
Abstract
The Ordivician-Khabour Formation from the Northern Thrust Zone of Iraqi-Kurdistan comprises between 500 and 800 m of alternating predominantly greenish-gray sandstones, siltstones, and shales. The succession has revealed an abundant ichnofossils characterized by 11 ichnogenus, namely: Helminthopsis, Gordia, Cruziana, Rusophycus, Monomorphichnus, Rhizocorallium, Thalassinoide, Planolite, Paleophycus, Deplocraterion, and Skolithose. Ethologically, the ichnofossils are distributed among domichnia and fodinichnia groups. This association of ichnofossils contains elements of the Skolithose and Cruziana ichnofacies. The presence of these ichnofacies indicates variations in energy of the sedimentary environment. High-energy condition with sandy-shifting substrate in foreshore zone is related to the Skolithos ichnofacies, while the Cruziana ichnofacies indicates unconsolidated-soft substrate developed under low-energy condition in the shoreface/offshore zone. These ichnogenera indicate shoreface-offshore zone of shallow-marine environment for the deposition of the rocks of the Khabour Formation.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
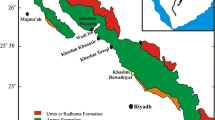
Strata belonging to the Khabour Formation crop out around Kaista and Nazdur villages, about 40 km northeast of Zakho city (Fig. 1). Several sedimentological studies have been made on the Khabour Formation in Iraqi-Kurdistan, and different sedimentary facies have been defined (e.g., Al-Bassam 2010). However, there is not yet a detailed study on the biogenic-sedimentary structures of these sediments. The study of biogenic-sedimentary structures in ancient sediments has provided important criteria that are commonly used to understand depositional environments recorded by sedimentary rocks. Published record of biogenic-sedimentary structures from the Ordovisian-Khabour Formation includes general descriptions and determinations of the stratigraphic age of formation (Seilacher 1963). Based on the sedimentary characteristics, the Khabour Formation is suggested to have deposited in a shallow to deeper marine environment, where a turbidite-affected deeper facies are recognized for the upper parts (Seilacher 1963; Buday 1980; Karim 2006; Al-Bassam 2010).

Geologic map of the Northern Thrust Zone, Iraqi-Kurdistan with location of the study area (Modified from Sissakian 2000)
The presence of evidence of organisms within the sediment is known as biogenic structures (ichnofossils) and is a very common feature in sedimentary rocks. Because they show minimal effects of compaction and of early diagenesis, ichnofossils may be the principal or only preserved biogenic evidence of pleoenvironmental conditions in thick stratigraphic intervals (Pemberton et al. 1992). especially in those dominated by nearshore facies. Therefore, biogenic structures provide critical data for the interpretation of ancient depositional environment. The existence of different trace fossil associations in the Khabour Formation has been recognized since the 1960s (e.g., Seilacher 1963). However, the implications and significance of theses associations have not been clear. The recognition that the Khabour Formation encompasses a wide variety of depositional settings (Al-Bassam 2010) allows the author to place the ichnological information within a palaeoenvironmental framework.
From the perspective of the analysis of sedimentary rocks, ichnofossils will often be more useful than fossil shells or bones because they are conclusive evidence that an animal lived there. They are indicators of depositional environment (Savrda 1995) and help to record sedimentation rate; where rates of sedimentation are high, the sea floor is covered by loose sandy or muddy material and a variety of ichnofacies occur according to the water depth, U-Shaped burrows and escape structures reflect more rapid sedimentation, while a complex feeding and grazing traces would reflect slow rates of sedimentation (Pemberton et al. 1992). Sediment exposed on the sea floor starts to consolidate if the rate of sedimentation is relatively slow and a firmground forms. Some organisms (such as bivalves, echinoids, and some sponges) are able to bore into rock to create dwelling traces, these indicate a break or very slow rate in sedimentation allowing seafloor cementation to take place (Nagendra et al. 2010).
In this study, the author uses ichnologic evidence from the out crops of the Khabour Formation in order to evaluate the paleoenvironmental implications of abundant members of the Skolithos and Cruziana ichnofacies in this formation.
Geological setting
Iraqi-Kurdistan is northeastern part of the Arabian Peninsula which is region of tectonic compression of particularly unstable area marked by convergent movements of the Arabian and Eurasian Plates that closed the Tethys paleo-ocean and formed the Northern Thrust Zone and the Zagross Fold Belt in the northeastern part of the Arabian Plate (Buday and Jassim 1987). Paleozoic formations in Kurdistan are cropping out on “restricted area” in the Northern Thrust Zone, definably, about 1-km south of Iraqi-Turkish border (Fig. 1).
The only surely ascertained formation belonging to the Cambro-Ordovician cycle in Iraq is the Khabour Formation (Jassim and Goff 2006). The Northern Thrust, which includes the studied area, is the widest in the west with asymmetrical domal culminations toward the south and occasionally the southern limbs are overturned (Fig. 1). The northern limbs of the structures have lower dips and are cut by reverse faults. The geologic structure of the studied area is characterized by a fold-thrust style which is developed along the plate margin during the Cretaceous and is characterized by thrusted anticlinal structures. Complicated faulting and folding, present mainly within the Northern Thrust Zone, has also caused much deformation of the sedimentary rocks. Deformation affected both Paleozoic and Cretaceous-Tertiary strata (Aqrawi et al. 2010). Minor faults are located perpendicular to the orientation of the main thrusts. The rocks exposed in the study area belong to the Paleozoic succession include five intracratonic sedimentary cycles predominated by siliciclastic or mixed siliciclastic-carbonate units (Jassim and Goff 2006). The Paleozoic cycles commence within the Ordovician with the deposition of the Khabour Formation. This was followed in Silurian times by the Akkas Formation and this is unconformably overlain by the Middle-Late Devonian to Early Carboniferous cycle, represented by the Chalki, Pirispiki, Kaista, Ora, and Harur formations. The overlying Permo-Carboniferous cycle is represented by the Ga’ara Formation. The uppermost cycle is late Permian in age and comprises the Chia Zairi Formation. The Paleozoic succession contains a series of muddy units distributed throughout the stratigraphy.
Khabour formation: lithology and depositional environment
The Paleozoic succession of Iraq is exposed in the extreme northern part of the country (Fig. 1) and can be traced south and west wards in the subsurface. The Khabour Formation, about 800-m exposed thickness of siliciclastic deposits in the type section, is the oldest rock unit exposed in Iraq dated as Ordovician. It was first described by Wetzel (1950) in the area North of Zakho city, in the valley of the Khabour River in the Kurdistan, where the formation got its name and where the type section is located. It is unconformably overlain by the Pirispiki Formation (Wetzel 1950).
The lithology, as described by Wetzel (1950). consists of alternation of thin-bedded, fine-grained sandstones, quartzites (Cruziana-rich), and silty micaceous shales, olive green to brown in color. The quartzites are generally cross-bedded, and the bedding planes are well surfaced with smooth films of greenish micaceous shales. Metamorphism is very slight in the thin-bedded shales with quartzites and almost unnoticeable in the thicker shale beds (Wetzel 1950). The sedimentary structures observed in the Khabour Formation at the exposed sections include laminations, ripple marks, load casts, and slump structures. The quartzite beds are also truncated by overlying beds and show fucoid markings, in filling trails and burrows, pitted surfaces and, other bedding-plane structures of unknown origin (Bellen et al. 1959; Al-Hadithi 1972; Isa’ac 1975). Based on the field observations of the Khabour Formation from the studied outcrop (Fig. 2a), the formation consists of cyclic alternations of fining upward sequences of sandstone-siltstone and silty micaceous shale with clear upper surfaces of individual cycles. Sandstones are thin-bedded to massive, silty, and are fine-grained, locally bioturbated (Fig. 2b). Ripple laminations, lenticular, and flaser beddings are observed in the sandstones. Siltstones are characterized by small-scale cross-stratification and wave ripples. The upper contact of the shale horizons is sharply terminated and bioturbated. Micaceous minerals form smooth surfaces with metallic luster marking the upper contact of each cycle. The quartzite is generally fine grained, often laminated, and occasionally bioturbated. The silty shale is ferruginous, slope-forming, ranging in thickness from few centimeters (in the lower part) to few meters (in the upper part).

Khabour Formation outcrop. a Road cut section shows cyclic alternations of sandstone-siltstone and silty micaceous shale. b Highly bioturbated (Thalassinoides ichnofossil) silty-sandstone bed obliterating all evidence of primary bedding and physical sedimentary structures
Based on Cruziana tracks throughout the section, the age was given by Wetzel (1950) as Cambro-Ordovician. However, in later works, the age was determined as Middle to Late Ordovician (Llandeilo) (Seilacher 1963). The Khabour formation is believed to have deposited in a tide- and storm-dominated shallow marine intertidal environment of mud- and silt-flats with occasional emergence (Bellen et al. 1959). Karim (2006) has suggested that the formation in north Iraq was deposited in a spectrum of environments including fluviatile, deltaic, shelf, slope, and deep marine. According to Jassim and Goff (2006). the formation, in outcrop sections, passes from littoral facies in the “Ora anticline” into a deeper marine turbidite facies in “Kaista anticline” (Khabour area). Al-Bassam (2010) suggested that the sequence of graded beddings of quartz sand, silt, and micaceous shale in Khabour Formation appears to have been deposited in marine environment ranging from intertidal to outer shelf realms.
Material and methods
Ichnofossil assemblages described in this paper have been collected from the southern limb of the Kaista anticline from the Northern Thrust Zone (Fig. 1). Road cut-sections were studied during the fieldwork (Fig. 2a). Ichnological data was gained through observation and description of the outcrops. Ichnological observations have been concentrated on the identification of ichnogenera, the intensity of bioturbation, and the distribution of trace fossil assemblages (Pemberton and Frey 1984). Material was difficult to collect without damage and therefore was mostly studied in the field. However, some samples were collected and sectioned to analyze the nature of burrow infills and shed light on taphonomic aspects. In describing trace fossils, especially trilobite traces, the author mainly follows terminology proposed by Crimes and Harper (1970). Frey (1975) and Seilacher (1970, 2007). The bioturbation index (BI) was quantified in the field and in the laboratory according to the Taylor and Goldring (1993). Based on Taylor and Goldring (1993). a bioturbation index is ranging from 0 (no bioturbation) to 6 (complete bioturbation). Hand-picked samples were collected, and a selection of them is housed at the Geology department, College of Science, Salahaddin University, Iraq.
Biogenic-sedimentary structures
Description of ichnofossils
The current study of the biogenic-sedimentary structures was based on the ichnogenera identification, description, and distribution. A total of 11 ichnogenera have been recognized in Khabour Formation.
The ichnogenus Helminthopsis (Heer 1877) has been found at the base of sandstone interbedded with silty-sandstone beds. This ichnite occurs as positive hyporelief, forming complex structural patterns, and oriented parallel to bedding plan. Helminthopsis composed of unbranched, irregularly winding or meandering, horizontal burrows or trails that do not touch or cross themselves (Fig. 3a and c). It is distinguished from Gordia by its tendency to meandering and the absence of self-overcrossing. Burrow diameter is constant throughout, typically alternating between winding straight course. Maximum observed length of the burrow is 150 mm and diameter is 6 mm. Burrow fill is massive.

Ichnogenera from the Khabour Formation. a Ichnogenus Helminthopsis (arrow). b Ichnogenus Gordia. c Ichnogenus Cruziana (Cr) with ichnogenus Helminthopsis (He). d Ichnogenus Rusophycus. e Ichnogenus Monomorphichnus (Mo) with undetermined trace (Un). f Ichnogenus Rhizocorallium
The ichnogenus Gordia (Emmons 1844) occurs mainly as positive hyporelief in the muddy sandstone beds. The burrow is composed of horizontal to sub-horizontal traces with sinuous, straight, or spiral forms (Fig. 3b). Usually, traces are intersecting and generate a “false truncation” pattern. In section, traces are approximately cylindrical, with uniform diameters ranging from 1 to 5 mm. No well-defined lining is observed, and filling material is slightly differing from host rock.
The ichnogenus Cruziana (d’Orbigny 1842) usually occurs in the finer grained portion of the sandstone facies interbedded with sandy mudstone, forming convex hyporelief traces. Cruziana traces consist of bilobate ridges with well-developed internal lobes separated by a relatively shallow median groove. The lobe is covered by discrete scratch marks arranged obliquely to the median groove (Fig. 3c). Trail trajectory slightly curved. The longest, bilobate trace is up to 150 mm long. Width of the complete structure varies from 35 to 125 mm and of the median groove is 5 mm in average.
The ichnogenus Rusophycus (Hall 1852) was found at the base of sandstone interbedded with silty-sandstone beds. This trace is preserved as convex epireliefs 50–110-mm long by 25–52-mm wide, with a poorly to moderately developed median furrow extending the entire length of the trace, dividing it into two lobes which bear imbricate or unevenly concentrated, weakly developed, evenly spaced scratching forming a very obtuse V-angle (Fig. 3d).
The ichnogenus Monomorphichnus (Crimes 1970) has not been commonly identified in Khabour Formation. It typically occurs in sandstone interbedded with sandy-mudstone facies. Monomorphichnus occurs as sets of isolated, straight to slightly curved parallel ridges separated from one another which are repeated laterally. The ridges vary in length from 8 to10 mm, in width from 1.5 to 2 mm and are 2–3 mm apart from each other (Fig. 3e). It is typically preserved as convex hyporelief; however, concave epirelief traces have also been observed.
The only example of ichnogenus Rhizocorallium (Zenker 1836) has rarely been found at the base of sandstone interbedded with sandy mudstone beds. This ichnite occurs as positive hyporelief, forming sub-horizontal to oblique U-shaped spreite burrow (Fig. 3f), although complete specimens are scarce. The spreiten structure is poorly preserved and dim. The burrow preserves some very faint scratch marks. Maximum burrow length varies between 150 and 18 mm and burrow width between 40 and 15 mm.
Ichnogenus Thalassinoides (Ehrenberg 1944) is preserved as hypichnial and enichnial traces and is mainly found at the top and locally in the center of sandstone and silty-sandtone beds, with smooth, unlined, three-dimensional, horizontal burrow system showing Y/T-shaped branching (Fig. 4a), horizontal network connected to surface by more or less vertical shaft (Fig. 2b). In section, traces are rounded to elliptical shape with massive-burrow fill, commonly similar to the host rock. Burrow-depth penetration is difficult to determine but the section of shaft on the bedding planes is 2–18 in diameter.

Ichnogenera from the Khabour Formation. a Ichnogenus Thalassinoides. b Ichnogenus Planolites (Pl) with ichnogenus Paleophycus (Pa). c Ichnogenus Diplocraterion. d Ichnogenus Skolithos (arrow)
Ichnogenus Planolites (Nicholson 1873) and ichnogenus Paleophycus (Hall 1847) are mainly found in sandstone with lamination beds. They are consisting of endichnial, straight to gently curved, unornamented, cylindrical unbranched horizontal to sub-horizontal burrows (Fig. 4b). Traces are found as convex hyporelief or concave epirelief. In section, traces range from constantly cylindrical to slightly ovalized, with 1–15 mm in diameter. Planolites ichnites are unlined and filled with material that is usually darker than the host rock, due to the high mud concentration (Fig. 4b). Paleophycus is less abundant, and is distinguished from Planolites by having a distinct wall lining (Fig. 4b) and sediment fill slightly different from the lithology of the host rock (Pemberton and Frey 1982).
The ichnogenus Diplocraterion (Torell 1870) is found at the top of the sandstone beds. It is regarded either as a dwelling (Cornish 1986; Ekdale and Lewis 1991) or as an equilibrium structure (Bromley 1996). Diplocraterion occurs as cylindrical, vertical, “U”-shaped burrows with 10–15 mm in diameter with spreite that are commonly preserved as endichnia in very fine- to medium-grained sandstone beds (Fig. 4c). Traces with the complete preserved section are rare, usually being found as two narrow cylinders at the bed tops, conjugated or not (Fig. 4c). Spreiten is generally bad defined.
Ichnogenus Skolithos (Haldeman 1840) is endichnial burrows and is mainly found at the top and locally in the center of sandstone beds, with straight, vertical to slightly inclined unbranched cylindrical tubes (Fig. 4d). Traces with good preservation are distinct and lined. In section, traces are rounded to elliptical shape with massive-burrow fill, commonly similar to the host rock. Burrow-depth penetration is difficult to determine but ring-like structure on the bedding planes with 2–15 mm in diameter.
Assemblages of ichnofossils
Ichnofossil assemblages are mainly reported on the soles and tops of sandstone and siltstone beds and locally within silty-sandstone beds in units where these sandstones are overlain by silty-shale beds. The abundance of burrows in individual beds thicker than 5 cm increases upwards, whereas in the thinner beds it tends to be constant. However, the ichnofossils of the Khabour Formation represent the open-marine assemblages and can be grouped into four main ichnofossil assemblages: Skolithose-Deplocraterion-Planolite-Paleophycus assemblage (IA 1), Skolithose-Thalassinoide-Planolite-Rhizocorallium assemblage (IA 2), Thalassinoide-Planolite-Cruziana-Monomorphichnus-Rusophycus assemblage (IA 3), and Cruziana-Helminthopsis-Gordia assemblage (IA 4).
The Skolithose-Deplocraterion-Planolite-Paleophycus assemblage (IA 1) is found in cross-bedding thick-sandstone beds. This assemblage is characterized by the predominance of vertical to sub-vertical burrows of Skolithos and Deplocraterion and not so abundant horizontal traces of the Planolite and Paleophycus ichnogenera. In the sandstone beds, which are embraced by this assemblage, the physical sedimentary structures clearly dominated over biogenic structures. Although overall ichnodiversity is moderate, individual beds commonly exhibit only one or two ichnogenus. The bioturbation disturbance grading of this assemblage is variable, although beds with this assemblage typically present bioturbation index 3. This assemblage is dominated by elements of the Skolithos ichnofacies.
The Skolithose-Thalassinoide-Planolite-Rhizocorallium assemblage (IA 2) is typical of laminated-sandstone beds, locally ripple-cross-laminated, quartzose, fine- to (very rarely) medium-grained sandstones. In contrast to Skolithose-Deplocraterion-Planolite-Paleophycus assemblage, feeding structures of deposit-feeders are more important components in this assemblage, presenting bioturbation indexes 3 and 4. Well-defined tiers are observed. This ichnofossil assemblage mixes elements of both the Skolithos and Cruziana ichnofacies.
The Thalassinoide-Planolite-Cruziana-Monomorphichnus-Rusophycus assemblage (IA 3) is recorded in laminated-sandstone beds and fine-grained quartzose silty sandstones. Although some dwelling traces of suspension-feeders may be present, but the assemblage is dominated by feeding structures of deposit feeders. Burrows are extremely compacted, displaying some flattened cross sections instead of circular or subcircular. The bioturbation index is typically 3–4 and, locally, 5. Bioturbation is commonly restricted to the top of sandstone beds, and suites are typically monospecific, although Planolite, Cruziana, and Monomorphichnus are present in places. The possible Rusophycus ichnite occurs in association with this assemblage. The association is an example of the Cruziana ichnofacies (Frey and Pemberton 1984). The high density of ichnofossils obliterates all evidence of primary bedding and physical sedimentary structures in most beds (Fig. 2b).
The Cruziana-Helminthopsis-Gordia assemblage (IA 4) occurs in the very fine-grained, quartzose sandstone interbedded with silty sandstones and siltstones beds. Horizontal and sub-horizontal traces of Cruziana and Helminthopsis, and inclined to sub-vertical traces of Gordia are predominant in this assemblage. The bioturbation is dominated by feeding structures of deposit feeders with index 2 and 3. The association represents an example of the Cruziana ichnofacies (Frey and Pemberton 1984).
Environmental significance: a discussion
The distribution of ichnofossils in the Ordovician-Khabour succession reflects the paleoecological conditions including the availability of oxygen and nutrient influx (e.g., Parcha et al. 2005; MacEachern et al. 2007) and indicates that the ichnocenosis is dominated by a high behavioral diversity ranging from suspension to deposit feeders (Seilacher 2007). However, the ichnofossil assemblages of the Khabour Formation characteristically display the development of Skolithos and Cruziana ichnofacies arranged in order from shorface to offshore in term of depositional systems (MacEachern et al. 2007).
Ichnofossil assemblages IA 1 and IA 2 are generally the members of the Skolithos ichnofacies and are characterized by trace fossils produced by suspension feeders. The presence of Planolite and Rhizocorallium ichnogenera with the Skolithose, Deplocraterion, and Thalassinoide ichnogenera (Table 1) represents distal-Skolithose ichnofacies and indicates middle to lower shoreface environment with aerobic condition (Bann and Fielding 2004; MacEachern et al. 2005, 2007). The sediment types and distinguished biogenic structures associated with the soft-sediment deformation structures indicate relatively high to moderate energy conditions, and shifting substrate have been exploited by the opportunistic animals in the shoreface/offshore systems. The Skolithos ichnofacies suggests that the depositional setting was relatively well oxygenated associated with relatively high levels of wave or current energy and typically occurred in well-sorted loose or shifting substrates. These conditions are interpreted to reflect deposition in a shoreface and sheltered foreshores, but similar conditions may occur also in high-energy shallow-water environments (MacEachern et al. 2005, 2007; Singh et al. 2008).
On the other hand, the association of the Thalassinoidem, Planolite, Cruziana, Monomorphichnus, and Rusophycus ichnogenera (IA 3) with the Helminthopsis and Gordia ichnogenera (IA 4) (Table 1) reflects the transition from proximal- to distal-Cruziana ichnofacies and indicates lower shoreface to offshore environment (MacEachern et al. 2005, 2007). However, the fine-grained sediments of the Khabour Formation embrace Cruziana, Planolites, Palaeophycus, Rhizocorallium, and Thalassinoides which are the member of the Cruziana ichnofacies. This sedimentary sequence also comprises of some facies crossing trace fossils like Planolites, Thalassinoide, and Helminthopsis. The Cruziana ichnofacies refers commonly to the unconsolidated poorly sorted muddy substrates shift into the shallow marine settings with uniform salinity (Pemberton et al. 1992). These conditions typically range from moderate energy levels, lying below fair-weather (minimum) wave base, but above storm wave base to lower energy levels in deeper, quieter waters (Pemberton et al. 1992; MacEachern et al. 2007; Singh et al. 2008). Therefore, it shifts into a shoreface to proximal offshore settings where the energy level is dropping and allows settling of fine-grained sediments. The horizontal forms of ichnologenera are a slightly impoverished Cruziana ichnofacies with an upward-increasing abundance of sub-horizontal and vertical forms typically associated with the Skolithos ichnofacies. The dominance of horizontal forms of Helminthopsis and Gordia which are developed at the water-sediment interface suggests very fine-grained unconsolidated substrate representing the low-energy offshore condition. The dominance of horizontal to sub-horizontal feeding structures of the Planolites and Palaeophycus indicates exploitation of organic detritus in sediment accumulated during relatively low-energy conditions. These ichnogenera, however, occur few centimeters below the sediment-water interface, suggesting unconsolidated substrate experiencing relatively moderate to low-energy shoreface/offshore conditions.
The analyzed interval records, from base (?) to top, a Cruziana ichnofacies zone, a mixed Cruziana-Skolithose ichnofacies zone, and a Skolithose ichnofacies zone (Table 1). This trend records progressively higher energy conditions from Cruziana ichnofacies to Skolithose ichnofacies. The presence of a mixed Cruziana-Skolithose ichnofacies in the middle interval may reflect changes in environmental energy levels, instead of water depth. As presented by numerous studies (e.g., Crimes et al 1981; Ekdale 1988; Buatois and Lopez Angriman 1992; Bromley 1992). ichnofacies distribution is controlled by local environmental factors such as salinity, oxygen concentration, substrate character, food supply, and sedimentation rate, rather than bathymetry. Thus, ichnofacies may be good indicators of the various relationships between the ichnofossils and environment.
The rarity of distinct biogenic structures in shales suggests low levels of bioturbation in these rocks. However, abundant casts of burrows on the lower surfaces of sandstone beds underlain by shale conversely indicate an intensive bioturbation of sales. Intense bioturbation of the shales is also suggested by their irregular, rough splitting, a feature typical of totally bioturbated deposits. Most probably, the rare occurrence of distinct bioturbation structures in shales was caused by the negligible contrast between burrow-fills and the host sediment (cf. Bromley 1996; Uchman 1999; Schieber 2003).
Conclusions
Ichnofossils are relatively diverse and abundant in the Khabour Formation and are mainly found in sandstone, silty-sandstone, and locally silty-shale lithologies. The lack of distinct biogenic structures in shales results from an insufficient contrast of burrows produced in these sediments while they possessed a soupy consistency. A total of 11 ichogenera has been identified, and they include the following: Helminthopsis, Gordia, Cruziana, Rusophycus, Monomorphichnus, Rhizocorallium, Thalassinoide, Planolite, Paleophycus, Deplocraterion, Skolithose, and one undeterminded. Ethologically, these ichnogenera are interpreted as dwelling and feeding activities of the infaunal organisms. The development of Skolithos and Cruziana ichnofacies indicates variations in energy of the depositional environment. Skolithos ichnofacies is related to the high-energy storm events and well-oxygenized sandy-shifting substrate in foreshore zone. On the other hand, Cruziana ichnofacies is composed of traces developed under low-energy and unconsolidated soft substrate stable environmental conditions in the shoreface/offshore zone.
References
Al-Bassam KS (2010) Petrology, geochemistry and depositional environment of the Khabour Formation in Ora and Khabour localities, Northern Iraq. Iraqi Bull Geol Min 6:71–94
Al-Hadithi TM (1972, revised by Pesel V 1974). Geological stage report on Sereru area, Northern Iraq. GEOSURV, internal report, no. 655
Aqrawi AAM, Goff J, Horbury AD, Sadooni FN (2010) The petroleum geology of Iraq. Scientific Press, UK
Bann KL, Fielding CR (2004) An integrated ichnological and sedimentological comparison of non-deltaic shoreface and subaqueous delta deposits in Permian reservoir units of Australia. In: McIlroy (ed) The application of ichnology to paleoenvironmental and stratigraphic analysis, vol 228. Geological Society of London, Special Publication, pp 273–310
Bellen RC, van Dunnington HV, Wetzel R, Morton D (1959) Lexique stratigraphique international. Asie, Fasc. 10a, Iraq. Paris, 3
Bromley RG (1992) Bioerosion: eating rocks for fun and profit. In: Maples CG, West RR (eds) Trace fossils (short courses in paleontology, no. 5). Paleontological Society, Knoxville, pp 121–129
Bromley RG (1996) Trace fossils: biology, taphonomy and applications, 2nd edn. Chapman and Hall, London
Buatois LA, Lopez Angriman AO (1992) The ichnology of a submarine braided channel complex: the Whisky Bay formation, Cretaceous of James Ross Island, Antarctica. Palaeogeogr Palaeoclimatol Palaeoecol 94:119–140
Buday T (1980) The regional geology of Iraq. Stratigraphy and Paleogeography, vol 1. Dar Al-Kutub Pub. Univ. of Mosul, Iraq, 445 p
Buday T, Jassim SZ (1987) The regional geology of Iraq: tectonism, magmatism, metamorphism. State establishment of geological survey and mineral investigation, Baghdad
Cornish FG (1986) The trace fossil Diplocraterion: evidence of animal-sediment interactions in Cambrian tidal deposits. Palaios 1:478–491
Crimes TP (1970) Trilobite tracks and other trace fosils from the upper Cambrian of North Wales. Geol J 7:47–68
Crimes TP, Harper JC (eds) (1970) Trace fossils. Geol J Special Issue 3, 547 p
Crimes TP, Goldring R, Homewood P, van Stuijvenberg J, Winkler W (1981) Trace fossil assemblages of deep-sea fan deposits, Gurnigel and Schlieren flysch (Cretaceous-Eocene). Eclogae Geol Helv 74:953–995
d’Orbigny A (1842) Voyage dans l’Amerique méridionale (le Bresil, la Repúblique argentine, le Patagonia, le Repúblique de Chile, le Repúbliique de Bolivie, le Repúblique de Perou) executé pendant les anee 1826, 1827, 1828, 1829, 1830, 1831, 1832 et 1833 . v. 3, pt 4, (Paleontologie): 1–188, París
Ehrenberg K (1944) Erganzende Bemerkungen zu den scinerzeit aus dem Miozan von Burgschleintz beschriebenen Gangkeren and Bauten dekapoder Krebse. Berl Palaeotol Z 23:354–359
Ekdale AA (1988) Pitfalls of paleobathymetric interpretations based on the trace fossil assemblages. Palaios 3:464–472
Ekdale AA, Lewis DW (1991) Trace fossils and paleoenvironmental control of ichnofacies in a late quaternary gravel and loess fan delta complex, New Zealand. Palaeogeogr Palaeoclimatol Palaeoecol 81:253–279
Emmons E (1844) The taconic system: based on observations in New York Massachusetts, Maine, Vermont and Rhode Island. Albany, Caroll and Cook. 68 p
Frey RW (ed) (1975) The study of trace fossils. Springer, Heidelberg
Frey RW, Pemberton SG (1984) Trace fossil facies model. In: Walker RG (ed) Facies models (2nd Ed.). Geoscience Canada, Reprint Series 1, 189–208
Haldeman SS (1840) Supplement to number one of a monograph of the Limniades, or fresh-water univalve shells of North America, containing description of apparently new animals in different classes, and names and characters of the subgenera in Paludina and Anculosa, Philadelphia, 1–3
Hall J (1847) Palaeontology of New York. C. van Benthuysen, Albany 1, 338 p
Hall J (1852) Palaeontology of New York. C. van Benthuysen, Albany 2, 368 p
Heer O (1877) Flora fossils Helvetiae. Die vorweltliche Flora der Schweiz. J. Wurster & Company, Zurich, 182 p
Isa’ac EA (1975) Geology of the Dafaf – Keshan area, Northern Thrust Zone, Iraq. GEOSURV, internal report, no. 659
Jassim SZ, Goff JC (eds) (2006) Geology of Iraq. Dolin, Prague and Moravian Museum, Brno, 340 p
Karim KH (2006) Comparison study between the Khabour and Tanjero formations from North Iraq. Iraqi J Earth Sci 6:1–12
MacEachern JA, Bann KL, Bhattacharya JP, Howell Jr CD (2005) Ichnology of deltas: organism responses to the dynamic interplay of rivers, waves, storms, and tides. In: Giosan L, Bhattacharya JP (eds) River deltas-concepts, models and examples. Society of Economic Paleontologists and Mineralogists Special Publication 83, 49–85
MacEachern JA, Pemberton SG, Gingras MK, Bann KL (2007) The ichnofacies paradigm: a fifty-year retrospective. In: Miller W (ed) Trace fossils: concepts, problems, prospects. Elsevier, Amsterdam, pp 52–77
Nagendra R, Patel SJ, Rakesh D, Reddy AN (2010) Bathymetric Significance of the ichnofossil assemblages of the Kulakkalnattam Sandstone, Ariyalur Area, Cauvery Basin. J Geol Soc India 76:525–532
Nicholson HA (1873) Contributions to the study of the errant Annelids of the older Paleozoic rocks. London, Proc R Soc 21:288–290
Parcha SK, Singh BP, Singh BP (2005) Palaeoecological signifi cance of ichnofossils from the Early Cambrian succession of the Spiti Valley, Tethys Himalaya, India. Curr Sci 88:158–162
Pemberton SG, Frey RW (1982) Trace fossil nomenclature and the Planolites-Palaeophycus dilemma. J Paleontol 56:843–881
Pemberton SG, Frey RW (1984) Ichnology of a storm-influenced shallow marine sequence: cardium Formation (Upper Cretaceous) at Seebe, Alberta. In: Stott DF (ed) Mesozoic of Middle North America. Canadian Society of Petroleum Geology, Memoir 9, pp 281–304
Pemberton SG, van Wagoner JC, Wach GD (1992) Ichnofacies of a wave-dominated shoreline. In: Pemberton SG (ed) Application of ichnology to petroleum exploration. Core Workshop, Society of Economic Palaeontologists and Mineralogists 17, pp 339–382
Savrda CE (1995) Ichnologic applications in paleoceanographic, paleoclimatic and sea-level studies. Palaios 10:565–577
Schieber J (2003) Simple gifts and buried treasures-implications of finding bioturbation and erosion surfaces in black shales. Sediment Rec 1:4–8
Seilacher A (1963) Kaledonischer Unterbau der Irakiden. Neuses Jahrb. Geol. Planet. Abt. Monatshefte, No. 10, 527–542
Seilacher A (1970) Cruziana stratigraphy of non-fossiliferous Palaeozoic sandstones. In: Crimes TP, Harper JC (eds) Trace fossils. Geol J Special Issue 3, pp 447–476
Seilacher A (2007) Trace fossil analysis. Springer, Berlin
Singh RK, Rodriguez-Tovar FJ, Ibotombi S (2008) Trace fossils of the upper Eocene—lower Oligocene transition of the Manipur Indo-Myanmar ranges (Northeast India). Turk J Earth Sci 17:821–834
Sissakian VK (2000) Geological map of Iraq. GEOSURV, Baghdad
Taylor A, Goldring R (1993) Description and analysis of bioturbation and ichnofabric. J Geol Soc Lond 150:141–148
Torell OM (1870) Petrificata Suecana Formationis Cambricae. Lunds Universitet Arsskrift 6:1–14
Uchman A (1999) Ichnology of the Rhenodanubian Flysch (Lower Cretaceous-Eocene) in Austria and Germany. Beringeria 25:65–171
Wetzel R (1950) Khabour Quartzite Formation. In: Bellen RC, van Dunnington HV, Wetzel R, Morton D (1959) Lexique Stratigraphique International. Asie, Fasc. 10a, Iraq. Paris, v.3, 333 p
Zenker JC (ed (1836) Historisch−topographisches Taschenbuch von Jena und seiner Umgebung besonders in naturwissenschaftlicher und medizinischer Beziehung. Wackenhoder, Jena. 338 p
Acknowledgments
The author would like to thank Prof. Ali Al-Jubory, Dr. Mohammad Al-Haj, and Mr. Mahdi Khairy for their help during the field work.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article

Cite this article
Shingaly, W.S. Biogenic-sedimentary structures of the Ordovician-Khabour formation from the Northern Thrust Zone, Kurdistan, Iraq. Arab J Geosci 9, 46 (2016). https://doi.org/10.1007/s12517-015-2126-7
Received:
Accepted:
Published:
DOI: https://doi.org/10.1007/s12517-015-2126-7