
Abstract
In the northwestern part of the Arabian Platform, the Adıyaman district (SE Turkey) displays some Lower-Middle Cenomanian deep-marine sediments of the Derdere Formation. These sediments are yielding abundant echinodermal remains, among which some thecal and brachial plates are assignable to roveacrinids. Routine microfacies analysis of the Lower-Middle Cenomanian part of the Derdere Formation revealed unexpected early Cenomanian assemblages of roveacrinoidal ossicles, comparable with those formerly reported further south in the Cenomanian-Turonian of the Arabian Platform. For the first time, genuine and undisputable Roveacrinidae are illustrated for SE Turkey. Seven borehole sections were scrutinized in search of microcrinoidal sections, most especially within carbonate microfacies. Within the scope of better constraining the position of the Lower-Middle Cenomanian, we had been compiling the successive occurrence of respective identified roveacrinid remains. These roveacrinidal assemblages are consisting in: Roveacrinus communis Douglas (1908) (= R. derdereensis Farinacci & Manni); Roveacrinus cf. alatus Douglas (1908); Roveacrinus spinosus Peck (1943); Roveacrinus sp.; Roveacrinidae indet.; Applinocrinus sp.; and Saccocomidae indet. These specimens provide significant clues to constrain the palaeogeographic reconstruction of Tethyan seaways, and represent potential fossil index candidates for the Lower Cenomanian stratigraphy of the Adıyaman district.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Cretaceous deposits are widely distributed over the Arabian Platform along the southern margin of the former Tethys Ocean. On the SE Turkish territory, Cretaceous exposures are well-known and have been mapped repeatedly over the past 50 years, particularly those of the Adıyaman district and surrounding areas. Only a few recent studies addressed the Lower-Middle Cretaceous lithostratigraphy, petrography, and sedimentological evolution of SE Turkey (Celikdemir et al. 1991; Mulayim et al. 2016).
In order to refine the stratigraphical framework of the Lower-Middle Cenomanian in this area, the microfacies analysis of the Derdere limestones evidenced the fluctuating but steady occurrence of echinodermal ossicles. Such crinoidal (“saccocomid-like”) microfacies were reported in internal unpublished reports by TPAO. Recent crinoidal microfacies were first published in SE Turkey in the Cenomanian Derdere Formation (Farinacci and Manni 2003).
Here, we focus our attention on the crinoid assemblages of this carbonate succession, with special attention to the most widespread but also most underestimated crinoid group, the Roveacrinida. We intend to demonstrate their potential for stratigraphic purposes, as examplified by their widespread occurrence in the Cretaceous succession of the Derdere Formation, and compare them to coeval faunas of adjacent areas.
Geologic setting and stratigraphic frame
SE Turkey is at the leading edge of a continent/continent collision where the Arabian Plate is converging on Eurasia at a pace of 18 ± 2 mm/year in a roughly north-northwesternward direction (McClusky et al. 2000). This collision is manifest in the convergent plate boundaries that are currently proximal to SE Turkey (Fig. 1). Events on these boundaries have largely controlled the Paleozoic, and particularly Late Cretaceous-Late Miocene, tectonics of SE Turkey.

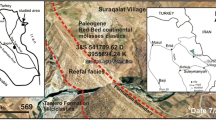
Geographical map of SE Turkey and location of studied borehole sections
The mid-Cretaceous Mardin Group carbonate platform of SE Turkey developed on the passive, northern edge of the Arabian Plate prior to the southward thrusting and subsequent folding during Late Cretaceous (Cater and Gillcrist 1994) (Fig. 1). The Lower Mesozoic basement is made of carbonates assigned to the Aptian-Campanian Mardin Group, some 700 m thick (Celikdemir et al. 1991). These carbonates were deposited on the southern passive margin of the Neo-Tethys and are subdivided into four formations (Tuna 1973). At its base is the Aptian-Albian Areban Formation, composed of limestones, dolomites, and interbedded shales and sandstones, all deposited in a restricted lagoonal to tidal-flat setting. This grades conformably up into the tidal-flat to subtidal carbonates of the Sabunsuyu Formation (Fig. 2). Conformably the organic-rich limestones at the base of the overlying Derdere Formation were deposited under relatively deeper marine, anoxic conditions, and are passing upward into a shallowing-upward sequence consisting of lagoonal to tidal-flat carbonates. Fossil-bearing and organic-rich carbonates of the unconformably overlying Karababa Formation were deposited under shallow marine and lagoonal conditions (Wagner and Pehlivanlı 1985; Celikdemir and Dulger 1990; Duran 1991; Perincek et al. 1991; Celikdemir et al. 1991; Mulayim et al. 2016).

Lithological and stratigraphical column of the Derdere Formation
According to Handfield et al. (1959) and Bryant (1960), the Derdere Formation in its type locality is transitional and lies conformably on the Sabunsuyu Formation and unconformably below the Karababa Formation (Kellogg 1960, 1961) (Fig. 2). From its type locality of the Korudag section, the Derdere Formation (73 m thick: Handfield et al. 1959) is composed of pale gray to brown, hard, medium-thick bedded, limestone, dolomite, and dolomitic limestones. Similarly, the lithostratigraphical succession of the Sabunsuyu section is similar to that in the type section. However, the formation here is thinner than in the Sabunsuyu area with only 209 m (Cros et al. 1999). The Derdere Formation has drawn most of the attention from the foraminiferal workers (Koyluoglu 1981; Coruh 1981, 1983; Erenler 1989) and the calcareous nannofossil researchers of SE Turkey (Ertug 1991). Preliminary biostratigraphic data (ongoing study) mostly based on benthonic foraminifers supported a Cenomanian age for the study area.
Lithostratigraphy and microfacies of the Lower-Middle Derdere Formation
Lower limestone unit
This unit consists of a thin-medium bedded, dark brown-beige, organic matter-rich, about 10–40-m thick limestone. This lower part of the Derdere Formation is particularly rich in calcispheres [e.g., Stomiosphaera sphaerica (Kaufmann), Calcisphaerula innominata Bonet (Fig. 3a), Pithonella ovalis (Kaufmann) (Fig. 3b), Pithonella sphaerica (Kaufmann); for original descriptions, see Kaufmann (1865)], planktonic foraminifers [Heterohelicidae, Hedbergellidae], roveacrinoids [Applinocrinus sp.], and ostracode fragments. The microfacies documents a wackestone-packstone texture with pelagic crinoids (Roveacrinidae indet.), calcispheres, and planktonic foraminifers (Fig. 2). The lowermost part of the succession consists of decimetric, bioclastic carbonate beds that mostly contain echinoderm ossicles (brachial and thecal plates of roveacrinoids, lateral plates of ophiuroids, and echinoid spines).

Microfacies photographs of the lower unit of the Derdere Formation. a Pithonella ovalis (Kaufmann); b Calcisphaerula innominata (Kaufmann); c, d MF1—calcisphere-planktonic foraminifer packstone, samples Bk-1.4.1 and K-1.1700, respectively; e, f MF2—calcisphere-roveacrinoid wackestone-packstone, samples S-2_2202_5x and S-2-2202.3, respectively; g, h MF3—bioclastic roveacrinoidal wackestone, calcareous benthonic foraminifers (Lenticulina sp.) with bioclasts in sample S-2-2274.2, and a transverse section of an echinoid spine in sample S-2-2274, respectively
Based on petrographic analysis and sedimentologic features of the lower limestone of the Derdere Formation, three microfacies are interpreted to represent mid- to outer-ramp environments.
Mf-1—calcisphere-planktonic foraminifer packstone
Calcispheres are the dominant skeletal grains in this microfacies, ranging from 60 to 80% in abundance. First, Pithonella ovalis and Calcisphaerula innominata are the major components. Secondly, the planktonic foraminifers are generally belonging to Families Heterohelicidae and Hedbergellidae. Calcispheres generally range in size from 0.05 to 0.15 mm while planktonic foraminifers range from 0.10 to 0.20 mm in size (Fig. 3c, d). Skeletal grains are randomly distributed in packstones. Dolomite occurs as a minor component (1 to 2%) in some samples. Some skeletal grains, especially planktonic foraminifers, show some dolomitic replacement. Both partial and complete replacements are observed. Matrix mainly consists in dark-brown micrite.
Mf-2—calcisphere-roveacrinid wackestone-packstone
The main constituents of this microfacies are calcispheres, roveacrinids, as well as planktonic foraminifers. Roveacrinids range from 5 to 30% in abundance. Planktonic foraminifers are 0.10 to 0.15 mm in size, and poorly preserved (Fig. 3e, f). Chambers of some planktonic foraminifers are completely filled in with dolomite at places. Dolomite occurs as rhombic crystals or patches which replace the chambers partially or completely. Chambers are generally neomorphosed and are filled in with sparite. Skeletal grains are evenly distributed in a micritic matrix of wackestone textures. Other subordinate grains are pelagic crinoid bioclasts. The matrix is generally composed of earthy to dark-brown micrite. The abundance of calcispheres along with roveacrinids, fine bioclasts, and mud-supported fabric all indicate a low-energy and deep-water environment.
Mf-3—bioclastic roveacrinoidal wackestone
This microfacies contains various bioclasts including roveacrinids, bivalves, as well as planktonic and some benthonic foraminifers and fine peloids (Fig. 3g, h). It displays no trace of organic matter. The presence of a micrite matrix evidences a low-energy environment. Disarticulation of bioclasts is due to bioturbation or storm activity. The combination of abundant and fragmented marine fauna, a micritic matrix, and texture suggests that the bioclastic wackestone facies was deposited in an open marine environment, probably below wave base.
Middle dolomite-dolomitic limestone unit
The middle unit, about 50–80-m thick, consists of dolomite and dolomitic limestone (Fig. 2), in medium-thick beds. It displays a bioclastic dolo-wackestone facies, characterized by bivalve fragments and benthonic foraminifers as the sole skeletal constituents within fine crystalline dolomite. Dolomite is characterized either by medium crystalline (20–100 μm), planar-e to planar-s type, or by fine crystalline mosaic especially when associated with bioclasts from a quiet environment. Dolomitization selectively affects matrix, or both matrix and grains simultaneously. Dolo-wackestone usually occurs in discrete beds within dolo-mudstone, although in a few places they grade into one another. The dolo-wackestone represents mud-supported fabrics from outer and middle ramp.
Identification of ossicles and recognition of section plane
The roveacrinoidal contribution to the Cretaceous limestones was reported in details by Ferré and Berthou (1993, 1994). The roveacrinoidal sections were only a few mentioned in the previous literature, most of the time gross identified as “Saccocoma limestones” [by comparison to seminal works on Jurassic rocks of Lombard (1937, 1945), and Verniory (1954, 1955, 1956, 1960, 1961, 1962)] or simply turned down by petroleum industry owing to the ignorance of their true nature and potentials. From this rather limited number of papers, it appears that the crinoidal component of carbonate rocks consists mostly of blooming populations of opportunistic roveacrinids; their mass occurrence and post-mortem accumulation are largely responsible for these very special microfacies scattering the Cretaceous carbonate series worldwide (e.g., Scott et al. 1977). The first formal evidence of roveacrinoid contribution to the Cretaceous carbonate factory in Turkey were brought by Cros et al. (1991) and Ferré et al. (1997). However, the first sections of genuine roveacrinidal affinity in SE Turkey were first illustrated by Farinacci and Manni (2003) with the erection of a new species: Roveacrinus derdereense FARINACCI & Manni, now and here regarded as a junior synonym of R. communis Douglas. Purported at first for roveacrinids (Ferré and Berthou 1993, 1994), the section orientation scheme was extended to saccocomids by Ferré and Dias-Brito (1999)—for extended typological comparison to Jurassic saccocomid sections, see Benzaggagh et al. (2018, for details and literature within). Meanwhile, Ferré and Granier (1997a, b, 2001) defined in length the orientation and taxonomic use of roveacrinid sections. Following that, the systematic assignment of Cretaceous roveacrinidal sections has been fully argumented for both the roveacrinids and the saccocomids (Ferré 1997; Ferré et al. 1999). We adopt this scheme in the following (Fig. 4).

Since these minute discarded or accumulated roveacrinid remains are not easily spotted in the field and require to be studied under a petrographic microscope and/or scanning electron microscopy (SEM), they are usually mentioned as microcrinoids or often misinterpreted as planktonic crinoids (instead of pelagic). The thecal size does not exceed a few millimeters; a complete specimen is about 5 cm wide. Roveacrinids are small articulate crinoids with five dichotomous arms, each displaying many brachial plates (up to three dozen in complete specimens). Their minute theca is devoid of any stem or anchoring device, and is built of two sets of plates, basal and dorsal, sometimes showing a prominent centrodorsal bulge. When exceptionally preserved, it displays an inner plate ring defining a double body cavity (Schneider 1987, 1989). Each roveacrinid species displays a distinctive architecture and bears a wide array of ornamental elements, such as a spine-like aboral element, simple bowls with or without processes, flanged or winged brachials, lateral processes, and flanges or spines (e.g., Schneider 1987, 1989; Jagt 1999; Hess 2015; Gale 2016).
Systematic paleontology
The supra-generic systematics follow the scheme proposed by Hess and Messing (2011), amended by Wright et al. (2017). All figured specimens and associated thin sections are housed in the collection of the Turkish Petroleum Corporation (TPAO)/Research Center (ARGEM), Ankara, under the abbreviation TPA.01 and catalogue numbers from TPA.01-BO01/09; TPA.01-BA01/03; TPA.01-BZ01; TPA.01-SV01/09; TPA.01-DK01/05; TPA.01-GN01/02; and TPA.01-KM01, respectively.
In the following, the term “cf.” refers to specimens, morphological features of which are comparable but not sufficient to clearly state a specific assignment.
Class Crinoidea Miller, 1821
Subclass Articulata VON Zittel, 1879
Order Roveacrinida Sieverts-Doreck, 1952
Family Roveacrinidae Peck, 1943
Genus Roveacrinus Douglas, 1908
Roveacrinus communis Douglas, 1908
(= Roveacrinus derdereensis MANNI, in FARINACCI & MANNI, 2003)
Plate 1, panels a, b

Microfacies (thin section) photographs of roveacrinid ossicles, Lower-Middle Cenomanian, Adıyaman area, SE Turkey. a Roveacrinus communis Douglas (= R. derdereensis Manni, in Farinacci & Manni), Obl/sub-TS-Rad and OblS-NBrn, core sample B-1_1484.1; b Roveacrinus communis Douglas, TS-Rad, Tg/LgS-Rad, and Lg/TgS-Rad, core sample S-2-2204.1; c Roveacrinus cf. alatus, OblS-Rad, core sample Bz-1-4-8; d Roveacrinus cf. alatus, (TS-Rad) and Roveacrinidae indet. (TS-NBrn), core sample S-2-2202.3; e accumulation of roveacrinoidal brachial plates (Roveacrinus cf. alatus), core sample D-1-2552.2; f Roveacrinus sp., TS-Rad and Obl/TS-IBr2, core sample Bz-1-4-7.1; g Roveacrinus sp., TS-Theca and OblS-I/IIBrn, core sample Bz-1-4-9.1; h Roveacrinus sp., OblS-Theca, core sample Bz-1-4-10.1
Material: Thin sections Rep. no. TPA.01-BA02 and TPA.01-SV08 (core samples B-1_1484.1 and S-2-2204.1, respectively).
Description: Plate 1, panel a depicts an oblique (transverse) section of a dorsal bowl (“complete theca” near its base, in the right part of the picture) along with some oblique sections of indeterminable brachial plates. This thecal section displays a sub-pentagonal rounded bowl flanked by five rather thick, lamellar radial expansions. The lack of reticulated pattern on these “alar” expansions excludes its identification as R. geinitzi Schneider, while their relatively thickness precludes their assignment to R. alatus Douglas.
Plate 1, panel b depicts various sections (transverse, transverse-longitudinal, and longitudinal-tangential) of isolated radial plates. The transverse section documents a rather angular outline of the radial edge characteristic of R. communis Douglas, devoid of any secondary ornamentation or wing-like radial expansion.
Given the outwards oblique orientation of radial articular facets, the highly conical, slender thecal silhouette, and the slightly ribbed longitudinal-radial ornamentation displayed by R. derdereensis Manni, this taxon should be considered as a junior synonym of R. communis DOUGLAS.
Occurrence: This taxon ranges from the Albian deposits of US Gulf Coast and Mexico (Peck 1943), through the Late Cretaceous chalks of Boreal Europe (Douglas 1908; Peck 1955; Rasmussen 1961), to the Paleogene of Poland (Salamon et al. 2010). In the Arabian Platform, it is recorded from Sabunsuyu Ravine in NE Turkey (Cros et al. 1991, 1999; Ferré et al. 1997), the Palmyrides in Syria, the coastal plains of Lebanon, and in the Cenomanian-Turonian deposits of Iran (Ferré, ongoing study).
Roveacrinus cf. alatus Douglas, 1908
Plate 1, panels c, e
Material: Thin sections Rep. no. TPA.01-BO09; TPA.01-SV01 and TPA.01-DK03 (core samples Bz-1-4-8; S-2-2202.3; and D-1-2552.2, respectively).
Description: Plate 1, panel c documents an oblique section of a lamellar radial expansion, typical of R. alatus. However, there is no radial structure to be affirmative for its specific assignment.
Plate 1, panel d shows a transverse section of an isolated, gracile radial plate and some transverse sections of indeterminable brachial plates displaying the typical wing-like radial expansion.
Plate 1, panel e illustrates an accumulation of roveacrinoidal brachial plates. Among these, some are showing the characteristic wing-like radial expansion.
This taxon with its special alar morphology is often found widespread in the Lower Cenomanian-Upper Turonian, in association with R. communis, over the central Tethys (Ferré et al. 1997, 2018) and the Boreal realm (Douglas 1908; Peck 1955; Rasmussen 1961).
Occurrence: This taxon with its special alar morphology is often found widespread in the Lower Cenomanian-Upper Turonian, in association with R. communis, over the central Tethys (Ferré et al. 1997, 2018) and the Boreal realm (Douglas 1908; Peck 1955; Rasmussen 1961).
Roveacrinus spinosus Peck, 1943
Plate 2, panels a, b

Microfacies (thin section) photographs of roveacrinid ossicles, Lower-Middle Cenomanian, Adıyaman area, SE Turkey. a Roveacrinus spinosus Peck, Lg/TgS-IBr1 and OblS-NBrn, core sample S-2-2202.4; b Roveacrinus spinosus Peck bearing typical spinose ornamentation (Lg/TgS-NBrn), and Roveacrinidae indet. (TgS-IBr1), core sample S-2-2202.5; c Applinocrinus sp. (Lg/TS-Rad) and Saccocomidae indet. (TgS-IBr2; OblS-NBrn), core sample Bz-1-4-7.3 (ex Bz-1-4.7); d Saccocomidae indet., indeterminable plates, core sample Bz-1-4-7; e Saccocomidae indet., Obl/TS-N/IIBrn/Rad, core sample B-1_1508.1; f Roveacrinidae indet., TgS-IBr2, core sample Bz-1-4-7.2; g Roveacrinidae indet., TgS-IBr1 and TgS-IBr2, core sample Bz-1-4-9.2 (ex Bz-1-4.9.10x); h Roveacrinidae indet., brachial plates, core sample Bz-1-4-10
Material: Thin sections Rep. no. TPA.01-SV09 and TPA.01-SV02 (core samples S-2-2202.4 and S-2-2202.5).
Description: Plate 2, panel a documents a “bifid” longitudinal-tangential section of a first primibrachial plate. This very special ornamentation is consistent with a spinose outward-directed ornamentation of proximal brachial plates below the articular facet, accompanied by oblique sections of indeterminable brachial plates also bearing “radial” ornament.
Plate 2, panel b documents a longitudinal-tangential section of a proximal brachial plate showing a spinose ornamentation. This feature is not consistent with a peculiar section orientation of the alar morphology of R. alatus but is in accordance with that characterizing R. spinosus from the Cenomanian of Texas.
Occurrence: This taxon is reported in the Albian Main Street and Cenomanian Grayson Formations of Texas, USA (Peck 1943; Rasmussen 1961; Hess 2015).
Roveacrinus sp.
Plate 1, panels f–h
Material: Thin section Rep. no. TPA.01-BO06; TPA.01-BO07; and TPA.01-BO08 (core samples Bz-1-4-7.1; Bz-1-4-9.1, and Bz-1-4-10.1).
Description: Plate 1, panel f displays a transverse section of an isolated radial showing the characteristic alar morphology, and an oblique-transverse section of a second primibrachial plate with an angular radial edge.
Plate 1, panel g illustrates an oblique section of primi- or secundi-brachial plate showing the characteristic alar morphology, associated with minute fragments of thin thecal wall (transverse sections).
Plate 1, panel h depicts an oblique section of a half theca showing an alar radial expansion.
Occurrence: In the present state of knowledge, R. alatus displays the same stratigraphical and geographical range than R. communis. However, this taxon dominates the roveacrinoidal assemblages in mud-supported sediments while R. communis dominates coarser-grained deposits. Genus Roveacrinus ranges from the Early Hauterivian of Spain (Ferré and Granier 1997a, b, 2000) to the Paleogene of Poland (Gorzelak et al. 2011).
Roveacrinidae indet.
Plate 1, panels d, e; Plate 2, panels b, f–h; Plate 3, panels a–h; Plate 4, panels a–h

Microfacies (thin section) photographs of roveacrinid ossicles, Lower-Middle Cenomanian, Adıyaman area, SE Turkey. a Roveacrinidae indet., TS-NBrn and TgS-IBr1, core sample B-1_1484.1; b Roveacrinidae indet., TS-NBrn, core sample D-1-2488.1; c Roveacrinidae indet., accumulation of broken brachial plates (NBrn), core sample D-1-2488.2; d Roveacrinidae indet., TS-Theca and Lg/TgS-NBrn, core sample G-1-2980; e Roveacrinidae indet., OblS-IBr2, core sample G-1-2992; f Roveacrinidae indet., TgS-IBr1, OblS-NBrn and TgS-IBr2, core sample S-2_2202_5x; g Roveacrinidae indet. (sturdy morphology), OblS-NBrn, core sample S-2_2206; h Roveacrinidae indet. (sturdy morphology), OblS-NBrn, core sample S-2_2206_5x

Microfacies (thin section) photographs of roveacrinid ossicles, Lower-Middle Cenomanian, Adıyaman area, SE Turkey. a Roveacrinidae indet. (sturdy morphology), OblS-NBrn, core sample S-2_2206_10x; b Roveacrinidae indet., TS-NBrn, core sample S-2-2202.1; c Indeterminable roveacrinoidal sections, core sample S-2-2274.1; d Roveacrinidae indet., IBr2 and NBrn, core sample B-1_1484.2; e Roveacrinidae indet., accumulation of brachial plates (OblS-NBrn), core sample Bk-1-1962.4; f Roveacrinidae indet., accumulation of brachial plates NBrn, core sample D-1-2552; g Roveacrinidae indet., TS-NBrn and TgS-IBr2, core sample D-1-2492; h Roveacrinidae indet., TgS-IBr2, core sample K-1-1630
Material: Thin sections Rep. no TPA.01-SV01/8; TPA.01-DK01/5; TPA.01-BO04/6; TPA.01-BA03; TPA.01-GK01/2; TPA.01-BZ01; and TPA.01-KM01 (core samples S-2-2202.3; D-1-2552.2; S-2-2202.5; Bz-1-4-7.2; Bz-1-4-9.2 (ex Bz-1-4.9.10x); Bz-1-4-10; B-1_1484.1; D-1-2488; D-1-2488.2; G-1-2980; G-1-2992; S-2_2202_5x; S-2_2206; S-2_2206_5x; S-2_2206_10x; S-2-2202.1; S-2-2274.1; B-1_1484.2; Bk-1-1962.4; D-1-2552; D-1-2492; and K-1-1630, respectively).
Description: All the illustrations mentioned above generally concern the many oblique sections of indeterminable brachial plates. For they do not usually bear significant features to reach a precise diagnosis (at a generic and/or specific level), they are left in open nomenclature until further additional data towards generic identification. However, they are clearly distinguishable from brachial plate sections of Saccocomidae for they do not display the flat, thin-shelled, wing-like “Saccocoma microfacies” but are rather sturdy and “ornamented.”
Occurrence: In the present state of knowledge, this family is reported from the lower Hauterivian (Ferré and Granier 2000) to the upper Maastrichtian (Jagt 1999; Gale 2016, 2017), with isolated occurrences in the Neogene of Poland (Salamon et al. 2010; Gorzelak et al. 2011). They are best-known from the mid-Cretaceous deposits (Albian-Cenomanian-Turonian) that have focused much of stratigraphers’ attention in search for paleobiological traces of anoxic events.
Family Saccocomidae Orbigny, 1852
Genus Applinocrinus Peck, 1973
(= Microcalamoides Bonet, 1956)
Applinocrinus sp.
Plate 2, panel c
Material: Thin section Rep. no. TPA.01-BO01 and TPA.01-BO02 (core sample Bz-1-4-7.3 (ex Bz-1-4.7)).
Description: This taxon is represented by a unique longitudinal transverse section of a thecal plate (Lg/TS-Rad), showing a faintly corrugated wall comparable to the so-called Microcalamoides (as junior synonym, and facial look-alike, see Ferré et al. 1999, 2018).
Occurrence: This isolated section clearly evidences post-mortem breakage and/or short transportation.
Saccocomidae indet.
Plate 2, panels c–e
Material: Thin sections Rep. no. TPA.01-BO01; TPA.01-BO02 and TPA.01-BA01 (core samples Bz-1-4-7.3 (ex Bz-1-4.7); Bz-1-4-7; and B-1_1508.1, respectively).
Description: Along with an isolated thecal plate section, Plate 2, panel c yields tangential sections of second primibrachial (TgS-IBr1) and indeterminable brachial plates (OblS-NBrn). Plate 2, panel d illustrates some tangential sections of indeterminable plates (those sections most likely recall radial ridges born by second primibrachial plates), while Plate 2, panel e shows arachnoid sections typical of outer ornamentation distal brachial plates (Obl/TS-N/IIBrn/Rad), but without any significant specific feature. Given the restricted taxonomical information born by these brachial plates, and the absence of definite microstructures on these sections, we decided to leave these saccocomid sections in open taxonomy til further additional knowledge.
Occurrence: Saccocomids are usually found in shallow, mud-supported bottom environment under low hydrodynamics and document more restricted and shallowing/shallower environmental conditions. Their radial plates sections are seldom found connected while their “winged” brachial plates (so-called Saccocoma- or Lombardia-facies) are found isolated and embedded in deeper hemipelagic mudstone carbonates.
Stratigraphic meaning and potential
The roveacrinoidal assemblages of SE Turkey display no direct stratigraphic inference of index species. Nevertheless, these assemblages are rather diversified and comparable with the other Lower Cenomanian assemblages found in adjacent countries. But we must stress out that these are less diversifed than the coeval assemblages from the US Gulf Coast (Peck 1943, 1948; Hess 2015; Gale 2017), and mainly consist in weakly ornamented species. Despite this lack of specific stratigraphic value, their recurring occurences can be used to define abundance levels (environmental and/or eustatic meaning) correlatable between boreholes on a intra-basinal scale, or at a wider scale (Arabian Platform, E-W Tethysian-wide correlations, etc.). Further works will have to focus on a detailed compiling of such abundance levels both in cores and field sections, within the scope of framing a comprehensive list of successive biostratigraphic events, among which roveacrinid abundance levels would have a lead part, as they are having nowadays in northern Africa (Benyoucef et al. 2017; Ferré et al. 2017).
Palaeo-environmental proxies and correlation to adjacent areas
All the study boreholes are scattered over SE Turkey; therefore, we can assume that the roveacrinoidal material, found as crumbled and disarticulated pieces within mudstone facies, provides fair and reliable insights about the roveacrinoidal population during the Early Cenomanian: the roveacrinoid skeletons were not transported far away, even stirred by weak bottom currents, and locally dismantled, and scattered within mud-supported sediments. Roveacrinids, as well as saccocomids, were hemipelagic to pelagic organisms, with possible escape response to escape predators. Subsequently to their early planktonic larval stage inducing a wide dispersal of any echinoderm brood, these roveacrinoidal relics can be regarded as potential biostratigraphic index species, and environmental proxies as well. Since they are usually found associated with calcisphere blooms, and their own abundance occurrences are positively correlated with those of first-level, surface carbonate producers (blooms of calcareous dinocysts, calcispheres, and heterohelicidids; Ferré 1997), they have been regarded as opportunistic organisms. The Upper Cretaceous rocks of SE Turkey show unexpectedly the dual presence of roveacrinids and saccocomids. Since roveacrinids have been found so far over the Arabian Platform more frequently in carbonate-grained, open-ramp marine environments (Fig. 4), their co-occurrence advocates for a mixed environment or, at least, supports that saccocomids were swept secondarily from a shallower but quieter environment. Furthermore, we may wonder about their potential role (HMC echinodermal plates as enhancing agent) in the dolomitization further south and the poronecrosis of potential reservoir rocks (Fig. 5).

Geographic distribution of roveacrinoid facies over SE Turkey
Conclusions
The finding of roveacrinoid ossicles in the Cenomanian deposits of the Adıyaman area more vividly supports their extensive presence and stratigraphic potential. The roveacrinoid assemblages of southeastern Turkey are very similar to those previously described from the NE Syria area (Ferré et al. 1997, ongoing study) but show noticeable discrete differences, among which the faint presence of saccocomids, and the high morphological diversity of roveacrinids.
Successive microfacies analyses of the Derdere Formation of the Adıyaman area in southeastern Turkey documented so far six roveacrinid taxa: Roveacrinus communis Douglas, 1908 (= R. derdereensis Manni, in FARINACCI & Manni); Roveacrinus cf. alatus Douglas, 1908; Roveacrinus spinosus Peck, 1943; Roveacrinidae indet.; Applinocrinus sp.; and Saccocomidae indet.; as well as many unidentifiable roveacrinoid remains.
Despite scarce or lacking data, some general trends regarding the quantitative distribution of roveacrinoid microfacies can be outlined. Because biogeographic distribution is controlled by many interconnected factors, a direct positive correlation appears probable between roveacrinoid population booms and calcisphere blooms. This positive link can easily be explained as a predator-prey relationship within the marine food chain: the passive suspensivorous filter-feeding roveacrinids feeding on the “snow” of calcisphere-secreting algae sinking onto the sea floor (Masters and Scott 1978, 1979).
Furthermore, within the framework of petroleum exploration (field observations or core analysis), some simple occurrence or accumulation levels can be used as baseline marker beds for local datation, inner-field or even inter-regional correlation depending on the relative abundance of roveacrinid remains.
As a result of their early planktonic life cycle, roveacrinids are found widespread in Tethysian oceanic settings and are thought to have dwelled from the outer-ramp to the upper-slope environments. Consequently, such abundance and/or accumulation levels are suitable for worldwide correlation, all correlatable with eustatic changes.
References
Benyoucef M, Mebarki K, Ferré B, Adaci M, Bulot L-G, Desmares D, Villier L, Bensalah M, Frau C, Ifrim C, Malti F-Z (2017) Litho- and biostratigraphy, facies patterns and depositional sequences of the Cenomanian-Turonian deposits in the Ksour Mountains (Saharan Atlas, Algeria). Cretac Res 78:34–55
Benzaggagh M, Homberg C, Ben Abdesselam-Mahdaouid S (2018) Saccocomid crinoid sections and biozones of the upper Jurassic in the western Tethyan realm. İn: Villaseñor Martínez A-B, Rosales Domínguez C, Olóriz-Sáez F (eds) 10th International Congress on the Jurassic System, San Luis Potosí, Mexico, Feb. 4-9, Abstracts Volume, Paleontologia Mexicana 3:20–24
Bonet F (1956) Zonification microfaunística de las calizas del Este de Mexico. Bol Asoc Mex Geól Petrol VIII(7–8):3–102
Bryant G-F (1960) Stratigraphic report of the Korudag area, Petroleum District VI, Southeast Turkey (American Overseas Petroleum (AMOSEAS) Report): Petrol İşleri Genel Müdürlüğü Teknik Arşivi, Kutu no: 323, Rapor no 14, 23s. [unpublished]
Cater J-M-L, Gillcrist J-R (1994) Karstic reservoirs of the mid-Cretaceous Mardin Group, SE Turkey: tectonic and eustatic controls on their genesis, distribution and preservation. J Pet Geol 17:253–278
Celikdemir E, Dulger S (1990) Güneydoğu Anadolu’da Mardin Grubu karbonatlarının stratigrafisi, sedimantolojisi ve rezervuar özellikleri. TPAO Rapor No: 2665. 93s. [unpublished]
Celikdemir E, Dulger S, Gorur N, Wagner C, Uygur K (1991) Stratigraphy, sedimentology, and hydrocarbon potential of the Mardin Group, SE Turkey. Spec Publ Eur Assoc Petrol Geosci 1:439–454
Coruh T (1981) Mardin Grubu karbonatlarının X. Bölge Bölükkonak-2/A, Alıçlı-2, Dodan-6M, Tavan-1, Kavikadağ-1 kuyularıyla; IX. Bölge Körkandil, Ballıkay ve Mehmet Yusuf ölçülmüş stratigrafik kesitlerindeki mikropaleontolojik incelenmesi: TPAO Araştırma Merkezi, Rapor no 403/A, 12s. [unpublished]
Coruh T (1983) XII. Bolge Çemberlitaş-4, 5, Adıyaman- 1 ve Durukaynak- 1 kuyularıyla, Karababa ve İnişdere ösk’larindaki Derdere ve Karababa formasyonları hakkında yeni mikropaleontolojik bulgular. TPAO Rapor No: 549. 16s. [unpublished]
Cros P, Dercourt J, Günay Y, Fourcade É, Bellier J-P, Lauer J-P, Manivit H, Kozlu H (1991) La plate-forme arabe en Turquie du sud:une rampe carbonatée albo-turonienne effondrée au Sénonien. Bull Centres Rech Explor Prod Elf-Aquitaine 15(1):215–237
Cros P, Dercourt J, Günay Y, Fourcade É, Bellier J-P, Lauer J-P, Manivit H, Kozlu H (1999) The Arabian Platform in Southeastern Turkey: an Albian-Cenomanian carbonate ramp collapsed during the Senonian. TAPG Bull 11(1):55–77
Douglas JA (1908) A note on some new chalk crinoids. Geol Mag 5(5):357–359
Duran O (1991) Beşikli, Tokaris, Bakacak ve Taşlik sahalarının stratigrafisi, sedimantolojisi ve rezervuar özellikleri. TPAO Rapor No: 1586. 43s. [unpublished]
Erenler M (1989) XI-XII. Bölge güney alanlarındaki kuyularda Mezosoyik çökel istifinin mikropaleontolojik incelenmesi: TPAO Araştırma Merkezi, Rapor no. 1364. 44s. [unpublished]
Ertug K (1991)Güneydoğu Anadolu Kretase yaşlı Mardin Grubu ve Karaboğaz formasyonu’nun palinostratigrafisi: TPAO Araştırma Merkezi, Rapor no, 1525. 49s. [unpublished]
Farinacci A, Manni R (2003) Roveacrinids from the Northern Arabian Plate in SE Turkey. Turk J Earth Sci 12:209–214
Ferré B (1997) “Lombardia”-facies and saccocomid-like sections in Cretaceous sediments: whose pieces? Mineralia Slovaca 29(4–5):336–337
Ferré B, Berthou P-Y (1993) Roveacrinidal remains from the Cotinguiba formation (Cenomanian-Turonian) of the Sergipe Basin (NE-Brazil). In: 13o Congresso Brasileiro de Paleontologia, 1o Simposio Paleontologico do Cone Sul, Sao Leopoldo, Brazil, Sept. 19–26. Boletim de Resumos, p 119
Ferré B, Berthou P-Y (1994) Roveacrinidal remains from the Cotinguiba formation (Cenomanien-Turonien) of the Sergipe Basin (NE-Brazil). Acta Geol Leopold XVIII(39/1):299–313
Ferré B, Dias-Brito D (1999) Microfacies recognition of Roveacrinid families Lombardia, Eothrix, Globochaete and other strains. In: Dias-Brito D, de Castro JC, Rohn R (eds) Boletim do 5o Simposio sobre o Cretaceo do Brasil, pp 651–659
Ferré B, Granier B (1997a) The Albian stemless microcrinoids (Roveacrinidae, Crinoidea) of the Congo Basin (Angola). Gaea Heidelbergensis 3:129–130
Ferré B, Granier B (1997b) Roveacrinus berthoui, nov. sp., the earliest representative of the family Roveacrinidae (Roveacrinida, Crinoidea) in the lower Hauterivian of Busot (Alicante, Spain). In: Final Regional Meeting of IGCP Project 362 “Tethyan and Boreal Cretaceous”, Stara Lesna, Slovakia, Sept. 29–Oct. 5, 1997, Mineralia Slovaca, Bratislava 29(4–5):338–339
Ferré B, Granier B (2000) Roveacrinus berthoui nov. sp., Early Hauterivian representative of Roveacrinidae (Roveacrinida, Crinoidea) of Busot (Alicante, Spain). Geol Carpath 51:101–107
Ferré B, Granier B (2001) Albian roveacrinids from the southern Congo Basin, off Angola. J S Am Earth Sci 14:219–235
Ferré B, Cros P, Fourcade É (1997) Tethyan Mid-Cretaceous (Cenomanian-Turonian) Roveacrinids (Roveacrinida, Crinoidea) as stratigraphical and paleobiogeographical tools. In: Final Regional Meeting of IGCP Project 362 “Tethyan and Boreal Cretaceous”, Stara Lesna, Slovakia, Sept. 29–Oct. 5, 1997, Mineralia Slovaca, Bratislava 29(4–5):267–268
Ferré B, Fernandez-Gonzalez M, Dias-Brito D (1999) New insight into Microcalamoides Bonet, 1956—revised systematics and subsequent bearings. In: Dias-Brito D, de Castro JC, Rohn R (eds) Boletim do 5o Simposio sobre o Cretaceo do Brasil, pp 661–668
Ferré B, Mebarki K, Benyoucef M, Villier L, Bulot L-G, Desmares D, Benachour H-B, Marie L, Sauvagnat J, Bensalah M, Zaoui D, Adaci M (2017) Roveacrinids (Crinoidea, Roveacrinida) from the Cenomanian-Turonian of Southwest Algeria (Saharan Atlas and Guir Basin). Ann Paléontol 103(3):185–196. https://doi.org/10.1016/j.annpal.2017.03.001
Ferré B, Granier B, Gorzelak P, Salamon M-A (2018) Cretaceous roveacrinids from Mexico revisited: overcoming the taxonomic misidentifications and subsequent biostratigraphic abuse. Bol Soc Geol Mex 70(2):499–530. https://doi.org/10.18268/BSGM2018v70n2a12
Gale A-S (2016) Roveacrinida (Crinoidea, Articulata) from the Santonian-Maastrichtian (Upper Cretaceous) of England, the US Gulf Coast (Texas, Mississippi) and southern Sweden. Pap Palaeontol 2:489–532
Gale A-S (2017) An integrated microcrinoid zonation for the lower Campanian chalks of southern England, and its implications for correlation. Cretac Res 87:312–357
Gorzelak P, Salamon M-A, Ferré B (2011) Pelagic crinoids (Roveacrinida, Crinoidea) discovered in the Neogene of Poland. Naturwissenschaften 98:903–908
Handfield R-W, Bryant G-F, Keskin C (1959) Measured section, Korudağ (American Overseas Petroleum). TPAO Arama Grubu, Arşiv no. 523. 10s. [unpublished]
Hess H (2015) Roveacrinids (Crinoidea) from the mid-Cretaceous of Texas: ontogeny, phylogeny, functional morphology and lifestyle. Swiss J Palaeont 134:77–107
Hess H, Messing C-G (2011) Treatise on Invertebrate Paleontology, Part T, Echinodermata 2 Revised, Crinoidea 3. XXIX+261 pp, University of Kansas Press, Lawrence, Kansas
Jagt J-M-W (1999) Late Cretaceous-Early Palaeogene echinoderms and the K/T boundary in the southeast Netherlands and northeast Belgium—Part 1: Introduction and stratigraphy, Part 2: Crinoids. Scr Geol 116:1–155
Kaufmann F-J (1865) Polythalamien des Seewerkalkes. In: Heer O (ed) Die Urwelt der Schweiz, pp 194–199
Kellogg H-E (1960) Stratigraphic report, Hazro area, Petroleum District V, SE Turkey. (American Overseas Petroleum (AMOSEAS) Report): Petrol İşleri Genel Müdürlüğü Teknik Arşivi, Kutu no: 126, Rapor no.1, 42s. [unpublished]
Kellogg H-E (1961) Regional stratigraphy and petroleum possibilities of Southeast Turkey. Petroleum District V, SE Turkey. (American Overseas Petroleum (AMOSEAS) Report): Petrol İşleri Genel Müdürlüğü Teknik Arşivi, Kutu no: 201, Rapor no.1, 29s. (TPAO Arama Grubu, Rapor no.767). [unpublished]
Koyluoglu M (1981) Körkandil ölçülmüş stratigrafik kesitinin Mezosoyik biyostratigrafisi: TPAO Araştırma Merkezi, Rapor no.362, 14s. [unpublished]
Lombard A (1937) Microfossiles d’attribution incertaine du Jurassique supérieur alpin. Eclogae Geol Helv 30:320–331
Lombard A (1945) Attribution de microfossiles du Jurassique supérieur alpin à des Chlorophycées. Eclogae Geol Helv 38:163–173
Masters B-A, Scott RW (1978) Microstructure, affinities and systematics of Cretaceous calcispheres. Micropaleontology 24:210–221
Masters B-A, Scott R-W (1979) Calcispheres and Nannoconids. In: Fairbridge RW, Jablonski D (eds) The Encyclopedia of Paleontology. Dowden, Hutchinson & Ross Inc., Stroudsburg, pp 167–169
McClusky S, Balassanian A, Barka C, Demir S, Ergintav I, Georgiev O, Gurkan M, Hamburger K, Hurst H, Kahle K, Kastens G, Kekelidze R, King V, Kotzev O, Lenk S, Mahmoud A, Mishin M, Nadariya A, Ouzounis D, Paradissis Y, Peter M, Prilepin R, Reilinger I, Sanli H, Seeger A, Tealeb M, Toksöz N, Veis G (2000) Global Positioning System constraints on plate kinematics and dynamics in the eastern Mediterranean and Caucasus. J Geophys Res 105(B3):5695–5719
Miller J-S (1821) A natura history of the Crinoidea or lily-shaped animals, with observation on the genera Asteria, Euryale, Comatula, and Marsupites. 150 pp, Bryan & Co. (eds), Bristol
Mulayim O, Mancini E, Çemen I, Yilmaz I-O (2016) Upper Cenomanian-Lower Campanian Derdere and Karababa formations in the Çemberlitaş oil field, southeastern Turkey: their microfacies analyses, depositional environments, and sequence stratigraphy. Turk J Earth Sci 25:46–63
Orbigny d’ AD (1852, for 1850) Prodrome de paléontologie stratigraphique universelle des animaux mollusques et rayonnés faisant suite au cours élémentaire de paléontologie et de géologie stratigraphique. vol 2, 427 pp., Masson (ed), Paris
Peck R-E (1943) Lower Cretaceous crinoids from Texas. J Paleontol 17(5):451–475
Peck R-E (1948) A Triassic crinoid from Mexico. J Paleontol 22(1):81–84
Peck R-E (1955) Cretaceous microcrinoids from England. J Paleontol 29(6):1019–1029
Peck R-E (1973) Applinocrinus, a new genus of Cretaceous microcrinoids and its distribution in North America. J Paleontol 47(1):94–100
Perincek D, Duran O, Bozdogan N, Coruh T (1991) Stratigraphy and paleogeographical evolution of the autochthonous sedimentary rocks in SE Turkey. Ozan Sungurlu Symp., Ankara, Turkey, Proceedings pp 274–305
Rasmussen H-W (1961) A monograph on the Cretaceous Crinoidea. Biol Skr K Dan Vid Selsk 12(1):1–428
Salamon M-A, Gorzelak P, Ferré B, Lach R (2010) Roveacrinids (Crinoidea, Echinodermata) survived the Cretaceous-Paleogene (K-Pg) extinction event. Geology 38:883–885
Schneider H-L (1987) Zur Kelchmorphologie und Systematik der Roveacrininae PECK, 1943 (Crinoidea, Oberkreide). Neues Jb Geol Paläontol Abh 175:181–206
Schneider H-L (1989) Zur Morphologie und Ontogenese von Roveacrinus geinitzi n. sp. (Crinoidea, Oberkreide). Neues Jb Geol Paläontol Abh 178:167–181
Scott R-W, Root S-A, Tenery J-H, Nestell M (1977) Morphology of the Cretaceous microcrinoid Poecilocrinus (Roveacrinidae). J Paleontol 51:343–349
Sieverts-Doreck H (1952). In: Moore RC, Lalicker CG, Fischer AG (eds), Invertebrate fossils. 766 pp, McGraw-Hill, New York
Tuna D (1973) VI. Bolge litostratigrafi adlamasinin aciklayici raporu: TPAO Rapor No.813. 131s. [unpublished]
Verniory R (1954) Eothrix alpina Lombard, Algue ou Crinoïde? Arch Sci Geneva 7(4):327
Verniory R (1955) Extension géographique et stratigraphique du genre Saccocoma Agassiz dans le Dauphiné méridional et en Provence. Arch Sci Geneva 8:225–226
Verniory R (1956) La création du genre Lombardia Brönnimann est-elle justifiée? Arch Sci Geneva 9(1):86
Verniory R (1960) Présence (et variétés) de Saccocoma tenella Goldfuss à Talloires (Haute-Savoie). Arch Sci Geneva 13:250–257
Verniory R (1961) Présence de Saccocoma quenstedti Doreck (in coll.) dans les gorges de la Méouge (Sisteron Provence). Arch Sci Geneva 14:315–320
Verniory R (1962) Une nouvelle forme de Saccocoma (Montbrand, Hautes-Alpes, France). Arch Sci Geneva 15:391–397
von Zittel K-A (1879, for 1876-1880) Handbuch der Paläontologie. Band I, Paläozoologie: Abt. 1. VII+765 pp, Oldenbourg R (ed), München & Leipzig
Wagner C, Pehlivanlı M (1985) Karst geological interpretation of the Mardin carbonates in the Çemberlitaş field. TPAO Report No 2051. 19s. [unpublished]
Wright D-F, Ausich W-I, Cole S-R, Peter M-E (2017) Phylogenetic taxonomy and classification of the Crinoidea (Echinodermata). J Paleontol 91(4):829–846. https://doi.org/10.1017/jpa.2016.141
Acknowledgements
The authors all wish to express their gratitude to the Turkish Petroleum Corporation (TPAO) and Research Center (ARGEM) for granting access to borehole material and permission to publish. We thank Dr. Robert W. Scott and an anonymous reviewer for their valuable suggestions and comments.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article

Cite this article
Mulayim, O., Yilmaz, I. & Ferré, B. Roveacrinid microfacial assemblages (Roveacrinida, Crinoidea) from the Lower-Middle Cenomanian of the Adıyaman area (SE Turkey). Arab J Geosci 11, 545 (2018). https://doi.org/10.1007/s12517-018-3901-z
Received:
Accepted:
Published:
DOI: https://doi.org/10.1007/s12517-018-3901-z