
Abstract
Plantain (Musa spp. AAB genome) is a major staple with an incredibly enormous potential for domestic and global trade that can serve as a vital source of income for many developing countries. The fruits are extraordinarily significant, providing an important and cheap source of dietary energy, vitamins, and several essential minerals to millions of consumers within the humid and sub-humid tropics. As molecular characterization is important for sustainable exploitation of crops, the extent of genetic and agro-morphological differentiation in a large Musa population of 109 accessions was evaluated using 10 microsatellite (SSR) markers and 14 morphotaxonomic descriptors, as a basis for selection of high yielding varieties. Measurement of DNA diversity delineated the plantains into 2 major groups and 8 distinct clusters, each with several sub-clusters. There was equally a very wide diversity detected in morphotaxonomic traits including pseudostem girth, number of bunches per plant, number of hands per bunch, number of fruits per bunch, number of fruits per hand, bunch weight, length of plant cycle, and flowering to harvest interval, which varied significantly (p < 0.05) amongst the accessions. Selection based on cluster analysis using standard morphological data also delineated the cultivars into 8 distinct clusters, with one of these containing samples that were most distant to all others and exhibited a 'double bunching' phenotype at fruiting; tacitly implying the presence of significant variability that symbolizes an excellent opportunity to bring about genetic improvement through selection of improved varieties. Principal component analysis based on morphotaxonomic traits revealed 7 attributes which contributed to 79.95% of total variation that was quite similar to the 71.87% detected using microsatellite fingerprinting. Correlation analysis between selected agronomic traits demonstrated that pseudostem girth was positively associated with bunch weight, number of hands per bunch, number of fruits per bunch, number of fruits per hand, and flowering to harvest interval across all accessions. Collectively, the data in this study suggest that plantain accessions with very unique traits expressing paramount horticultural significance are available within the diverse germplasm evaluated and are currently being collected for future breeding purposes to broaden the genetic base of this important crop.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Plantain (Musa spp. AAB genome) is a giant perennial herb of the genus Musa. It is one of the most versatile food crops in the tropics, where it is seen as a very important component of local food security and provides a substantial amount of income to local farming communities through internal trade (Crouch et al. 1998a). The crop is one of the few most important suppliers of dietary energy in the wet tropical and sub-tropical regions of the world, where it is cultivated and provides essential nutrition for more than half a billion people. The fruits (fingers) are of extraordinary significance, providing an important and cheap source of dietary carbohydrates, vitamins, and several essential minerals including potassium, sodium, phosphorus and iron. Presently, sub-Saharan Africa produces more plantain than the rest of the world (FAOSTAT 2006). In fact, in many areas of the continent plantain and a few other crops such as cassava and yam constitute either a year-round or seasonal basic staple foods. Plantain equally serves as a raw material in many rural-based cottage industries including the production of wine (Ssebuliba et al 2000), vinegar and local beer, which is important nutritionally as it is described to be very rich in vitamin B due to the high yeast content in the brew. Aside from these, plantain is equally converted into high quality flour when the green (unripe) fruit is peeled, and the pulp dried and powdered. Such flour is known to be a very good source of energy and usually preferred by diabetics as it has been demonstrated to be more digestible than that derived from any of the common cereals (Brisibe and Ekanem 2019).
There are currently about one hundred and sixteen (116) different cultivars of plantain which are maintained by farmers under field conditions in different parts of Africa, but the frequency of occurrence of these cultivars vary from one country to the other as well as between different agro-ecological zones within the same country (Swennen 1997; Ogazi 1996). Up until recently, identification of this wide selection of plantain cultivars on the continent was traditionally based on morphological attributes as phenotypic description has proven very useful for distinguishing the large diversity existing amongst plantain cultivars in the tropics (Tezanas du Montcel 1987; Jarret and Gawel 1995). However, this method of distinction does not adequately provide a clear revelation of the close genetic and horticultural relationships that exist amongst the different plantain varieties cultivated due, in part, to frequent somatic mutations and morphological changes that are occasioned by the environment, which poses a major obstacle towards proper identification of all the cultivars (Kaemmer et al. 1992) on the continent.
Thus far, progress in plantain production in Africa has depended to a large extent on the ability to select high yielding cultivars from a segregating population (IITA 2007). And given that the estimation of genetic diversity in a crop species is a prerequisite for its improvement, there is an inspiring possibility that different plantain cultivars can be improved once genetic variability has been ascertained using appropriate selection indices. However, as the expected genetic response to selection is determined by heritability and the variability of the traits for which the crop selected for cultivation are to be made relative to the farming systems prevalent in a given socio-economic environment, there is no doubt that an accurate knowledge of genetic diversity and relationships among plantain collections in any preserved germplasm is essential and important for the establishment, management and warranty of long-term success of plantain improvement programmes through breeding (Gwag et al. 2010).
Plantain cultivars show considerable variations for many horticultural traits such that different criteria including pedigree records, morphological traits and DNA marker technology (Weising et al. 2005) have been used in the past to estimate genetic diversity prevalent within the species. These criteria are relevant even today as morphological descriptors have remained the major indices for varietal selection by farmers in the field. In spite of this, however, there is still general paucity of information on genetic diversity amongst plantain varieties cultivated. For example, there is practically no scientific information on pseudostem characteristics, bunch sizes, shape of fruits as well as the yield potential of plantain varieties predominantly maintained by farmers in the field. Consequently, no robust selection and breeding programmes have been undertaken in the past with a view to conserving and further improving high yielding plantain cultivars already in existence.
Like with banana (Musa spp. ABB genome), different plantain producing areas currently suffer from newly emerging pests and diseases and rapidly changing environmental conditions with adverse effects. In the absence of locally adapted resistant varieties and a general lack of characterized germplasm that could be used as potential parents for breeding purposes, farmers would need to extensively use pesticides, which threaten the sustainability of not only the crop but also the environment (Pennisi 2010). There is therefore an urgent need for selection of accessions with a wider diversity of genetically improved traits with more robust disease resistance, increased productivity and better adaptability to a wider range of growing conditions. In consideration of the fact that an understanding of the variability amongst different plantain varieties would be desirable for setting up of an efficient strategy for breeding improved cultivars and support the choice of parents that can be used for regeneration, a very robust appreciation of the genetic and morphotaxonomic diversity of available resources will be of paramount importance. Consequently, the current study was designed to provide details on the diversity available, based on information generated through molecular fingerprinting and variations in agro-morphological traits within a large plantain germplasm collection, as a foundation for selection and conservation of genetically superior cultivars that can be used for further breeding and improvement of this economically important staple crop.
Materials and methods
Plant material and DNA extraction
A total of one hundred and nine (109) Musa (AAB genome) accessions from different localities spread across the humid forest zone in south-eastern Nigeria, which were predominantly available and maintained in an in situ germplasm collection under farmers’ field conditions, were evaluated during the production cycles between 2012 and 2016. Global Positioning System (GPS Etrex Legend, Garmin model) was catalogued to identify the longitude, latitude and altitude of the sites where the accessions were sampled but these details are not provided in the current study due to the large size (supplementary information available on request).
For extraction of genomic DNA, young leaf tissue was initially collected from each of the plantain accessions, packaged and outsourced to a commercial biotechnology laboratory according to the modified CTAB method of Saghai-Maroof et al. (1984) with 2% (v/v) β-mercaptoethanol (added just before use) and 2% (w/v) polyvinylpyrrolidone, followed by RNase treatment for the removal of contaminating RNA.
PCR amplification and marker visualization
Polymerase chain reaction mixture containing 100 ng of template DNA, 2.5 mM of MgCl2, 100 µM of dNTPs, 1X Taq buffer, 20 micromoles of 10-mer primer, and 1 unit of Taq DNA polymerase was used in an Eppendorf tube. Distilled and deionised water was added to this mixture to make a total volume of 25 μl in each tube. DNA amplification was performed in a Mastercycler Gradient Thermocycler (Eppendorf, Hamburg, Germany) using a thermal profile of 94 °C for 5 min (10 cycles), 72 °C for 2 min (10 cycles) and a final extension at 72 °C for 10 min (10 cycles). The samples were cooled at 4 °C.
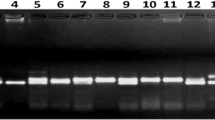
The amplification products were visualized in an agarose gel (1.2%) containing ethidium bromide (for electrophoresis at 125 V) in a 1X Tris-boric acid-EDTA buffer solution. The electropherograms were documented using a gel AlphaImager documentation analysis system (Alpha Innotech, USA). Bands were compared with 100 and 500 bp DNA standard markers (BioGene, USA). All amplifications were performed thrice with genomic DNA isolated independently to confirm the reproducibility.
Primers designed and used for the study
Ten simple sequence repeat (SSR) or microsatellite primer pairs were designed based on the reports of Crouch et al. (1998b), Lagoda et al. (1998) and Hippolyte et al. (2010) for Musa species. The primers, whose forward and reverse sequences and type of repeat motifs are presented in Table 1, were sourced from ‘wet lad’ of Operon Technology (USA) and used for this study. The sourced primer pairs were screened for polymorphism against all 109 plantain accessions. Primer pairs that produced distinct bands and patterns with good quality of amplicons and reproducibility were selected and used for molecular fingerprinting study.
SSR fragment analysis
The fragment sizes in base pairs for each accession across SSR markers were converted to binary data, where alleles were transformed into presence (1) or absence (0) of a microsatellite band. The genetic diversity indices such as number of alleles per locus, gene diversity and polymorphism information content (PIC) were estimated using GenAlex software. The cluster pattern of the genetic diversity was constructed using DARWIN 5.0 software.
Determination of morphotaxonomic and yield traits
Visual observations indicated that there were significant differences between the 109 accessions in terms of morphotaxonomic attributes. Consequently, the International Board for Plant Genetic Resources (IBPGR 1988) descriptor list for Musa species was used to characterize all the accessions in the current study. Experimental field data using fourteen morphological descriptors that were taxonomically most informative for characterization of the pseudostem, leaf, floral parts, bunches, hands, and fruits following procedures outlined for sampling in the IBPGR descriptor list for Musa species were collected. Six replicates were taken to represent an accession in each sampling site for key morphological attributes and yield-related traits such as pseudostem (plant) girth, pseudostem height, number of fruits (fingers) per hand, number of hands per bunch and bunch weight as the mean value of four measurements per replicate. Qualitative traits including pseudostem colour, fruit skin colour, fruit pulp colour, bunch size, shape of cross section of fruit and shape of fruit were determined by careful observation and comparison with the IBPGR descriptor list and Royal Horticultural Standard colour chart according to Agoreyo et al. (2008). Mature fruits were carefully harvested and measurements taken for all agro-morphological and yield-related traits. A sharp kitchen knife was used to cut fruits to separate bunches (where there were more than one), hands and fruits. A conventional weighing balance (calibrated in kg) was used to measure bunch weight while a digital camera (Nikon, Japan) was used for taking photographs of the different parameters for qualitative observations. The length of production cycle (in months) and flowering to harvest interval (in days) for each accession were also recorded.
Principal component analysis
The general linear model procedure was specified for ANOVA (at p < 0.05%). Multivariate examination was performed using principal component analysis (PCA) to determine the relative importance of traits accounting for variability among the 109 plantain accessions. These statistical analyses were undertaken using the computer software Genstat, version 6.0 (ICRAF).
Cluster analysis
A cluster analysis specifying the nearest relative or neighbour option based on the Euclidean distances was used to explore relationships among the accessions. Clusters were graphically represented in a dendrogram by using Unweighted Pair Group Method with Arithmetic Means (UPGMA), which was implemented in DARwin software according to Perrier and Jacquemoud-Collet (2006).
Data processing and analysis
The data collected for all the traits observed above were subjected to descriptive statistics, one way analysis of variance (ANOVA) and correlation analysis for quantitative traits, while frequency distribution was adopted for qualitative traits.
Results
Characterization of microsatellite marker polymorphism in plantain accessions (n = 109)
According to the standardized platform set for molecular characterization of Musa germplasm based on microsatellite (SSR) markers, six out of the 10 primer pairs (Table 1) used in this study for DNA fingerprinting of the 109 plantain accessions were informative and polymorphic. These markers produced well-defined and reproducible bands, which were used to calculate the number of unique alleles, range of band size, polymorphism information content (PIC), and other indices exhibited by the SSR primers for all the accessions. Based on the characterization data generated, the molecular information for all accessions were developed and used for analysis of the principal components.
As presented in Table 2, genetically different groups of accessions were detected. The data revealed that five principal components contributed a total of 71.87% to the variation observed among the plantain accessions. Principal component (PC) 1 with an Eigen value of 0.06566 contributed the highest percentage of 23.14% to the total variability observed. Five accessions (No. 17, 19, 25, 33, and 62) were the major contributors to this component, with each having a loading value of 0.6709. Principal component (PC) 2, on the other hand, had an Eigen value of 0.05626 and contributed 19.83% to the total variability observed. The major contributors to this component were also five accessions (No. 8, 11, 14, 22, and 95), with each having an equal loading score of 0.4904. Principal component (PC) 3 presented an Eigen value of 0.04699, while contributing 16.56% to the total variation observed. Four accessions (No. 7, 23, 31 and 54) were the major contributors to this component. While accessions number 23 and 54 each had a loading value of 0.7005, the remaining two, that is, 7 and 31 had a loading score of 0.5348, respectively.
Table 3 equally showed that principal component (PC) 4 had an Eigen value of 0.02640 and contributed about 9.31% to the total variations observed in the large plantain germplasm. Five accessions (No. 39, 67, 94, 101, and 105) were the major loading contributors to this component, with loading values of 0.2839 (for accessions 39 and 105), 0.3141 (for 67 and 94), and 0.4834 (for 101), respectively. Principal component (PC) 5 was the least contributing component to the overall variations observed. This component had an Eigen value of 0.0861 and contributed only 3.03% to the total variations observed. The major contributors to this principal component and their loading values were accessions number 91 (0.2794), 93 (0.3144), and 100 (0.3730), respectively.
Establishment of genetic relationships for the plantain accessions based on the microsatellite (SSR) data generated
With the microsatellite or SSR fingerprinting data generated, it was possible to delineate the genetic landscape of all the 109 plantain accessions into major groups, clusters and sub-clusters, respectively, on the basis of their relationships as presented in Figs. 1, 2, and 3. The dendrogram, based on UPGMA cluster analysis, resulted in relatively clear grouping of all the accessions into two major varietal groups of False Horn and True Horn plantains, respectively. The False Horn plantain collections represented the larger of the two groups and were composed of more genetic resources, which were clearly separated from those of the True Horn plantains. The dendrogram equally clustered all the 109 accessions into eight distinct or major clusters (designated as C1–C8). Major cluster 1 had five sub-clusters with each grouped on the basis of the molecular peculiarities of the accessions. Similarly, major cluster 2 had three sub-clusters which were also grouped based on their molecular peculiarities. It is obvious from these data that formation of the different groups, clusters and sub-clusters reiterated the presence of diverse genetic variability within and between the plantain germplasm. It is equally intriguing that the establishment of these molecular delineations are in conformity with those detected using the morphotaxonomic descriptors of the plantain accessions, which are well documented in the later part of this paper.

Scatter plots for 109 plantain accessions (Musa spp. AAB genome) based on microsatellite marker-derived data

One hundred and nine (109) accessions of plantain showing 8 groupings based on microsatellite marker-derived data

Dendrogram for 109 plantain accessions showing 2 groups and 8 major clusters based on microsatellite marker-derived data
Variation in morphological (quantitative) characteristics and yield-attributing traits
In situ evaluation of all 109 accessions for four consecutive years in a second series of experiments that compared the mean ± standard error, minimum and maximum values, and coefficient of variation generated from the population with indications of variations in some of the morphological characteristics and yield-related traits are shown in Table 3. The analysis of divergence demonstrated that while pseudostem height and number of leaves per plant at harvest did not vary significantly, differences in pseudostem girth and length of plant cycle were significant (p < 0.05) amongst the 109 accessions. The height of the plants (pseudostem) in most of the study sites generally ranged between 1.83 and 3.63 m with a coefficient of variation of 13.48%. The pseudostem girth usually ranged between 38.97 and 59.13 cm, with a coefficient of variation of 8.67%. The number of leaves per plant at harvest was between 5 and 9 with a coefficient of variation of 10.13%. Usually, the length of the plant cycle for the varieties evaluated ranged between 12 and 18 months with a variation coefficient of 13.05%.
Regarding yield-related traits, only one bunch was usually harvested per plant though plants in one of the study sites showed inflorescence dichotomization which resulted in the production of ‘double bunches’ on the same pseudostem at fruiting with a coefficient of variation of 0.035%. The number of hands per bunch counted for the accessions usually ranged between 3 and 13. The number of fingers per hand ranged between 4 and 8 with a coefficient of variation of 17.62% while the number of fingers per bunch ranged between 15 and 72 with an average of 51.69 and a coefficient of variation of 36.21%. Overall, the average weight of the plantain bunches was 13.07 kg, though individual weight of bunches ranged between 5.02 and 26.76 kg with a coefficient of variation of 43.15%. The flowering to harvest interval showed an average of 75 days and usually ranged from 43 to 84 days.
Cluster analysis based on morphological characteristics and yield-attributing traits of plantain accessions (n = 109)
Like with the data generated during molecular characterization of the accessions, results of the delineation of all 109 plantains based on morphotaxonomic characterization suggested that two major groups of Musa cultivars (False Horn and True Horn types) could be identified. In order to classify the accessions based on similarity indices of the agro-morphological traits, a single linkage cluster analysis using Nei’s (1978) genetic distance was used to generate eight clusters (Fig. 4) with specific characteristics as presented in Table 4. The proximity Euclidean distances between the 8 cluster groups generated are provided in Table 5. Each of the clusters can be described as follows:

Dendrogram revealing 2 groups and genetic distances amongst eight (8) clustered groups of plantain cultivars using Euclidean distances generated from morphotaxonomic data (FHP = False Horn plantain; THP = True Horn plantain)
Cluster 1 (n = 9)
Plantain accessions in this cluster were characterized by the presence of usually only one big individual bunch per plant. Morphologically, they were of the True Horn type with a pseudostem that was purple in colour. The individual unripe fruits (or fingers) of bunches were usually pale green in colour, with a tetragonal cross section and creamy pulp. The crop had an average planting cycle of 14 months, an average of 7 leaves per plant and an average pseudostem height and girth of 2.61 m and 46.81 cm, respectively, at harvest.
In terms of yield, these plants produced an average bunch weight of 23.43 kg, about 10 hands per bunch, and a mean of 7 and 64 big fingers per hand and bunch, respectively. The plants usually had a flowering to harvest interval of 69 days.
This cluster was observed to be most distant from clusters 4 (distance = 1303.1) and 5 (distance = 2054.8) but was closest to clusters 2 (distance = 52.7) and 3 (distance = 41.7), respectively, as shown in Table 5.
Cluster 2 (n = 8)
These accessions were also characterized by big bunch sizes with only one bunch per plant. They were predominantly found in plantations alongside tree crops, which provided good organic matter to the soil. Morphologically, the plants in this cluster were of the False Horn type, with greyish pseudostem and fruits with creamy pulp colour and a triangular cross section. The unripe fruits of these cultivars were light yellow in colour and plants had an average planting cycle of 13 months. The cultivars showed an average of 6 leaves per plant at harvest with an average pseudostem height and girth of 2.48 m and 44.53 cm, respectively.
In terms of yield, these plants produced an average bunch weight of 21.78 kg, a mean of 10 hands per bunch, a mean of 6 and 58 fingers per hand and bunch, respectively, and a flowering to harvest interval of 74 days. The cluster was most distant from clusters 4 (distance = 884.8) and 5 (distance = 1514.1), while closest to cluster 3 (distance = 40.1).
Cluster 3 (n = 8)
The accessions in this cluster were characterized by big individual bunches, which was usually one per plant. The plants were found predominantly in localities where the soil had high organic matter and humus content. Morphologically, the plants were of the giant True Horn type, with brownish pseudostem and fruits with a triangular cross section and creamy white pulp while the unripe fruits were deep green in colour. Generally, the plants in this cluster had an average planting cycle of 15 months with an average of 7 leaves per plant at harvest. The plants also had an average pseudostem height and girth of 2.55 m and 48.77 cm, respectively.
In terms of yield, these plants showed an average bunch weight of 24.96 kg, average of 12 hands per bunch, average of 7 and 59 fingers per hand and bunch, respectively, and a flowering to harvest interval of 74 days. This cluster was most distant from cluster 4 (distance = 1048.4) and cluster 5 (distance = 1686.4), while closest to cluster 1 (distance = 52.7) and cluster 2 (distance = 40.1), respectively..
Cluster 4 (n = 11)
These accessions were characterized by small individual bunches, with only one bunch per plant. Morphologically, the plants were of the small False Horn type, with purplish pseudostem and fruits that had a tetragonal cross section and milky white pulp. The unripe fingers were pale green in colour while the plants had an average planting cycle of 14 months and 7 leaves per plant at harvest. The plants had an average pseudostem height and girth of 2.41 m and 47.92 cm, respectively.
In terms of yield, these plants produced an average bunch weight of 8.39 kg, average of 5 hands per bunch, an average of 5 and 32 fingers per hand and bunch, respectively, and a flowering to harvest interval of 75 days. This cluster was most distant from cluster 1 (distance = 1303.1) and cluster 3 (distance = 1048.4), while closest to cluster 5 (distance = 104.2).
Cluster 5 (n = 16)
These accessions were also characterized by small individual bunches, but bore some peculiar features such as the presence of a dichotomous inflorescence that produced 2 bunches per plant at fruiting. Morphologically, the plants were of the small False Horn plantain type, with purplish pseudostem, fruits that were pentagonal in cross section and creamy pulp in colour. The unripe fingers of these cultivars were pale green in colour while the plants had the longest planting cycle of 17 months amongst all the cultivars evaluated in the current study. The cultivars presented an average of 6 leaves per plant at time of harvest with an average pseudostem height and girth of 2.81 m and 48.29 cm, respectively.
In terms of yield, these accessions showed an average bunch weight of 7.81 kg, with an average of 4 hands per bunch, an average of 4 and 23 fingers per hand and bunch, respectively, and an interval of 64 days from flowering to harvesting. This cluster was most distant from cluster 1 (distance = 2054.8) and cluster 3 (distance = 1686.4), while closest to cluster 4 (distance = 104.2).
Cluster 6 (n = 26)
The accessions in this cluster were characterized by medium-sized individual bunches, with only one bunch per plant. Morphologically, they were of the False Horn plantain type too, with greyish pseudostem and a pentagonal cross section of the fruits that had pulp that were cream in colour. The unripe fruits were deep green in colour and the plants had an average of 13 months of planting cycle. The cultivars showed an average of 7 leaves per plant at time of harvest with an average pseudostem height and girth of 2.80 m and 46.14 cm, respectively.
In terms of yield, these accessions demonstrated a mean bunch weight of 12.35 kg, average of 8 hands per bunch and a mean of 6 and 48 fingers per hand and bunch, respectively, and an interval of 67 days from flowering to harvesting. The cluster was most distant from cluster 5 (distance = 686.1) and cluster 1 (distance = 418.7), while closest to cluster 7 (distance = 70.8).
Cluster 7 (n = 15)
These accessions were characterized by medium-sized individual fruits too, with only one bunch per plant. Morphologically, they were of the False Horn type, with brownish pseudostem and fruits that were triangular in cross section and pulp that was milky white in colour. The unripe fruits of the plants were light yellow in colour while the plants had an average of 15 months of planting cycle. The accessions recorded an average of 7 leaves per plant remaining at harvest with an average pseudostem height and girth of 2.75 m and 48.99 cm, respectively.
In terms of yield, these cultivars showed an average bunch weight of 11.88 kg, average of 8 hands per bunch and an average of 6 and 55 fingers per hand and bunch, respectively, and an interval of 77 days from flowering to harvesting. The cluster was most distant from cluster 5 (distance = 1089.7) and closest to cluster 8 (distance = 43.1).
Cluster 8 (n = 16)
The accessions in this cluster were characterized by medium-sized individual bunches, with only one bunch per plant. Morphologically, the plantains were of the True Horn type, with purplish pseudostem and fruits that had a tetragonal cross section and pulp that was creamy white in colour. The unripe fruits of these cultivars were pale green in colour with an average of 14 months of planting cycle. The accessions showed an average of 6 leaves per plant remaining at harvest and an average pseudostem height and girth of 2.85 m and 51.83 cm, respectively.
In terms of yield, these cultivars showed an average bunch weight of 13.22 kg, average of 10 hands per bunch, an average of 7 and 6 fingers per hand and bunch, respectively, and an interval of 71 days from flowering to harvesting. This cluster was most distant from cluster 5 (distance = 1522.3) and closest to cluster 7 (distance = 43.1) as presented in Fig. 4.
Variation in morphological and yield-related traits amongst cluster groups of plantain accessions
Now, in an attempt to demonstrate the degree of divergence amongst the 8 cluster groups of plantain, analysis of variance was conducted separately for each of the agro-morphological traits with quite intriguing results. While differences in some of the traits evaluated were not significant amongst the cluster groups, others including pseudostem girth, flowering to harvest interval and length of plant cycle demonstrated significant (p < 0.05) variations. For example, while plants in cluster 3 recorded the widest average pseudostem girth of 51.77 cm, those in cluster 5 produced the smallest average girth of 30.29 cm. Moreover, while accessions in cluster 3 required 74 days for fruit filling or flowering to harvest interval, those in cluster 5 required 64 days to complete fruit filling. In addition, while the plantain varieties in clusters 1, 2 and 6 required only an average of 13 months, those in cluster 5 showed the longest duration of 17.36 months to complete their planting cycles. Interestingly, there was no significant (p < 0.05) variation amongst the plants in all the cluster groups for pseudostem height. Average number of leaves per plant at harvest also did not differ (p > 0.05) significantly among the cluster groups. Often, a maximum of 7 but usually a minimum of 6 leaves per plant were observed among plants in all the cluster groups.
Unlike the data generated for the morphological characteristics, those for yield-related traits among the 8 cluster groups presented a completely different picture. While certain traits including number of hands per bunch, number of fruits per hand, number of fruits per bunch, bunch weight and flowering to harvest interval varied significantly (p < 0.05), however, there were no significant differences in the number of bunches per plant among the 8 distinct cluster groups of plantain. Generally, plants in 7 out of the 8 cluster groups had only one bunch except for those in cluster 5 that presented 2 bunches at maturity. The number of hands per bunch revealed significant (p < 0.05) differences among the cluster groups with accessions in cluster 3 having the highest number of hands (12.44) per bunch while those in clusters 4 and 5 had the least number of 5.09 and 4.50 hands per bunch, respectively. The number of fingers per hand also showed significant (p < 0.05) variations among the cluster groups. While accessions in cluster 3 had an average of 8.33 fruits per hand, those in cluster 5 had the least number of 4.87 fingers. Similarly, the number of fingers per bunch also differed significantly among the cluster groups. Cluster 1 produced the highest number of 64.88 fingers per bunch while cluster 5 produced the least number of fingers (23.06), respectively. Significant (p < 0.05) variation was also observed in bunch weight among the cluster groups. The heaviest mean bunch weight of 24.96 kg was obtained from plants in cluster 3 while those in cluster 5 produced the least mean bunch weight of 7.81 kg.
Principal component analysis of morphotaxonomic traits of the plantain accessions (n = 109)
With the exclusion of the four highly correlated yield-related traits of bunch weight, number of hands per bunch, number of fruits per hand and number of fruits per bunch, a multivariate evaluation based on principal component analysis was equally performed by considering 10 related agro-morphological descriptors (Table 6). High morphological variations were recorded among the accessions evaluated. The majority of the descriptors were significantly correlated and were thus used as a basis for comparison with the contributions made by the SSR fingerprinting data to the variations observed among the 109 plantain accessions. Intriguingly, with the morphotaxonomic traits the first seven principal component axes accounted for 79.95% of total variability, which is reasonably similar and consistent with the 71.87% detected with the five principal components obtained using molecular or DNA fingerprinting to partition the accessions. The first principal component axis, which made up 16.70% of total variation, was positively correlated with pseudostem girth (0.595), length of plant cycle (0.601), flowering to harvest interval or fruit filling (0.476) and area of fruit cross section (0.544).
The second principal component axis, accounting for 14.37% of total variation, was strongly correlated with pseudostem girth (0.395), length of plant cycle (0.432), flowering to harvest interval (0.445) and area of fruit cross section (0.432). The number of leaves at harvest (0.451), length of plant cycle (0.525) and area of finger cross section (0.888) were strongly correlated with the third principal component axis which accounted for 12.48% of total variation.
The fourth principal component axis, making up 10.03% of total variation, was strongly correlated with pseudostem height (0.733), length of plant cycle (0.472), area of fruit cross section (0.521) and finger skin colour (0.443). The fifth principal component axis constituting only 9.14% of the total variation observed among the accessions was strongly correlated with pseudostem girth (0.459), number of leaves at harvest (0.465), area of finger cross section (0.444) and fruit skin colour (0.500). The sixth principal component axis making up 8.65% of total variation was mainly correlated with pseudostem height (0.344), pseudostem girth (0.333), number of leaves at harvest (0.522), length of plant cycle (0.671) and flowering to harvest interval (0.597).
Correlation analysis amongst the morphological characteristics and yield-related traits
In a bid to establish the degree of relatedness amongst and between the morphological characteristics and yield-related traits, a correlation analysis was undertaken. Data presented in Table 7 show significant and positive correlations between the number of hands per bunch and number of fingers per bunch (r = 0.86, p < 0.05), number of hands per bunch with number of fingers per hand (r = 0.71, p < 0.05) and number of hands per bunch with bunch weight (r = 0.79, p < 0.05). The number of fingers per hand was also highly correlated with the number of fingers per bunch (r = 0.93, p < 0.05) and the number of fingers per hand with bunch weight (r = 0.40, p < 0.05). The number of fingers per bunch also correlated positively with bunch weight (r = 0.57, p < 0.05).
The pseudostem girth correlated positively with some yield-related traits such as number of fingers per hand (r = 0.83, p < 0.05), number of fingers per bunch (r = 0.75, p < 0.05), number of hands per bunch (r = 0.76, p < 0.05), bunch weight (r = 0.63, p < 0.05), and flowering to harvest interval (r = 0.57, p < 0.05). The number of leaves per plant at harvest not only correlated positively with pseudostem height (r = 0.03, p < 0.001), but also with bunch weight (r = 0.04, p < 0.001). Conversely, some morphological traits correlated negatively with some yield-related traits. For example, the number of leaves per plant correlated negatively with pseudostem height (r = − 0.01, p < 0.05), length of cycle (r = − 0.15, p < 0.05), number of bunches per plant (r = − 0.02, p < 0.05), number of hands per bunch, number of fingers per hand, number of fingers per bunch (r = − 0.01, p < 0.05) and with flowering to harvest interval (r = − 0.02, p < 0.05). Not surprisingly, pseudostem girth negatively correlated with length of plant cycle (r = − 0.07, p < 0.05). On the other hand, pseudostem height was also negatively correlated with length of plant cycle (r = − 0.19, p < 0.05), number of hands per bunch (r = − 0.05, p < 0.05), bunch weight (r = − 0.08, p < 0.05), and flowering to harvest interval (r = − 0.14, p < 0.05).
Length of plant cycle negatively correlated with number of hands per bunch (r = − 0.20, p < 0.05), number of fruits per hand (r = − 0.17, p < 0.05), number of fruits per bunch (r = − 0.19, p < 0.05), number of bunches per plant (r = − 0.14, p < 0.05) and flowering to harvest interval (r = − 0.13, p < 0.05). Also, number of bunches per plant was negatively correlated with number of hands per bunch (r = − 0.57, p < 0.05), number of fingers per hand (r = − 0.63, p < 0.05), number of fruits per bunch (r = − 0.64, p < 0.05), bunch weight (r = − 0.40, p < 0.05) and flowering to harvest interval (r = − 0.12, p < 0.05). Flowering to harvest interval correlated negatively with number of hands per bunch (r = − 0.01, p < 0.05), number of fruits per hand (r = − 0.11, p < 0.05) and number of fruits per bunch (r = − 0.09, p < 0.05).
Discussion
The data presented in the current study clearly demonstrated that a huge amount of genetic diversity as well as variations in morphotaxonomic traits was harboured by the 109 plantain accessions evaluated. For instance, just as there were differences in the number of DNA amplification bands with the microsatellite markers used these variations were also significant to enable the delineation of the accessions into 8 distinct cluster groups. This is an indication that enough simple sequence repeat polymorphism existed to discriminate between the individual Musa accessions, perhaps on account of the multiplicity of mutations that have been generated over the years following the high rate of varietal multiplication by local farmers in the field. This is equally buttressed by the pseudostem girth, which also varied significantly amongst all the accessions evaluated just like has been reported by Phillipe et al. (2012) and Dzomeku et al. (2004). Moreover, the fact that most of the yield-related traits also presented significant variations among the accessions is an additional confirmation of the presence of extreme diversity. For example, while the number of bunches borne per plant was usually one (or two in a few cases), however, other yield-related traits including the number of hands per bunch, number of fruits per hand, number of fruits per bunch, and bunch size varied significantly. Furthermore, weight of the individual bunches also varied significantly among the accessions, perhaps on account of the variations observed in pseudostem girth since these two traits were seen to be highly correlated. Interestingly, while the accessions with small bunches had an average weight of 5.02 kg, those with the big bunches weighed much more (26.76 kg on the average), thus corroborating results reported previously by Dzomeku et al. (2004), Vuylsteke et al. (1996), Agoreyo et al. (2008), Swennen (1997) and Kaemmer et al. (1992).
Following the significant diversity that was observed among the 109 plantain accessions in terms of morphological and yield-related traits evaluated, this was also used as a basis for their classification into eight major cluster groups, with some very unique features. For example, while accessions in clusters 1, 2 and 3 that were delineated on the basis of morphotaxonomic traits generally produced big bunches with curved fingers, those in clusters 4 and 5 had bunches that were small in size with flat fingers, and plants in the three remaining clusters (that is, 6, 7 and 8) had medium-sized bunches with flat fingers. Interestingly, the heavy bunch weight identified amongst the accessions in clusters 1, 2, and 3, is an indication of high fruit yield and quality by these plants. Their large fruit sizes is also an indication that these accessions have a superior and larger yield potential, which can be selected and exploited for sustenance of commercial plantain production both for agro-industrial processing and the fresh market within the country as well as for export. In addition, the high yield potential observed amongst these plantain varieties can further serve as a strong motivation for public investment in technology development which can equally be seen as major incentives for farmers to adopt them for commercial-scale cultivation. Above all of these and most significantly, the diversity in bunch weight and fruit size identified amongst the 8 cluster groups in this study also provides an exceptional opportunity for selection of high yielding parents among the clusters that can be used for breeding of elite cultivars with quite outstanding genetic qualities of horticultural significance. As with other interesting characters with simple inheritance, such cultivars can be further improved through in vitro micropropagation techniques, which can potentially allow the introgression of very important genes including, but not necessarily limited to, those for pest and disease resistance through genetic transformation of the plants. On the contrary, the small group of plants from cluster 5, which produced ‘double bunches’ on a single pseudostem at fruiting that was separated from the bulk of the other clusters, suggests that this group may harbour genetic resources with uniquely rare and fascinating alleles with good horticultural properties for widening the genetic base of plantain breeding populations.
Accordingly, a greater effort will be directed in the future at collecting such varieties presenting inflorescence dichotomization events and subjecting them to serious molecular characterization, as they did not receive much attention in the current study because of the rather inconsistent heritability of this rare but fascinating trait in the ratoon crops. This implies that though selection for the ‘double-bunching’ trait (Odeigah 1997; Tenkouano 2000; Ekanem and Brisibe 2018; Brisibe and Ekanem 2019) observed in cluster 5 may not be reflective of an inspiring horticultural significance at the moment, nevertheless, it may provide a basis for understanding the wider variation for inter-cluster genetic dissimilarities, which can be utilized for direct use as parents in hybridization programmes in the future to develop high yielding plantain cultivars.
Of equal interest and significance in the current study are the details generated from the multivariate analyses, which revealed that five and seven principal component axes were responsible for 71.87% and 79.95% of total variability observed on the basis of the data generated from molecular fingerprinting and agro-morphological evaluation, respectively. For this study, the most important morphological traits of horticultural significance included pseudostem height, pseudostem girth, bunch size, number of leaves at harvest and length of plant cycle. For qualitative traits, those found to be significant included pseudostem colour, finger skin colour, pulp colour, shape of cross section of the finger, and shape of the finger. Interestingly, most of these traits remained the same at the different growth stages of the plant and as such enables the identification of the accessions even at earlier stages of development such as when they were suckers. For yield-related traits, the number of bunches per plant, bunch weight, number of hands per bunch, number of fingers per bunch, and number of fingers per hand were considered the most important to be selected for plantain varietal improvement. The highly significant positive correlation observed between these yield-related traits such as number of fingers per bunch and bunch weight, number of hands per bunch and bunch weight, number of fingers per hand, number of fingers per bunch, number of bunches per plant and pseudostem height can be further examined and utilized for the selection of plantain cultivars with improved or desired traits such as big and heavy bunches for improved commercial value.
Conclusions
The large Musa germplasm in the study area displayed a very impressive diversity with significant genetic variation for a multitude of morphotaxonomic traits. The results of the principal component analyses revealed that there were highly significant differences among the 109 plantain accessions evaluated and these variations both in the microsatellite fingerprinting measurements as well as morphotaxonomic identification could be used as basis for delineation of all the accessions into major cluster groups, therefore supporting the fact that the two sets of data could be used either individually or collectively to compress all the major morphological descriptors and yield-related traits of the plantain accessions to five and seven principal components, respectively, with a cumulative contribution of 71.87% (for SSR fingerprinting) and 79.95% (for morphotaxonomic traits) of total variability. These results therefore demonstrate the utility of cluster analysis and PCA in partitioning the genetic variation among plantain accessions and in the identification of different accessions of plantains, which would serve as potential sources of unique breeding material for future crop improvement. The study also provides guidance for future analysis of genetic diversity in plantain accessions using other equally reliable molecular markers such as single nucleotide polymorphism (SNP) to facilitate efficient management and utilization of the available germplasm. On the basis of these findings, 8 elite accessions (one from each of the 8 major cluster groups using data generated from both SSR fingerprinting and morphotaxonomic demarcations) that were identified to perform well under the particular geo-climatic conditions of the area have been collected and are being conserved and maintained continuously as a unique genetic-resource pool for utilization in future breeding programmes of plantain.
References
Agoreyo BO, Golden KD, Brown SE (2008) Analysis of genetic variability among plantain cultivars (Musa paradisiaca L) using arbitrarily primed PCR technique. Afr J Biotech 7:1041–1045
Brisibe EA, Ekanem NG (2019) Ploidy level and nucleotide variations in inflorescence dichotomous cultivars of Musa species (AAB genome). BMC Genomics 20:713. https://doi.org/10.1186/s12864-019-6083-5
Crouch JH, Vulysteke D, Ortiz R (1998a) Perspectives on the application of biotechnology to assist the genetic enhancement of plantain and banana (Musa spp.). J Biotechnol 1:1–18
Crouch HK, Crouch JH, Jarret RL, Cregan PB, Ortiz R (1998b) Segregation at microsatellite loci in haploid and diploid gametes of Musa. Crop Sci 38:211–217
Dzomeku BM, Bam R, Ankomah AA, Yeboah DK (2004) Preliminary study on the performance of some Musa hybrids in Ghana. J Ghana Sci Assoc 6:90–94
Ekanem NG, Brisibe EA (2018) Inflorescence developmental polymorphism and its horticultural significance in plantain (Musa spp. AAB genome). Afr J Biotech 17:1–8
FAO (2006) Production yearbook 2006. Food and Agriculture Organization, Rome, Italy
Gwag JG, Dixit A, Park YJ, Ma KH, Kwon SJ, Cho GT, Lee GA, Lee SY, Kang HK, Lee SH (2010) Assessment of genetic diversity and population structure in mungbean. Genes Genomics 32:299–308
Hippolyte I, Bakry F, Seguin M, Gardes L, Rivallan R, Risterucci AM, Jenny C, Perrier X, Carreel F, Argout X, Piffanelli P, Khan IA, Miller RNG, Pappas GJ, Mbe´guie´-A-Mbe´guie D, Matsumoto T, De Bernardinis V, Huttner E, Kilian A, Baurens FC, D’Hont A, Cote F, Courtois B, Glaszmann JC (2010) A saturated SSR/DArT linkage map of Musa acuminata addressing genome rearrangements among bananas. BMC Plant Biol 10:65. https://doi.org/10.1186/1471-2229-10-6
IBPGR (1988) International Board for Plant Genetic Resources. Lists for Quantitative and Qualitative Measurements of Growth and Yield Attributes in Plants, Rome, Italy
International Institute for Tropical Agriculture (2007) Plantain development trials in Ghana. IITA Res Bull 4:12–16
Jarret RL, Gawel N (1995) Molecular markers, genetic diversity and systematics in Musa. In: Gowen S (ed) Bananas and Plantains. Chapman and Hall, London, UK, pp 66–83
Kaemmer D, Afza R, Weising K, Kahl G, Novak FJ (1992) Oligonucleotide and amplification fingerprinting of wild species and cultivars of banana (Musa spp.). Biotechnology 10:1030–1035
Lagoda PJL, Noyer JL, Dambier D, Baurens FC, Grapin A, Lanaud C (1998) Sequence tagged microsatellite site (STMS) markers in Musaceae. Mol Ecol 7:657–666
Nei M (1978) Estimation of average heterozygosity and genetic distance from a small number of individuals. Genetics 89:583–590
Odeigah PGC (1997) “Double bunching” in a plantain germplasm in Nigeria: a new genotype. Genet Resour Crop Evol 44:1–3
Ogazi PO (1996) Plantain: production, processing, utilization pp.305. Paman and Associates Limited, Uku–Okigwe
Pennisi E (2010) Armed and dangerous. Science 327:804–805
Perrier X, Jacquemoud-Collet JP (2006) DARwin software https://darwin.cirad.fr/
Phillipe K, Ambang Z, Begoude D, Ten HM, KuataAmougou JA (2012) Evaluation of the sensitivity of two plantain varieties essong and big ebanga to the nematode Radopholus similis. Afr J Biotech 11:14755–14760
Saghai-Maroof MA, Soliman KM, Jorgenson RA, Allard RW (1984) Ribosomal DNA spacer length polymorphism in barley: Mendelian inheritance, chromosomal location and population dynamics. Proc Natl Acad Sci USA 81:8014–8018. https://doi.org/10.1073/pnas.81.24.8014
Ssebuliba R, Vuylsteke D, Hartman J, Makumbi D, Talengera D, Rubaihayo P, Magambo S, Nuwagala L, Nahamya P, Karamura E (2000) Towards improving highland bananas. Uganda J Agric Sci 5:36–38
Swennen R (1997) Plantain cultivation under West African conditions: a reference manual. International Institute for Tropical Agriculture, Ibadan, Nigeria, Amarin Printing Group Co. Ltd Thailand 24 p
Tenkouano A (2000) Persistence and horticultural value of inflorescence dichotomy in plantain. HortScience 35:933–936
Tezanas du Montcel H (1987) Plantain and bananas. Macmillan Publisher Ltd, London, UK, pp 4–6
Vuylsteke DR, Swennen RL, De Langhe EA (1996) Field performance of somaclonal variants of plantain (Musa spp., AAB Group). J Am Soc Hort Sci 121:42–46
Weising K, Nybom H, Wolff K, Kahl G (2005) DNA Fingerprinting in plants: principles, methods, and applications, 2nd edn. CRC Press, London
Author information
Authors and Affiliations
Contributions
EAB conceived the study, interpreted the data, and wrote the manuscript; MGU designed and performed experiments, and identified and maintained elite cultivars based on agronomic data.
Corresponding author
Ethics declarations
Conflict of interest
It is declared that there is no conflict of interest associated with the publication of this paper.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
About this article

Cite this article
Brisibe, E.A., Ubi, G.M. Microsatellite fingerprinting and analysis of intra-population divergence in morphotaxonomic traits in a large Musa (AAB genome) germplasm. Physiol Mol Biol Plants 26, 1973–1988 (2020). https://doi.org/10.1007/s12298-020-00877-0
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s12298-020-00877-0