
Abstract
This study was designed to investigate the effects of amlodipine (AM), a dihydropyridine calcium channel blocker, on the oxidative damage induced by diazinon (DZN) in the rat cortex and cerebellum. Forty-two rats were randomly divided into six groups. The rats were treated intraperitoneally with normal saline (group 1), AM (9 mg/kg; group 2), DZN (32 mg/kg; group 3) and different doses of AM (3, 6, and 9 mg/kg; groups 4, 5, and 6, respectively) with DZN. After 14 days, the cerebellum and cortex tissues were removed for biochemical and histological experiments. DZN significantly decreased acetylcholinesterase activity (AChE; 57%, p < 0.001 and 39.1%, p < 0.05), depleted total antioxidant capacity (TAC; 46.2%, p < 0.01 and 44.7%, p < 0.05), and increased lactate dehydrogenase activity (LDH; 96%, p < 0.001 and 202%, p < 0.001), nitric oxide (NO; 130%, p < 0.001 and 74.4%, p < 0.001), and lipid peroxidation levels (LPO; 35.6%, p < 0.001 and 128.7%, p < 0.001), in the cerebellum and cortex tissues, respectively. In addition, DZN induced structural alterations in the cerebellum and cortex. Following AM administration, a remarkable improvement was observed in LDH activity and some of the oxidative markers, such as NO and LPO; however, no significant changes were found in AChE activity when the DZN group was compared with the AM-treated groups. This study suggests that AM may prevent DZN-induced neurotoxicity via improvement of the oxidative/antioxidant balance in the cerebellum and cortex tissues.
Similar content being viewed by others
Avoid common mistakes on your manuscript.
Introduction
The cerebellum and cortex are the most effective parts of the brain involved in motor and balance regulation. The cerebral cortex is separated into four different lobes—the frontal, parietal, temporal, and occipital—each of which are responsible for processing various types of sensory information [1]. In addition, studies have demonstrated that the cerebellum is a simple coordinator of somatic motor and autonomic function. Cerebral cortex involvement in a broad spectrum of functions and disorders such as language, learning, memory, attention-deficit/hyperactivity disorder, schizophrenia, and autism has been indicated [2,3,4].
Organophosphorus compounds (OPCs) are the most commonly used pesticides in the world. Several studies have shown that exposure with these compounds can be associated with some neurological diseases such as neurobehavioral disorder and Alzheimer’s disease [5, 6]. For instance, monocrotophos can exert its neurobehavioral disorder by affecting noncholinergic functions involving serotonergic and dopaminergic systems related with enhanced oxidative stress in cerebellum and cortical regions [7].
Among the known OPCs, diazinon (DZN) (O,O-diethyl O (2-isopropyl-6-methyl-4-pyridinyl phosphorothioate) is a synthetic organophosphate that is widely used in agriculture [8]. The key mechanism of acute toxication with DZN, like other OPCs, is irreversible inhibition of acetylcholinesterase (AChE) activity in the synaptic terminals of the nervous [9]. In addition, the occurrence of oxidative stress and mitochondrial dysfunction as a result of chronic exposure to OPCs is associated with neurodegenerative disorders and neurobehavioral deficits [10].
Under pathologic conditions, increase in intracellular calcium leads to the activation of proteolytic enzymes and nitric oxide synthase, as well as the production of free radicals [11, 12]. These agents can react with cellular macromolecules and thereby lead to oxidative system damage in DNA, proteins, and lipids [13]. Several studies have confirmed the antioxidant effects of dihydropyridine calcium channel blockers under in vitro and in vivo conditions [10, 14, 15]. Amlodipine (AM) is a lipophilic dihydropyridine calcium channel blocker that inhibits L-type calcium channels in various organs, including the brain [16].
Based on existing evidence, the present study was designed to evaluate the effects of AM on oxidative damage induced by DZN in the cortex and cerebellum tissue of Wistar rats.
Materials and Methods
Animals
Forty-Two Wistar male rats (200–250 g), were obtained from the animal house of Hamadan University of Medical Science (HUMS). The rats were quarantined in a room with an ambient temperature of 22–25 °C and a 12:12 light/dark cycle and fed pellet diet and water ad libitum for 1 week prior to use. The study protocol was approved by HUMS Ethics Committee with the code number of IR.UMSHA.REC.1394.509.
Experimental Design
The rats were divided randomly into six groups of seven. Animals were treated intraperitoneally for fourteen consecutive days as follows: Group 1 (negative control) received normal saline. Group 2 received 9 mg/kg of AM. Group 3 (positive control) received 32 mg/kg DZN, and groups 4–6 were treated with 3, 6, and 9 mg/kg of AM, respectively, 1 h before DZN (32 mg/kg) administration. 24 h after treatment, animals were anesthetized by diethyl ether and perfused transcardially with 100 ml of 0.9% sodium chloride. After surgery, the rats’ brains were removed for isolation of cortex and cerebellum tissues [17]. A portion of these tissues was homogenized in tris buffer (pH 7.4). The tissue homogenate (10%, w/v) were centrifuged at 3000g for 10 min at 4 °C. Then, the supernatants were removed and kept at −80 °C until the biochemical examinations.
Acetylcholinesterase Assay
The AChE activity was assayed by measuring the yellow color generated from the reaction between Ellman’s reagent and the thiocholine obtained from the hydrolysis of acetylthiocholine iodide, at 412 nm using a microplate reader (Synergy HTX, Biotek, USA). The results were expressed as percent of control [18].
Lipid Peroxidation Assay
The lipid peroxidation (LPO) level was measured by thiobarbituric acid reactive substances (TBARS) method. Briefly, tissue homogenate were mixed with TBA (0.2%) in H2SO4 (0.05 M) and heated in a boiling water bath for 30 min. Thiobarbituric acid-reactive substances were extracted by n-butanol and its absorbance was assayed at 532 nm. The results were expressed as nmol/mg protein [19].
Total Antioxidant Capacity Assay
Total antioxidant capacity (TAC) was assayed by measuring their ability to reduce Fe3+ to Fe2+. The complex Fe2+-TPTZ gives a blue color with a absorbance at 593 nm. The results were expressed as nmol/mg protein [20].
Nitric Oxide Assay
Tissue nitric oxide (NO) was determined using Griess reagent (1% sulfanilamide, 0.1% NED, and 2.5% phosphoric acid). The results were expressed as µmol/mg protein [10].
Lactate Dehydrogenase Assay
Lactate dehydrogenase (LDH) activity was assayed by determining the rate of oxidation of NADH using an enzymatic colorimetric kit (Pars Azmoon Co., Tehran, Iran), at 340 nm. The results were expressed as U/mg protein.
Protein Assay
Protein content was determined in the crude homogenate of liver according to Bradford method using bovine serum albumin (BSA) as a standard.
Histopathological Examination
After cardiac perfusion, the animals were decapitated and then the brain was removed. The isolated tissues were hold in 4% formaldehyde at 25 °C for at least 2 h to be extremely fixed. The paraffin-embedded block was prepared by automatic tissue processor and sectioned to a thickness of 4–µm. Finally, the tissues were stained with Hematoxylin and Eosin (H&E) dye and were examined for histopathological changes [17].
Data Analysis
The data are expressed as the mean ± standard deviation (SD) and analyzed by Graph Pad Prism software, version 6.0. The statistical difference of the values was determined by one-way analysis of variance (ANOVA) with Tukey’s test for multiple comparisons. A p value of less than 0.05 was considered statistically significant.
Results
Effect of Amlodipine on AChE Activity in Cortex and Cerebellum
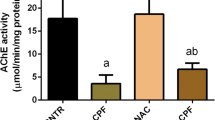
As shown in Fig. 1, a significant decrease in AChE activity was observed in DZN-exposed groups compared to the control group (cortex: 57%, p < 0.001 and cerebellum: 39.1%, p < 0.05). However, no significant changes were found in this marker when DZN group was compared with AM-treated groups.

Effects of AM on AChE activity in cortex and cerebellum tissues in male rats. Results are expressed as mean ± SD, N = 7 for each group. ≠≠≠p < 0.001, and ≠≠p < 0.01 versus control group (cortex); +p < 0.05 versus control group (cerebellum). DZN diazinon, AM amlodipine, AChE acetylcholinesterase
Effect of Amlodipine on LPO Level in Cortex and Cerebellum
As shown in Fig. 2, LPO level in cortex and cerebellum significantly increased after administration of DZN compared to control group (35.6%, p < 0.001 and 128.7%, p < 0.001, respectively). All doses of AM were able to decrease LPO level in the cortex tissue (0.36%, p < 0.001; 26.2%, p < 0.001 at doses 3, 6 mg/kg and 22.1%, p < 0.01 at dose 9 mg/kg) compared to DZN-exposed rats. In addition, AM administration at doses of 6 and 9 mg/kg significantly improved LPO level of cerebellum in comparison with DZN group (41.2%, p < 0.01 and 37.9%, p < 0.01, respectively).

Effects of AM on LPO level in cortex and cerebellum of male rats. Results are expressed as mean ± SD, N = 7 for each group. +++p < 0.001 versus control group (cortex). ≠≠≠p < 0.001 versus control group (cerebellum). aap < 0.01, aaap < 0.001 versus DZN group (cortex). bbp < 0.01 versus DZN group (cerebellum). DZN diazinon, AM amlodipine, LPO lipid peroxidation
Effect of Amlodipine on TAC in Cortex and Cerebellum
As shown in Fig. 3, DZN significantly reduced TAC of the cortex (46.2%, p < 0.01) and cerebellum (44.7%, p < 0.05). Administration of AM at the dose of 6 mg/kg significantly improved TAC level in cortex (46.8%, p < 0.05) and cerebellum (48.7%, p < 0.05) tissues, while in other doses (3 and 9 mg/kg) changes were not significant.

Effects of AM on TAC level in cortex and cerebellum of male rats. Results are expressed as mean ± SD, N = 7 for each group. ≠≠p < 0.01 versus control group (cortex). +p < 0.05 versus control group (cerebellum). ap < 0.05 versus DZN group (cortex). In cerebellum tissue; bp < 0.01 versus DZN group (cerebellum). DZN diazinon, AM amlodipine, TAC total antioxidant capacity
Effect of Amlodipine On NO Level in Cortex and Cerebellum
As shown in Fig. 4, administration of DZN causes a remarkable increase in NO level compared to the control group in cortex (130%, p < 0.001) and cerebellum (74.4%, p < 0.001). Following AM administration, a significant decrease was observed in cortex NO level in doses of 6 mg/kg (39%, p < 0.01) and 9 mg/kg (32%, p < 0.05) of AM compared to the DZN group. AM treatment at dose of 6 mg/kg significantly prevented NO production in cerebellum tissue (38%, p < 0.05).

Effects of AM on NO level in cortex and cerebellum tissues of male rats. Results are expressed as mean ± SD, N = 7 for each group. +++p < 0.001 versus control group (cortex). ≠≠≠p < 0.001 versus control group (cerebellum). ap < 0.05, aap < 0.01 versus DZN group (cortex). bp < 0.05 versus DZN group (cerebellum). DZN diazinon, AM amlodipine, NO nitric oxide
Effect of Amlodipine on LDH Activity in Cortex and Cerebellum
As shown in Fig. 5, DZN causes a remarkable increase in LDH activity of cortex (96%, p < 0.001), and cerebellum (202%, p < 0.001) compared to the control group. Following AM administration, a significant decrease was observed in its enzyme activity in treatment groups compared to the DZN group.

Effects of AM on LDH activity in cortex and cerebellum tissues of male rats. Results are expressed as mean ± SD, N = 7 for each group. +++p < 0.001 versus control group (cortex); ≠≠≠p < 0.001 versus control group (cerebellum). ap < 0.05, aap < 0.01 versus DZN group (cortex). bp < 0.05 versus DZN group (cerebellum). DZN: diazinon, AM amlodipine, LDH lactate dehydrogenase
The Effects of Amlodipine on Histopathological Changes
The histopathological examinations showed some pathological alterations in cortex and cerebellum tissues, including vascular congestion, lymphocytes accumulation, vascular and glial cells alteration, in DZN group. AM prevented some of the histopathological changes of DZN such as decrease the number of lymphocytes and glial cells changes in these tissues (Table 1 and Fig. 6).

Photomicrographs of rat cortex and cerebellum tissues in different groups. For cortex tissue: a negative control group, b AM (9 mg/kg) group, c DZN (32 mg/kg) group, d DZN (32 mg/kg) + AM (3 mg/kg) group, e DZN (32 mg/kg) + AM (6 mg/kg) group, and f DZN (32 mg/kg) + AM (9 mg/kg) group. For cerebellum tissue: g negative control group, h AM (9 mg/kg) group, i DZN (32 mg/kg) group, j DZN (32 mg/kg) + AM (3 mg/kg) group, k DZN (32 mg/kg) + AM (6 mg/kg) group, and l DZN (32 mg/kg) + AM (9 mg/kg) group. Original magnification of all images is × 40. DZN diazinon, AM amlodipine
Discussion
In the current study, we showed that exposure to DZN (32 mg/kg) induced significant changes in oxidant/antioxidant balance and also cellular architecture, indicating the early involvement of the cortex and cerebellum in DZN toxicity. In addition, these findings explained the role of AM in improving oxidative damage induced by DZN.
It is well known that the main mechanism of toxicity of OPCs is inhibition of the enzyme AChE and subsequent accumulation of the neurotransmitter acetylcholine in the synaptic terminals of nervous system [8, 21, 22]. Therefore, significant reduction of AChE activity in the cortex and cerebellum tissues indicates the occurrence of neurotoxicity in DZN-exposed groups.
In this study, we showed that DZN could induce significant changes in LPO, index of oxidative degradation of lipids, which indicates increases in reactive radicals in cortex and cerebellum tissue. Overall, the brain tissue exhibits a high level of oxidative metabolism and polyunsaturated fatty acids [23], and consumes about 20% of the cardiac output [24]. Therefore, this tissue is particularly sensitive to oxidative damage caused by OPCs that its vulnerability may be different in various areas of the brain tissue. As our results shows, we found that the LPO level in the cerebellum was significantly higher than that in cortex tissue. Afolabi et al. [25] have shown that the level of polyunsaturated fatty acids in the cerebellum is higher than that in cortex tissue. Therefore, the difference in LPO levels between these two tissues may be associated with high levels of polyunsaturated fatty acids in cerebellum tissue.
The present study showed that DZN decreased the TAC content in the cortex and cerebellum. In agreement with our findings, previous studies have shown that OPCs altered the brain levels of antioxidative agents such as antioxidant enzymes, including glutathione peroxidase (GPx), catalase (CAT), and copper-zinc superoxide dismutase (Cu/Zn SOD), in animal models [26, 27]. The activity of these enzymes could be related with the alteration of antioxidant capacity in different areas of the brain.
Following the administration of AM, the LPO and TAC levels were significantly improved in animals exposed to DZN, and the best response was observed with the AM dose of 6 mg/kg. The antioxidant properties of AM could be associated with its chemical structure, which facilitates proton-donating and resonance-stabilization mechanisms that, in turn, inhibit the oxidative stress process [10, 28]. In addition, AM-antioxidative effects can be attributed to the preservation of superoxide dismutase activity [28].
In OPCs-induced toxicity, excessive influx of calcium due to overstimulation of muscarinic acetylcholine receptors (mAChRs) and/or N-methyl-d-aspartate (NMDA) receptors could be considered as an important cause of neuronal cell death [29]. Moreover, intracellular calcium overload leads to excessive oxidative stress and inflammatory responses [30]. Hirooka et al. [31] showed that AM administration could be effective in improvement of oxidative damage on stroke-prone spontaneously hypertensive rats. Also, Sendrowski et al. [32] reported that both T-type and L-type calcium channel blockers had protective effects against glutamate toxicity in cultured hippocampal neurons. Therefore, in addition to the antioxidative properties of AM, it also controlled the influx of calcium into the neuron cells and prevented oxidative injury in the cortex and cerebellum tissue.
As our data shows, DZN caused an increased in NO content of cerebellum and cortex. NO, a key signaling molecule, is synthesized from l-arginine by enzyme nitric oxide synthase (NOS) which exists in different cells [33]. In nitrosative stress, the high levels of NO is produced by the increased expression of NOS in some of the nerve cells such as astrocytes and microglia cells following their stimulation via inflammatory cytokines and/or reactive oxygen species. NO can react with the superoxide anion, resulting in the generation of the highly reactive peroxynitrite anion (ONOO−) [34]. The role of nitrosative stress in the development of OPCs neurotoxicity is confirmed through inhibition of endgenous NO generation by the selective neuronal NOS inhibitor or the non-selective NOS. They were able to reduce brain oxidative stress caused by acute toxicity of malathion in animal model [35]. Our findings show that treatment with AM results in decreased brain NO levels, which could indicate its neuroprotective effects on cerebellum and cortex.
In current study, a significant increase in LDH activity was observed in cerebellum and cortex tissues. LDH is an index of anaerobic metabolism that may also be the cause oxidative damage induced by DZN in cortex and cerebellum tissues. Previously, Nili-Ahmadabadi et al. [10] reported an increase in LDH activity in hippocampus tissue following OPCs toxication. It has been observed that metabolic modifications can occur after exposure to OPCs, such as interference with the glycolytic pathway and the tricarboxylic acid cycle (TCA), and also alterations in cellular mitochondrial respiration [36]. Consequently, mitochondrial dysfunction may cause aerobic metabolism disruption and notably enhance oxidative damage [37]. Calcium regulation plays a key role in cellular mitochondrial function. Mamou et al. [38] showed that AM improved mitochondrial function and preserved its structure in ventricular cardiomyocytes during ischemia–reperfusion in the pig. Therefore, it seems that AM can prevent increase in LDH activity in the cortex and cerebellum of DZN-exposed animals; this further demonstrates its role in intracellular calcium regulation, and consequently, improving mitochondrial function.
Histopathological alterations in various groups confirm our biochemical findings. In microscopic examination, significant changes in cellular architecture were observed in the cortex and cerebellum of DZN-treated rats that improved following administration of AM.
Conclusion
Taken together, these findings show that AM exerts its neuroprotective effects after DZN toxicity through improvement of cellular aerobic respiration and oxidant/antioxidant balance. These therapeutic effects may be associated with the role of calcium channel blockers in intracellular calcium regulation and the antioxidant properties of AM. In addition, AM may play a key role in preventing the neurotoxic effects of OPCs through inhibition of nitrosative stress in brain cells. Our findings could shed light on the management of the neurotoxic effects of OPCs in exposed patients.
References
Gudienė D, Burba B. Mental disorders and their relation to brain lesion location: diagnostical problems. Medicina (Kaunas). 2003;39:114–21.
Bugeon S, Lafi S, Beurrier C, Sahu S, Runge K, Schaller F, et al. Morphofunctional deficits in the cerebral cortex of NeuroD2 mutant mice are associated with autism/schizophrenia-like behaviors. BioRxiv: 2018:296889.
Fenlon LR, Liu S, Gobius I, Kurniawan ND, Murphy S, Moldrich RX, et al. Formation of functional areas in the cerebral cortex is disrupted in a mouse model of autism spectrum disorder. Neural Dev. 2015;10:10.
Herzfeld DJ, Pastor D, Haith AM, Rossetti Y, Shadmehr R, O’shea J. Contributions of the cerebellum and the motor cortex to acquisition and retention of motor memories. Neuroimage. 2014;98:147–58.
Sankhwar ML, Yadav RS, Shukla RK, Pant AB, Singh D, Parmar D, et al. Impaired cholinergic mechanisms following exposure to monocrotophos in young rats. Hum Exp Toxicol. 2012;31:606–16.
Yadav S, Singh M, Yadav R. Organophosphates induced Alzheimer’s disease: an epigenetic aspect. J Clin Epigenet. 2016;2:1158–2472.
Sankhwar ML, Yadav RS, Shukla RK, Singh D, Ansari RW, Pant AB, et al. Monocrotophos induced oxidative stress and alterations in brain dopamine and serotonin receptors in young rats. Toxicol Ind Health. 2016;32:422–36.
Nili-Ahmadabadi A, Alibolandi P, Ranjbar A, Mousavi L, Nili-Ahmadabadi H, Larki-Harchegani A, et al. Thymoquinone attenuates hepatotoxicity and oxidative damage caused by diazinon: an in vivo study. Res Pharm Sci. 2018;13:500–8.
Hassani S, Maqbool F, Salek-Maghsoudi A, Rahmani S, Shadboorestan A, Nili-Ahmadabadi A, et al. Alteration of hepatocellular antioxidant gene expression pattern and biomarkers of oxidative damage in diazinon-induced acute toxicity in Wistar rat: a time-course mechanistic study. EXCLI J. 2018;17:57–71.
Nili-Ahmadabadi A, Ali-Heidar F, Ranjbar A, Mousavi L, Ahmadimoghaddam D, Larki-Harchegani A, et al. Protective effect of amlodipine on diazinon-induced changes on oxidative/antioxidant balance in rat hippocampus. Res Pharm Sci. 2018;13:368–76.
Magi S, Castaldo P, Macrì ML, Maiolino M, Matteucci A, Bastioli G, et al. Intracellular calcium dysregulation: implications for Alzheimer’s disease. Biomed Res Int 2016. https://doi.org/10.1155/2016/6701324.
Saternos HC, AbouAlaiwi WA. Signaling interplay between primary cilia and nitric oxide: a mini review. Nitric Oxide 2018;80:108–12.
Nili-Ahmadabadi A, Pourkhalili N, Fouladdel S, Pakzad M, Mostafalou S, Hassani S, et al. On the biochemical and molecular mechanisms by which malathion induces dysfunction in pancreatic islets in vivo and in vitro. Pestic Biochem Physiol. 2013;106:51–60.
Pronobesh C, Dagagi A, Pallab C, Kumar W. Protective role of the calcium channel blocker amlodipine against mitochondrial injury in ischemia and reperfusion injury of rat liver. Acta Pharm. 2008;58:421–8.
Fares H, DiNicolantonio JJ, O’Keefe JH, Lavie CJ. Amlodipine in hypertension: a first-line agent with efficacy for improving blood pressure and patient outcomes. Open Heart. 2016;3:e000473.
Ritz B, Rhodes SL, Qian L, Schernhammer E, Olsen JH, Friis S. L-type calcium channel blockers and Parkinson disease in Denmark. Ann Neurol. 2010;67:600–6.
Zhao Q, Wang X, Chen A, Cheng X, Zhang G, Sun J, et al. Rhein protects against cerebral ischemic-/reperfusion-induced oxidative stress and apoptosis in rats. Int J Mol Med. 2018;41:2802–12.
Ozarowski M, Mikolajczak PL, Piasecka A, Kujawski R, Bartkowiak-Wieczorek J, Bogacz A, et al. Effect of Salvia miltiorrhiza root extract on brain acetylcholinesterase and butyrylcholinesterase activities, their mRNA levels and memory evaluation in rats. Physiol Behav. 2017;173:223–30.
Nili-Ahmadabadi A, Tavakoli F, Hasanzadeh G, Rahimi H, Sabzevari O. Protective effect of pretreatment with thymoquinone against Aflatoxin B1 induced liver toxicity in mice. Daru. 2011;19:282–7.
Zeinvand-Lorestani H, Nili-Ahmadabadi A, Balak F, Hasanzadeh G, Sabzevari O. Protective role of thymoquinone against paraquat-induced hepatotoxicity in mice. Pestic Biochem Physiol. 2018;148:16–21.
Judge SJ, Savy CY, Campbell M, Dodds R, Gomes LK, Laws G, et al. Mechanism for the acute effects of organophosphate pesticides on the adult 5-HT system. Chem Biol Interact. 2016;245:82–9.
Yadav RS. Biomarkers: an essential gizmo in pesticide toxicity. Biomarkers. 2016;2:9.
Garbarino VR, Orr ME, Rodriguez KA, Buffenstein R. Mechanisms of oxidative stress resistance in the brain: lessons learned from hypoxia tolerant extremophilic vertebrates. Arch Biochem Biophys. 2015;576:8–16.
Fischer GW, Torrillo TM, Weiner MM, Rosenblatt MA. The use of cerebral oximetry as a monitor of the adequacy of cerebral perfusion in a patient undergoing shoulder surgery in the beach chair position. Pain Pract. 2009;9:304–7.
Afolabi O, Sulaiman O, Adeleke G, Wusu D. Acetylcholinesterase activity and oxidative stress indices in cerebellum, cortex and hippocampus of rats exposed to lead and manganese. Int J Bio Res. 2016;4:157–64.
Kazi AI, Oommen A. Monocrotophos induced oxidative damage associates with severe acetylcholinesterase inhibition in rat brain. Neurotoxicology. 2012;33:156–61.
Cemek M, Büyükokuroğlu ME, Büyükben A, Aymelek F, Özcan L. Effects of vitamin E and selenium on tissue bio-element status in organophosphate toxicity of rats. Pestic Biochem Physiol. 2010;98:9–18.
Godfraind T, Salomone S. Ambiguities in dietary antioxidant supplementation compared to calcium channel blockers therapy. Front Pharmacol. 2015;6:10.
Chen Y. Organophosphate-induced brain damage: mechanisms, neuropsychiatric and neurological consequences, and potential therapeutic strategies. Neurotoxicology. 2012;33:391–400.
Liu Z, Zhou T, Ziegler AC, Dimitrion P, Zuo L. Oxidative stress in neurodegenerative diseases: from molecular mechanisms to clinical applications. Oxid Med Cell Longev. 2017;2017:2525967.
Hirooka Y, Kimura Y, Nozoe M, Sagara Y, Ito K, Sunagawa K. Amlodipine-induced reduction of oxidative stress in the brain is associated with sympatho-inhibitory effects in stroke-prone spontaneously hypertensive rats. Hypertens Res. 2006;29:49–56.
Sendrowski K, Rusak M, Sobaniec P, Ilendo E, Dabrowska M, Bockowski L, et al. Study of the protective effect of calcium channel blockers against neuronal damage induced by glutamate in cultured hippocampal neurons. Pharmacol Rep. 2013;65:730–6.
Förstermann U, Sessa WC. Nitric oxide synthases: regulation and function. Eur Heart J. 2011;33:829–37.
Moylan S, Berk M, Dean OM, Samuni Y, Williams LJ, O’neil A, et al. Oxidative and nitrosative stress in depression: why so much stress? Neurosci Biobehav Rev. 2014;45:46–62.
Abdel-Salam OM, Youness ER, Mohammed NA, Yassen NN, Khadrawy YA, El-Toukhy SE, et al. Nitric oxide synthase inhibitors protect against brain and liver damage caused by acute malathion intoxication. Asian Pac J Trop Med. 2017;10:773–86.
Pearson JN, Patel M. The role of oxidative stress in organophosphate and nerve agent toxicity. Ann N Y Acad Sci. 2016;1378:17–24.
Karami-Mohajeri S, Abdollahi M. Mitochondrial dysfunction and organophosphorus compounds. Toxicol Appl Pharmacol. 2013;270:39–44.
Mamou Z, Chahine M, Rhondali O, Dehina L, Chevalier P, Descotes J, et al. Effects of amlodipine and perindoprilate on the structure and function of mitochondria in ventricular cardiomyocytes during ischemia-reperfusion in the pig. Fundam Clin Pharmacol. 2015;29:21–30.
Acknowledgements
This work was financially supported (Grant No. 9412187262) by the Vice-chancellor of Research and Technology, Hamadan University of Medical Sciences, Hamadan, Iran.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
The authors declare that there is no conflict of interest.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
About this article

Cite this article
Ataei, S., Abaspanah, S., Haddadi, R. et al. Therapeutic Potential of Dihydropyridine Calcium Channel Blockers on Oxidative Injury Caused by Organophosphates in Cortex and Cerebellum: An In Vivo Study. Ind J Clin Biochem 35, 339–346 (2020). https://doi.org/10.1007/s12291-019-00830-3
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s12291-019-00830-3