinescu MC, Bassetti MA, Deaconu F, Head MJ, Gorini C, Do Couto D, Rubino JL, Auxietre JL, Floodpage J (2015) Lago Mare episodes around the Messinian-Zanclean boundary in the deep southwestern Mediterranean. Marine and Petroleum Geology 66:55–70. https://doi.org/10.1016/j.marpetgeo.2015.04.002
Popescu SM, Melinte-Dobrinescu MC, Suc JP, Do Couto D (2017) Ceratolithus acutus Gartner and Bukry 1974 (= C. armatus Müller 1974), calcareous nannofossil marker of the marine reflooding that terminated the Messinian salinity crisis: Comment on “Paratethyan ostracods in the Spanish Lago-Mare: More evidence for interbasinal exchange at high Mediterranean sea level” by Stoica et al., 2016. Palaeogeogr Palaeoclimatol Palaeoecol 441: 854–870. Palaeogeogr Palaeoclimatol Palaeoecol 485: 986-989. https://doi.org/10.1016/j.palaeo.2016.07.011
Raffi I, Flores JA (1995) Pleistocene through Miocene calcareous nannofossils from eastern equatorial Pacific Ocean (Leg 138). In: Pisias NG, Mayer LA, Janecek TR, Palmer-Julson A, Van Andel TH (Eds.), Proceedings of the Ocean Drilling Program, Scientific Results, Vol. 138: 233-286.
Raffi I, Mozzato C, Fornaciari E, Hilgen FJ, Rio D (2003) Late Miocene calcareous nannofossil biostratigraphy and astrochronology for the Mediterranean region. Micropaleontology 49:1–26
Raffi I, Backman J, Fornaciari E, Pälike H, Rio D, Lourens L, Hilgen F (2006) A review of calcareous nannofossil astrobiochronology encompassing the past 25 million years. Quatern Sci Rev 25:3113–3137. https://doi.org/10.1016/j.quascirev.2006.07.007
Rakic El Bied K, Benson RH (1996) La stratigraphie à Haute Résolution: Théorie et application au Néogène supérieur du Maroc. Notes et Memoires du Service Géologique du Maroc 383:5–50
Riding R, Braga JC, Martin JM, Sanchez-Almazo IM (1998) Mediterranean Messinian salinity crisis: constraints from a coeval marginal basin, Sorbas, southern Spain. Marine Geology 146:1–20. https://doi.org/10.1016/S0025-3227(97)00136-9
Rouchy JM (1980) La genèse des évaporites messiniennes de Méditerranée: un bilan. Bull Cent Rech Pau-SNPA 4:511–545
Rouchy JM (1982a) La crise évaporitique messinienne de méditerranée: nouvelles propositions pour une interprétation génétique. Bull Mus nation Hist natur, Paris, c (3-4): 1- 52.
Rouchy JM (1982b) La genèse des évaporites Messiniennes de Méditerranée. Mém Muséum nat Hist natur, Paris, Sér C, Sciences de la Terre (L):1-280.
Rouchy JM, Caruso A (2006) The Messinian salinity crisis in the Mediterranean Basin: a reassessment of the data and an integrated scenario. Sediment. Geol. 188/189:35–67. https://doi.org/10.1016/j.sedgeo.2006.02.005
Rouchy JM, Saint Martin JP (1992) Late Miocene events in the Mediterranean as recorded by carbonate-evaporite relations. Geology 20(7):629–632. https://doi.org/10.1130/00917613(1992)020<0629:LMEITM>2.3.CO;2
Rouchy JM, Orszag-Sperber F, Bizon G, Bizon JJ (1980) Mise en évidence d’une phase d’émersion fini-messinienne dans le bassin de Pissouri, Chypre: une modalité de passage Miocène–Pliocène en Méditerranée orientale. C R Acad Sci, Paris 291:729–732
Rouchy JM, Chaix C, Saint Martin JP (1982) Importance et implications de l’existence d’un récif corallien messinien sur le flanc sud du Djebel Murdjadjo (Oranie Algérie). C R Acad Sci Paris 294:813–816
Rouchy JM, Pierre C, Et-Touhami M, Kerzazi K, Caruso A, Blanc-Valleron MM (2003) Late Messinian to early Pliocene changes in the Melilla Basin (NE Morocco) and their relations to Mediterranean evolution. Sediment Geol 163:1–27. https://doi.org/10.1016/S0037-0738(03)00157-X
Rouchy JM, Caruso A, Pierre C, Blanc-Valleron MM, Bassetti MA (2007) The end of the Messinian salinity crisis: evidences from the Chelif Basin (Algeria). Palaeogeography, Palaeoclimatology, Palaeoecology 254:386–417. https://doi.org/10.1016/j.palaeo.2007.06.015
Roveri M, Gennari R, Lugli S, Manzi V (2009) The Terminal Carbonate Complex: the record of sea-level changes during the Messinian salinity crisis. GeoActa 8:63–77
Roveri M, Manzi V, Bergamasco A, Falcieri FM, Gennari R, Lugli S, Schreiber BC (2014a) Dense shelf water cascading and Messinian canyons: A new scenario for the Mediterranean salinity crisis. American Journal of Science 314:751–784. https://doi.org/10.2475/05.2014.03
Roveri M, Flecker R, Krijgsman W, Lof J, Lugli S, Manzi V, Sierro FJ, Bertini A, Camerlenghi A, De Lange C, Govers R, Hilgen FJ, Hübscher C, Meijer PT, Stoica M (2014b) The Messinian salinity crisis: past and future of a great challenge for marine sciences. Mar Geol 352:25–58. https://doi.org/10.1016/j.margeo.2014.02.002
Roveri M, Gennari R, Lugli S, Manzi V, Minelli N, Reghizzi M, Riva A, Rossi ME (2016) The Messinian salinity crisis: Open problems and possible implications for Mediterranean petroleum systems. Petroleum Geoscience 22:283–290. https://doi.org/10.1144/petgeo2015-089
Roveri M, Lugli S, Manzi V, Reghizzi M, Rossi FP (2020) Stratigraphic relationships between shallow-water carbonates and primary gypsum: insights from the Messinian succession of the Sorbas basin (Betic Cordillera, Southern Spain). Sedim Geol 404(105678):1–18. https://doi.org/10.1016/j.sedgeo.2020.105678
Rubino JL, Haddadi N, Camy-Peyret J, Clauzon G, Suc JP, Gorini C (2010) Messinian Salinity Crisis expression along North African Margin. Cairo (Egypt) SPE Conf. 129526-PP: 1-4.
Ruggieri G (1962) La serie marine pliocenica e quaternaria della Val Marecchia. Atti Acad Sci Lett Arti Palermo 19:1–169
Ruggieri G, Sprovieri R (1974) The lacustrine faunas in Sicily and the desiccation theory of the Messinian Salinity Crisis. Lavori Inst Geol Univ Palermo 13:3–6
Ruggieri G, Sprovieri R (1976) Messinian salinity crisis and its paleogeographical implications. Palaeogeogr Palaeoclimatol Palaeoecol 20:13–21
Sadran G (1952) Les roches cristallines du littoral Oranais. Publ 19è Congr Géol Intern, Alger. Monographies régionales, 1ère sér, n° 18: 1-84.
Sadran G (1958) Les formations volcaniques tertiaires et quaternaires du Tell oranais. Publ Serv Carte Géol Algérie, nouv Sér 18:1–533
Saint Martin JP (1990) Les formations récifales coralliennes du Miocène supérieur d’Algérie et du Maroc. Mém Mus nat Hist nat Paris 56:1–366
Saint Martin JP, Rouchy JM (1990) Les plates-formes carbonatées messiniennes en Méditerranée occidentale: leur importance pour la reconstitution des variations du niveau marin au Miocène terminal. Bull Soc géol Fr (8), VI, 1: 83-94.
Saint Martin JP, Chaix C, Moissette P (1983) Le Messinien récifal d’Oranie (Algérie): une mise au point. C R Acad Sc Paris 297(2):545–547
Saint Martin JP, Cornée JJ, Conesa G, Bessedik M, Belkebir L, Mansour B, Moissette P, Anglada R (1992) Un dispositif particulier de plate-forme carbonatée messinienne: la bordure méridionale du bassin du Bas-Chelif Algérie. C R Acad Sci, Paris 2(315):1365–1372
Saoudi N (1982) Pliocène et Pléistocène inférieur et moyen du Sahel d’Alger. Thèse 3è Cycle, Aix-Marseille III University, 277 p.
Satour L, Lauriat-Rage A, Belkebir L, Bessedik M (2013/2014) Biodiversity and taphonomy of bivalve assemblages of the Pliocene of Algeria (Bas Chelif Basin). Arab J Geosci 7:5295–5308. https://doi.org/10.1007/s12517-013-1154-4
Snel E, Mărunţeanu M, Meulenkamp JL (2006) Calcareous nannofossils biostratigraphy and magnetostratigraphy of the Upper Miocene and Lower Pliocene of the Northern Aegean (Orphanic Gulf-Strymon Basin areas), Greece. Palaeogeogr Palaeoclim Palaeoecol 238:125–150. https://doi.org/10.1016/j.palaeo.2006.03.022
Sprovieri R (1993) Pliocene-early Pleistocene astronomically forced planktonic foraminifera abundance fluctuations and chronology of Mediterranean calcareous plankton bio-events. Rivista italiana di paleontologia e stratigrafia 99(3):371–414. https://doi.org/10.13130/2039-4942/8903
Sprovieri R, Sprovieri M, Caruso A, Pelosi N, Bonom, S, Ferraro L (2006) Astronomic forcing on the planktonic foraminifera assemblage in the Piacenzian Punta Piccola section (southern Italy). Paleoceanography 21, PA4204: 1-21. https://doi.org/10.1029/2006PA001268.
Suc JP, Do Couto D, Melinte-Dobrinescu MD, Macaleţ R, Quillévéré F, Clauzon G, Csato I, Rubino JL, Popescu SM (2011) The Messinian Salinity Crisis in the Dacic Basin (SW Romania) and early Zanclean Mediterranean–Eastern Paratethys high sea-level connection. Palaeogeogr Palaeoclimatol Palaeoecol 310(3/4):256–272. https://doi.org/10.1016/j.palaeo.2011.07.018
Suc JP, Popescu SM, Do Couto D, Clauzon G, Rubino JL, Melinte-Dobrinescu MC, Quillévéré F, Brun JP, Dumurdžanov N, Zagorchev V, Lesić V, Tomićţ D, Sokoutis D, Meyer B, Macalet R, Rifelj H (2015) Marine gateway vs. fluvial stream within the Balkans from 6 to 5 Ma. Marine and Petroleum Geology 66:231–245. https://doi.org/10.1016/j.marpetgeo.2015.01.003
Tchouar L (2013) Etude des dinoflagellés de la série mio-pliocène du Télégraphe de Sidi Brahim (Bassin du Chélif, Algérie nord-occidentale): Systématique et Paléoécologie. Magister thesis, Oran 2 University; 1-111.
Theodoridis S (1984) Calcareous nannofossil biozonation of the Miocene and revision of the helicoliths and discoasters. Utrecht Micropaleontological Bulletin 32:271 p
Thomas G (1985) Géodynamique d’un bassin intramontagneux. Le bassin du Bas Chélif occidental durant le mio-plio-quaternaire. Thèse Doctorat d’Etat, Pau & Pays de l’Adour University: 1-594.
Thunell RC (1979) Climatic evolution of the Mediterranean Sea during the last 5.0 million years. Sediment Geol 23:67–79
Van Couvering JA, Castradori D, Cita MB, Hilgen FJ, Rio D (2000) The base of the Zanclean Stage and of the Pliocene Series. Episodes 23(3):179–187
Yassini I (1973) Nouvelles données stratigraphiques sur la limite Pliocène inférieur- Pliocène moyen (Plaisancien-Aslien) dans la région d’Alger. Rev Micropaleontol 16(4):229–248
Yassini I (1975) Planktonic foraminiferal biozonation of Neogene deposits in the “Sahel” of Algiers, Algeria. Rivista Ital Paleontol 81(1):89–120
Zachariasse WJ (1975) Planktonic foraminiferal biostratigraphy of the late Neogene of Crete (Greece). Utrecht Micropaleont Bull 11:1–171
Acknowledgments
This study was performed within the framework of the doctoral training of 3rd Cycle “Geology of Marine and Continental Environments: Integrated Stratigraphy, Chronology and Dynamics of Paleoenvironments.”
This work is carried out thanks to the support of the DGRSDT. It fits into the CNEPRU (n° E04N01UN310220150001) and PRFU (E0401UN310320200001) projects of the Ministry of Higher Education and Scientific Research. Jean-Pierre Suc and the anonymous reviewers are thanked for their suggestions, English corrections, and substantial enrichments of the manuscript. The authors wish to thank Sonatrach and Total companies for allowing us to use the seismic profile produced in the Chelif basin. The local authorities of Ouled Maallah were of great help to us in our numerous trips in the field. The authors warmly thank M. Miliani and F-Z Bessedik for their invaluable assistance.
Author information
Authors and Affiliations
Corresponding author
Additional information
Responsible editor: Marina Rabineau
Rights and permissions
About this article

Cite this article
Osman, M.K., Bessedik, M., Belkebir, L. et al. Messinian to Piacenzian deposits, erosion, and subsequent marine bioevents in the Dahra Massif (Lower Chelif Basin, Algeria). Arab J Geosci 14, 684 (2021). https://doi.org/10.1007/s12517-021-06481-0
Received:
Accepted:
Published:
DOI: https://doi.org/10.1007/s12517-021-06481-0