
Abstract
Granule neuron precursors (GNPs) proliferate under the influence of Sonic hedgehog (Shh) that is secreted by Purkinje neurons during early postnatal cerebellar development. To investigate microRNA (miRNA) function in this developmental process, we conditionally deleted the Dicer1 gene under the activity of human glial fibrillary acidic protein (hGFAP) promoter. We report that Dicer1-ablated GNPs display decreased proliferation and survival at early postnatal stages and that the proliferation defect of mutant GNPs cannot be rescued by treatment of an Shh agonist in vitro as assayed by 5-bromo-2′-deoxyuridine (BrdU) pulse labeling and Shh target gene expression detection. Further analysis reveals that the expression of distinct cell cycle regulator genes including cell cycle inhibitor, CDKN1a (p21), selectively increases in Dicer1-ablated GNPs. Subsequently, we demonstrate that miR-17-5p exhibits high expression level in the developing cerebellum and that transfection of a synthetic miR-17-5p mimic downregulates p21 protein expression in GNPs and promotes proliferation of GNPs in culture. Therefore, Dicer1 ablation impairs Shh-induced GNP proliferation by disrupting the expression of distinct cell cycle regulator genes that are targets of miR-17∼92 cluster members. This study establishes a molecular link between miRNAs and cell cycle progression in the proliferating GNPs during normal cerebellar development and may facilitate miRNA application in treating medulloblastoma.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Cerebellar granule neuron precursors (GNPs) originate from the rhombic lip (RL) at embryonic stages. In the early postnatal mouse cerebellum, GNPs proliferate rapidly in the external granule layer (EGL) under the influence of Sonic hedgehog (Shh) secreted by the underlying Purkinje neurons in the Purkinje cell layer (PCL), and their differentiated progenies migrate inward along Bergmann glial fibers to form the internal granule layer (IGL) [1, 2]. Purkinje neurons are the main source of Shh in the developing cerebellum and critical in regulating GNP proliferation and differentiation [3, 4]. Shh signaling is initiated by its binding to a receptor complex containing Patched (Ptch) and Smoothened (Smo) and subsequently accomplished by the Gli family of transcription factors. While Gli2 and Gli3 are the major mediators of Shh signaling, Gli1 is a direct downstream target and, therefore, a readout of Shh signaling in the developing cerebellum [5]. N-myc and cyclin D1 are among the downstream transcriptional targets of Shh signaling pathway [6, 7], and the latter promotes cell cycle progression through binding and activating cyclin-dependent kinases (CDKs). Normal cell cycle progression is also modulated by CDK inhibitors such as the CIP/KIP family, which includes CDKN1a/p21WAF1/CIP1 (p21), CDKN1b (p27), and CDKN1c (p57). p21 is considered as a universal inhibitor of cell cycle progression [8], and it exerts its function not only by inhibiting cyclin/CDK complexes, but also by binding and inhibiting proliferating cell nuclear antigen (PCNA), degrading retinoblastoma protein (pRB), and inhibiting cyclin B1 [9].
MicroRNAs (miRNAs), a class of small non-coding RNAs, recognize the 3′-untranslated region (3′-UTR) of their target mRNAs through imperfect base pairing. MiRNAs usually function by either degrading the targets or inhibiting their translation [10–12]. Dicer is an RNase enzyme and indispensable for producing mature miRNAs, and therefore, Dicer knockout mice have been widely used as models to study miRNA functions [13]. For example, Dicer1 has been demonstrated to play crucial roles in different regions and cell types during development of the central nervous system (CNS) [14–21]. In the adult cerebellum, Dicer1 ablation in astroglia [22] and Purkinje neurons [23] leads to degeneration of cerebellar cell types associated with ataxic walking behavior of the mice. In addition, Dicer1 conditional knockout in the early embryonic mid-brain and hindbrain region affects its proper patterning and results in ablation of the entire cerebellum [24, 25]. Our group also reported previously that Dicer1 is essential to proper Notch1 signaling and the development of Bergmann glial cell type in the early postnatal cerebellum [26]. The development of cerebellar GNPs requires normal Dicer1 function, and certain miRNAs have been identified to regulate proliferation, differentiation, and survival of these cells [19, 27–29]. However, the exact mechanisms of miRNAs regulating GNP development, especially the Shh-induced proliferation of these cells, remain elusive.
zIn the present study, we again took advantage of a human glial fibrillary acidic protein (hGFAP)-Cre-mediated Dicer1 conditional knockout mouse to examine the effect of miRNAs in the developing cerebellar GNPs. In this mouse line, GNPs, but not Purkinje neurons, are targeted by the Cre activity [26]. We show that GNP proliferation and survival are reduced in the absence of Dicer1, and this is accompanied by a blockade of Shh signaling in the mutant GNPs. Furthermore, we identified distinct cell cycle regulator genes including cyclin D1, p21, and p57 being upregulated in Dicer1-ablated GNPs, suggesting that these genes are preferentially controlled by miRNAs among other cell cycle regulator genes. We further showed that all the three genes mentioned above are predicted targets of the miR-17∼92 cluster and that manipulating a member of this miRNA cluster, miR-17-5p, can modulate the expression level of p21 and proliferation of GNPs in culture. Therefore, miRNAs, especially the miR-17∼92 cluster members, target cell cycle regulator genes and modulate cell cycle progression in the developing cerebellar GNPs.
Materials and Methods
Mouse Lines and Genotyping
The generation and genotyping of hGFAP-Cre;Dicer1 f/f mice (referred to as mutants) were carried out as previously described [26]. All animal use and studies were approved by the Institutional Animal Care and Use Committee of West China 2nd Hospital, Developmental & Stem Cell Institute, Sichuan University, Chengdu, People’s Republic of China. All procedures were carried out in accordance with the approved guidelines.
Immunohistochemistry, Immunocytochemistry, and Fluorescence Microscopy
Cerebella were collected after intracardiac perfusion with 4 % paraformaldehyde (PFA). The tissues were then sagittally sectioned into 40-μm thickness on a vibratome and postfixed in 4 % PFA overnight at 4 °C. Tissue sections or fixed cell cultures were incubated with monoclonal antibodies against β-III tubulin (TuJ1, mouse IgG, 1:500, Covance), Pax6 (mouse IgG, 1:10, DSHB), 5-bromo-2′-deoxyuridine (BrdU) (rat IgG, 1:200, Accurate), and NeuN (mouse IgG, 1:400, Life Technology) and polyclonal antibodies against Pax6 (rabbit IgG, 1:200, Proteintech), calbindin D-28K (rabbit IgG, 1:500, ImmunoStar), GFAP (rabbit IgG, 1:500, DAKO), cleaved caspase-3 (rabbit IgG, 1:1000, Cell Signaling), β-gal (chicken, 1:1000, Abcam), and Ki67 (rabbit IgG, 1:500, Millipore), followed by appropriate species-specific secondary antibodies (Molecular Probes). 4′,6-Diamidino-2-phenylindole (DAPI) (10 μg/ml, Sigma) was included in the secondary antibody incubations to label nuclei. The tissue sections or cultured cells were then mounted and analyzed by conventional (Nikon Eclipse Ti) fluorescence microscopy.
In Vivo BrdU Pulse Labeling
The procedure for BrdU (Sigma) pulse labeling has been described previously [26]. Briefly, for pulse labeling of proliferating GNPs, BrdU solution was injected peritoneally into postnatal day 5 (P5) mice at 100 mg/kg body weight. Mice were sacrificed 30 min after injection. BrdU staining was performed as described [26].
Total RNA Extraction and qRT-PCR
The procedures for total RNA extraction and quantitative reverse transcriptase PCR (qRT-PCR) analysis have been described previously [26]. Primer sequences for qRT-PCR are shown in Table 1. The relative gene expression levels were normalized to that of the housekeeping gene GAPDH. MiRNA-specific primers were purchased from Guangzhou Ribobio Co., Ltd. (Guangzhou, People’s Republic of China). The relative expression level of individual miRNAs was analyzed according to manufacturer’s instructions and normalized to that of U6 snRNA.
Cerebellar Cell Cultures, GNP Purification, Shh Agonist Treatments, and miRNA Transfection
For acute cerebellar cultures, cerebella were dissected from P5–8 mice. After removal of meninges, the whole tissue was minced using surgical blades, digested with trypsin-EDTA (Life Technology), and dissociated into single cells by pipetting. Cells (1 × 105) were applied to coverslips precoated with poly-d-lysine (50 μg/ml, Sigma) and laminin (20 μg/ml, Life Technology) in a serum-free medium containing DMEM/F12, penicillin/streptomycin, and B27 (Life Technology) [30]. After 4 h of incubation at 37 °C with 5 % CO2, the cells were fixed with 4 % PFA and subjected to immunostaining. GNPs were purified from P5–8 cerebella by Percoll gradient centrifugation according to a modified procedure described previously [30, 31]. The purified GNPs were then plated on a laminin-coated substrate and allowed to recover for 4 h before being harvested with TRIzol® reagent for RNA extraction and subsequent qRT-PCR analysis.
For the GNP proliferation assay, dissociated cells were prepared from P5–8 cerebella and cultured for 22 h in a serum-free medium (DMEM/F12, B27, and penicillin/streptomycin) in the presence of BrdU (10 μg/ml) and either Shh agonist (100 nM, Curis, Cur-0199567) [32] or DMSO (vehicle). The cultures were then fixed with 4 % PFA and subjected to immunostaining by standard protocols. For gene expression analysis, Percoll-purified GNPs were prepared from P5–8 cerebella and cultured for 4 h in the presence of either Shh agonist (100 nM, Curis, Cur-0199567) or DMSO (vehicle) before being harvested in TRIzol® reagent for RNA extraction and subsequent qRT-PCR.
For miRNA transfection, Percoll-purified GNPs were prepared from P5–8 cerebella and allowed to adhere on a laminin-coated substrate for 1 h before being transfected with a synthetic miR-17-5p mimic or scrambled control (synthesized by Guangzhou Ribobio Co., Ltd., People’s Republic of China) using GenoFect™ transfection reagent (Geno Biotech, Beijing, People’s Republic of China) according to the manufacturer’s instructions. We routinely achieve over 60 % of transfection efficiency with this method as revealed by a fluorescence-labeled RNA probe [33]. The cultures were harvested at 16 h after transfection for qRT-PCR and western blot analysis.
Western Blot Analysis
Cerebellar tissues or cultures were harvested either in SDS sample buffer and boiled for 5 min or in TRIzol® reagent, and protein portions from TRIzol® reagent were isolated according to the manufacturer’s instructions (Life Technology). Western blot analysis was performed as previously described [20, 26]. The primary antibody used was anti-Shh (rabbit IgG, 1:750, Proteintech). Anti-β-actin (rabbit IgG, 1:1000, Sigma) was used to normalize sample loadings. Quantification of relative protein expression levels was done by measuring signal intensity of target bands on a LI-COR Odyssey® Infrared Imaging System and normalizing to that of β-actin.
Statistical Analysis
All quantifications were done on sections from at least three animals unless indicated otherwise. For quantifications on cell cultures and western blots, data were collected from at least three biological replicates. The data were presented as means ± standard deviation. Statistical analysis was performed in Microsoft Excel using Student’s t test.
Results
Dicer1 Ablation Induces Loosely Packed Granule Neurons in the IGL of the Postnatal Cerebellum
To investigate miRNA function in the developing cerebellum, we previously generated a mouse line with conditional Dicer1 gene knockout under the activity of hGFAP-Cre transgene. The hGFAP-Cre-mediated recombination is first active in the RL at embryonic day 13.5 (E13.5) and predominates in GNPs of the EGL and subsequent granule neurons of the IGL during cerebellar development [26]. The hGFAP-Cre;Dicer1 f/f mice (referred to as mutants in this study) can survive until 3 weeks after birth but exhibit a walking deficit and a smaller cerebellum [26]. To examine granule neurons in the postnatal cerebellum, we performed immunostaining with an antibody against NeuN, a mature neuronal marker. We found that differentiated granule neurons revealed by anti-NeuN antibody were positioned correctly in the IGL of mutant cerebella at P16, yet their cell bodies were loosely packed in contrast to compact sheets of granule neurons in controls with a clear boundary between the molecular layer (ML) and IGL (Fig. 1a, b). In addition, we often found “holes” in the loosely packed P16 mutant IGL by anti-NeuN staining (Fig. 1c). These NeuN-negative holes were likely due to the absence of cells rather than less neuronal maturation, since they were devoid of cell bodies as revealed by negative DAPI staining, but filled with GFAP+ astrocytic processes (Fig. 1c, c’, c”, arrows).

Loosely packed IGL in the Dicer1-ablated cerebellum. The structural phenotype of cerebellar IGL was examined by NeuN immunostaining on control (ctrl) (a) and Dicer1-ablated (mut (mutant)) (b) cerebellar sections of P16 mice. c The enlarged region (box in b) shows that NeuN− “holes” in the mutant cerebellar IGL are devoid of DAPI staining but filled with GFAP+ processes (arrows). Scale bars 100 μm
Dicer1 Ablation Results in Reduced GNP Proliferation and Survival in the Developing Cerebellum
The reduction of cerebellar size prompted us to examine cell proliferation in the mutant cerebellum during development, especially in the GNPs where Cre-mediated Dicer1 deletion strongly occurs. By immunohistochemistry, we found that the intensity of anti-Ki67 staining in the mutant EGL appeared normal during embryonic stages (data not shown) but started to decrease at P1 in the anterior lobes (Fig. 2a, arrows) suggesting a decrease of GNP proliferation. This is unlikely due to a defect in tangential migration of Dicer1-ablated GNPs, because the thickness of the mutant EGL in the anterior lobes is comparable to that of controls as revealed by DAPI staining (Fig. 2a, insets). Later during development, the Ki67+ EGL was much less developed in the mutant cerebellum at P5, especially in the anterior lobes (Fig. 2a, arrows), accompanied by shorter lobes and malformation of folia (Fig. 2a). However, the Ki67+ cells in the mutant parenchyma showed a higher density than those in controls (Fig. 2a), and we think that this was likely due to a “crowding” effect rather than increased proliferation. To further confirm the proliferation defect of Dicer1-ablated cerebella, we also quantified the percentage of Ki67+ cells in acute P5 cerebellar primary cultures. Consistently, we found a significant decrease in the percentage of Ki67+ cells among total, as well as Pax6+, cells in mutants confirming a reduction of proliferation in mutant GNPs (Fig. 2b). To check if Dicer1 ablation affects the cell cycle dynamics of proliferating GNPs, we performed a 30-min BrdU pulse labeling experiment on P5 mice to exclusively label the proliferating GNPs in S-phase. The cell proliferation rate was analyzed on GNPs of the central lobes where Ki67 expression was not much affected in mutants. Our result showed that the percentage of cells in S-phase (BrdU+) among Ki67+ GNPs in the EGL significantly decreased in mutants especially in the anterior lobes (L1, L2, and L4) (Fig. 2c), indicating a defected cell cycle parameter of proliferating GNPs upon Dicer1 deletion. Therefore, our conclusion is that Dicer1 ablation leads to a reduced GNP proliferation at early postnatal stages.

Dicer1-ablated GNPs exhibit reduced proliferation and survival in the developing cerebellum. (a) Immunostaining on P1 and P5 cerebellar parasagittal sections showed reduced Ki67 expression in the mutant (mut) EGL compared with the control (ctrl), especially in the anterior lobes (arrows). Higher-magnification images showed a normal cell number (DAPI) in the anterior EGL of P1 mutant (insets). Scale bars 100 μm. b The percentage of Ki67+ cells among both total cells (DAPI) and GNPs (Pax6+) in acutely isolated P6 cerebellar cultures was also significantly reduced in mutants compared with controls (n = 3 for both mutants and controls). c A 30-min BrdU pulse labeling analysis showed a significant reduction in the percentage of BrdU+ among Ki67+ cells in the anterior lobes (L1, L2, and L4) in P6 mutants compared with controls (n = 3 for both mutants and controls). The overlay image showed a representative double-immunostaining of Ki67 and BrdU (inset). d Immunostaining with an anti-cleaved caspase-3 antibody showed increased apoptotic cells in P1 mutant (mut) cerebellar sections compared with controls (ctrl), especially in the EGL. The apoptotic cells in the EGL were TuJ1− and Pax6+ (insets). e The numbers of apoptotic cells in the EGL and in total area per section were significantly higher in the mutant cerebellum at both P1 and P5 (n = 3 for both mutant and control). DAPI was used to label nuclei. EGL external granule layer. *P < 0.05; **P < 0.01 by Student’s t test
Next, we examined whether apoptosis contributed to the reduction in cerebellar size in mutants. Indeed, staining with an anti-cleaved caspase-3 antibody showed a significant increase of apoptotic cells in the P1 and P5 mutant cerebella compared with controls (Figs. 2d and 3e), especially in the EGL where GNPs (Pax6+ and TuJ1−) reside (Fig. 2d, insets). Therefore, Dicer1 ablation also leads to increased apoptosis most strikingly in GNPs of the early postnatal cerebellum.

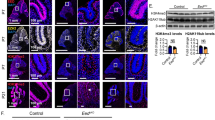
Mispositioned Purkinje neurons and yet sufficient Shh expression in the Dicer1-ablated cerebellum. a Calbindin-expressing Purkinje neurons exhibit an aligning defect in the mutant (mut) cerebellum at P5 resulting in a multicell layer PCL (arrow) in contrast to a single-cell layer PCL in the control (ctrl). At P16, this aligning defect remains with ectopic Purkinje neurons occasionally seen in the IGL and WM (arrowhead). Scale bar 40 μm. b Gene expression levels of Dicer1 and Shh in P4 cerebellar tissues (n = 5 for both mutant and control) were analyzed by qRT-PCR. c Protein levels of Shh were also compared by western blot analysis between these P4 cerebellar tissues, and levels were normalized by β-actin. PCL Purkinje cell layer, n.s. no significance; **P < 0.01 by Student’s t test
Dicer1 Ablation Induces Misaligned Purkinje Neuron Without Altering Shh Expression in the Developing Cerebellum
During normal development, calbindin-expressing Purkinje neurons align as a single-cell layer underneath the EGL starting from P3 and secrete Shh to regulate development of the adjacent proliferative GNPs. We therefore set out to examine the behavior of Purkinje neurons, the source of secreted Shh [3, 4, 34], in mutants. Interestingly, immunostaining showed that Purkinje neurons were mispositioned at P5 in mutant cerebella where they appeared as a multilayer of cells instead of a single-cell layer as seen in controls (Fig. 3a, arrow). This mispositioning phenotype of Purkinje neurons disrupted close association between these cells and GNPs and could potentially alter the concentration of Shh protein received by the latter cells in the EGL. At P16, most Purkinje neurons in mutants aligned in the PCL with some misplaced probably due to their inability to reach the PCL (Fig. 3a, arrowhead). Note that Purkinje neurons are not targeted by hGFAP-Cre [26] and, therefore, genetically normal in the mutant mouse and that the observed phenotypic defect of these cells is likely due to non-cell-autonomous mechanisms.
Since Shh plays a critical role in regulating GNP proliferation during cerebellar development [3, 4, 35], we examined whether Shh signaling was altered in mutant cerebella. The gene expression level of Shh signaling components was compared between mutant and control at P4 by qRT-PCR and western blot analysis. We first confirmed a significant reduction of Dicer1 expression in mutant cerebellar tissues by qRT-PCR (Fig. 3b). At the secreted factor level of the signaling pathway, we observed a similar level of both Shh mRNA and protein at P4 between mutant and control (Figs. 3b and 4c), suggesting that a sufficient amount of this factor is produced in the mutant cerebella. However, the misalignment of Purkinje neurons in mutant cerebella raised the question whether Shh protein is delivered efficiently to the proliferating GNPs that reside on top.

Dicer1-ablated GNPs fail to increase proliferation in response to Shh agonist treatment in culture. a A 4-h Shh agonist (100 nM) treatment was performed in Percoll gradient-purified GNPs, and gene expression response was analyzed by qRT-PCR and compared between mutants and controls (n = 3 for both mutant and control). Furthermore, the acute cerebellar cultures from P6 control and mutant mice were subjected to a 22-h treatment of Shh agonist (100 nM) or vehicle control in the presence of BrdU. Immunostainings with specific antibodies were performed to characterize mutant (b–i), as well as control, cerebellar cultures. DAPI (blue) was used to label nuclei. GNP proliferation (j), differentiation (k), and apoptosis (l) upon this Shh agonist treatment were examined and quantified by immunostainings. *P < 0.05; **P < 0.01 by Student’s t test (Color figure online)
Dicer1-Ablated GNPs Fail to Respond to Shh Agonist Treatment in Vitro
The hGFAP-Cre targets multiple cerebellar cell types including GNPs and Bergmann glia [26], and thus, it is hard to distinguish the specificity of Dicer1-deletion-mediated effect on GNPs, i.e., cell-autonomous vs. non-cell-autonomous effect. Therefore, we mainly applied purified GNPs in the following experiments to characterize miRNA function in these cells in cultures where cell-cell interaction is minimized, which is likely to reveal cell-autonomous mechansims. To test mutant GNP’s response to Shh directly, we subjected these cells to treatment of a Shh agonist [32] in vitro. GNPs respond to the Shh agonist by increasing target gene expression and cell proliferation in culture [3]. We isolated these cells from early postnatal cerebella by Percoll gradient centrifugation and exposed them to the Shh agonist in vitro. First, gene expression was measured by qRT-PCR after a 4-h exposure of the Shh agonist (100 nM). As expected, the expression of Gli1, Gli2, and Patched1 in control GNPs increased by this treatment with both Gli1 and Patched1 being transcriptional targets of Shh signaling [5, 36] (Fig. 4a). However, expression of these genes in mutant GNPs was not increased upon Shh agonist incubation, suggesting a blockade of Shh signaling pathway in the mutant GNPs (Fig. 4a). Reduced Dicer1 level confirms the mutant genotype, and similar Pax6 expression indicates an equivalent GNP purification efficiency between mutant and control samples (Fig. 4a). Second, GNP proliferation was assessed by their ability to incorporate BrdU in the presence of Shh agonist in a 22-h assay. GNPs were identified by their anti-Pax6 immunoreactivity, while anti-BrdU antibody marked proliferating cells. In mutant cerebellar cultures, we confirmed that Cre-mediated recombination (as revealed by β-gal positivity) occurs in more than 50 % of Pax6+ GNPs (Fig. 4b) and TuJ1+ neurons (Fig. 4c), most of which are granule neurons as they are also Pax6+ (Fig. 4d). We also confirmed BrdU labels dividing cells in these cultures as indicated by a proliferation marker Ki67 (Fig. 4e) and that the majority of BrdU+ cells are Pax6+ GNPs (Fig. 4f). Furthermore, apoptotic cells that are positive for cleaved caspase 3 do not show colocalization with any of these abovementioned markers (Fig. 4g–i), which ensures the specificity of our analysis on survived mutant GNPs, but not the ones undergoing apoptosis. Upon Shh signaling activation, our results showed that the percentage of BrdU+ cells among Pax6+ GNPs significantly increased in control but not in mutant (Fig. 4j), and the percentage of TuJ1+ cells among Pax6+ GNPs decreased in control but not in mutant (Fig. 4k). These results indicated that, while Shh agonist treatment promotes cell proliferation and inhibits neuronal differentiation of control GNPs, its effect is blocked in Dicer1-ablated GNPs suggesting a critical function of miRNAs in mediating Shh signaling pathway. In addition, this percentage of proliferating GNPs was higher under untreated conditions (vehicle) in control compared with that in mutant (Fig. 4j), which is consistent with our in vivo data of decreased GNP proliferation in mutant mice (Fig. 3). In the same factor treatment experiment, we also checked the Shh agonist effect on GNP survival and found that increased percentage of apoptotic GNPs in mutant cerebellar cultures as identified by immunopositivity of both caspase 3 and Pax6 could not be rescued by a 22-h Shh agonist treatment (Fig. 4i). Furthermore, this result also indicated that cell apoptosis does not contribute to the blockade of Shh signaling in Dicer1-ablated GNPs. In conclusion, these combined in vitro results demonstrated that GNPs fail to respond to Shh signaling activation and do not show promoted proliferation in the absence of miRNAs.
Dicer1-Ablated GNPs Display Increased Expression of Distinct Cell Cycle Regulator Genes
Decreased proliferation rate in vivo and blocked Shh agonist-stimulated proliferation of mutant GNPs in vitro prompted us to examine their cell cycle parameters in more detail. Percoll gradient-purified GNPs were obtained and allowed to recover in a serum-free medium for 4 h before RNAs were extracted and analyzed by qRT-PCR. We first showed that Pax6 expression is similar between mutant and control, which indicated an equivalent number of GNPs in the samples (Fig. 5a). Instead of doing an extensive gene profile screening to identify responsible targets for the mutant GNP defect, we decided to first concentrate on genes associated with cell cycle. Among cell cycle-related genes examined, only p21, p57, and cyclin D1 showed a significant increase in mutant GNPs compared with control GNPs (Fig. 5a), suggesting their regulation by miRNAs in the developing GNPs. Furthermore, the expression of Gli1 and Gli2, two critical components of Shh signaling pathway, showed no difference in purified GNPs (Fig. 5a), indicating that their expression is not affected by Dicer1 ablation.

Dicer1-ablated cerebellar GNPs show elevated expression level of distinct cell cycle-related genes that are predicted targets of the miR-17∼92 cluster. a Expression levels of cell cycle-related genes in Percoll-purified GNPs (n = 3 for both mutant and control) were analyzed by qRT-PCR. b Diagram showing miRNAs from the miR-17~92 cluster target the 3′-UTRs of distinct cell cycle-related genes. c Genomic arrangement (not to scale) of the miRNA clusters that have been shown to target p21 mRNA. Multiple miRNAs (labeled by filled shape) from each cluster contain the same seed sequence that targets p21 mRNA. d Expression levels of selected mature miRNAs that are predicted to target p21 were analyzed by qRT-PCR in P4 cerebellar tissues (n = 5 for both mutant and control). e Expression levels of selected mature miRNAs that are predicted to target p21 were also analyzed by qRT-PCR in P4-purified cerebellar GNPs (n = 3 for both mutant and control). *P < 0.05; **P < 0.01 by Student’s t test. N.D. not detected
MiR-17-5p-Mediated Regulation of p21 Expression and Cell Proliferation in GNPs
Among the cell cycle-related genes that we examined, p21 is most drastically upregulated upon Dicer1 ablation (Fig. 5a). To gain molecular insights as to how p21 expression is regulated by miRNAs, we set out to identify potential miRNA candidates that target the p21 mRNA in the developing GNPs. The miRNA cluster miR-17∼92 has been involved in cerebellar development and medulloblastoma formation [29, 37–39]. Interestingly, three genes that show enhanced expression in Dicer1-ablated GNPs all contain conserved binding sites of miRNA members of the miR-17∼92 cluster and therefore predicted targets of these miRNAs (Fig. 5b). Notably, miR-17-5p targets two of the three upregulated genes including p21, suggesting a particularly important function of this miRNA in mediating GNP proliferation (Fig. 5b). In fact, miR-17-5p shares the same seed sequence with members of the same and several other miRNA clusters (Fig. 5c), indicating that these individual miRNAs may play redundant roles during cerebellar development. Since several previous studies have described p21 as one of the major cell cycle inhibitors targeted by miRNAs in embryonic stem cells (ESC) and cancer [40–43], we took a different approach to investigate further, that is, to examine the expression of these known miRNAs to see which, if any, of these known miRNA regulators are present in the developing cerebella and, more importantly, to see which one(s) of them is knocked down in our Dicer1-ablated mice. We picked one miRNA from each individual clusters that have been shown to target p21 in other systems (Fig. 5c) and examined their expression, assuming that members of a miRNA cluster are spliced and co-expressed from one polycistronic RNA molecule. Again, we performed a qRT-PCR analysis on P7 cerebellar tissues and found that miR-17-5p, miR-106b-5p, and miR-106a-5p are present in the P7 cerebellum in a high to low order in terms of their expression level as judged by their delta Ct values in the amplification curve (data not shown). In addition, the expression of these miRNAs is dramatically reduced in mutant (Fig. 5d), suggesting their possible contribution to the defected proliferation phenotype of mutant GNPs. MiR-294-3p and miR-302 showed low and undetectable expression, respectively, in cerebellar tissues, and no significant downregulation of these miRNAs was observed in mutant compared with control (Fig. 5d). Furthermore, we also confirmed the expression of miR-17-5p and miR-106a-5p in purified GNPs and their drastic reduction in expression in mutant GNPs (Fig. 5e).
Since miR-17-5p is the most highly expressed miRNA that is predicted to target p21 in the developing cerebellum, we next asked if changing the cellular level of miR-17-5p could regulate p21 expression. Ideally, the most relevant experiment would be to overexpress miR-17-5p in mutant GNPs and see if this could rescue their phenotype. However, the extremely low yield of mutant GNPs after Percoll purification makes them very hard to sustain over our transfection procedure. Therefore, to test proof of principle, we transfected control GNPs with a synthetic miR-17-5p mimic to increase the miR-17-5p level in these cells and examined their behavior. Indeed, we were able to detect a significant decrease of p21 protein level by western blot analysis upon transfection with the miR-17-5p mimic in control GNPs (Fig. 6a). In order to check if reduced p21 protein level upon transfection is associated with a possible increase in cell proliferation, we performed a BrdU pulse labeling in the same experiment and found a significant increase of BrdU+ cells among the Pax6+ GNPs upon transfection with the miR-17-5p mimic compared with control transfection (Fig. 6b). In the meantime, we also examined the expression of proliferation genes such as Ki67 and PCNA by qRT-PCR and detected significant increases in the expression of these genes upon transfection with the miR-17-5p mimic (Fig. 6c). Therefore, transfection with the miR-17-5p mimic can regulate cellular level of p21 protein and promote GNP proliferation in culture. Similarly to our result, overexpression of the entire miR-17∼92 cluster has been shown to promote proliferation of isolated GNPs in culture [38, 39]. In addition, in consistence with our observation, miR-17-5p binding sites on the 3′-UTR of p21 were predicted and validated by luciferase assay in both human [44–46] and mouse [40, 41, 44].

MiR-17-5p regulates p21 expression and cell proliferation of GNPs in vitro. a Percoll-purified cerebellar GNPs from P7 wild-type mice were transfected with a synthetic miR-17-5p mimic or control. Western blot analysis was performed to detect changes of P21 protein level, and the result was quantified by fluorescent intensity of the protein bands after being normalized to that of β-actin. b Proliferations of wild-type cerebellar GNPs were analyzed by BrdU pulse labeling upon miR-17-5p mimic transfection. c Gene expression of proliferation markers was also examined by qRT-PCR upon miR-17-5p mimic transfection. The results are from at least three independent experiments. *P < 0.05 by Student’s t test
Discussion
In this study, we conditionally deleted the Dicer1 gene and examined the effect of loss of miRNAs in GNPs during cerebellar development. Our data show that the smaller cerebellum seen in Dicer1 mutants results from a series of cellular defects including decreased proliferation and increased apoptosis in GNPs at early postnatal stages, which is accompanied by the formation of a loosely packed IGL later during development. The Dicer1-ablated GNPs fail to show enhanced proliferation upon Shh agonist treatment in vitro and exhibit impaired Shh response as assayed by expression of Shh downstream target genes. Furthermore, we report that, along with other cell cycle regulator genes, p21 is significantly upregulated in Dicer1-ablated GNPs and may be a major target of miRNAs in regulation of GNP proliferation. Finally, we provide evidence that the highly expressed miR-17∼92 cluster may regulate several cell cycle regulators during GNP cell proliferation in the early postnatal cerebellum. Our report is the first to link the miR-17∼92 cluster to cell cycle regulator genes in the developing cerebellar GNPs and highlight that miRNAs are indispensable to normal development of these cells, in addition to their roles in the formation of medulloblastoma.
It has been reported that miRNAs target multiple components of the Shh signaling pathway [47]. Ferretti et al. screened miRNAs that are differentially expressed between human MB tissues with high and low Shh signaling levels and identified miR-125b, miR-326, and miR-324-5p as acting as suppressors of Shh pathway components such as Smo and Gli1. The authors further demonstrated that the expression of these miRNAs increased along with granule neuron in vitro differentiation and promoted granule neuron maturation and inhibited proliferation of their precursors when overexpressed [47]. Consistently, our data also underline the importance of miRNA involvement in modulating GNP proliferation and Shh signaling via the Dicer1-ablated condition. Furthermore, we particularly value the Dicer knockout model as a nice screening system for identifying miRNA-regulated gene expression under physiological conditions, because genes that are normally suppressed by miRNAs will likely go up in their expression in the absence of Dicer1. We think that cell cycle regulator genes p21, p57, and cyclin D1 identified in our analysis are genes within that category and belong to miRNA-sensitive (MRS) target genes as we previously defined [20]. However, this screening method is not exclusive and we cannot rule out that other cell cycle-related genes are also regulated by miRNAs. In fact, p27 is reportedly regulated by miR-221 and miR-222 in cancer cells [48–50], and components of the Shh signaling pathway such as Smo and Gli1 are also targets of miRNAs in MBs as mentioned above. However, our screening did not reveal those genes. One possible explanation is that functionally opposing miRNAs exist in vivo and that ablation of both positive and negative types of regulatory miRNAs induces no net change of target gene expression. Alternatively, miRNAs identified in disease models may not be relevant under physiological conditions. In addition, the extensiveness of regulation by miRNAs vs. genetic control in the expression of a gene may determine the positivity of detection [20]. The fact that p21 showed most robust upregulation of expression in the absence of Dicer1 indicates that it is heavily regulated by miRNAs as a good example of MRS target genes in the proliferating GNPs and may serve as a potential miRNA-facilitated drug target for treating MB.
MiRNA-regulated expression of p21 has been described in other biological systems [40–43]. Although Dicer1 ablation in cerebellar GNPs has been described recently [19, 21, 33, 51], we are the first to show that distinct cell cycle regulator genes including p21 are preferentially upregulated in the absence of Dicer1 (Fig. 5). Furthermore, we provide evidence that miR-17-5p from the miR-17∼92 cluster regulates p21 expression in developing GNPs and promotes GNP proliferation when overexpressed (Fig. 6). Other miRNAs such as miR-106b in the miR-17-5p family with the same seed sequence are expected to have similar function in GNP proliferation as in other biological system [52]. Previous reports indicate that miR-17∼92 cluster is a transcriptional target of N-myc [38], the expression of which is promoted by Shh signaling [6]. In addition, miR-17∼92 cluster has been shown to have an upregulated expression level and to be involved in the aberrant proliferation in MB [38, 39], but its genetic targets have not been identified in the cerebellum thus far [53]. Here, we show that miR-17-5p targets p21, a universal cell cycle inhibitor, to modulate its expression. Cyclin D1 and p57 are also predicted targets of the miR-17 family and show higher expression levels in the isolated GNPs of mutant. Therefore, miR-17 family members function to modulate multiple targets including both positive (cyclin D1) and negative (p21 and p57) regulators to facilitate normal cell cycle control in the developing cerebellar GNPs. However, we do not exclude the possible indirect mechanisms of miRNA-mediated regulation of GNP proliferation. In fact, both miR-17-5p and miR-106b have been shown to regulate cell proliferation by interacting with other signaling pathways [54–56]. Furthermore, miR-22 was recently reported to regulate cell cycle length of proliferating GNPs by targeting an N-myc-interacting transcription factor Max [27]. These authors further provide evidence that miR-22 regulates Max expression level in the GNPs and promoter activity of N-myc/Max downstream genes [27]. Since miR-17∼92 cluster is a transcriptional target of N-myc [38], miR-22 may act upstream of miR-17∼92 cluster members. Therefore, these miRNAs function at different levels of the signaling pathways to coordinate regulation of cell cycle progression of the developing cerebellar GNPs. Interestingly, this study also identified miR-17, miR-19b, and miR-18a of the miR-17∼92 cluster as downregulated miRNAs in the BMP2-treated GNPs when Shh signaling is antagonized [27]. This is consistent with our observation in the current report that miRNAs including miR-17-5p are critical components of a functional Shh response in the GNPs. The downregulated miRNAs in BMP2 treatment were not characterized further in terms of their potential gene targets in the GNPs [27]; however, it would be interesting to check if cell cycle regulator genes such as p21 are upregulated in their experimental condition similarly to our results in Dicer1-ablated GNPs.
Increased apoptosis in Dicer1-deleted GNPs mimics observations reported in other regions of the CNS. For example, increased cell death of neural progenitors has been observed in Dicer1-deleted developing forebrains, and late progenitors are more sensitive to Dicer1 ablation than the early ones [15]. These authors speculated that miRNA functions are more profound when cells are at a stage of altering their state, i.e., from proliferating progenitors to postmitotic progenies. Similarly in our study, Cre-mediated recombination occurs in the RL as early as E13.5 and the embryonic EGL during subsequent stages, yet we did not observe drastic apoptosis until after birth when GNPs initiate their differentiation program and inward migration. Another possible reason for the absence of apoptotic GNPs at embryonic stages in our Dicer1 ablation mouse model is that, as we observed in our previous report, miRNA depletion does not fully occur until perinatal stages during development even though hGFAP promoter activity can be detected in the cerebral cortex as early as E13.5 [20]. Consistent with this notion, Nestin (an earlier expressed gene than hGFAP)-Cre-mediated Dicer1 ablation is able to induce GNP apoptosis in the RL of the embryonic cerebellum as early as E14.5 [21]. A contradictory finding on Dicer1-ablated GNP apoptosis is from a recent study where Atoh1 (an RL-specific gene)-Cre-mediated Dicer1 ablation does not induce GNP apoptosis at all as determined by TUNEL staining [19]. Notably, while Nestin and hGFAP promoters are active in multiple cerebellar cell types including GNPs, Atoh1 promoter is more specific to granule neuron lineage [57]. This raises the question whether the observed GNP apoptosis in our Dicer1 ablation mouse model is due to a cell-autonomous effect, although we can also detect increased apoptosis in Dicer1-ablated cerebellar cultures where cell-cell interaction is minimized (Fig. 2). A more recent report demonstrated a substantial GNP apoptosis in the Atoh1 (Math1)-Cre-mediated Dicer1-deletion mouse [51]. These authors further provided evidence that Dicer is essential for resolving replication-associated DNA damage in GNPs and that apoptosis seen with Dicer deficiency in these cells can be partially rescued by deletion of p53 [51]. Interestingly, we also observed increased gene expression of p53 in purified mutant GNPs in our study (data not shown). Thus, further investigation is needed to clarify these controversial findings. Nevertheless, miRNAs are essential to cell survival since knockdown of a single miRNA (miR-29) induces massive cell death in different regions of the CNS including cerebellum [28].
Conclusions or Summary
In conclusion, our study provides molecular insights into miRNA-mediated regulation of cerebellar GNP proliferation and identifies cell cycle regulator genes such as p21 as important miRNA-regulated targets for the cell cycle progression of these cells. Although miRNAs have been implicated in MB and GNP proliferation, our loss of function system provides unique evidence that miRNAs are indispensable to Shh-stimulated GNP proliferation during normal cerebellar development. In addition, understanding gene targets of functional miRNAs in the GNPs will mechanistically facilitate therapeutic use of these small RNA molecules [58] in treating MB.
References
Sotelo C. Cellular and genetic regulation of the development of the cerebellar system. Prog Neurobiol. 2004;72(5):295–339.
Wang VY, Zoghbi HY. Genetic regulation of cerebellar development. Nat Rev Neurosci. 2001;2(7):484–91.
Dahmane N, Ruiz i Altaba A. Sonic hedgehog regulates the growth and patterning of the cerebellum. Development. 1999;126(14):3089–100.
Wechsler-Reya RJ, Scott MP. Control of neuronal precursor proliferation in the cerebellum by Sonic Hedgehog. Neuron. 1999;22(1):103–14.
Corrales JD, Rocco GL, Blaess S, Guo Q, Joyner AL. Spatial pattern of sonic hedgehog signaling through Gli genes during cerebellum development. Development. 2004;131(22):5581–90.
Kenney AM, Cole MD, Rowitch DH. Nmyc upregulation by sonic hedgehog signaling promotes proliferation in developing cerebellar granule neuron precursors. Development. 2003;130(1):15–28.
Oliver TG, Grasfeder LL, Carroll AL, Kaiser C, Gillingham CL, Lin SM, et al. Transcriptional profiling of the Sonic hedgehog response: a critical role for N-myc in proliferation of neuronal precursors. Proc Natl Acad Sci U S A. 2003;100(12):7331–6.
Xiong Y, Hannon GJ, Zhang H, Casso D, Kobayashi R, Beach D. p21 is a universal inhibitor of cyclin kinases. Nature. 1993;366(6456):701–4.
Mirzayans R, Andrais B, Scott A, Murray D. New insights into p53 signaling and cancer cell response to DNA damage: implications for cancer therapy. J Biomed Biotechnol. 2012;2012:170325.
Bartel DP. MicroRNAs: genomics, biogenesis, mechanism, and function. Cell. 2004;116(2):281–97.
Stefani G, Slack FJ. Small non-coding RNAs in animal development. Nat Rev Mol Cell Biol. 2008;9(3):219–30.
Cao X, Yeo G, Muotri AR, Kuwabara T, Gage FH. Noncoding RNAs in the mammalian central nervous system. Annu Rev Neurosci. 2006;29:77–103.
Bernstein E, Kim SY, Carmell MA, Murchison EP, Alcorn H, Li MZ, et al. Dicer is essential for mouse development. Nat Genet. 2003;35(3):215–7.
Davis TH, Cuellar TL, Koch SM, Barker AJ, Harfe BD, McManus MT, et al. Conditional loss of Dicer disrupts cellular and tissue morphogenesis in the cortex and hippocampus. J Neurosci. 2008;28(17):4322–30.
De Pietri Tonelli D, Pulvers JN, Haffner C, Murchison EP, Hannon GJ, Huttner WB. miRNAs are essential for survival and differentiation of newborn neurons but not for expansion of neural progenitors during early neurogenesis in the mouse embryonic neocortex. Development. 2008;135(23):3911–21.
Dugas JC, Cuellar TL, Scholze A, Ason B, Ibrahim A, Emery B, et al. Dicer1 and miR-219 are required for normal oligodendrocyte differentiation and myelination. Neuron. 2010;65(5):597–611.
Zhao X, He X, Han X, Yu Y, Ye F, Chen Y, et al. MicroRNA-mediated control of oligodendrocyte differentiation. Neuron. 2010;65(5):612–26.
Zheng K, Li H, Zhu Y, Zhu Q, Qiu M. MicroRNAs are essential for the developmental switch from neurogenesis to gliogenesis in the developing spinal cord. J Neurosci. 2010;30(24):8245–50.
Constantin L, Wainwright BJ. MicroRNAs promote granule cell expansion in the cerebellum through Gli2. Cerebellum. 2015;14(6):688–98.
Zhang C, Ge X, Liu Q, Jiang M, Li MW, Li H. MicroRNA-mediated non-cell-autonomous regulation of cortical radial glial transformation revealed by a Dicer1 knockout mouse model. Glia. 2015;63(5):860–76.
Zindy F, Lee Y, Kawauchi D, Ayrault O, Merzoug LB, Li Y, et al. Dicer is required for normal cerebellar development and to restrain medulloblastoma formation. PLoS One. 2015;10(6):e0129642 .PMCID: 4474721
Tao J, Wu H, Lin Q, Wei W, Lu XH, Cantle JP, et al. Deletion of astroglial Dicer causes non-cell-autonomous neuronal dysfunction and degeneration. J Neurosci. 2011;31(22):8306–19.
Schaefer A, O’Carroll D, Tan CL, Hillman D, Sugimori M, Llinas R, et al. Cerebellar neurodegeneration in the absence of microRNAs. J Exp Med. 2007;204(7):1553–8.
Huang T, Liu Y, Huang M, Zhao X, Cheng L. Wnt1-cre-mediated conditional loss of dicer results in malformation of the midbrain and cerebellum and failure of neural crest and dopaminergic differentiation in mice. J Mol Cell Biol. 2010;2(3):152–63.
Soukup GA, Fritzsch B, Pierce ML, Weston MD, Jahan I, McManus MT, et al. Residual microRNA expression dictates the extent of inner ear development in conditional Dicer knockout mice. Dev Biol. 2009;328(2):328–41.
Kuang Y, Liu Q, Shu X, Zhang C, Huang N, Li J, et al. Dicer1 and MiR-9 are required for proper Notch1 signaling and the Bergmann glial phenotype in the developing mouse cerebellum. Glia. 2012;60(11):1734–46.
Berenguer J, Herrera A, Vuolo L, Torroba B, Llorens F, Sumoy L, et al. MicroRNA 22 regulates cell cycle length in cerebellar granular neuron precursors. Mol Cell Biol. 2013;33(14):2706–17 .PMCID: 3700129
Roshan R, Shridhar S, Sarangdhar MA, Banik A, Chawla M, Garg M, et al. Brain-specific knockdown of miR-29 results in neuronal cell death and ataxia in mice. RNA. 2014;20(8):1287–97 .PMCID: 4105753
Zindy F, Kawauchi D, Lee Y, Ayrault O, Ben Merzoug L, McKinnon PJ, et al. Role of the miR-17 approximately 92 cluster family in cerebellar and medulloblastoma development. Biol Open. 2014;3(7):597–605 .PMCID: 4154296
Li H, Berlin Y, Hart RP, Grumet M. Microtubules are critical for radial glial morphology: possible regulation by MAPs and MARKs. Glia. 2003;44(1):37–46.
Hatten ME. Neuronal regulation of astroglial morphology and proliferation in vitro. J Cell Biol. 1985;100(2):384–96.
Li H, Han YR, Bi C, Davila J, Goff LA, Thompson K, et al. Functional differentiation of a clone resembling embryonic cortical interneuron progenitors. Dev Neurobiol. 2008;68(14):1549–64.
Swahari V, Nakamura A, Baran-Gale J, Garcia I, Crowther AJ, Sons R, et al. Essential function of dicer in resolving DNA damage in the rapidly dividing cells of the developing and malignant cerebellum. Cell Rep. 2016;14(2):216–24 .PMCID: 4716786
Wallace VA. Purkinje-cell-derived Sonic hedgehog regulates granule neuron precursor cell proliferation in the developing mouse cerebellum. Curr Biol. 1999;9(8):445–8.
Corrales JD, Blaess S, Mahoney EM, Joyner AL. The level of sonic hedgehog signaling regulates the complexity of cerebellar foliation. Development. 2006;133(9):1811–21.
Goodrich LV, Johnson RL, Milenkovic L, McMahon JA, Scott MP. Conservation of the hedgehog/patched signaling pathway from flies to mice: induction of a mouse patched gene by Hedgehog. Genes Dev. 1996;10(3):301–12.
Murphy BL, Obad S, Bihannic L, Ayrault O, Zindy F, Kauppinen S, et al. Silencing of the miR-17∼92 cluster family inhibits medulloblastoma progression. Cancer Res. 2013;73(23):7068–78 .PMCID: 3857104
Northcott PA, Fernandez LA, Hagan JP, Ellison DW, Grajkowska W, Gillespie Y, et al. The miR-17/92 polycistron is up-regulated in sonic hedgehog-driven medulloblastomas and induced by N-myc in sonic hedgehog-treated cerebellar neural precursors. Cancer Res. 2009;69(8):3249–55.
Uziel T, Karginov FV, Xie S, Parker JS, Wang YD, Gajjar A, et al. The miR-17∼92 cluster collaborates with the Sonic Hedgehog pathway in medulloblastoma. Proc Natl Acad Sci U S A. 2009;106(8):2812–7.
Ivanovska I, Ball AS, Diaz RL, Magnus JF, Kibukawa M, Schelter JM, et al. MicroRNAs in the miR-106b family regulate p21/CDKN1A and promote cell cycle progression. Mol Cell Biol. 2008;28(7):2167–74.
Wong P, Iwasaki M, Somervaille TC, Ficara F, Carico C, Arnold C, et al. The miR-17-92 microRNA polycistron regulates MLL leukemia stem cell potential by modulating p21 expression. Cancer Res. 2010;70(9):3833–42.
Wang Y, Baskerville S, Shenoy A, Babiarz JE, Baehner L, Blelloch R. Embryonic stem cell-specific microRNAs regulate the G1-S transition and promote rapid proliferation. Nat Genet. 2008;40(12):1478–83.
Dolezalova D, Mraz M, Barta T, Plevova K, Vinarsky V, Holubcova Z, et al. MicroRNAs regulate p21(Waf1/Cip1) protein expression and the DNA damage response in human embryonic stem cells. Stem Cells. 2012;30(7):1362–72.
Hong L, Lai M, Chen M, Xie C, Liao R, Kang YJ, et al. The miR-17-92 cluster of microRNAs confers tumorigenicity by inhibiting oncogene-induced senescence. Cancer Res. 2010;70(21):8547–57.
Trompeter HI, Abbad H, Iwaniuk KM, Hafner M, Renwick N, Tuschl T, et al. MicroRNAs MiR-17, MiR-20a, and MiR-106b act in concert to modulate E2F activity on cell cycle arrest during neuronal lineage differentiation of USSC. PLoS One. 2011;6(1):e16138.
Fontana L, Fiori ME, Albini S, Cifaldi L, Giovinazzi S, Forloni M, et al. Antagomir-17-5p abolishes the growth of therapy-resistant neuroblastoma through p21 and BIM. PLoS One. 2008;3(5):e2236.
Ferretti E, De Smaele E, Miele E, Laneve P, Po A, Pelloni M, et al. Concerted microRNA control of Hedgehog signalling in cerebellar neuronal progenitor and tumour cells. EMBO J. 2008;27(19):2616–27.
Galardi S, Mercatelli N, Giorda E, Massalini S, Frajese GV, Ciafre SA, et al. miR-221 and miR-222 expression affects the proliferation potential of human prostate carcinoma cell lines by targeting p27Kip1. J Biol Chem. 2007;282(32):23716–24.
Gillies JK, Lorimer IA. Regulation of p27Kip1 by miRNA 221/222 in glioblastoma. Cell Cycle. 2007;6(16):2005–9.
le Sage C, Nagel R, Egan DA, Schrier M, Mesman E, Mangiola A, et al. Regulation of the p27(Kip1) tumor suppressor by miR-221 and miR-222 promotes cancer cell proliferation. EMBO J. 2007;26(15):3699–708.
Constantin L, Constantin M, Wainwright BJ. MicroRNA biogenesis and hedgehog-patched signaling cooperate to regulate an important developmental transition in granule cell development. Genetics. 2016;202(3):1105–18 .PMCID: 4788112
Gibcus JH, Kroesen BJ, Koster R, Halsema N, de Jong D, de Jong S, et al. MiR-17/106b seed family regulates p21 in Hodgkin’s lymphoma. J Pathol. 2011;225(4):609–17.
Roussel MF, Hatten ME. Cerebellum development and medulloblastoma. Curr Top Dev Biol. 2011;94:235–82.
Gong C, Qu S, Lv XB, Liu B, Tan W, Nie Y, et al. BRMS1L suppresses breast cancer metastasis by inducing epigenetic silence of FZD10. Nat Commun. 2014;5:5406.
Shen G, Jia H, Tai Q, Li Y, Chen D. miR-106b downregulates adenomatous polyposis coli and promotes cell proliferation in human hepatocellular carcinoma. Carcinogenesis. 2013;34(1):211–9.
Yu F, Lu Z, Huang K, Wang X, Xu Z, Chen B, et al. MicroRNA-17-5p-activated Wnt/beta-catenin pathway contributes to the progression of liver fibrosis. Oncotarget. 2016;7(1):81–93 .PMCID: 4807984
Wang VY, Rose MF, Zoghbi HY. Math1 expression redefines the rhombic lip derivatives and reveals novel lineages within the brainstem and cerebellum. Neuron. 2005;48(1):31–43.
Yan H, Hong P, Jiang M, Li H. MicroRNAs as potential therapeutics for treating spinal cord injury. Neural Regen Res. 2012;7(17):1352–9.
Acknowledgments
We are very grateful to Dr. Alexandra L. Joyner for her helpful comments and critical reading of this manuscript. This work was supported by grants from the National Natural Science Foundation of China (30971633 and 31171045), the Department of Science and Technology of Sichuan Province (Young Scientific Innovation Team in Neurological Disorders grant 2011JTD0005), and the Program for Changjiang Scholars and Innovative Research Team in University (PCSIRT) (IRT0935).
Author Contributions
Q.L., M.J., and H.L. designed the research; Q.L., M.J., Y.K., X.S., J.L., and H.L. performed the research; Q.L., M.J., M.W.L., and H.L. analyzed the data; and H.L. wrote the paper.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of Interest
The authors declare that they have no conflict of interest.
Additional information
Qian Liu and Mei Jiang contributed equally to this work.
Rights and permissions
About this article

Cite this article
Liu, Q., Jiang, M., Kuang, Y. et al. Dicer1 Ablation Impairs Responsiveness of Cerebellar Granule Neuron Precursors to Sonic Hedgehog and Disrupts Expression of Distinct Cell Cycle Regulator Genes. Cerebellum 16, 450–461 (2017). https://doi.org/10.1007/s12311-016-0821-x
Published:
Issue Date:
DOI: https://doi.org/10.1007/s12311-016-0821-x