
Abstract
Identifications of the benthic foraminiferal assemblages (Gavelinella, Stensioeina, and Brotzenella) from one stratigraphic section measured from the platform of the uppermost Turonian-upper Campanian successions of Gebel Duwi Range, Quseir Region, allowed the recognition of seven biozones and two subzones. The Duwi Basin through the uppermost Turonian to upper Campanian increased in depth, the benthic components indicating a time of the Egyptian stable continental shelf. The gradual increase in depth had occurred with no dramatic structure or tectonic changes. The benthic foraminiferal content reveals significant correlates of biostratigraphical and paleoecological changes of the same interval through Eastern, Western, and Central European provinces, and Tethyan epicontinental facies. The benthic foraminiferal contents are of diversified epibenthic and endobenthic taxa suggesting epicontinental inner to outer shelf depositional paleoenvironments and mesotrophic habitat.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Calcareous benthic foraminifera are important tools to date the Cretaceous platform carbonates, where the ammonites and/or planktonic foraminifera are absent or very rare. However, benthic foraminifera are sensitive to environmental changes and, therefore, are facies dependent (Gusic et al. 1988; Velic and Vlahovic 1994; Cause et al. 2003).
The stratigraphic ranges of Late Cretaceous benthic foraminifera may not correspond to their true evolution (appearance-extinction), and they should be used for biostratigraphic correlations with great care, even in environments of long-lasting carbonate platforms (Gusic et al. 1988; Cause et al. 2003). Although several benthic foraminiferal stratigraphic distributions and biozonations have been suggested especially for local areas, a refined, standard biozonation, which can be applied for the Mediterranean region, has not been able to be created for the Upper Cretaceous up till now due to the limitations mentioned above.
Walker and Miller (1992) illustrated that the epifaunal morphotypes are only able to live within a well-oxygenated marine environment. However, infaunal morphotypes can live in an oxygen-deficient environment. Aerobic taxa include morphogroups with a low surface-area-to-volume ratio, while anaerobic taxa have a high surface-area-to-volume ratio (Kaiho 1991). The morphogroup analysis of calcareous benthic assemblages is widely applied for paleoenvironmental studies (e.g., Koutsoukos and Hart 1990; Frenzel 2000; Setoyama et al. 2011, 2017).
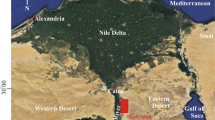
This paper aims to establish detailed benthic foraminiferal zonation of the Upper Cretaceous (uppermost Turonian-upper Campanian) epicontinental to neritic sediments of Gebel Duwi Range, Quseir Region (Fig. 1), and correlate it with the adjacent epicontinental platforms.

Geological map of Gebel Duwi Range (modified after Khalil and McClay 2002)
The basic idea of morphogroup analysis of the foraminiferal fauna in the present study in the Duwi basin allowed us to do some further and more detailed paleoecological investigations.
Geological setting
The study area, 500 km south of Suez and 10 km inland from the Red Sea, along Quseir-Qeft Road, contains the largest preserved remnants of Late Cretaceous to Early Tertiary cover along the Egyptian Red Sea coast. These 600-m-thick platform sediments are preserved as outliers down faulted along trends typical of the Eastern Desert (Said 1962, 1990) (Fig. 1).
Fault trends are most prominent in the northwest-trending, 40-km-long Gebel Duwi fault block and appear to represent reactivated Precambrian structures (Khalil and McClay 2002). Red Sea–related structures began to disrupt the area in Oligocene to early Miocene time. The Cenozoic faulting hierarchy involves a pattern of northeast-tilted, northwest-trending blocks. Tilting was followed by an eastward shift of tectonic activity to form the main Red Sea basin (Akkad and Noweir 1980). A post-tilting regional erosion surface developed during mid-Miocene quiescence and has since suffered minor disruption. Possible continuations of the same tilted fault block structural style controlled by basement grain should be of interest for offshore petroleum exploration.
Upper Cretaceous epicontinental basins of the southern platform of Egypt, not deeper than 100–150 m, accumulated predominantly siliciclastic and carbonate sediment facies of marls, calcareous clays, limestones, phosphate granules, and chert. The interval of uppermost Turonian-upper Campanian is about 21 m thick of condensed phosphorite bearing succession represented by the Atcham phosphate (A-Beds) informal member of the Duwi Formation (Said 1962, 1990).
The Duwi Formation is one of the important stratigraphic units in the geology of the Cretaceous Epoch in Egypt. It is considered a marker unit of the Central and Southern Egypt. It was laid down in extremely shallow epicontinental-neritic seas which flaked the southern margin of the Tethys Ocean in Egypt (Kerdany and Cherif 1990; Glenn and Arthur 1990; El Gammal and Orabi 2019a).
Lithostratigraphy
Gebel Duwi area comprises siliciclastic Nubian facies overlain by the following formations: Quseir (Cenomanian), Duwi (upper Cenomanian-upper Campanian), Dakhla (upper Campanian-Lower Paleocene), Tarawan (Middle to Upper Paleocene), Esna (Upper Paleocene-Lower Eocene), and Thebes (Lower Eocene) (Issawi et al. 1969).
The Duwi Formation attains 77 m thick at its type locality (Gebel Duwi Range), where Said (1962, 1990) classified locally the Duwi Formation on the basis of its mineable phosphate beds into Lower Hamadat unit (or Hamadat or C-Beds), Middle Duwi phosphate Bed (or B-Beds), and Upper Phosphate Atcham Beds (or A-Beds). El Gammal and Orabi (2019a, 2019b) studied in detail the lithobiostratigraphy of Quseir Fm. and Duwi Fm. (C-Beds and B-Beds) and assigned these formations to Cenomanian-upper Turonian, and the same authors assigned the Atcham unit (A-Beds) to upper Turonian-lower Campanian (Fig. 2).


Latest Turonian-Late Campanian benthic foraminiferal species distribution chart, upper part of the Duwi Formation, Gebel Duwi, Egypt. Benthonic foraminifera zones from older to younger: 1: Gavelinella vesca/Gavelinella ammonites zone; 2: Gavelinella praeinferasantonica/Protostensioeina granulata zone; 3: Gavelinella thalmanni/Stensioeina exsculpta zone; 4: Gavelinella vombensis/Gavelinella praestelligera/Stensioein perfecta zone, a subzone: Gavelinella compressa/Stensioeina perfecta/Neoflabellina suturalis, b subzone: Gavelinella praestelligera/Stensioeina gracilis; 5: Gavelinella clementiana/Stensioeina pommerana zone; 6: Brotzenella monterelensis/Gavelinella costulata zone; 7: Brotzenella taylorensis zone
Lithologically, the Atcham unit (A-Beds) is 20 m thick and consists of marly limestone, marl, shale beds, and a thin dolomitic oyster band near the top. The Atcham unit is capped by a conglomeratic phosphatic bed (50 cm thick); it marks the top of the all Duwi Formation section, which is overlying the Dakhla Formation.
Materials and methods
About 80 samples were collected from 21-m-thick A-Beds (Atcham Beds) at decimeter intervals, with closer sampling across the Coniacian, Santonian, and Campanian strata of the Duwi Formation at the Gebel Duwi stratigraphic section and examined to establish the biostratigraphic framework to date and detect depositional environments of the epicontinental basin of the Duwi Region (Fig. 2). The samples were disaggregated in water with diluted H2O2, washed through a 63-μm sieve, and dried at 50 °C. All the representative specimens were mounted on microslides for permanent record and identification.
Benthic foraminiferal biozonation (Fig. 2) is established based on the first occurrence (FO) of the taxa by the rules recommended by the North American Stratigraphic Code (NASC 1983) and International Stratigraphic Guide (Murphy and Salvador 1994).
Picked benthic specimens are grouped, identified, and scanned (SEM micrographs) using a scanning electron microscope of Alexandria University (JSM-IT200, Series Jeol) (Figs. 3, 4, and 5). The original samples are preserved with the author’s collections and deposited at the Menoufia University museum collection.

Scale bar = 100 μm. (1a, b, c, d, e, f) Gavelinella ammonoides, (2a, b, c, d, e) G. clementiana, (3a, b, c) G. costulata, (4a, b) G. costata, (5) G. compressa, (6) G. dakotensis, (7) G. kelleri, (8a, b, c, d) G. lorneiana, (9a, b, c, d) G. pertusa, (10a, b) G. praeinfrasantonica, (11) G. sandidgei, (12a, b) G. praestelligera, and (13a, b, c, d) G. stelligera

Scale bar = 100 μm. (1) Gavelinella thalmanni, (2a, b, c, d) G. tumida, (3a, b) G. vesca, (4a, b, c) G. vombensis, (5) Protostensioneina granulate, (6a, b) Stensioneina exsculpta, (7a, b) S. gracilis, (8) S. perfecta, (9a, b) S. pommerana, (10a, b) Anomalina henbesti, (11a, b) A. umbonifera, (12) Anomalinoides nakkadyi, (13) Berthelina intermedia, (14a, b) Brotzenella monterelensis, (15a, b) B. taylorensis, (16) Chilostomella ovoidea, (17a, b) Cibicidoides eriksdalensis, (18) Cibicides cooensis, (19) C. micrus, and (20) Clavulina plummerae

Scale bar = 100 μm. (1a, b, c, d) Discorbis pseudoscopos, (2a, b, c) Gyroidinoides globosus, (3a, b) Lenticulina muensteri, (4) L. pseudomamilligera, (5) L. pseudosecans, (6a, b) L. rotulata, (7) L. secans, (8) L. cf. subangulata, (9) L. evoluta, (10) Nuttalinella florealis, (11) N. ripleyensis, (12) Lenticulina pondi, (13a, b) Planulina austinana, (14) P. texana, (15a, b) Sitella colonensis, (16a, b) Valvulineria californica var. appressa, (17a, b) V. californica var. obesa, (18) Dentalina solvate, (19) Dorothia bulletta, (20) Ellipsoglandulina compacta, (21) E. ellisi, (22) Ellipsonodosaria decurta, (23) Frondocularia cordata, (24) Neoflabellina buticula, (25) N. suturalis, (26a, b, c, d) Eouvigerina aspera, (27) Gaudryina austinana, (28) G. rugose, (29a, b) Reusella pseudospinulosa, (3) Vaginulina cretacea, (31) Nodosaria alternistriata, (32) N. incostans, (33) N. pauperecula, (34) N. limonensis, (35a, b) Loxostomum cushmani, (36a, b) Marginulina austinana, (37a, b) Orthokarstenia clarki, (38) Praebulimina ovulum, (39a, b) P. prolixa longa, (40a, b) Pseudovigerina cretacea, (41) Reusella spinulosa, (42) Saracenaria triangularis, (43) Spiroplectamina henryi, and (44) S. laevis
Benthic foraminiferal biostratigraphy
The present work provides and establishes 7 benthic biozones and 2 subzones for the first time in the area under consideration, which correlated with the planktonic foraminiferal zonation of El Gammal and Orabi (2019a, 2019b) (Fig. 2). The suggestion benthic foraminiferal zones can be correlated with the same observation of Benyamovskiy (2008a, 2008b) and Dubicka and Peryt (2014) in Eastern, Western, and Central European provinces including Russian and south Russian basins.
The main benthic foraminiferal markers represented by the Gavelinella, Stensioeina, and Brotzenella genera at the Duwi section support the obtained stratigraphic subdivisions and correlation with other Tethyan sections. In general, benthic foraminiferal assemblages are flooded almost uniform through most of the stages and substages of epicontinental, evolved, diversity, and comparisons (Hart et al. 1989; Dubicka and Peryt 2014; Peryt and Dubicka 2015).
Both Gavelinella and Stensioeina genera allowed us to establish 7 benthic foraminiferal zones, where these genera have a wide biogeographical distribution covering Eastern and Western European provinces, Ukraine, Russia, southern Russia, and others (Koch 1977; Hart et al. 1989; Olfer'ev and Alekseev 2003; Walaszczyk et al. 2004; Kopaevich et al. 2007; Olferev et al. 2007; Bailey et al. 2009; Dubicka and Peryt 2012, 2014; Bragina et al. 2016; Dubicka 2016; Dubicka et al. 2017; Vishnevskaya et al. 2018).
Benthic foraminifera of the studied interval of the Duwi Formation are rich, well preserved, and diverse of both Gavelinella and Stensioeina genera which are characterized by the complete condensed sedimentary record and correlated with the comparable planktonic foraminiferal standard zones (Fig. 2). The composition of the benthic foraminifers is very similar across the entire epicontinental European basins and Gebel Duwi section (i.e., southern Tethyan epicontinental basin) giving an opportunity of direct correlations between regions.
Gavelinella vesca/Gavelinella ammonoides zone
Category
Concurrent range zone
Stage
Upper Turonian
Definition
Assemblage of Gavelinella vesca and G. ammonoides as the first appearance of upper Turonian Marginotruncana schneegansi zone and overlying the oyster banks of the Duwi FM.
Association
The nominate zone plus Berthelina intermedia, Gavelinella dakotensis, and Sitella coloensis co-occurred with the planktonic Marginotruncana schneegansi, M. paraconcavata, Planoheterohelix moremani, and Dicarinella canaliculate assemblage.
Remarks
Benyamovskiy (2008a) named this interval partially as Gavelinella ammonoides/G. moniliformis zone (LC4) of upper Turonian of the Eastern European Province (EEP). Gavelinella ammonoides (Reuss) is the upper Turonian form in eastern and western European provinces (Dubicka and Peryt 2014). Berthelina intermedia (Berthelin) is widely recorded worldwide from Albian and upper Turonian sediments (Tyszka 2006), as well as Gavelinella vesca (Bykova), which is of upper Turonian-Coniacian in age (Dubicka and Peryt 2014).
Gavelinella praeinfrasantonica/Protostensioeina granulata zone
Category
Concurrent range zone
Stage
Lower Coniacian
Definition
First occurrence (FO) of Gavelinella praeinfrasantonica, Protostensioeina granulata, and Gavelinella lorneiana at the lowermost base of the Coniacian of Duwi section.
Association
Gavelinella vesca, G. kelleri, and Berthelina intermedia
Remarks
Dubicka and Peryt (2014) introduced Protostensioeina as a new genus of the distinct trend in the evolution of the Stensioeina spp., which is known from the Turonian to mid-Santonian of the European epicontinental basin. This zone is characterized by the first occurrence of rich Gavelinella kelleri, G. lorneiana, and G. praeinfrasantonica. The first occurrence of Protostensioeina granulata in the Duwi material referred to the first occurrence of Turonian age as suggested by Dubicka and Peryt (2014). Edwards (1981) considered Gavelinella praeinfrasantonica was evolved from the Gavelinella vombensis lineage. The determination of the basal Coniacian boundary based on benthic foraminifera is problematic here due to very rare benthic species at the uppermost Turonian of the Duwi Formation.
Gavelinella thalmanni/Stensioneina exsculpta zone
Category
Concurrent interval zone
Stage
Upper Coniacian
Definition
First occurrence of Gavelinella thalmanni and Stensioeina exsculpta, Praebulimmina ovulum, and Sitella colonensis within the upper Coniacian interval of the Duwi Formation. The upper limit of this zone is marked by the first appearance of Neoflabellina suturalis as marked by the Coniacian/Santonian boundary (El Gammal and Orabi 2019a).
Association
This zone contains Gavelinella vesca, G. praeinfrasantonica, G. lorneiana, G. ammonoides, Berthelina intermedia, Protostensioeina granulate, Clavulina plummerae, some nodosariids, Lenticulina evoluta, L. rotulata, and L. cf. subangulata (Fig. 2).
Remarks
Koch (1977), Bailey et al. (1983), Hart et al. (1989), Dubicka and Peryt (2014), and others detected the presence of Stensioneina exsculpta in the middle Coniacian of epicontinental beds of Europe. Many transitional forms of S. exsculpta occurred in the upper Santonian (Dubicka and Peryt 2014); in the present work, the same observation is recognized in the Duwi section.
Gavelinella vombensis/Gavelinella praestelligera/Stensioeina perfecta zone
Category
Concurrent zone
Stage
Santonian
Definition
First occurrence of Gavelinella vombensis, G. praestelligera, G. compressa, Stensioeina perfecta, and Neoflabellina suturalis
Association
Gavelinella sandidgei, G. tumida, G. thalmanni, Cibicidoides eriksdalensis, Gaudryina austinana, G. rugosa var. spinulosa, Anomalina henbesti, Cibicides cooensis, Valvulineria californica appressa, var. californica obessa, Chilostomella ovoidea, and Lenticulina pseudosecans. The lower limit of this zone is marked by the first appearance of Neoflabellina suturalis of the Coniacian/Santonian boundary (Fig. 2).
Remarks
Olferev et al. (2007) used both Gavelinella stelligera and Stensioeina pommerana as a partial zone of upper Santonian. Dubicka in Walaszczyk et al. (2016) used the Gavelinella vombensis/Stensioeina perfecta zone and the Gavelinella praeinfrasantonica zone as the middle Santonian of extra Carpathian of Poland. Vishnevskaya et al. (2018) recorded this interval as the Gavelinella stelligera zone covering upper Santonian. All intervals of both Stensioeina exsculpta and Neoflabellina suturalis are the most accurate of the Santonian Stage of the Duwi section. Neoflabellinids have a wide geographical distribution in the Tethys, Boreal realm, North America, Russian platform, Western Carpathians, European provinces, Northern Spain (Lamolda et al. 2014), and Tunisia in Northern Africa (Elamri et al. 2018). Neoflabellina suturalis is considered here as the first best proxy zonal species for the Coniacian/Santonian boundary.
This zone can be locally divided into the following subzones as:
Lower subzone (a)
Gavelinella compressa/Stensioeina perfecta/Neoflabellina suturalis as first appearance case of lower to middle Santonian
Upper subzone (b)
Gavelinella praestelligera/Stensioeina gracilis (as first appearance taxon within the interval)
Gavelinella clementiana/Stensioeina pommerana zone
Category
Concurrent interval zone
Stage
Lower Campanian
Definition
First occurrence of Gavelinella clementiana and Stensioeina pommerana with the extinctions of Gavelinella vombensis and Stensioeina perfecta of the lower zone
Association
Stensioeina gracilis, Lenticulina evoluta, L. muensteri, L. secans, Planulina austinana, P. texana, Reussella spinulosa, Vaginulina cretacea, Valvulineria californica obessa, and V. californica appressa (Fig. 2).
Remarks
Olfer'ev and Alekseev (2003), Olferiev and Alekseev 2005), Benyamovskiy (2008b), Dubicka in Walaszczyk et al. (2016), and Vishnevskaya et al. (2018) used Gavelinella clementiana, Stensioeina pommerana, and Sten. gracilis as marker species covering the early Campanian interval.
This interval zone is characterized by high flooding of the first occurrence Orthokarstenia clarki which is considered a good marker of the early Campanian of the Duwi section as well as the first occurrence of Euvigerina aspera (Bragina et al. 2016). The uppermost boundary of this zone is characterized by the extinction of most Gavelinella spp. as G. Pertusa and G. tumida.
The first occurrence of Gavelinella clementiana and Stensioeina pommerana was associated with extinctions of all dicarinellids (Dicarinella concavata and D. asymetrica) at the same level of the first occurrence of Globotruncanita elevata, together marking the Santonian/early Campanian boundary event (Fig. 6). This result is in agreement with Dubicka et al. (2017) at Bocieniec, Southern Poland, and Eastern European Platform (Vishnevskaya et al. 2018).

Sea-level changes vs. planktonic and benthic foraminiferal biozones, chronozones and biostratigraphic events, Coniacian-late Campanian succession of the Duwi Fm., Gebel Duwi, Egypt. Time scale adopted after assuming a constant sedimentation rate, Tethyan planktonic foraminiferal zones. Peaks and troughs on the regional sea-level curves of Egypt according to Haq et al. (1987) and Lüning et al. (1998)
Brotzenella monterelensis/Gavelinella costulata zone
Category
Concurrent zone
Stage
Middle-upper Campanian
Definition
First occurrence of Gavelinella costulata and Brotzenella monterelensis and first occurrence of Neoflabellina buticula and Frondicularia cordata.
Association
First occurrence of Neoflabellina buticula and Frondicularia cordata; Gavelinella clementiana, Stensioeina gracilis, S. pommerana, Gyroidinoides globosus, Lenticulina muenstri, L. spachholtz Nodosaria paupercula, Loxostomum cushmani, Nuttallinella florealis, N. ripleyensis, Orthokarstenia clarki, Planulina austinana, P. texana, Praebulimina prolixa, Saracenaria triangularis, Spiroplectammina henryi, S. laevis, Vaginulina cretacea, Anomalina nakkadyi, A. umponifera, Cibicides micrus, Discorbis pseudosecans, Dentalina solvate, Dorothia bulletta, Ellipsoglandulina compacta, E. ellisi, and Ellipsonodosaria decurta (Fig. 2).
Remarks
There is general agreement among all authors to consider the Brotzenella monterlensis as marker taxon of the lowermost upper Campanian of the Boreal realm and Tethyan provinces (Hart et al. 1989; Olfer'ev and Alekseev 2003; Olferiev and Alekseev 2005; Olferev et al. 2007; Benyamovskiy 2007; Walaszczyk et al. 2016; Vishnevskaya et al. 2018). The Neoflabellina buticula is known from the upper Campanian layers of Western and Eastern European provinces (Koch 1977).
Brotzenella taylorensis zone
Category
Interval zone of the nominate taxon
Stage
Upper Campanian
Definition
First occurrence of Brotzenella taylorensis (Carsey)
Association
Neoflabellina buticula, Anomalina nakkadyi, Gyroidinoides globosus, and Stensioeina pommerana
Remarks
Olfer'ev and Alekseev (2003), Olferiev and Alekseev 2005), Naidin and Beniamovskiy (2006), Benyamovskiy (2007), Olferev et al. (2007), and Vishnevskaya et al. (2018) are in agreement to consider the Brotzenella taylorensis taxon as marking the upper Campanian. Brotzenella taylorensis was widespread in the Maastrichtian sediments of the East European province (Benyamovskiy 2008a). Neoflabellina buticula is known from the upper Campanian of Western and Eastern European provinces (Koch 1977). The disappearance of Gavelinella clementiana is considered a criterion for defining the Campanian/Maastrichtian boundary (Odin and Lamaurelle 2001; Beniamovski et al. 2012). Gyroidinoides globosus is characteristic and significant species of stratigraphic importance for upper Campanian (Bragina et al. 2016) and the Brotzenella taylorensis zone may be correlated with the planktonic Globotruncana aegyptiaca zone.
Foraminiferal morphogroups
Benthic foraminifera are sensitive to environmental changes which can be reflected in the composition of the assemblages. The distribution, abundance, and even evolution of the benthic foraminifera are controlled by several parameters such as depth, the flux of organic nutrients to the seafloor, oxygen levels of the bottom, salinity, food quality and quantity, substrate, temperature, and others (Koutsoukos and Hart 1990; Nagy 1992; Frenzel 2000; Nagy et al. 2000; Setoyama et al. 2011; Setoyama et al. 2017; Stojanova and Petrov 2018).
Because most of our samples are closely spaced, we focus on general trends rather than changes within each interpreted foraminiferal biozone. The Atchan Member or A-Beds of the upper beds of the Duwi Formation at Gebel Duwi section yielded 4 agglutinated genera and more than 30 calcareous benthic genera.
Calcareous foraminifera morphogroups
A morphogroup scheme for calcareous benthic foraminifera used in this study is after Stojanova and Petrov (2018). The greatest variety of test morphologies is observed in the Duwi section, where 6 morphogroups were described.
-
1.
Rounded trochospiral morphogroup (RT)
It includes trochospiral species and broadly rounded periphery: Valvulineria, Berthelina, Anomalina, and Planulina. This morphogroup comprises epifaunal active herbivores, detrivores, omnivores, and bactrivores. Planulina and Valvulineria inhabited inner shelf to upper slope environments, i.e., deep to middle shelf environments (Olsson 1988). Valvulineria and abundant Gavelinella are considered as the strongest opportunistic fauna (Tyszka 2009).
-
2.
Plano-convex trochospiral morphogroup (PT)
It has trochospiral tests, having a flat umbilical side and narrowly rounded to sharp periphery: Cibicidoides, Cibicides, Discorbis, Gavelinella, Protostensioeina, Stensioeina, Brotzenella, and Nuttallinella. This morphogroup is typical aerobic and eutrophic to mesotrophic environments, and most of these taxa are epifaunal forms. These taxa have been identified in the middle-outer shelf to upper slope deposits (Coccioni and Galeotti 1993; Widmark 1997; Grunert 2011). Berthelina and Gavelinella are common examples of this group.
-
3.
Lenticular morphogroup (L)
Species from this group display biconvex morphology with sharply angled or keeled periphery: Lenticulina and Saracenaria. This morphogroup is dominated by active deposit feeders, representing eutrophic to mesotrophic conditions and epifaunal to deep infaunal microhabitats. These taxa can be found in different environmental conditions, from sublittoral to upper bathyal and aerobic to dysaerobic conditions (Widmark 1997; Rückheim et al. 2006).
-
4.
Tapered and cylindrical morphogroup (T/C)
Rectilinear and straight uniserial, biserial, and triserial tests are included in this morphogroup. Species from this group display a round, oval, or triangular cross section, and parallel or subparallel sides such as Nodosaria, Dentalina, Sitella (Buliminella), Chilostomella, Ellipsoglandulina, Loxostomum, Marginulia, Ellipsonodosaria, Reussella, Pseudovigerina, Vaginulina, Eouvigerina, Orthokarstenia, and Praebulimina. Life position shows that they are shallow to deep infaunal, deposit feeders, and live in the neritic to upper bathyal, aerobic to dysaerobic and mesotrophic to eutrophic environment conditions (Rückheim et al. 2006; Grunert 2011). This morphogroup seems to be indicative of a well-oxygenated environment and seems to have preferred meso-eutrophic living conditions. Elongated multiserial forms are commonly found in areas with increased organic flux (Nagy et al. 1995, 1997).
-
5.
Flattened tapered morphogroup (FT)
The taxa of this morphogroup are characterized by an ovate to compressed cross section, and parallel to subparallel sides, and include uniserial, biserial, and palmate tests; they are represented by Frondicularia and Neoflabellina. This morphogroup is characterized by shallow infaunal detrivores and scavengers’ taxa (Stojanova and Petrov 2018).
-
6.
Agglutinated morphogroup
The agglutinated foraminiferal taxa are rare throughout the section such as some sporadic Gaudryina, Dorothia, Clavulina, and Spiroplectammina. The agglutinated morphogroup includes the elongated or sub-cylindrical test shape; these taxa are shallow to deep infaunal and active deposit feeders with mesotrophic to eutrophic conditions, which are present in a wide range of marine environments from inner shelf to upper bathyal (Rückheim et al. 2006). This assemblage appears to be moderately low levels of oxygen. The presence of large-sized agglutinants and low species diversity indicates constant temperature and stagnant water conditions (Haynes 1981).
Result and discussion
1. Distribution of the morphogroups and genera: Most of the benthic foraminiferal fauna that is present in Duwi Basin exists today in the shelf to upper bathyal environments. The dominating morphogroups in the most representative Duwi section (T/C, PT, RT, RP, and FT) are recorded predominantly in modern marine basins (Murray 1991; Khare et al. 1995; Alperin et al. 2011) and ancient seas (Frenzel 2000; Szydło 2005; Reolid et al. 2008; Setoyama et al. 2011, 2017) .
Assemblages in a mesotrophic, well-oxygenated environment would be characterized by a mixture of forms occupying different habitats and different feeding strategies since both organic matter and oxygen levels are high enough to support all the epifaunal, shallow, and deep infaunal forms. Various studies show that these patterns in the model can be observed in recent benthic foraminiferal assemblages (Jorissen et al. 1995; Kaminski et al. 1995; Gooday and Rathburn 1999; Heinz et al. 2001; Fontanier et al. 2005).
The population of the Duwi benthic fauna is dominated by epifaunal morphotypes of rounded trochspiral, plano-convex trochspiral, biconvex trochospiral/planispiral, flattened tapered, lenticular and tapered and cylindrical, besides agglutinated forms. The relative abundance of morphogroups in assemblages reflect environmental changes through time (Corliss 1985; Jones and Charnock 1985).
2. Uppermost Turonian-lower Santonian: Uppermost Turonian and lower Coniacian characterized by low-diversity assemblages. Several authors have studied the behavior of Recent benthic foraminiferal faunas under extreme conditions (food, oxygen), and the results show that under oxygen stress, low-diversity assemblages tend to occur, characterized by strong dominance of few species (Van der Zwaan and Jorissen 1991; Barmawidjaja et al. 1991; Sjoerdsma and Van der Zawan 1992; Sen Gupta and Machain-Castillo 1993).
The presence of Gavelinella, Cibicidoides, Gaudryina, Anomalina, Valvulineria, Chilostomella, and Lenticulina in the Santonian sediments. These morphogroups (PT, RT, T/C, and L) are found in aerobic and eutrophic to mesotrophic environments. Most of these taxa are epifaunal forms and active deposit feeders, and have been identified in the middle-outer shelf to upper slope deposits (Coccioni and Galeotti 1993; Widmark 1997; Grunert 2011). Valvulineria inhabited deep to middle shelf environments (Olsson 1988), where elongated Chilostomella is found in areas with increased organic flux (Nagy et al. 1995, 1997).
Benthic foraminiferal assemblages of the lower Santonian are characterized by abundant agglutinated foraminifera (Gaudryina austinana, G. rugosa var. spinulosa). These results refer to an increase in paleodepth from an inner shelf to an outer shelf during this interval (Nagy et al. 1995, 1997).
The abundant phosphatic grains in the upper part of the Santonian marl layers support high productivity and/or some tolerance to subsurface oxygen depletion (Polivoda et al. 2011).
In upper Coniacian, Santonian, and lower Campanian, Lenticulina lives within a wide range of depths in the substrate because the genus tolerates a wide range of microhabitats from epifaunal to deep infaunal (Murray 1991; Tyszka 1994).
3. Lower Campanian: The benthic foraminiferal assemblages Orthokarstenia, Stensioeina, and Lenticulina are rich and well-preserved in the lower Campanian indicating further deepening of the Duwi Basin. The dominant C/T morphogroup seems to be indicative of a well-oxygenated environment and has preferred meso-eutrophic trophic conditions. Elongated multiserial forms are commonly found in areas with increased organic flux (Nagy et al. 1995, 1997).
4. Middle Campanian: The flooded of Orthokarstenia clarki in the overall number of middle Campanian species indicate mesotrophic to more oligotrophic environmental conditions (Nagy et al. 1995, 1997). This indicates an open marine, probably outer shelf to slightly upper bathyal environment up to the late Campanian of the benthic Brotzenella taylorensis zone.
In the middle-upper Campanian of the Duwi section (Brotzenella monterelensis/Gavelinella costulata zone), the benthic foraminferal assemblages are of low diversity with Spiroplectammina henryi, S. laevis, and Dorothia bulletta. This assemblage is typical of restricted environments with dysoxic conditions. Shallow infaunal foraminifera (Dentalina, Nodosaria) live in depths of less than 5 cm (Olóriz et al. 2006). The test form is generally elongated. Very low numbers of infaunal foraminifers thriving best in environments associated with relatively high organic matter flux (Bernhard 1986) supports this interpretation.
5. Upper Campanian: The well-preserved foraminiferal assemblage and increased numbers of Sitella, Vaginulina, Cibicides, Gyroidinoides, and Brotzenella indicate good stratification of the water column and a tropical to sub-tropical climate during the studied upper Campanian (Leckie 1989; Huber et al. 1995). These point out to stable environmental conditions, a stable thermocline and other parameters indicating the presence of an open marine outer shelf to slightly upper bathyal environments at least at the upper boundaries of the Duwi Formation at Gebel Duwi Basin.
The presence of Dorothia and Spiroplectammina in upper Campanian indicates marine environments from the inner shelf to upper bathyal; these faunas appear to be typical of moderately low levels of oxygen (Rückheim et al. 2006). The absence or a decrease in number of Spiroplectammina, nodosariids, Pseudovigerina, and Reussella in late Campanian is interpreted to indicate some cooling events associated with deep upwelling water circulation (Bejaoui et al. 2019).
Furthermore, in late Campanian, the genus Gyroidinoides commonly occurred in sediments of the oligotrophic well-oxygenated environment (Dubicka and Peryt 2011) as of late Campanian marker of the Duwi section (Brotzenella taylorensis zone), not as dysoxic bottom water conditions (Friedrich et al. 2003; Friedrich 2009).
6. The Bolivinoides and Orthokarstenia genera: Genus Bolivinoides are completely absent from uppermost Turonian, Coniacian to upper Campanian in the Duwi section, which coincides with Eastern and Western European provinces. The Bolivinoides lineages are rich and diverse in deeper zones with enough supply of nutrients and oxygen to sustain mesotrophic conditions in deeper environments (Alve and Goldstein 2003, 2010; Bejaoui et al. 2019). Orthokarstenia clarki is found in marly facies of the Quseir region with both Gavelinella and Stensioeina. The flooding occurrence of Orthokarstenia clarki through the Campanian interval of the Duwi Basin indicates middle to outer shelf to slightly deeper than 100 m. A similar case was interpreted by Kaminski and Gradstein (2005) showing Orthokarstenia acme zones in the modern North Atlantic water. The absence of Bolivinoides, Bolivina, and even Bolivinopsis genera were documented by several authors in the Maastrichtian sediments of southern Egyptian province (Hewaidy and Cherif 1984; Hewaidy 1996). Meanwhile, the Egyptian deeper northern paleogeographic province is characterized by flooding of both Bolivinoides and Orthokarstenia (El-Deeb and El-Gammal 1994, 1997).
7. Gavelinella, Stensioeina, and Brotzenella genera: The Gavelinella, Stensioeina, and Brotzenella genera of the Late Cretaceous co-occurred within the Duwi epicontinental basin. The Gavelinella and Stensioeina of the Coniacian Stage of the Duwi section together constitute up to 60% of the total benthic foraminiferal content and slightly decrease into the Santonian Stage most probably due to the Oceanic Anoxic Event (OEA3) (Fig. 6). This event is inferred and documented in the first appearance of Dicarinella asymetrica of early Santonian, which co-occurred with Neoflabellina suturalis and Cibicidoides eriksdalensis and extinctions to Marginotruncana and Dicarinella at the latest Santonian and Early Campanian (El Gammal and Orabi 2019a). The Late Campanian Event (LCE) (Jarvis et al. 2002; Thibault et al. 2012; Voigt et al. 2012) in many Tethyan cases may correlate with the planktonic Globotruncana aegyptiaca zone, which co-occurred with Brotzenella monterelensis and Brotzenella taylorensis (Fig. 6).
Concluding remarks
1. Identifications of the benthic foraminiferal assemblages (Gavelinella, Stensioeina, and Brotzenella) from the upper Turonian-upper Campanian successions of Gebel Duwi Range, Quseir Region, allowed the recognition of seven biozones and two subzones. The benthic foraminiferal contents are of diversified epibenthic and endobenthic taxa suggesting epicontinental inner to outer shelf depositional paleoenvironments and mesotrophic habitat.
2. Uppermost Turonian and lower Coniacian characterized by low-diversity assemblages. The behavior of recent benthic foraminifera under extreme conditions (food, oxygen) with low-diversity assemblages tend to occur, characterized by strong dominance of a few species.
3. The presence of Gavelinella, Cibicidoides, Gaudryina, Anomalina, Cibicides, Valvulineria, Chilostomella, and Lenticulina in the Santonian sediments, which belong to morphogroup plano-convex trochospiral, rounded trochospiral, tapered and cylindrical and lenticular (PT, RT, T/C, and L), is typical if aerobic and eutrophic to mesotrophic environments. Most of these taxa have been identified in the middle-outer shelf to upper slope deposits.
4. The benthic foraminiferal assemblages Orthokarstenia, Stensioeina, and Lenticulina are rich and well-preserved in the Early Campanian indicating further deepening of the Duwi Basin.
5. The flood of Orthokarstenia clarki in the overall number of middle Campanian species indicates mesotrophic to more oligotrophic environmental conditions and an open marine, probably outer shelf to slightly upper bathyal environment up to the late Campanian of benthic Brotzenella taylorensis zone.
6. The well-preserved foraminiferal assemblage and increased numbers of Sitella colonensis, Vaginulina, Cibicides, Gyroidinoides, and Brotzenella, which indicate good stratification of the water column and a tropical to sub-tropical climate during the studied upper Campanian.
7. The related morphotypes characterized by an abundance of the epifaunal forms and presence of some infaunal morphotypes of the well-oxygenated environment indicate a mesotrophic habitat. The Duwi benthic fauna are characterized by a mixture of forms occupying different habitats and employing different feeding strategies since both organic matter and oxygen levels are high enough to support epifaunal to deep infaunal microhabitats.
8. The Duwi Basin through the uppermost Turonian to upper Campanian increased in depth, the benthic components indicating a time of the Egyptian stable continental shelf. The gradual increase in depth had occurred with no dramatic structure or tectonic changes at least in southern Egypt.
References
Akkad MK, Noweir AM (1980) Geology and lithostratigraphy of the Arabian Desert orogenic belt between latitudes 25° 35῍ and 26° 30῍. Permagon Press, New York, pp 127–135
Alperin MI, Cusminsky GC, Bernasconi E (2011) Benthic foraminiferal morphogroups on the argentine continental shelf. J For Res 41(2):155–166
Alve E, Goldstein ST (2003) Propagule transport as a key method of dispersal in benthic foraminifera (Protista). Limnol Oceanogr 48:2163–2170
Alve E, Goldstein ST (2010) Dispersal, survival and delayed growth of benthic foraminiferal propagules. J Sea Res 63:36e51
Bailey HW, Gale AS, Mortimore RN, Swiecicki A, Wood CJ (1983) The Coniacian-Maastrichtian stages of the United Kingdom, with particular reference to southern England. Newsl Stratigr 12:19–42
Bailey HW, Hart MB, Swiecicki A (2009) Evolutionary lineages of benthic foraminifera in the chalk seas of NW Europe and their application to problem solving. In: Demchuk TD, Gary AC (eds) Geological problem solving with microfossils: a volume in honor of Garry D. Jones, vol 93. SEPM Special Publication, pp 233–249
Barmawidjaja, D.M., Jorisson, F. J. Puskaric, S., Van der Zawan, G., J., 1991. Microhabitat selection by benthic foraminifera in the northern Adriatic Sea. In: D. M. Barmawidjaja (ed), Studies in living and fossil foraminifers from seasonally productive Regions. Geol. Ultraiectina, 82: 15–55
Bejaoui A, Saidi E, Zaghbib-Turki D (2019) Small benthic foraminiferal biostratigraphy and palaeoecology during the Campanian-Maastrichtian transition in North-Western Tunisia. Turkish J Earth Sci 28:500–530
Beniamovski VN, Alekseev AS, Ovechkinac MN, Vishnevskayaa VS, Podgaetskiie AV, Pronin VG, V. G. (2012) Upper Campanian–Lower Maastrichtian sections of the northwestern Rostov region. Article 1. Description, paleontological assemblages, and lithobiostratigraphy. Stratigraphy and Geological Correlation, 2012 20(4):346–379
Benyamovskiy VN (2007) Benthic foraminifera-based zonal scheme of the upper cretaceous of the European Paleogeographic Area (EPA) as an evidence of stages of evolutionary development. In: Cretaceous system of Russia and adjacent areas: problems of stratigraphy and paleogeography, vol 2006. SO EAGO, Saratov, pp 26–27
Benyamovskiy VN (2008a) Infrazonal biostratigraphy of the Upper Cretaceous in the East European province based on benthic foraminifers, part 1: Cenomanian-Coniacian. Stratigr Geol Correl 16(3):257–266
Benyamovskiy VN (2008b) Infrazonal biostratigraphy of the Upper Cretaceous in the East European province based on benthic foraminifers, part 2: Santonian-Maastrichtian. Stratigr Geol Correl 16(5):515–527
Bernhard JM (1986) Characteristic assemblages and morphologies for anoxic organic rich deposits: Jurassic through Holocene. J Foraminifer Res 16:207–215
Bragina LG, Beniamovsk VN, Kopaevich LF (2016) Radiolarians, foraminifers, and biostratigraphy of the Coniacian–Campanian deposits of the Alan-Kyr Section, Crimean Mountains. Stratigraphy and Geological Correlation, 2016 24(1):39–57
Cause E, Bernaus JM, Boix C, Calonge A, Perez R (2003) Upper cretaceous shallow benthic biozones (KSBZ): a preliminary report. In: AAPG international conference Barcelona, Spain, pp 21–24
Coccioni R, Galeotti S (1993) Orbitally induced cycles in benthonic foraminiferal morphogroups and trophic structure distribution patterns from the L Albian Amadeus segment (Central Italy). J Micropaleontol 12:227–239
Corliss BH (1985) Microhabitats of benthic foraminifera within deep-sea sediments. Nature 314:435–438
Dubicka, Z., 2016. Benthic foraminiferal biozonation. In: Walaszczyk, I., Dubicka, Z., Olszewska-Nejbert, D., Remin, Z. (Eds.), Integrated biostratigraphy of the Santonian through Maastrichtian (upper cretaceous) of the extra-Carpathian, Poland. Acta Geologica Polonica, 66(3), pp. 313–350
Dubicka Z, Peryt D (2011) Integrated biostratigraphy of Upper Maastrichtian chalk at Chełm (SE Poland). Ann Soc Geol Pol 81:185–197
Dubicka Z, Peryt D (2012) Latest Campanian and Maastrichtian palaeoenvironment changes: implications from an epicontinental sea (SE Poland and western Ukraine). Cretaceous Res 37:272–284
Dubicka Z, Peryt D (2014) Classification and evolutionary interpretation of late Turonian–early Campanian Gavelinella and Stensioeina (Gavelinellidae, benthic foraminifera) from western Ukraine. J Foraminifer Res 44:151–176
Dubicka Z, Jurkowska A, Thibault N, Razmjooei MJ, Wojcik K et al (2017) An integrated stratigraphic study across the Santonian/Campanian boundary at Bocieniec, southern Poland: a new boundary stratotype candidate. Cretac Res 80:61–85
Edwards PG (1981) The foraminiferid genus Gavelinella in the Senonian of Northwest Europe. Palaeontology 24:391–416
El Gammal RMH, Orabi HO (2019a) Coniacian-late Campanian planktonic events in the Duwi formation, Red Sea region, Egypt. J Geol Geophys 8:1,16
El Gammal RMH, Orabi HO (2019b) New findings of the Cenomanian-Turonian successions in the Quseir region, Red Sea, Egypt. J Geol Geophys 8:3.13
Elamri Z, Sherif F, Zaghbib-Turki D (2018) Benthic foraminiferal paleoenvironmental reconstruction from the Upper Coniacian-Lower Campanian succession in north-western Tunisia. Rev Micropaleontol 61:23–36
El-Deeb WZM, El-Gammal RMH (1994) Contributions to the biostratigraphy of Sudr and Esna formations in southwestern Sinai, Egypt. Egypt J Petrol 3:65–84
El-Deeb WZM, El-Gammal RMH (1997) Foraminiferal paleoecologic study of the Maastrichtian -Ypresian succession at Southwestern Sinai Area, Egypt. M.E.R.C. Ain Shams Univ., Earth Sci. Ser. 11:37–50
Fontanier C, Jorissen FJ, Chaillou G, Anschutz P, Grémare A, Griveaud C (2005) Live foraminiferal faunas from a 2800 m deep lower canyon station from the Bay of Biscay: faunal response to focusing of refractory organic matter. Deep-Sea Research I 52:1189–1227
Frenzel P (2000) Die benthischen Foraminiferen der Rügener Schreibkreide (Unter-Maastricht, NE-Deutschland). Neue Paläontologischen Abhandlungen 3:1–361
Friedrich O (2009) Benthic foraminifera and their role to decipher paleoenvironment during mid-Cretaceous oceanic anoxic events–the “anoxic benthic foraminifera” paradox. Rev Micropaleontol 53:175–192
Friedrich O, Reichelt K, Herrle JO, Lehmann J, Pross J, Hemleben C (2003) Causes for the formation of the Late Aptian Niveau Fallot black shales in the Vocontian Basin (SE France): evidence from planktic and benthic foraminifera and stable isotopes. Mar Micropaleontol 49:65–85
Glenn CR, Arthur MA (1990) Anatomy and origin of a Cretaceous phosphorite-greensand giant, Egypt. Sedimentology 37:123–154
Gooday AJ, Rathburn AE (1999) Temporal variability in living deep-sea benthic foraminifera: a review. Earth Sci Rev 46(1–4):187–212
Grunert P (2011) Integrated facies analysis and stratigraphy of the Early Miocene north Alpine foreland basin (Upper Austria). Unpublished Ph.D. Thesis, Karl Franzens Universität Graz
Gusic I, Jelaska V, Velic I (1988) Foraminiferal assemblages, facies, and environments in the upper cretaceous of the island of Brac, Yugoslavia. Revue de Palébiologie 2(Benthos’86):447–456
Haq BU, Hardenbol J, Vail PR (1987) Chronology of fluctuating sea levels since the Triassic. Science 235:1156–1167
Hart MB, Bailey HW, Crittenden S, Fletcher BN, Price RJ, Swiecicki A (1989) Cretaceous. In: Jenkins DG, Murray JW (eds) Stratigraphical atlas of fossil foraminifera, 2nd edn. Ellis Horwood Ltd, Chichester, pp 273–371
Haynes JR (1981) Foraminifera. Macimilian Publishers Ltd, London, 433p
Heinz P, Kitazato H, Schmiedl G, Hemleben C (2001) Response of deep-sea benthic foraminifera from the Mediterranean Sea to simulated phytoplankton pulses under laboratory conditions. J Foraminifer Res 31:210–227
Hewaidy AA (1996) Factors controlling the distribution of the foraminiferal genera Othokarstenia Dietrich and Bolivinoides Cushman in Egypt. Geol Soc Egypt, Spec Publ No 2(1996):1–12
Hewaidy AA, Cherif OH (1984) Contribution to the bathymetric variations of the Late Cretaceous sea over the Abu Tartur area by using foraminifera. Annals of the Geological Survey of Egypt 15:231–241
Huber BT, Hodell DA, Hamilton CR (1995) Middle-Late Cretaceous of the southern high latitudes: stable isotopic evidence for minimal equator-to-pole thermal gradients. Geol Soc Am Bull 107:1164–1191
Issawi B, El-Hinnawi M, Francis M, Mehanna A (1969) Contribution to the structure and phosphate deposits of Qusseir area: Egypt. Organ Geol Res Min Geol Survey 50:35
Jarvis I, Mabrouk A, Moody RTJ, de Cabrera S (2002) Late Cretaceous (Campanian) carbon isotope events, sea-level change and correlation of the Tethyan and Boreal realms. Palaeogeogr Palaeoclimatol Palaeoecol 188(3–4):215–248
Jones RW, Charnock MA (1985) “Morphogroups” of agglutinating foraminifera. Their life position, feeding habits and potential applicability in (paleo) ecological studies. Rev Paléobiol 4:311–320
Jorissen FJ, Stigter HC, Widmark JGW (1995) A conceptual model explaining benthic foraminiferal microhabitats. Mar Micropaleontol 26:3–15
Kaiho K (1991) Global changes of Paleogene aerobic/anaerobic benthic foraminifera and deep-sea circulation. Palaeogeogr Palaeoclimatol Palaeoecol 83:65–85
Kaminski MA, Gradstein FM (2005) Atlas of Paleogene cosmopolitan deep-water agglutinated foraminifera. Grzybowski Foundation special publication 10:574 +vii pp
Kaminski MA, Boersma A, Tyszka J, Holbourn AEL (1995) Response of deep-water agglutinated foraminifera to dysoxic conditions in the California borderland basins. In: Kaminski MA et al (eds) Proceedings of the Fourth International Workshop on Agglutinated Foraminifera, Kraków, Poland. September 12-19, 1993. Grzybowski Foundation spec. Public, vol 3, pp 131–140
Kerdany, M. T., Cherif, O., 1990. Mesozoic. Chapter 22, in Geology of Egypt, (1990), Said, R. (Edi.). 1990 A. Balkema, Rotterdam, Netherlands, p.727
Khalil SM, McClay K (2002) Extentional fault related folding, northwestern Red Sea, Egypt. J Struc Geol 24:743–762
Khare N, Sinha R, Rai AK, Nigam R (1995) Distributional pattern of benthic foraminiferal morpho-groups in the shelf region of Mangalore: environmental implications. Indian J Mar Sci 24:162–165
Koch W (1977) Stratigraphie der Oberkreide in Nordwestdeutschland (Pompeckjsche Scholle). Teil 2. Biostratigraphie in der Oberkreide und Taxonomie von Foraminiferen. Geologisches Jahrbuch A 38:11–123
Kopaevich LF, Beniamovski VN, Sadekov AY (2007) Middle Coniacian-Santonian foraminiferal bioevents around the Mangyshlak Peninsula and Russian platform. Cretaceous Res 28:108–118
Koutsoukos EAM, Hart MB (1990) Cretaceous foraminiferal morphogroup distribution patterns, palaeocommunities and trophic structures: a case study from the Sergipe Basin, Brazil. Trans R Soc Edinburgh Earth Sci 81(3):221–246
Lamolda MA, Paul CRC, Peryt D, Pons JM (2014) The global boundary stratotype and section point (GSSP) for the base of the Santonian stage, “Cantera de Margas”, Olazagutia, northern Spain. Episodes 37:2–13
Leckie RM (1989) A paleocenographic model for the early evolutionary history of planktonic foraminifera. Palaeogeogr, Palaeoclimatolol, Palaeoecol 73:107–138
Lüning S, Marzouk AM, Morsi AM, Kuss J (1998) Sequence stratigraphy of the Upper Cretaceous of south-east Sinai. Egypt: Cretaceous Res 19:153–196
Murphy MA, Salvador A (1994) International Subcommission Stratigraphic Classification of IUGS, International Commission on Stratigraphy, International Stratigraphic Guide - an abridged version. Episodes, Vol. 22, no. 4
Murray JW (1991) Ecology and palaeoecology of benthic foraminifera. In: Longman Scientific and Technical. John Wiley and Sons Inc., Essex, New York, pp 1–329
Nagy J (1992) Environmental significance of foraminiferal morphogroups in Jurassic North Sea deltas. Palaeogeogr Palaeoclimatol Palaeoecol 95:111–134
Nagy J, Gradstein FM, Kaminski MA, Holbourn AE (1995) Foraminiferal morphogroups, palaeoenvironments and new taxa from Jurassic to cretaceous strata of Thakkhola, Nepal. In: Kaminski MA, Geroch S, Gasiński MA (eds) Proceedings of the Fourth International Workshop on Agglutinated Foraminifera, vol 3. Grzybowski Foundation special publication, pp 181–209
Nagy J, Kaminski MA, Johnsen K, Mitlehner AG (1997) Foraminiferal, palynomorph, and diatom biostratigraphy and paleoenvironments of the Tork Formation: a reference section for the Paleocene–Eocene transition in the western Barents Sea. In: Hass HC, Kaminski MA (eds) Contributions to the micropaleontology and paleoceanography of the northern North Atlantic, vol 5. Grzybowski Foundation Special Publication, pp 15–38
Nagy J, Kaminski MA, Kuhnt W, Bremer MA (2000) Agglutinated foraminifera from neritic to bathyal facies in the Palaeogene of Spitsbergen and the Barents Sea. In: Hart MB, Kaminski MA (eds) Proceedings of the Fifth International Workshop on Agglutinated Foraminifera, vol 7. Grzybowski Foundation special publication, pp 333–361
Naidin DP, Beniamovskiy VN (2006) The Campanian–Maastrichtian stage boundary in the Aktulagai section (North Caspian depression). Stratigr Geol Correl 14(4):433–443
North American Commission on Stratigraphic Nomenclature (NACSN) (1983) North American stratigraphic code. Am Assoc Pet Geol Bull 67(5):841–875
Odin GS, Lamaurelle AM (2001) The global Campanian-Maastrichtian stage boundary. Episodes 24:229–238
Olfer'ev AG, Alekseev AS (2003) Zonal stratigraphical scale of the Upper Cretaceous of the east-European platform: stratigraphy and geological correlation. 11:172–198
Olferev AG, Beniamovski VN, Vishnevskaya VS, Ivanov AV, Kopaevich LF, Pervushov EM, Seltser VB, Tesakova EM, Kharitonov VM, Shcherbinina EA (2007) Upper Cretaceous deposits in the north west of Saratov oblast, part 1: Litho- and biostratigraphic analysis of the Vishnevoe section. Stratigr Geol Correl 15:610–655
Olferiev AG, Alekseev AS (2005) Stratigraphic scheme of the Upper Cretaceous sediments of the east European platform. Explanatory Note, Moscow: Paleontol. Inst. RAN
Olóriz F, Reolid M, Rodríguez-Tovar FJ (2006) Approaching trophic structure in Late Jurassic neritic shelves: a western Tethys example from southern Iberia. Earth−Sci Rev 79:101–139
Olsson RK (1988) Foraminiferal modeling of sea-level change in the Late Cretaceous of New Jersey. In: Wilgus CK, Hastings BS, Kendall CGSC, Posamentier HW, Ross CA, Van Wagoner JC (eds) Sea-level changes: an integrated approach, 289-297. The Society of Economic Paleontologists and Mineralogists. Special publication, Tulsa, OK, p 42
Peryt D, Dubicka Z (2015) Foraminiferal bioevents in the upper Campanian to lowest Maastrichtian of the Middle Vistula River section, Poland. Geological Quarterly 59:814–830
Polivoda AS, Abramovich S, Almogi-LabinL A, Schineider-Mor A, Feinsttein S, Puttmann W, Berner Z (2011) Palaeoenvironments of the latest Cretaceous oil shale sequence, Sothern Tethys, Israel, as an integral part of the prevailing upwelling system. Palaeogeogr, Palaeoclimatolol, Palaeoecol 305:93–108
Reolid M, Rodriguez-Tovar F, Nagy J, Oloriz F (2008) Benthic foraminiferal morphogroups of mid to outer shelf environments of the Late Jurassic (Prebtic zone, southern Spain): characterization of biofacies and environmental significance. Palaeogeogr Palaeoclimatol Palaeoecol. 261:280–299
Rückheim S, Bornemann A, Mutterlose J (2006) Planktic foraminifera from the mid-Cretaceous (Barremian-early Albian) of the North Sea Basin: Palaeoecological and palaeoceanographic implications. Mar Micropaleontol 58:83–102
Said R (1962) The geology of Egypt. Elsevier, Amsterdam
Said R., 1990. The geology of Egypt. Balkema, Rotterdam. pp721
Sen Gupta BK, Machain-Castillo ML (1993) Benthic foraminifera in oxygen-poor habitats. Mar Micropaleontol 20:3–4
Setoyama E, Kaminski MA, Tyszka J (2011) The Late Cretaceous–Early Paleocene palaeobathymetric trends in the southwestern Barents Sea-palaeoenvironmental implications of benthic foraminiferal assemblage analysis. Palaeogeogr Palaeoclimatol Palaeoecol 307:44–58
Setoyama E, Kaminiski MA, Tyszka J (2017, 2017) Late Cretaceous–Paleogene foraminiferal morphogroups as palaeoenvironmental tracers of the rifted Labrador margin, northern proto-Atlantic. In: Kaminski MA, Alegret L (eds) Proceedings of the Ninth International Workshop on Agglutinated Foraminifera, vol 22. Grzybowski Foundation special publication, pp 179–220
Sjoerdsma PG, Van der Zawan GJ (1992) Simulating the effect of changing organic flux and oxygen content on the distribution of benthic foraminifers. Mar Micropaleont 19:163–180
Stojanova V, Petrov G (2018) Paleoecological significance of benthic foraminiferal fauna from the Ovče Pole Basin, Republic of Macedonia. Geologica Macedonica 32(1):45–57
Szydło A (2005) Benthic foraminiferal morphogroups and tapho-nomy of the Cieszyn beds (Tithonian-Neocomian, Polish Outer Carpathians). Studia Geol Polonica 124:199–214
Thibault N, Husson D, Harlou R, Gardin S, Galbrun B, Huret E, Minoletti F (2012) Astronomical calibration of upper Campanian–Maastrichtian carbon isotope events and calcareous plankton biostratigraphy in the Indian Ocean (ODP Hole 762C): Implication for the age of the Campanian–Maastrichtian boundary. Palaeogeogr Palaeoclimatol Palaeoecol 337–338:52–71
Tyszka J (1994) Response of Middle Jurassic benthic foraminiferal morphogroups to dysoxic/anoxic conditions in the Pieniny Klippen Basin, Polish Carpathians. Palaeogeogr, Palaeoecol, Palaeoclimatol 110:55–81
Tyszka J (2006) Taxonomy of Albian Gavelinellidae (Foraminifera) from the Lower Saxony Basin, Germany. Palaeontology 49(6):1303–1334
Tyszka J (2009) Foraminiferal response to seasonality modulated by orbital cycles in the Cretaceous mid-latitudes: the Albian record from the Lower Saxony Basin. Palaeogeogr Palaeoclimatol Palaeoecol 276:148–159
Van der Zwaan GJ, Jorissen FJ (1991) Biofacial patterns in river-induced shelf anoxia. In: Tyson RV, Pearson TH (eds) Modern and ancient continental shelf anoxia, vol 58. Geol. Soc. London, Spec Publ, pp 65–82
Velic I, Vlahovic I (1994) Foraminiferal assemblages in the Cenomanian of the Buzet–Savudrija area (northwestern Istria, Croatia). Geol Croatica 47(1):25–43
Vishnevskaya VS, Kopaevich LF, Beniamovskiia VN, Ovechkinac MN, M.N. (2018) The correlation of the Upper Cretaceous zonal schemes of the eastern European platform based on Foraminifera, Radiolaria, and Nannoplankton. Mosc Univ Geol Bull 73(2):131–140
Voigt S, Gale AS, Jung C, Jenkyns HC (2012) Global correlation of Upper Campanian - Maastrichtian successions using carbon-isotope stratigraphy: development of a new Maastrichtian timescale. Newsl Stratigr 45(1):25–53
Walaszczyk I, Kopaevich LF, Olfer'ev AG (2004) Inoceramid/foraminiferal succession of the Turonian and Coniacian (Upper Cretaceous) of the Briansk Region (Central European Russia). Acta Geol Polon. 54(4):597–609
Walaszczyk I, Dubicka Z, Olszewska-Nejbert D, Remin Z (2016) Integrated biostratigraphy of the Santonian through Maastrichtian (Upper Cretaceous) of extra-Carpathian Poland. Acta Geol Pol 66(3):313–350
Walker SE, Miller W (1992) Organism—substrate relations. Toward a logical terminology: PALAIOS 7:236–238
Widmark JGV (1997) Deep-sea benthic foraminifera from Cretaceous-Paleogene boundary strata in the South Atlantic -taxonomy and paleoecology. Fossils Strata 43:1–94
Acknowledgments
First and foremost, the authors want to sincerely thank Prof. Dr. Jaroslaw Tyszka, Institute of Geological Sciences, Polish Academy of Sciences, Poland, for his helpful discussion and revision of the manuscript. The authors are grateful to the anonymous reviewer for his very helpful comments on the manuscript. We greatly appreciate the organizations that funded our research.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
The authors declare that they have no competing interests.
Additional information
Responsible Editor: Attila Ciner
Rights and permissions
About this article

Cite this article
Orabi, O.H., El Gammal, R.M. Biostratigraphic and paleoecological significance of benthic foraminiferal morphogroups in the uppermost Turonian–upper Campanian of Gebel Duwi Range, Red Sea, Egypt. Arab J Geosci 14, 168 (2021). https://doi.org/10.1007/s12517-020-06353-z
Received:
Accepted:
Published:
DOI: https://doi.org/10.1007/s12517-020-06353-z