
Abstract
One stratigraphic section named Nimbolook succession in the northwest of Qayen city was measured and sampled. Based on identified foraminifera Berthelina intermedia assemblage zone, Rotalipora appenninica and Biticinella subbreggiensis interval zones were proposed. These biozones coincide with global biozones. According to foraminifera fauna, an Early–Late Albian age was suggested for this study. Palaeoecological studies led to the recognition of three benthic foraminifera (MG1–3) and two planktonic morphogroups (MG4–5). In the lower and middle part of the section, the abundance of benthic and agglutinated foraminifera (MG1–3) is more than planktonic foraminifera (MG4–5), respectively. Calcareous benthic morphogroups could be attributed to aerobic, shallow to slightly deep water, and eutrophic to mesotrophic environment. Agglutinated benthic morphogroups suggested that aerobic, mesotrophic, to eutrophic environments are characteristic by a dominance of arenaceous shallow infaunal specimens which are active deposit feeders. Planktonic foraminifera morphogroups with trochospiral and smooth test have been seen near the surface water, aerobic to semi-aerobic environment. Increasing planktonic foraminifera and specially keeled species could be a suggested recovery in paleoenvironmental conditions.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Based on geological structure, Iran has been divided into several structural units. For the understanding and good description of these areas, it is necessary to know these structural units, characterized by a relatively unique record of stratigraphy, tectonics, magmatic activities, orogenic events, and overall geological style. The first systematic geological studies in Iran started in the late 1960s. Stocklin (1968) suggested several basins for the first time. Later, using mostly the NIOC database and important investigations, Nabavi (1976), Eftekharnejad (1981), and Berberian and King (1981) classified Iran into several different structural zones (units) namely Zagros, Sanandaj-Sirjan, Central Iran, Kopet Dagh, Alborz, and Eastern Iran. However, in recent years, new interpretations and models have been offered regarding the geological setting of Iran (Nogol-e-Sadat and Almasian 1993; Alavi 1991; Aghanabati 2009). Eastern Iran can be divided into two parts namely Lut block and Flysch or colored mélange of Zabol–Baluch Zone. Lut block located to the west of Zabol–Baluch Zone is the main body of Eastern Iran. Lut block extends for about 900 km in a north–south direction. Lut block is part of the eastern microcontinental area of the Central Iran. Cretaceous sediments comprised of two different facies. In some parts of south Birjand, Cretaceous succession occurs in carbonate form and belongs to Early to Late Cretaceous (Aghanabati 2009). Flysch Zone (Zabol–Baluch) is located between Lut block to the west and Helmand (in Afghanistan) to the east. Flysch Zone is highly deformed and tectonized and consists of thick deep-sea sediments (Fig. 1).

Iranian major tectono-sedimentary units (redrawn from Berberian and King 1981): stable area, Arabian Precambrian platform in southwest and Turanian Hercynian plate in northeast (1); Zagros, including Zagros foredeep, main sector of the marginal active fold belt peripheral to stable area and High Zagros (2); Alborz Mountains (3); Central Iran lying between the two marginal active fold belts (4); Talesh, Armenian Late Hercynian belt with a possible continuation to Iranian Talesh Mountain (5); ZabolBaluch and Makran post ophiolite flysch trough (6); Kopet Dagh folded belt and foredeep (7)
Recent detailed geological studies of the area were carried out by Fauvelet and Eftekhar-nezhad (1990), Berthiaux et al. (1991), and Raisossadat et al. (2011, 2014). The rudists were studied by Raissosadat and Skelton (2005) and Khazaei et al. (2011). The Orbitolina limestones were studied by Babazadeh et al. (2010) and Zarei et al. (2014).
The main aims of this study are to determine the benthic and planktonic foraminifera from the Late Aptian to Early Cenomanian interval of the Nimbolook succession at the east of Lut block in Eastern Iran for the first time. The biostratigraphic studies and applying to suggestion biozones, classification of foraminifera contents to morphogroups, and interpretation of the paleoechological conditions based on them are the other aims of this study.
Materials and methods
In order to study foraminifera, a total of 60 samples both hard and soft were collected from Nimbolook section. The shale and marl samples were crushed to little particles (2–3-cm diameters), soaked in tap water with diluted hydrogen peroxide 10 % during 48 h, and then washed, and the residue was sieved at 125, 80, and 60 μm; then, all of foraminifera were picked under a binocular microscope, while SEM was used at Ferdowsi Mashhad University for taxonomic identification of foraminifera assemblages. The real thickness has been calculated, and stratigraphical section was drawn by using computer software packages. Excel software has also been used for drawing charts and graphs.
Stratigraphy
The Cretaceous succession anywhere in the East and Central Iran begins with coarse grain sediments that gradually change to fine grain including sandstone, limestone, shale, marl, and thick-bedded limestone.
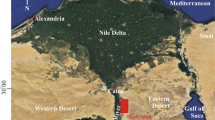
The measured section of Nimbolook is located about 30 km northwest of Qayen. For access to the measured section from Qayen–Gonabad road, a track road passable by car is used for about 9 km in the southwest of Nimbolook village. The exact location of the section is at 33° 52′ 9 N latitude and 59° 00′ 49 E longitude (Fig. 2). The Nimbolook succession with 222-m thickness conformably overlies and underlies the limestone beds and could be divided into four units (Fig. 3). The first unit consists of
-
1.
16 m of gray to light gray medium-bedded limestone beds, in the base containing bivalvia.
-
2.
20 m of gray to greenish gray sandy limestone with few meters of thin-bedded greenish gray sandstone was partly seen at the base of this unit.
-
3.
122-m alternation of light brown thin-bedded shale and marl gray to greenish gray medium-bedded sandy limestone.
-
4.
64-m massive to medium-bedded limestone of this unit that changed to green to greenish gray, weathered color brown thin-bedded limestone (Fig. 4)

Geographical names and their positions, location of measured section

A field photograph showing outcrop of the section

Stratigraphical column of Nimbolook section
Assemblage of foraminifera and biostratigraphy
Bed-by-bed sampling at the Nimbolook succession for biostratigraphic studies provides a firm basis for a biozonation in the Late Aptian–Early Cenomanian age. This study led to the identification of 12 genera and 13 species of benthic foraminifera as well as 13 genera and 18 species of planktonic foraminifera, making a total of 6 genera and 6 species.
Many biozonations have been proposed by different authors, based on the planktonic foraminifera in Tethys realm. The most important are Caron (1985), Bolli (1966), Hardenbol et al. (1998), and Premoli-Silva and Verga (2004). In this study, the three biozones suggested coincide with global biozones. The following benthic foraminifera Berthelina baltica, Berthelina intermedia, Dorothia sp., Gaudryina rugosa, Guttulina communis, Gyroidinoides infracretaceus, Lenticulina macrodisca, Lenticulina saxocretacea, Lenticulina subalata, Laevidentalina communis, Laevidentalina cf. soluta, Marginulina sp., Marssonella sp., Mayncina orbignyi, Pseudonodosaria sp., Orthokarstenia shastaensis, Saracenaria triangularis, and Saracenaria sp. and planktonic foraminifera Biticinella subbreggiensis, Favusella washitensis, Muricohedbergella delrioensis, Pseudothalmanninella subticinensis, Planomalina buxtorfi, and Rotalipora appenninica were identified, respectively, in the studied section. Based on these foraminifera, three biozones have been suggested (Fig. 5) namely (1) B. intermedia assemblage zone, (2) B. subbreggiensis interval zone, and (3) R. appenninica interval zone (Fig. 5).
-
1.
B. intermedia assemblage zone

Foraminifera range chart, proposed biozonation and stratigraphical column in Nimbolook section
This zone is an assemblage zone that was introduced by Holbourn and Kaminski (1997). B. intermedia is mentioned by Moullade (1974) and Weidich (1990) from Late Aptian to Cenomanian age, while Holbourn and Kaminski (1997) believed that this species starts from Albian.
This biozone is defined based on first appearance of B. intermedia and is recorded from 40–102 m of studied section (Fig. 5). The assemblage foraminifera in this biozone are Marssonella sp., G. communis, G. rugosa, L. communis, Dorothia sp., B. intermedia, G. infracretaceus, O. shastaensis, L. saxocretacea, L. macrodisca, Saracenaria sp. 1, and Saracenaria sp. 2. Based on these assemblage fauna, the Early Albian age has been suggested.
-
2.
B. subbreggiensis interval zone
This biozone was introduced by Postuma (1971) as interval zone for Middle Albian–Late Albian age. Researchers like Van Hinte (1976), Postuma (1971), Sigal (1977), and Premoli-Silva and Verga (2004) considered it as Middle Albian. This biozone includes 25 m (102 to 127) of the studied section. The lower boundary of this biozone is defined based on the first appearance of B. subbreggiensis, and the upper boundary of this biozone is based on the first appearance of R. appenninica (Fig. 5). The assemblage foraminifera in this biozone are Marssonella sp., L. saxocretacea, L. subalata, L. macrodisca, Saracenaria sp. 2, G. communis, and L. communis.
-
3.
R. appenninica interval zone
This zone is an interval zone introduced by Bronnimann (1952). Postuma (1971) placed this zone in the lower most part of Cenomanian, while others such as Bolli (1966), Sigal (1977), Carron (1985), and Premoli-Silva and Verga (2004) placed it in Late Albian. This biozone is defined based on the first appearance of R. appenninica up to the first appearance of Rotalipora brotzeni. The thickness of this zone is 33 m (127 to 160) (Fig. 5). The assemblage fauna includes P. buxtorfi, F. washitensis, M. delrioensis, P. subticinensis, B. baltica, Pseudonodosaria sp., Marginulina sp., L. subalata, Saracenaria sp. 1, Saracenaria sp. 2, and L. communis.
Foraminifera morphogroups
Previous studies of benthic morphological groups by Koutsoukos and Hart (1990) and Nagy (1992) revealed strong relationship between environmental conditions and morphotypes. According to the study of Corliss and Chen (1988), Koutsoukes and Hart (1990), and Coccioni and Galeotti (1993), there is an acceptable relationship between test morphology and inferred habitat of foraminifera fauna. Friedrich et al. (2003) is of the opinion that benthic foraminifera assemblages as well as stable isotope data can be used to interpret the oxygen content of bottom waters, organic matter flux to the sea floor, and sea-level changes during the Late Aptian time. In their study, Erbacher et al. (1998) stated that benthic foraminifera can serve as a sensitive tool to provide a better understanding of the conditions at the sea floor, during the Lower Cretaceous time.
The planktonic morphogroups are dependent on sea-level changes. Eicher and Worstell (1970) and Eicher (1967) have shown that the globigernid and globular morphotypes were the first planktonic foraminifera that appeared in transgression and the last to disappear during regression. The globular forms such as Globigerina inhabit surface waters and are keeled bioconvex and keeled plano-convex such as globorotallids which inhabit deeper waters (Hart and Bailey 1979; Wiedmann et al. 1982; Boudagher Fadel 2015).
Another approach is the successive appearance and disappearance of species or genera along gradients, e.g., water depth, with other factors also influencing the distribution of species. These may include the rise and fall of oxygen minimum zones (Jarvis et al. 1988; Leary et al. 1989; Koutsoukos and Hart 1990; Hart 1999) or nutrient supply (Premoli-Silva and Sliter 1999).
The foraminifera assemblage of the Nimbolook succession allowed three benthic (MG1–3) and two planktonic (MG4–5) morphogroups to be distinguished (Fig. 6). This classification is based on morphology, life position, feeding habitat, and environment of foraminifera.

Distribution of morphogroups of foraminifera in Nimbolook section
Morphogroup 1 (MG1)
This morphogroup consists of taxa with calcareous test and planispiral to trochospiral shape test which is typical for aerobic and eutrophic to mesotrophic environments. Most of these taxa are epifaunal forms and active deposit feeders that have been identified in the middle–outer shelf to upper slope deposits (Coccioni and Galeotti 1993; Lowery et al. 2014). B. baltica, B. intermedia, and Gyroidinoides infracretacea are the common examples of this group (Fig. 7 (1–3)). G. infracretaceus seems to be mainly controlled by paleobathymetry. The species is characteristic of outer shelf to middle bathyal depths in Albian strata of the South Atlantic Ocean (Kochhann et al. 2014).

Some selected SEM photographs of foraminifera that classified in morphogroups. Berthelina intermedia, sample no. MN 4-10, scale 100 μm (1a–1c); Anomalina ammonoides, sample no. MN4-29, scale 100 μm (2a–2c); Gyroidinoides infracretaceus, sample no. MN 4-12, scale 100 μm (3a and 3b); Dorothia sp., sample n. MN 4-15, scale 100 μm (4a and 4b); Gaudryina rugosa, sample no. MN 4-15, scale 100 μm (5); Marssonella sp., sample no. MN 4-17, scale 100 μm (6); Orthokarstenia shastaensis, sample no. MN 4-15, scale 100 μm (7); Laevidentalina communis, sample no. MN 4-25, scale 100 μm (8); Laevidentalina cf. soluta, sample no. MN 4-2, scale 100 μm (9); Marginulina sp., sample no. MN 4-29, scale 100 μm (10)
Morphogroup 2 (MG2)
This morphogroup is divided into two submorphogroups including the calcareous and agglutinated benthic foraminifera with elongate or subcylindrical test shape which is indicative of the following characteristic of environmental conditions. All these taxa are potentially shallow to deep infaunal, active deposit feeder, and mesotrophic to eutrophic conditions, which are present in a wide range of marine environments from inner shelf to upper bathyal (Ruckheim et al. 2006; Bindiu and Filipescu 2011). These faunae seem to be typical for moderately low levels of oxygen. Dorothia sp., G. rugosa, and Marssonella sp., could be classified as agglutinated benthic foraminifera in this morphogroup. In some taxa with elongated flattened to straight periphery, life position is shallow to deep infaunal, deposit feeder, neritic to upper bathyal, aerobic to dysaerobic, and mesotrophic to eutrophic environment conditions (Ruckheim et al. 2006). The most taxa with calcareous test belonging to this group are O. shastaensis, L. communis, L. cf. soluta, Marginulina sp., and Pseudonodosaria sp. This morphogroup seems to be well oxygenated and to prefer meso-eutrophic living conditions (Fig. 7 (4–10)).
Morphogroup 3 (MG3)
The foraminifera fauna was dominated by taxa such as Saracenaria sp. 1, Saracenaria sp. 2, G. communis, L. macrodisca, L. subalata, and L. saxocretacea, which could have the following specific characteristics: calcareous, biconvex test shape, active deposit feeders, and eutrophic to mesotrophic conditions as is the case for epifaunal to deep infaunal assemblages. They can be found in a wide range of environment from sublithoral to upper bathyal and aerobic to dysaerobic conditions (Ruckheim et al. 2006) (Fig. 8).

Some selected SEM photographs of foraminifera that classified in morphogroups. Lenticulina macrodisca, sample no. MN 4-17, scale 100 μm (1a–1c); Lenticulina saxocretacea, sample no. MN 4-21, scale 100 μm (2a–2c); Saracenaria sp. 1, sample no. MN 4-21, scale 100 μm (3a and 3b); Saracenaria sp. 2, sample no. MN 4-12, scale 100 μm (4); Lenticulina subalata, sample no. MN 4-25, scale 100 μm (5a–5c); Guttulina communis, sample no. MN 4-21, scale 100 μm (6a and 6b)
The diversity and specific distribution of benthic assemblage foraminifera are mainly influenced by oceanic circulation patterns, oxygenation and nutrient availability, surface fertility, changes in depth of the CCD, and variations within the seawater chemistry, such as pH (Van der Zwaan et al. 1999). The preservation of benthic foraminifera in Nimbolook section varies from good to poor throughout the studied interval.
Planktonic morphogroup (MG4)
This morphogroup includes trochospiral forms which are living in surface or near surface and suspension feeding (Price and Hart 2002; Lowery et al. 2014). They live in shallow epicontinental sea in a pelagic, aerobic, highly eutrophic environment. M. delrioensis, F. washitensis, and B. subbreggiensis are examples of these morophogroups (Leckie 1987; Kochhann et al. 2013) (Fig. 9). The oxygen isotope data supports the view that the Cenomanian globular foraminifera, such as the genus Muricohedbergella, inhabited near-surface waters that are rich in organic material (Caron and Homewood 1982; Leckie 1989; Premoli-Silva and Sliter 1999; Price and Hart 2002).

Some selected SEM photographs of foraminifera that classified in morphogroups. Muricohedbergella delrioensis, sample no. MN 4-37, scale 100 μm (1a–1c); Favusella washitensis, sample no. MN 4-37, scale 100 μm (2a–2c); Muricohedbergella delrioensis, sample no. MN 4-34, scale 100 μm (3a–3c); Biticinella subbreggiensis, sample no. MN 4-29, scale 100 μm (4a and 4b)
From the standpoint of view of paleobiogeogaphy and distribution pattern, species of Muricohedbergella belong to shallow and open marine (Hart and Bailey 1979; Leckie 1985). According to Jarvis et al. (1988) and Corfield et al. (1990), small planktonic forms such as Muricohedbergella lived near the surface and bigger forms such as Rotalipora in deeper depth. Muricohedbergella planispira is a proxy for normal salinity surface and low-oxygen waters (Eicher and Worstell 1970; Leckie 1987).
Keller and Pardo (2004) studied foraminifera of the Pueblo basin of Colorado and concluded that the abundance of M. delrioensis coincides with increases of oxygen. This species lived in rich nutrient and normal salinity sea. Fisher and Arthur (2002) believed that hedbegellids lived in different levels of water such that M. delrioensis is located under M. planispira.
Planktonic morphogroup (MG5)
This morphogroup includes trochospiral forms floating in nearly deep water of upper part of bathyal to lower part of surface water (Norris and Wilson 1998). Pseudothalmanninella, Rotalipora, and Planomalina are the most abundant specimens belonging to this morphogroup (Fig. 10). According to Keller and Pardo (2004) and Leckie (1987), normal salinity and aerobic and oligotrophic environment are the characters in this habitat.

Some selected SEM photographs of foraminifera that classified in morphogroups. Rotalipora appenninica, sample no. MN 4-37, scale 100 μm (1a–1c); Planomalina buxtorfi, sample no. MN 4-39, scale 100 μm (2a–2c); Pseudothalmanninella subticinensis, sample no. MN 4-29, scale 100 μm (3a–3c)
Discussion and conclusion
For the first time in eastern margin of Lut block, foraminifera contents of Albian deposits have been investigated that led to the following results.
The Nimbolook succession has a thickness of about 222 m at the studied section including sandstone, limestone, shale, and marl. It is conformably overlain, underlies the limestone beds, and could be divided into four units.
The stratigraphic distribution of the 13 benthic and 6 planktonic identified foraminiferal species enabled to suggest three biozones as follows: (1) B. intermedia assemblage zone, (2) B. subbreggiensis interval zone, and (3) R. appenninica interval zone. They coincide with the global biozones. Based on foraminifera fauna, an Early–Late Albian age is suggested for the studied area.
Palaeoecological conditions have been reconstructed. This study lead to the recognition of three benthic foraminifera and two planktonic morphogroups. The preservation of benthic species shows relatively good to moderate in lower and middle part of Nimbolook section.
A detailed analysis of the studied succession appeared a high abundance of the benthic morphogroups (MG1–3) in the lower and middle part and the planktonic morphogroups (MG4–5) in the upper part of the section, respectively. The first morphogroup is epifauna, the second is infauna, and the third is assemblage of epifauna and infauna morphogroups. It seems that living conditions for benthic morphogroups were better than in the upper part of the section.
The most abundant species of calcareous and agglutinate benthic foraminifera in morphogroups 1 to 3 are Berthelina, Lenticulina, and Laevidentalina; Dorothia; and Gaudryina, respectively. Muricohedbergella and Rotalipora are the most common species in planktonic foraminifera that belong to morphogroups 4 and 5.
Because of the planktonic morphogroups (MG4–5) that lived in deeper water to compare with benthic morphogroups (MG1–3), it could be concluded that the depth of basin from lower to upper of succession increased.
According to the identified morphogroups in Nimbolook succession, during the Early–Late Albian–Early Cenomanian time, this section exposited in shallow to fairly deep basin.
References
Aghanabati A (2009) Geology of Iran geological survey of Iran. In Persian
Alavi M, compiler (1991) Tectonic map of the Middle East: geological survey of Iran scale1: 5,000,000
BabazadehS A, Raisossadat SN, Ahrari F (2010) Biostratigraphy and evolutionary study of the Cretaceous orbitolinids in the sedimentary deposits of east Lut, south west of Qayen. Sedimentary Facies 3(1):1–10 English Abstract
Berberian M, King GCP (1981) Towards a palaeogeography and tectonic evolution of Iran. Can J Earth Sci 18:210–265
Berthiaux A, Christmann P, Fauvelet E, Hatrival JN, Vaslet D, Vahdati Daneshmand A (1991) Quadrangle geological map of Qayen Scale: 1/250000 geological survey of Iran
Bindiu R, Filipescu S (2011) Agglutinated foraminifera from the northern TarcăuNappe. Studia UBB Geologia 56(2):31–41
Bolli HM (1966) Zonation of Cretaceous to Pliocene marine sediments based on planktonic foraminifera. Boletin Informativo Asociacion Venezolana de Geologia. MineriayPetroleo 9:9–32
BouDagher-Fadel MK (2015) Biostratigraphic and geological significance of planktonic foraminifera, Second Edition. UCL Press
Bronnimann P (1952) Globigerinidae from the upper Cretaceous (Cenomanian-Maestrichtian) of Trinidad. Bull Am Paleontol 34:5–71
Caron M (1985) Cretaceous Planktic foraminifera. In: Bolli H.M, Saunders J, Perch-Nielsen K, (eds) Plankton Stratigraphy 1: 17–86, Cambridge University
Caron M, Homewood P (1982) Evolution of early planktic foraminifers. Mar Micropaleontol 7:453–462
Coccioni R, Galeotti S (1993) Orbitally induced cycles in benthonic foraminiferal morphogroups and trophic structure distribution patterns from the Late Albian Amadeus Segment central Italy. J Micropalaeontol 12:227–239
Corfield RM, Hall MA, Brasier MD (1990) Stable isotope development for foraminiferal habitats during the development of the Cenomanian/Turonianoceanic anoxic event. Geology 18:175–178
Corliss BH, Chen C (1988) Morphotype patterns of Norwegian Sea deep sea benthic foraminifera and ecological implications. Geology 16:716–719
Eftekharnejad J (1981) Tectonic division of Iran with respect to sedimentary basins. J Iranian Petroleum Soc 82:19–28 [in Persian]
Eicher DL (1967) Foraminifera from Belle Fourche shale and equivalents Wyoming and Montana. J Paleontol 41(1):167–188
Eicher DL, Worstell P (1970) Cenomanian and Touronian foraminifera from the Great Plains United States. Micropaleontology 16:269–324
Erbacher J, Gerth W, Schmiedl G, Hemleben C (1998) Benthic foraminiferal assemblages of late Aptian-early Albian black shale intervals in the Vocontian Basin SE France. Cretac Res 19:805–826
Fauvelet E, Eftekhar-nezhad J (1990) Explanatory text of the Qayen, Quadrangle map 1: 250,000. Geological Survey of Iran 200 pp
Fisher CG, Arthur MA (2002) Water mass characteristics in the Cenomanian US western interior seaway as indicated by stable isotopes of calcareous organisms. Palaeogeogr Palaeoclimatol Palaeoecol 188:189–213
Friedrich O, Reichelt K, Herrle JO, Lehmann J, Pross J, Hemleben C (2003) Formation of the Late Aptian Niveau Fallot black shale in the Vocontian Basin SE France: evidence from foraminifera, palynomorphs, and stable isotopes. Mar Micropaleontol 49:65–85
Hardenbol J, Thierry J, Farley MB, Jacquin T, De Gracian-sky P-C, Vail PR (1998) Cretaceous sequence chronostratigraphy. In: De Graciansky P-C, Hardenbol J, Jacquin T, Vail PR (eds) Mesozoic and Cenozoic Sequence Stratigraphy of European Basins. Special Publication Society Sedimentary Geology 60 (chart)
Hart MB (1999) The evolution and biodiversity of Cretaceous planktonic Foraminiferida. Geobios 32(2):247–255
Hart MB, Bailey HW (1979) The distribution of planktonic Foraminiferida in the Mid-Cretaceous of NW Europe. Int Union Geol Sci Seri A 6:527–542
Holbourn A, Kaminski M (1997) Lower Cretaceous deep-water foraminifera of Indian Ocean. Grzybowski Foundation Special Publication No 4:172 p
Jarvis I, Carson GA, Cooper MKE, Hart MB, Leary PN, Tocher BA, Horne D, Rosenfeld A (1988) Microfossil assemblages and the Cenomanian-Turonian Late Cretaceous oceanic anoxic event. Cretac Res 9:3–103
Keller G, Pardo A (2004) Age and paleoenvironment of the Cenomanian–Turonian global stratotype section and point at Pueblo Colorado. Mar Micropaleontol 51:95–128
Khazaei A, Raisossadat SN, Asadi S (2011) Rudist bivalves (Requieniidae family) in Early Cretaceous sediments from SW Qayen Eastern Iran, Paleobiogeographic aspects. Sedimentary Facies 3(2):52–67 English Abstract
Kochhann KGD, Koutsoukos DAM, Fauth G, Sial AN (2013) Aptian–Albian planktic foraminifera from DSDP site 365564 (offshore Angola): biostratigraphy, paleoecology and palebroceanographic significance. J Foraminifer Res 43(4):443–463
Kochhann KGD, Koutsoukos EAM, Fauth G (2014) Aptian-Albian benthic foraminifera from DSDP site 364 (offshoreAngola): a paleoenvironmental and paleobiogeographic appraisal. Cretac Res 48:1–11
Koutsoukos EAM, Hart MB (1990) Cretaceous foraminiferal morphogroup distribution patterns palaeocommunities and trophic structures: a case study from the Sergipe Basin, Brazil. Trans R Soc Edinb Earth Environ Sci 81:221–246
Leary PN, Carson GA, Cooper MKE, Hart MB, Horne D, Jarvis I, Rosenfeld A, Tocher BA (1989) The biotic response to the late Cenomanian oceanic anoxic event integrated evidence from Dover SE England. J Geol Soc Lond 146:311–317
Leckie RM (1985) Foraminifera of the Cenomanian-Turonian boundary interval Greenhorn formation Rock Cabyon Atlantic Peublo Colorado. In: Pratt LM, Kauffman EG, and Zelt F (eds.) Fine-Grained Deposits and Biofacies of the Cretaceous Western Interior Seaway Evidence of Cyclic Sedimentary Processes. Soc. Econ. Min. and Paleontol. Field Trip Guide book No. 4, 139–149
Leckie RM (1987) Paleoecology of mid-Cretaceous foraminifera, a comparison of open ocean and epicontinental sea assemblages. Micropaleontology 33:164–176
Leckie RM (1989) Apaleoceanographic model for the early evolutionary history of planktonic foraminifera. Palaeogeogr Palaeoclimatol Palaeoecol 73:107–138
LoweryCh M, Corbett MJ, Leckie RM, Watkins D, Romero AM, Pramudito A (2014) Foraminiferal and nannofossilpaleoecology and paleoceanography of the Cenomanian–Turonian eagle ford shale of southern Texas. Palaeogeogr Palaeoclimatol Palaeoecol 413:49–65
Moullade M (1974) Zones de Foraminifères de Crétacéinférieurmésogéen. C R Acad Sci Paris Ser D 278:1813–1816
Nabavi MH (1976) An introduction to geology of Iran. Geological survey of Iran (In Persian)
Nagy J (1992) Environmental significance of foraminiferalmorphogroups in Jurassic North Sea deltas. Palaeogeogr Palaeoclimatol Palaeoecol 95:111–134
Nogole-Sadat MAA, Almasian M (1993) Tectonic map of Iran. Scale 1:1,000,000. Geological Survey of Iran
Norris RD, Wilson PA (1998) Low-latitude sea-surface temperatures for the mid-Cretaceous and the evolution of planktic foraminifera. Geology 26(9):823–826
Postuma JA (1971) Manual of planktonic foraminifera. Elsevier Publishing Company
Premoli Silva I, Sliter WV (1999) Cretaceous paleoceanography evidence from planktonic foraminiferal evolution. Geol Soc Am Spec Pap 332:301–328
Premoli Silva I, and Verga D (2004) Practical manual of Cretaceous planktonic foraminifera.International School on planktonic foraminifera 3 course Cretaceous Universities of Perugia and Milan Perugia 283 pp
Price GD, Hart MB (2002) Isotopic evidence for early to mid-Cretaceous ocean temperature variability. Mar Micropaleontol 46:45–58
Raisossadat SN, Skelton PW (2005) First record of rudist fauna from the Qayen area Eastern Iran. 7th International Cretaceous Symposium, Neuchatel, Switzerland
Raisossadat SN, Mosavinia A, Khazaei AR, Asadi Sh, (2011) Stratigraphy of Cretaceous deposits in Southwest of Qayen area (East of Iran). Proceeding of the 5th Symposium of Iranian Paleontological Society [English Abstract]
Raisossadat SN Asadi Sh, Zarei H, Khazaei AR, Mortazavi M, Motamedalshariati M, Mosavinia A, Moloudi D, Yazdi Moghadam M (2014) Stratigraphy paleoecology and paleobiogeography of Cretaceous deposits in the east of Lut Block (Qayen area). Proceeding of 18th Symposium of Geological Society of Iran, 600–610, Tarbiat Modarres University, Iran
Rückheim S, Bornemann A, Mutterlose J (2006) Planktic foraminifera from the mid-Cretaceous (Barremian–Early Albian) of the North Sea basin. Palaeoecological and palaeoceanographic implications. Mar Micropaleontol 58:83–102
Sigal J (1977) Essai de Zonation du Cr’etac’eme’diterraneen a l’aide des foraminifers planctoniques. Géol Mediterr 4:99–108
Stocklin J (1968) Structural history and tectonics of Iran: a review. Am Assoc Pet Geol Bull 52:1229–1258
Van der Zwaan GJ, Duijnstee IAP, den Dulk M, Ernst SR, Jannink NT, Kouwenhoven TJ (1999) Benthic foraminifers: proxies or problems? A review of paleoecologicalconcepts. Earth-Sci Rev 46:213–236
Van Hinte JE (1976) A Cretaceous time scale. Am Assoc Pet Geol Bull 60(4):498–516
Weidich KF (1990) Die kalkalpine unterkreide und ihre foraminiferen fauna. Zitteliana 17:1–312
Wiedmann J, Butt A, Einsele G (1982) Cretaceous stratigraphy environment and subsidence history at the Moroccan continental margin. In: von Rad U et al (eds) Geology of the northwest African continental margin. Springer, Berlin, pp. 336–395
Zarei H, Raisossadat SN Yazdi Moghadam M, Mortazavi Mehrizi M, (2014) Lower Cretaceous paleoecology in the Qomenjan section, southwest Qayen, based on benthic foraminifera. Proceeding of the 8th Symposium of Iranian Paleontological Society, 21–22 May 2014, Zanjan University, Zanjan, Iran
Acknowledgments
Comments by two anonymous reviewers were highly appreciated and significantly improved this manuscript. We thank Dr. Brian Huber and Ann Holbourn for their comments in identifying some specimens and providing some useful papers. We appreciate the Department of Geology, University of Birjand for providing laboratory facilities and some logistic support in the course of the field work.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article

Cite this article
Motamedalshariati, M., Raisossadat, S.N., Moluodi, D. et al. Foraminifera biozonation and morphogroups from Nimbolook section, east margin of Lut block, Iran. Arab J Geosci 9, 720 (2016). https://doi.org/10.1007/s12517-016-2709-y
Received:
Accepted:
Published:
DOI: https://doi.org/10.1007/s12517-016-2709-y