
Abstract
Chronic obstructive pulmonary disease (COPD) is the leading cause of human death worldwide. Currently available therapies for COPD mainly relieve symptoms and preserve lung function, suggesting the need to develop novel therapeutic or preventive regimens. Because chronic inflammation is a mechanism of emphysematous lesion formation and because adenosine A3 receptor signaling and peroxisome proliferator-activated receptor gamma (PPARγ) regulate inflammation, we investigated the effect of LJ-529, a selective adenosine A3 receptor agonist and partial PPARγ agonist, on inflammation in vitro and elastase-induced pulmonary emphysema in vivo. LJ-529 markedly ameliorated elastase-induced emphysematous lesion formation in the lungs in vivo, as indicated by the restoration of pulmonary function, suppression of airspace enlargement, and downregulation of elastase-induced matrix metalloproteinase activity and apoptotic cell death in the lungs. LJ-529 induced the expression of PPARγ target genes, the activity of PPARγ and several cytokines involved in inhibiting inflammation and inducing anti-inflammatory M2-like phenotypes. Moreover, LJ-529 did not exhibit significant cytotoxicity in normal cell lines derived from various organs in vitro and induced minimal changes in body weight in vivo, suggesting no overt toxicity of LJ-529 in vitro or in vivo. These results indicate the potential of LJ-529 as a novel therapeutic/preventive agent for emphysema with limited toxicity.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Chronic obstructive pulmonary disease (COPD) is one of the leading causes of human death worldwide, and the socioeconomic burden of COPD is expected to gradually increase (Lozano et al. 2012; Guarascio et al. 2013; Barnes et al. 2015; Tachkov et al. 2017). COPD is characterized by chronic destructive changes in the lungs that cause gas exchange disturbance, and chronic bronchitis and emphysema are the main symptoms of COPD (Barnes et al. 2015; Van Tho et al. 2015). In patients with emphysema, the alveolar walls are irreversibly destroyed, and the airspace is abnormally enlarged (Horio et al. 2017). Several factors, such as α1-antitrypsin deficiency, oxidative stress, aging, and chronic inflammation, are involved in the pathogenesis of emphysema (Gooptu et al. 2009; Barnes et al. 2015). Several exogenous insults, including smoking and air pollutants, activate macrophages and neutrophils and produce reactive oxygen species and various proteases, leading to tissue damage (Sharafkhaneh et al. 2008; Barnes et al. 2015). Elastin fragments produced by matrix degradation cause chemoattraction and autoimmunity, leading to the further enhancement of pulmonary inflammation (Lee et al. 2007; Sharafkhaneh et al. 2008). Inhaled bronchodilators, anticholinergics, β2-adrenergic agonists, corticosteroids, antibiotics and antioxidants have been used for to relieve symptoms, including respiratory difficulty, and preserve lung function for the treatment of emphysema/COPD (Celli 2018). Although these regimens can alleviate disease progression and cause clinically significant benefits for patients with COPD (Celli 2018), it is difficult to completely cure COPD/emphysema using these agents in most cases. Therefore, novel therapeutics for the treatment or prevention of emphysema are urgently needed.
Previous reports have suggested the association of adenosine signaling with the regulation of inflammation and pulmonary homeostasis and its implication in chronic pulmonary disorders such as asthma and COPD (Zhou et al. 2009; Haskó and Cronstein 2013). Adenosine binds to one of four G protein-coupled adenosine receptors (A1AR, A2AAR, A2BAR, and A3AR). Of these, the adenosine A3 receptor (A3AR) is associated with the regulation of ischemia, inflammation, and tumor growth and is considered a target for the development of therapeutic agents (Borea et al. 2015). We have developed potent and selective agonists and antagonists of A3AR (Jeong et al. 2003, 2006, 2008).
Previous studies have demonstrated the role of peroxisome proliferator-activated receptor (PPAR) signaling in the regulation of inflammation (Daynes and Jones 2002). Among the isoforms of PPAR (PPARα, PPARβ/δ, and PPARγ), PPARγ plays an important role in the regulation of gene expression linked to several pathologic conditions (Tontonoz and Spiegelman 2008). PPARγ agonists exert anti-inflammatory actions by suppressing the production of pro-inflammatory cytokines in macrophages (Tontonoz and Spiegelman 2008) and inducing the polarization of monocytes to the M2 phenotype (Bouhlel et al. 2007) and differentiation into regulatory T cells (Housley et al. 2009). The modulation of the transcriptional activity of nuclear factor-kappa B (NF-κB) is regarded as the mechanism underlying the anti-inflammatory role of PPARγ (Kapadia et al. 2008; Remels et al. 2009). Notably, agonistic PPARγ activation ameliorates pulmonary emphysema in experimental models by suppressing inflammation, inhibiting pathogenic lung dendritic cells and chemokine expression, inducing the polarization of macrophages to the M2 phenotype, and clearing apoptotic neutrophils (Lea et al. 2014; Shan et al. 2014; Solleti et al. 2015). Therefore, the activation of PPARγ may be an effective strategy for controlling COPD/emphysema. To discover novel potential compounds that suppress the development of emphysema, we employed a large chemical library consisting of compounds from various classes. According to our previous report demonstrating the identification of LJ-529 [2-chloro-N6-(3-iodobenzyl)-5′-N-methylcarbomoyl-4′-thioadenosine] as a partial PPARγ agonist (Yu et al. 2017), here we investigated the inhibitory effect of LJ-529 on elastase-induced pulmonary emphysema. Our findings showed that LJ-529 significantly suppressed the elastase-induced deregulation of pulmonary function and lung destruction, matrix metalloproteinase activity, and apoptosis in the lungs. LJ-529 induced the expression of several PPARγ target genes and several cytokines involved in inhibiting inflammation and inducing anti-inflammatory M2-like phenotypes in macrophages in the murine lungs. Moreover, LJ-529 exhibited minimal cytotoxicity in several normal cell lines derived from various organs in vitro and induced no obvious toxic effects in vivo. These findings highlight the potential of LJ-529 as a novel agent for the treatment of patients with emphysema.
Materials and methods
Reagents
LJ-529 was synthesized as described previously (Jeong et al. 2003). Porcine pancreatic elastase (PPE), 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT), and other chemicals were purchased from Sigma-Aldrich (St. Louis, MO, USA) unless otherwise specified.
Cell culture
Human retinal pigment epithelial (RPE) cells were kindly provided by Dr. Jeong Hun Kim (Seoul National University, Seoul, Republic of Korea). The murine hippocampal neuronal cell line HT-22 was kindly provided by Dr. Dong Gyu Jo (Sungkyunkwan University, Suwon, Republic of Korea). Human bronchial epithelial (HBE) cells were kindly provided by Dr. John Minna (University of Texas Southwestern Medical Center, Dallas, TX, USA). The murine macrophage cell line RAW 264.7 was kindly provided by Dr. Sang Kook Lee (Seoul National University, Seoul, Republic of Korea). RPE, HT-22, and RAW 264.7 cells were cultured in DMEM supplemented with 10% FBS and antibiotics (all from Welgene, Inc., Gyeongssan-si, Republic of Korea). HBE cells were cultured in keratinocyte-SFM (Invitrogen, Grand Island, NY, USA) supplemented with 5 ng/ml recombinant epidermal growth factor (EGF), 50 μg/ml bovine pituitary extract, and antibiotics. The cells were incubated at 37 °C with 5% CO2 in a humidified atmosphere.
Cell viability assay
Cells (2 × 103 cells/well in 96-well plates) were treated with LJ-529 for three days. The cells were incubated with an MTT solution for 4 h at 37 °C. The formazan products were dissolved in DMSO, and the absorbance was measured at 570 nm. The data are presented as percentage of the control group.
Real-time PCR
Total RNA was isolated from frozen lung tissues using a phenol–chloroform extraction method, reverse-transcribed using a first-strand cDNA synthesis kit (TransGen Biotech, Beijing, China), and analyzed by real-time PCR using a SYBR Green-based qPCR master mix solution (Enzynomics, Daejeon, Republic of Korea) and gene-specific primers. The primer sequences used for the PCR analyses are listed in Table 1. The thermocycler conditions were as follows: preincubation at 95 °C for 15 min; 50 cycles of 95 °C for 10 s, 60 °C for 15 s, and 72 °C for 20 s; and melting curve analysis to determine reaction specificity. The quantification or relative mRNA expression was performed using the comparative cycle threshold (CT) method as described previously (Livak and Schmittgen 2001).
Animal experiment
The animal experiment was performed according to a protocol approved by the Seoul National University Institutional Animal Care and Use Committee. Mice were provided standard mouse chow and water ad libitum and housed in temperature- and humidity-controlled facilities under a 12-h light/12-h dark cycle. Eight-week-old FVB mice were administered vehicle [20% DMSO dissolved in sterile distilled water containing 20% polyethylene glycol (PEG)] or LJ-529 (50 μg/kg) by oral gavage 6 times per week for 6 weeks. One week after drug treatment, 0.25 units of PPE was intratracheally instilled into the lungs of the mice. Body weight changes were monitored during the treatment. Changes in pulmonary function in vehicle- and LJ-529-treated mice were analyzed using the FlexiVent system (Scireq, EMKA Technologies, Montreal, Canada) (Vanoirbeek et al. 2010) according to the protocol recommended by the manufacturer. The mice were euthanized by inhalation of an overdose of isoflurane, and the lungs were excised after perfusion with ice-cold PBS and then embedded in OCT compound (Sakura Finetek, USA, Inc., Torrance, CA, USA). Frozen blocks were used for further analyses. H&E-stained tissues were used to determine structural changes in the lungs and histological changes in the kidneys, liver, and brain. The quantification of structural changes was determined by calculating the mean linear intercept (MLI), as previously described (Dunnill 1962; Chen et al. 2010).
In situ zymography
Dried cryosections of the lungs were incubated with fluorescein-conjugated DQ-gelatin (Thermo Fisher Scientific, Carlsbad, CA, USA) diluted in low-gelling temperature agarose for 3 h at room temperature. Fluorescence was observed under a fluorescence microscope, and the sections were photographed.
TUNEL staining
TUNEL staining was performed using the TMR Red In Situ Cell Death Detection Kit (Roche Applied Science, Penzberg, Germany) according to the manufacturer’s provided protocol.
Immunofluorescence staining
Cryosections (8 μm) were prepared for immunofluorescent analysis. The sections were fixed in 4% paraformaldehyde (PFA) for 30 min followed by permeabilization in 0.2% triton X-100 for 15 min at room temperature. The slides were incubated with anti-CD206 antibody (Santa Cruz Biotechnology, Dallas, TX, USA) or anti-Arginase 1 antibody (Cell Signaling Technology, Danvers, MA, USA) at 4 °C overnight. After washing, sections were incubated with alexa flour-conjugated secondary antibody for 1 h at room temperature. The nuclei were stained with DAPI and analyzed by confocal microscope (LSM 700; Carl Zeiss Microscopy, Jena, Germany).
Luciferase reporter gene assays
Raw264.7 cells were seeded in a 24-well plate and transiently transfected with PPARγ, PPRE-luciferase and the CH110-Lac-Z plasmid vector using jetPRIME transfection reagent (Polyplus Transfection, Illkirch, France). Luciferase reporter gene assays were performed using the p.j.k Luciferase Assay Kit (PJK GmbH, Kleinbittersdorf, Germany). The PPARγ and PPRE-luciferase reporter plasmid vector were kindly provided by Professor Keon Wook Kang of Seoul National University (Seoul, Korea).
Migration assay
Migration assays were performed in a Boyden chamber (Corning, Inc., Corning, NY, USA) with 8-μm inserts coated with gelatin. MLE12 cells were seeded into the lower wells. After 24 h, Raw 264.7 cells were loaded into the upper wells. Cells were incubated for 20 h. After the incubation, the cells were fixed with 100% cold methanol and stained with hematoxylin solution. The number of stained cells was counted using a microscope.
Statistical analysis
The data are presented as the mean ± SD. Statistical significance of difference was determined using a two-tailed Student’s t-test or one-way analysis of variance with Dunnett’s multiple comparisons test using GraphPad Prism 8 (GraphPad Software, Inc., La Jolla, CA, USA). A P value less than 0.05 was considered statistically significant.
Results
Inhibitory effect of LJ-529 on elastase-induced pulmonary emphysema
Based on the inhibitory effect of PPARγ agonists on pulmonary emphysema (Lea et al. 2014; Shan et al. 2014), we examined the effect of LJ-529 on elastase-induced emphysema in mice (Mahadeva and Shapiro 2002). As shown in Fig. 1a, 1 week after LJ-529 treatment, porcine pancreatic elastase (PPE) was instilled into the murine lungs, and the mice were treated with LJ-529 for an additional 4 weeks. Because pulmonary dysfunction is a main characteristic of emphysema (Barnes et al. 2015), we examined whether treatment with LJ-529 prevents the PPE-mediated deregulation of lung function by determining the compliance and tissue elastance of the lungs (Papandrinopoulou et al. 2012). In PPE-instilled mice, a reduction in lung tissue elastance caused elevated lung compliance, and treatment with LJ-529 significantly restored the PPE-induced deregulation of lung function in mice (Fig. 1b). Consistently, microscopic analysis of the mean linear intercept (MLI) of H&E-stained lung tissues also showed that LJ-529 treatment significantly suppressed PPE-induced airspace enlargement (Fig. 1c). These results suggest that LJ-529 has the capacity to suppress elastase-induced pulmonary emphysema.

Suppression of elastase-induced emphysema by the oral administration of LJ-529. a Schematic diagram of the experimental schedule. b Inhibition of elastase-induced pulmonary dysfunction by treatment with LJ-529 (50 μg/kg). Changes in lung function were monitored using the FlexiVent system. c Alleviation of elastase-mediated pulmonary destruction in the lungs of LJ-529-treated mice. Right. Quantification of structural changes in the airspace in each treatment group. The bars represent the mean ± SD; *P < 0.05 and ***P < 0.001 compared with the PPE-treated group. PPE porcine pancreatic elastase, LJ LJ-529, MLI mean linear intercept
Inhibitory effect of LJ-529 on elastase-induced apoptotic death of alveolar epithelial cells
Based on the importance of the protease-antiprotease imbalance in the development of emphysema (Sharafkhaneh et al. 2008) and the activation of matrix metalloproteinases by elastase (Ferry et al. 1997), we next investigated the effects of LJ-529 on elastase-induced gelatinase activity. We observed a marked increase in gelatinase activity in the lungs of PPE-instilled mice, and treatment with LJ-529 significantly suppressed PPE-induced gelatinase activity (Fig. 2a). In addition, the PPE-induced increase in the number of TUNEL-positive cells in the lungs, an indicator of PPE-mediated lung damage, was also significantly attenuated by treatment with LJ-529 (Fig. 2b). These results indicate the suppressive effect of LJ-529 on emphysema formation.

Inhibitory effects of LJ-529 on elastase-induced increases in matrix metalloproteinase activity and apoptotic cell death in the lungs. a Attenuation of elastase-mediated matrix metalloproteinase (MMP) activity by treatment with LJ-529. MMP activity was determined by using fluorescein-conjugated DQ-gelatin. Right. Quantification of gelatinase activity versus that in vehicle-treated controls. b Inhibition of elastase-mediated apoptotic cell death in the lungs by treatment with LJ-529, as determined by TUNEL staining of lung tissue cryosections. Right. Quantification of the number of TUNEL-positive cells versus that in vehicle-treated controls. The bars represent the mean ± SD; *P < 0.05, **P < 0.01 and ***P < 0.001 versus the PPE-treated group. PPE porcine pancreatic elastase, LJ LJ-529
Induction of anti-inflammatory cytokines by treatment with LJ-529
Macrophages play an important role in the development of emphysema upon exposure to causative factors (Yamasaki and Eeden 2018). M1 macrophages are classical pro-inflammatory macrophages that produce high levels of pro-inflammatory cytokines and reactive oxygen and nitrogen species (Yamasaki and Eeden 2018). Macrophages polarized to the M2 type are known to mediate damage repair by secreting anti-inflammatory cytokines, such as IL4 and IL10 (Krzyszczyk et al. 2018). Therefore, we examined whether LJ-529 modulates the expression of several factors associated with the polarization of macrophages to the M2 type and the inhibition of inflammation under pro-inflammatory environments in vitro. To mimic in vivo experimental conditions, murine macrophage cells were stimulated with lipopolysaccharide (LPS), a potent activator of innate immunity (Raetz and Whitfield 2002) which is also known to induce elastase production in macrophages (Duc Dodon and Vogel 1985), in the presence or absence of LJ-529 (1 μM). In LPS-stimulated RAW 264.7 cells, the expression of anti-inflammatory cytokines such as Il4 and Il10 was significantly decreased, which was substantially restored by treatment with LJ-529 (Fig. 3a). The Tgfb1 mRNA expression was moderately increased upon stimulation with LPS, which was presumably due to context-dependent roles of TGF-β1 in innate immunity (Wahl 2007). Consistent with the effect on the expression of Il4 and Il10, treatment with LJ-529 significantly enhanced Tgfb1 expression in LPS-stimulated macrophage cells (Fig. 3a). Moreover, LJ-529 significantly ameliorated LPS-mediated downregulation of markers of M2 macrophages, including Arg1 (encodes arginase 1, an enzyme involved in nitric oxide metabolism (Yang and Ming 2014) and Mrc1 (encodes CD206 the mannose receptor C-type 1 present on the surface of M2 macrophages (Trombetta et al. 2018) (Fig. 3b). Consistent with in vitro results, we observed the transcription of Il4, Il10, and Tgfb1 was restored or enhanced in the lungs of LJ-529-treated mice compared with those of PPE-instilled mice (Fig. 3c). Moreover, LJ-529 treatment significantly alleviated elastase-induced downregulation of the mRNA levels of Arg1 and Mrc1 in the lungs (Fig. 3d). We also asked whether LJ-529 treatment affects M1 macrophage parameters including TNF-α, IL-1β, and iNOS. Unlike M2 macrophage parameters, M1 macrophage parameters did not show significant differences by LJ-529 in the lungs (Fig. 3e). These results suggest that the inhibitory effects of LJ-529 on PPE-induced emphysema development are mediated by its anti-inflammatory actions in M2 macrophages rather than regulating pro-inflammatory actions of M1 macrophages.

Upregulation of anti-inflammatory cytokines and a M2 macrophage-related marker in the lungs from LJ-529-treated mice. a–d Changes in the transcription of cytokines and markers of M2 macrophages in murine macrophages (a and b) and in the lungs (c and d) of each treatment group were determined by real-time PCR. e Changes in the M1 macrophage parameters in the lungs of each treatment group were determined by real-time PCR. f The recruitment of M2 macrophages in the lungs of each treatment group were determined by immunofluorescence analysis. Right. Quantification of the recruitment of CD206- or Arginase 1-positive cells in each treatment group versus vehicle-treated controls. g Changes of the macrophages migration were determined by migration assay. The bars represent the mean ± SD; *P < 0.05, **P < 0.01, and ***P < 0.001 versus the PPE-treated group. PPE porcine pancreatic elastase, LJ LJ-529
Next, we examined whether M2 macrophages are recruited by LJ-529 treatment in the lungs. Treatment of LJ-529 resulted in the enhanced recruitment of M2 macrophages, as determined by the increase of the CD206+ and Arg1+ cells in the lungs (Fig. 3f). Unexpectedly, we found that the recruitment of CD206-positive M2 macrophages is upregulated by elastase instillation in the lungs. According to previous studies, CD206 is generally expressed by dendritic cells, and lymphatic or endothelial cells as well as M2 macrophages (Haque et al. 2019). And also, CD206 positive cells can be enhanced in the elastase-induced extensive injuries as a result of autonomous resolution mechanism (Shibata et al. 2018; Yan et al. 2019). Both differentiation and recruitment of macrophages are likely responsible for PPE-induced pathologies and LJ-529-induced suppression against the pathologies. We next evaluated the role of LJ-529 in macrophage migration by using Boyden chambers. LPS-stimulated alveolar epithelial cells promoted macrophage migration. Moreover, LJ-529 treatment more significantly enhanced macrophage migration than LPS-stimulated epithelial cells (Fig. 3g). Collectively, these findings suggest that LJ-529 exhibit anti-inflammatory actions by inducing recruitment of macrophages and their differentiation into M2 in an inflammatory alveolar space.
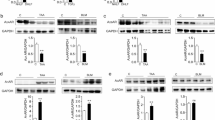
Induction of PPARγ target genes by treatment with LJ-529
LJ-529 has been shown to activate the PPARγ signaling pathway (Yu et al. 2017). Based on the role of PPARγ signaling in the regulation of inflammation (Kapadia et al. 2008; Martin 2010), we assessed whether LJ-529 increases the expression of target genes of the PPARγ signaling pathway. We found that LJ-529 upregulated the expression of PPARγ target genes, including Adrp (Fig. 4a), Cd36 (Fig. 4b), Abcg1 (Fig. 4c), and Lpl (Fig. 4d), in the lungs of mice. We next asked whether LJ-529 treatment enhances PPARγ activity. To assess PPARγ activity following LJ-529 treatment, we performed simultaneous co-transfection of PPARγ and PPRE-Luc and then conducted PPRE luciferase reporter assay. LJ-529 significantly increased PPARγ activity inhibited by LPS stimulation (Fig. 4e). These results suggest that the anti-inflammatory effect of LJ-529 (the inhibitory effect of LJ-529 on elastase-induced pulmonary emphysema) is mediated through the induction of PPARγ activity.

Activation of PPARγ by treatment with LJ-529. a–d Expression of PPARγ target genes in the lungs of mice, as determined by real-time PCR. e PPARγ activation was assessed by PPRE-Luc reporter assay in murine macrophages. The bars represent the mean ± SD; *P < 0.05 and **P < 0.01 compared with the indicated control group. PPE porcine pancreatic elastase, LJ LJ-529
Minimal toxic effects of LJ-529 in vitro and in vivo
We determined the toxicity of LJ-529 in vitro and in vivo. LJ-529 had minimal effects on the viability of three normal epithelial cell lines derived from the lung bronchus (HBE), retinal pigment epithelium (RPE), and hippocampus (HT-22). The viability of these cells was minimally affected by the drug even at a concentration of 1 μM, which induced a transcriptional increase in anti-inflammatory and M2 macrophage-associated markers in macrophage cells (Fig. 5a). Consistent with previous findings indicating no side effects of LJ-529 in in vivo experiment (Chung et al. 2006), the administration of LJ-529 caused no significant body weight changes in the mice (Fig. 5b). Moreover, histological examination also revealed that the administration of LJ-529 did not cause any changes in major organs, such as the brain, kidneys, and liver (data not shown). Therefore, although additional in vitro and in vivo investigations are needed for a precise determination of the toxicity of LJ-529, these findings indicate that LJ-529 has minimal toxicity and suggest the potential for LJ-529 as an efficacious agent for attenuating pulmonary emphysema with limited toxicity.

Minimal toxic effects of LJ-529 in vitro and in vivo. a LJ-529 had no significant cytotoxic effects in normal cell lines from various organs. HBE, RPE, and HT-22 cells were treated with vehicle (DMSO) or LJ-529 (0.1 or 1 μM) for 3 days. Cell viability was determined by the MTT assay. b Body weight changes in each treatment group during the animal experiment. PPE porcine pancreatic elastase, LJ LJ-529
Discussion
The socioeconomic burden of COPD has increased globally. However, therapeutic options for COPD are still limited, and symptomatic alleviation using inhaled bronchodilators or corticosteroids is the main therapeutic approach for COPD/emphysema in the clinic. Therefore, novel therapeutic or preventive agents for COPD/emphysema are urgently needed. In the present study, we aimed to discover new drugs with a safe toxicity profile that effectively suppress the development of emphysema. We demonstrated herein that LJ-529 is a drug that inhibits elastase-induced pulmonary emphysema in mice without overt toxicity. We further demonstrated that LJ-529 significantly suppressed the expression of anti-inflammatory cytokines through the activation of the PPARγ signaling pathway. These results suggest the potential of LJ-529 as an efficacious agent for the treatment of emphysema with minimal toxicity.
As chronic inflammation is one of the crucial pathogenic causes of the development of pulmonary emphysema, the inhibition of the inflammatory response is considered a logical approach for the treatment of COPD (Cazzola et al. 2012). Indeed, the oral phosphodiesterase 4 (PDE4) inhibitor roflumilast displays anti-inflammatory effects and has been approved for the treatment of COPD in the clinic (Wedzicha et al. 2016). However, most anti-inflammatory agents, which have various modes of action, have not reached clinical application due to a lack of effectiveness, low potency, side effects, and/or toxicity (Wedzicha et al. 2016). Therefore, it is necessary to develop potent drugs with safe toxicity profiles for the treatment of COPD/emphysema. PPARγ is considered a cellular target for the development of anti-COPD/emphysema agents due to its role in the negative regulation of inflammation. PPAR agonists exhibit excellent inhibitory effects on inflammation and COPD; however, several drawbacks, including the need for a high dose and increased cardiovascular risk in patients with diabetes (Cazzola et al. 2012), limit their use in the clinic. In this regard, our study may be of importance for the development of drugs for treating patients with emphysema.
First, our study identified a potential drug that suppresses elastase-induced pulmonary emphysema, a chronic disease that requires long-term treatment. This effect was achieved by the administration of a very low dose (50 μg/kg) of LJ-529 by oral gavage, suggesting the high potency of oral LJ-529 for suppressing emphysema. In a previous study on the antitumor effect of LJ-529 in breast cancer, the intraperitoneal administration of LJ-529 for up to 1 month did not cause toxicity in mice (Chung et al. 2006). Consistently, LJ-529 did not cause significant cytotoxicity in a panel of normal cell lines derived from various organs. More importantly, mice administered LJ-529 did not exhibit detectable changes in body weight or side effects or toxicity in various organs. Hence, our data showing the high potency and oral bioavailability of LJ-529 as well as its limited toxicity indicate its use for the treatment of emphysema.
Second, our study proposes LJ-529 as a novel PPARγ agonist with clinical applicability. A3AR agonists have been shown to suppress bleomycin-induced pulmonary inflammation and fibrosis (Morschl et al. 2008) and lung ischemia–reperfusion injury (Mulloy et al. 2013) through its anti-inflammatory activities. Consistently, the A3AR agonist IB-MECA also inhibits dextran sulfate sodium (DSS)-induced colitis in mice via modulating the NF-κB signaling pathway (Ren et al. 2015). Interestingly, the A3AR modulator has been found to act as a partial peroxisome proliferator activated receptor gamma (PPARγ) agonist and PPARδ antagonist (Yu et al. 2017). A recent study reported that an A3AR agonist, LJ529, has the capacity to act as a partial PPARγ agonist (Jeong et al. 2003; Yu et al. 2017). Previous studies have shown that PPARγ exerts anti-inflammatory effects via inhibiting the expression of inflammatory cytokines (Martin 2010) and modulating the transcriptional activity of NF-κB (Kapadia et al. 2008; Remels et al. 2009). Furthermore, previous studies have suggested that PPARγ exerts anti-inflammatory properties by inducing differentiation into M2 macrophages (Bouhlel et al. 2007) and the stimulation of A3 receptors can support survival of anti-inflammatory monocytes (Haskó and Pacher 2012). Hence, we reasoned that the inhibitory effects of LJ-529 on pulmonary emphysema and the induction of anti-inflammatory gene expression are attributed to the activation of PPARγ signaling. Indeed, in our cell line and animal models, LJ-529 upregulated the expression of several PPARγ target genes and enhanced PPARγ activity, as indicated by the augmentation of PPRE-luciferase reporter gene. These findings suggest that both PPARγ and A3AR are potential mechanisms by which LJ-529 inhibits the development of pulmonary emphysema by triggering differentiation into M2 macrophages via PPAR-γ activity and supporting survival of anti-inflammatory monocytes through the stimulation of A3 receptors.
In the present study, we used porcine pancreatic elastase (PPE)-induced emphysema mouse model. Unlike cigarette smoke-induced emphysema mouse model, the PPE-induced emphysema mouse model requires less than on month to induce disease development and has been used to screen multiple reagents within 1 month. Despite the advantage, the main weakness of the PPE-induced model is lack of all the featured clinical signs of COPD and emphysema, including changes in body weight (Wright et al. 2008; Antunes and Rocco 2011; Rodrigues et al. 2017). Therefore, further studies are warranted to validate the efficacy of LJ-529 in animal models, wherein the disease development is induced by chronic exposure to cigarette smoking.
In summary, the present study demonstrates the potential of LJ-529 as an inhibitor of elastase-induced emphysema with limited toxicity. Further investigation of the biological activities and toxicity of LJ-529 in various preclinical and clinical settings is warranted. In addition, extensive studies are required to elucidate the detailed mechanisms underlying the LJ-529-mediated blockade of pulmonary emphysema.
References
Antunes MA, Rocco PR (2011) Elastase-induced pulmonary emphysema: insights from experimental models. An Acad Bras Cienc 83(4):1385–1396. https://doi.org/10.1590/s0001-37652011005000039
Barnes PJ, Burney PG, Silverman EK, Celli BR, Vestbo J, Wedzicha JA, Wouters EF (2015) Chronic obstructive pulmonary disease. Nat Rev Dis Primers 1:15076. https://doi.org/10.1038/nrdp.2015.76
Borea PA, Varani K, Vincenzi F, Baraldi PG, Tabrizi MA, Merighi S, Gessi S (2015) The A3 adenosine receptor: history and perspectives. Pharmacol Rev 67(1):74–102. https://doi.org/10.1124/pr.113.008540
Bouhlel MA, Derudas B, Rigamonti E, Dièvart R, Brozek J, Haulon S, Zawadzki C, Jude B, Torpier G, Marx N, Staels B, Chinetti-Gbaguidi G (2007) PPARgamma activation primes human monocytes into alternative M2 macrophages with anti-inflammatory properties. Cell Metab 6(2):137–143. https://doi.org/10.1016/j.cmet.2007.06.010
Cazzola M, Page CP, Calzetta L, Matera MG (2012) Emerging anti-inflammatory strategies for COPD. Eur Repir J 40(3):724–741. https://doi.org/10.1183/09031936.00213711
Celli BR (2018) Pharmacological therapy of COPD: reasons for optimism. Chest 154(6):1404–1415. https://doi.org/10.1016/j.chest.2018.07.005
Chen ZH, Lam HC, Jin Y, Kim HP, Cao J, Lee SJ, Ifedigbo E, Parameswaran H, Ryter SW, Choi AM (2010) Autophagy protein microtubule-associated protein 1 light chain-3B (LC3B) activates extrinsic apoptosis during cigarette smoke-induced emphysema. Proc Natl Acad Sci USA 107(44):18880–18885. https://doi.org/10.1073/pnas.1005574107
Chung H, Jung JY, Cho SD, Hong KA, Kim HJ, Shin DH, Kim H, Kim HO, Shin DH, Lee HW, Jeong LS, Kong G (2006) The antitumor effect of LJ-529, a novel agonist to A3 adenosine receptor, in both estrogen receptor-positive and estrogen receptor-negative human breast cancers. Mol Cancer Ther 5(3):685–692. https://doi.org/10.1158/1535-7163.MCT-05-0245
Daynes RA, Jones DC (2002) Emerging roles of PPARs in inflammation and immunity. Nat Rev Immunol 2(10):748–759. https://doi.org/10.1038/nri912
Duc Dodon M, Vogel SN (1985) Analysis of effects of lipopolysaccharide and interferon on murine macrophages: modulation of elastase secretion in vitro. Infect Immun 49(3):709–714. https://doi.org/10.1128/IAI.49.3.709-714
Dunnill MS (1962) Quantitative methods in the study of pulmonary pathology. Thorax 17(4):320–328. https://doi.org/10.1136/thx.17.4.320
Ferry G, Lonchampt M, Pennel L, de Nanteuil G, Canet E, Tucker GC (1997) Activation of MMP-9 by neutrophil elastase in an in vivo model of acute lung injury. FEBS Lett 402(2–3):111–115. https://doi.org/10.1016/s0014-5793(96)01508-6
Gooptu B, Ekeowa UI, Lomas DA (2009) Mechanisms of emphysema in alpha1-antitrypsin deficiency: molecular and cellular insights. Eur Respir J 34(2):475–488. https://doi.org/10.1183/09031936.00096508
Guarascio AJ, Ray SM, Finch CK, Self TH (2013) The clinical and economic burden of chronic obstructive pulmonary disease in the USA. Clinicoecon Outcomes Res 5:235–245. https://doi.org/10.2147/CEOR.S34321
Haque ASMR, Moriyama M, Kubota K, Ishiguro N, Sakamoto M, Chinju A, Mochizuki K, Sakamoto T, Kaneko N, Munemura R, Maehara T, Tanaka A, Hayashida JN, Kawano S, Kiyoshima T, Nakamura S (2019) CD206+ tumor-associated macrophages promote proliferation and invasion in oral squamous cell carcinoma via EGF production. Sci Rep 9(1):14611. https://doi.org/10.1038/s41598-019-51149-1
Haskó G, Cronstein B (2013) Regulation of inflammation by adenosine. Front Immunol 4:85. https://doi.org/10.3389/fimmu.2013.00085
Haskó G, Pacher P (2012) Regulation of macrophage function by adenosine. Arterioscler Thromb Vasc Biol 32(4):865–869. https://doi.org/10.1161/ATVBAHA.111.226852
Horio Y, Ichiyasu H, Kojima K, Saita N, Migiyama IT, Fujii K, Niki T, Hirashima M, Kohrogi H (2017) Protective effect of Galectin-9 in murine model of lung emphysema: involvement of neutrophil migration and MMP-9 production. PLoS ONE 12(7):e0180742. https://doi.org/10.1371/journal.pone.0180742
Housley WJ, ÓConor CA, Nichols F, Puddington L, Lingenheld EG, Zhu L, Clark RB (2009) PPARgamma regulates retinoic acid-mediated DC induction of Tregs. J Leukoc Biol 86(2):293–301. https://doi.org/10.1189/jlb.1208733
Jeong LS, Jin DZ, Kim HO, Shin DH, Moon HR, Gunaga P, Chun MW, Kim YC, Melman N, Gao ZG, Jacobson KA (2003) N6-substituted D-4'-thioadenosine-5'-methyluronamides: potent and selective agonists at the human A3 adenosine receptor. J Med Chem 46(18):3775–3777. https://doi.org/10.1021/jm034098e
Jeong LS, Lee HW, Kim HO, Jung JY, Gao ZG, Duong HT, Rao S, Jacobson KA, Shin DH, Lee JA, Gunaga P, Lee SK, Jin DZ, Chun MW, Moon HR (2006) Design, synthesis, and biological activity of N6-substituted-4'-thioadenosines at the human A3 adenosine receptor. Bioorg Med Chem 14(14):4718–4730. https://doi.org/10.1016/j.bmc.2006.03.030
Jeong LS, Pal S, Choe SA, Choi WJ, Jacobson KA, Gao ZG, Klutz AM, Hou X, Kim HO, Lee HW, Lee SK, Tosh DK, Moon HR (2008) Structure-activity relationships of truncated D- and l-4'-thioadenosine derivatives as species-independent A3 adenosine receptor antagonists. J Med Chem 51(20):6609–6613. https://doi.org/10.1021/jm8008647
Kapadia R, Yi JH, Vemuganti R (2008) Mechanisms of anti-inflammatory and neuroprotective actions of PPAR-gamma agonists. Front Biosci 13:1813–1826. https://doi.org/10.2741/2802
Krzyszczyk P, Schloss R, Palmer A, Berthiaume F (2018) The role of macrophages in acute and chronic wound healing and interventions to promote pro-wound healing phenotypes. Front Physiol 9:419. https://doi.org/10.3389/fphys.2018.00419
Lea S, Plumb J, Metcalfe H, Spicer D, Woodman P, Fox JC, Singh D (2014) The effect of peroxisome proliferator-activated receptor-γ ligands on in vitro and in vivo models of COPD. Eur Respir J 43(2):409–420. https://doi.org/10.1183/09031936.00187812
Lee SH, Goswami S, Grudo A, Song LZ, Bandi V, Goodnight-White S, Green L, Hacken-Bitar J, Huh J, Bakaeen F, Coxson HO, Cogswell S, Storness-Bliss C, Corry DB, Kheradmand F (2007) Antielastin autoimmunity in tobacco smoking-induced emphysema. Nat Med 13(5):567–569. https://doi.org/10.1038/nm1583
Livak KJ, Schmittgen TD (2001) Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 25(4):402–408. https://doi.org/10.1006/meth.2001.1262
Lozano R, Naghavi M, Foreman K, Lim S, Shibuya K, Aboyans V, Abraham J, Adair T, Aggarwal R, Ahn SY, Alvarado M, Anderson HR, Anderson LM, Andrews KG, Atkinson C, Baddour LM, Barker-Collo S, Bartels DH, Bell ML, Benjamin EJ, Bennett D, Bhalla K, Bikbov B, Bin Abdulhak A, Birbeck G, Blyth F, Bolliger I, Boufous S, Bucello C, Burch M, Burney P, Carapetis J, Chen H, Chou D, Chugh SS, Coffeng LE, Colan SD, Colquhoun S, Colson KE, Condon J, Connor MD, Cooper LT, Corriere M, Cortinovis M, de Vaccaro KC, Couser W, Cowie BC, Criqui MH, Cross M, Dabhadkar KC, Dahodwala N, De Leo D, Degenhardt L, Delossantos A, Denenberg J, Des Jarlais DC, Dharmaratne SD, Dorsey ER, Driscoll T, Duber H, Ebel B, Erwin PJ, Espindola P, Ezzati M, Feigin V, Flaxman AD, Forouzanfar MH, Fowkes FG, Franklin R, Fransen M, Freeman MK, Gabriel SE, Gakidou E, Gaspari F, Gillum RF, Gonzalez-Medina D, Halasa YA, Haring D, Harrison JE, Havmoeller R, Hay RJ, Hoen B, Hotez PJ, Hoy D, Jacobsen KH, James SL, Jasrasaria R, Jayaraman S, Johns N, Karthikeyan G, Kassebaum N, Keren A, Khoo JP, Knowlton LM, Kobusingye O, Koranteng A, Krishnamurthi R, Lipnick M, Lipshultz SE, Ohno SL, Mabweijano J, MacIntyre MF, Mallinger L, March L, Marks GB, Marks R, Matsumori A, Matzopoulos R, Mayosi BM, McAnulty JH, McDermott MM, McGrath J, Mensah GA, Merriman TR, Michaud C, Miller M, Miller TR, Mock C, Mocumbi AO, Mokdad AA, Moran A, Mulholland K, Nair MN, Naldi L, Narayan KM, Nasseri K, Norman P, O'Donnell M, Omer SB, Ortblad K, Osborne R, Ozgediz D, Pahari B, Pandian JD, Rivero AP, Padilla RP, Perez-Ruiz F, Perico N, Phillips D, Pierce K, Pope CA 3rd, Porrini E, Pourmalek F, Raju M, Ranganathan D, Rehm JT, Rein DB, Remuzzi G, Rivara FP, Roberts T, De León FR, Rosenfeld LC, Rushton L, Sacco RL, Salomon JA, Sampson U, Sanman E, Schwebel DC, Segui-Gomez M, Shepard DS, Singh D, Singleton J, Sliwa K, Smith E, Steer A, Taylor JA, Thomas B, Tleyjeh IM, Towbin JA, Truelsen T, Undurraga EA, Venketasubramanian N, Vijayakumar L, Vos T, Wagner GR, Wang M, Wang W, Watt K, Weinstock MA, Weintraub R, Wilkinson JD, Woolf AD, Wulf S, Yeh PH, Yip P, Zabetian A, Zheng ZJ, Lopez AD, Murray CJ, AlMazroa MA, Memish ZA (2012) Global and regional mortality from 235 causes of death for 20 age groups in 1990 and 2010: a systematic analysis for the Global Bruden of Disease Study 2010. Lancet 380(9859):2095–2128. https://doi.org/10.1016/S0140-6736(12)61728-0
Mahadeva R, Shapiro SD (2002) Chronic obstructive pulmonary disease * 3: experimental animal models of pulmonary emphysema. Thorax 57(10):908–914. https://doi.org/10.1136/thorax.57.10.908
Martin H (2010) Role of PPAR-gamma in inflammation. Prospects for therapeutic intervention by food components. J Mutat Res 690(1):57–63. https://doi.org/10.1016/j.mrfmmm.2009.09.009
Morschl E, Molina JG, Volmer JB, Mohsenin A, Pero RS, Hong JS, Kheradmand F, Lee JJ, Blckburn MR (2008) A3 adenosine receptor signaling influences pulmonary inflammation and fibrosis. Am J Respir Cell Mol Biol 39(6):697–705. https://doi.org/10.1165/rcmb.2007-0419OC
Mulloy DP, Sharma AK, Fernandez LG, Zhao Y, Lau CL, Kron IL, Laubach VE (2013) Adenosine A3 receptor activation attenuates lung ischemia-reperfusion injury. Ann Thorac Surg 95(5):1762–1767. https://doi.org/10.1016/j.athoracsur.2013.01.059
Papandrinopoulou D, Tzouda V, Tsoukalas G (2012) Lung compliance and chronic obstructive pulmonary disease. Pulm Med 2012:542769. https://doi.org/10.1155/2012/542769
Raetz CR , Whitfield C (2002) Lipopolysaccharide endotoxins. Annu Rev Biochem 71:635-700. https://doi.org/10.1146/annurev.biochem.71.110601.135414
Remels AH, Langen RC, Gosker HR, Russell AP, Spaapen F, Voncken JW, Schrauwen P, Schols AM (2009) PPARgamma inhibits NF-kappaB-dependent transcriptional activation in skeletal muscle. Am J Physiol Endocrinol Metab 297(1):E174–183. https://doi.org/10.1152/ajpendo.90632.2008
Ren T, Tian T, Feng X, Ye S, Wang H, Wu W, Qiu Y, Yu C, He Y, Zeng J, Cen J, Zhou Y (2015) An adenosine A3 receptor agonist inhibits DSS-induced colitis in mice through modulation of the NF-kB signaling pathway. Sci Rep 5:9047. https://doi.org/10.1038/srep09047
Rodrigues R, Olivo CR, Lourenco JD, Riane A, Cervilha DAB, Ito JT, Martins MA, Lopes FDTQDS (2017) A murine model of elastase- and cigarette smoke-induced emphysema. J Bras Pneumol 43(2):95–100. https://doi.org/10.1590/S1806-37562016000000179
Shan M, You R, Yuan X, Frazier MV, Porter P, Seryshev A, Hong JS, Song LZ, Zhang Y, Hilsenbeck S, Whitehead L, Zarinkamar N, Perusich S, Corry DB, Kheradmand F (2014) Agonistic induction of PPARγ reverses cigarette smoke-induced emphysema. J Clin Invest 124(3):1371–1381. https://doi.org/10.1172/JCI70587
Sharafkhaneh A, Hanania NA, Kim V (2008) Pathogenesis of emphysema: from the bench to the beside. Proc Am Thorac Soc 5(4):475–477. https://doi.org/10.1513/pats.200708-126ET
Shibata S, Miyake K, Tateishi T, Yoshikawa S, Yamanishi Y, Miyazaki Y, Inase N, Karasuyama H (2018) Basophils trigger emphysema development in a murine model of COPD through IL-4-mediated generation of MMP-12-producing macrophages. Proc Natl Acad Sci USA 115(51):13057–13062. https://doi.org/10.1073/pnas.1813927115
Solleti SK, Simon DM, Srisuma S, Arikan MC, Bhattacharya S, Rangasamy T, Bijli KM, Rahman A, Crossno JT Jr, Shapiro SD, Mariani TJ (2015) Airway epithelial cell PPARγ modulates cigarette smoke-induced chemokine expression and emphysema susceptibility in mice. Am J Physiol Lung Cell Mol Physiol 309(3):L293–304. https://doi.org/10.1152/ajplung.00287.2014
Tachkov K, Kamusheva M, Pencheva V, Mitov K (2017) Evaluation of the economic and social burden of chronic obstructive pulmonary disease (COPD). Biotechnol Biotechnol Equip 31:855–861. https://doi.org/10.1080/13102818.2017.1335616
Tontonoz P, Spiegelman BM (2008) Fat and beyond: the diverse biology of PPARgamma. Annu Rev Biochem 77:289–312. https://doi.org/10.1146/annurev.biochem.77.061307.091829
Trombetta AC, Soldano S, Contini P, Tomatis V, Ruaro B, Paolino S, Brizzolara R, Montagna P, Sulli A, Pizzorni C, Smith V, Cutolo M (2018) A circulating cell population showing both M1 and M2 monocyte/macrophage surface markers characterizes systemic sclerosis patients with lung involvement. Respir Res 19(1):186. https://doi.org/10.1186/s12931-018-0891-z
Van Tho N, Ogawa E, le Trang TH, Ryujin Y, Kanda R, Nakagawa H, Goto K, Fukunaga K, Higami Y, Seto R, Wada H, Yamaguchi M, Nagao T, le Lan TT, Nakano Y (2015) A mixed phenotype of airway wall thickening and emphysema is associated with dyspnea and hospitalization for chronic obstructive pulmonary disease. Ann Am Thorac Soc 12(7):988–996. https://doi.org/10.1513/AnnalsATS.201411-501OC
Vanoirbeek JA, Rinaldi M, De Vooght V, Haenen S, Bobic S, Gayan-Ramirez G, Hoet PH, Verbeken E, Decramer M, Nemery B, Janssens W (2010) Noninvasive and invasive pulmonary function in mouse models of obstructive and restrictive respiratory diseases. Am J Respir Cell Mol Biol 42(1):96–104. https://doi.org/10.1165/rcmb.2008-0487OC
Wahl SM (2007) Transforming growth factor-beta: innately bipolar. Curr Opin Immunol 19(1):55–62. https://doi.org/10.1016/j.coi.2006.11.008
Wedzicha JA, Calverley PM, Rabe KF (2016) Roflumilast: a review of its use in the treatment of COPD. Int J Chron Obstruct Pulmon Dis 11:81–90. https://doi.org/10.2147/COPD.S89849
Wright JL, Cosio M, Churg A (2008) Animal models of chronic obstructive pulmonary disease. Am J Physiol Lung Cell Mol Physiol 295(1):L1–15. https://doi.org/10.1152/ajplung.90200.2008
Yamasaki K, Eeden SFV (2018) Lung macrophage phenotypes and functional responses: role in the pathogenesis of COPD. Int J Mol Sci. https://doi.org/10.3390/ijms19020582
Yang Z, Ming XF (2014) Functions of arginase isoforms in macrophage inflammatory responses: impact on cardiovascular diseases and metabolic disorders. Front Immunol 5:5333. https://doi.org/10.3389/fimmu.2014.00533
Yan H, Hu Y, Akk A, Ye K, Bacon J, Pham CTN (2019) Interleukin-12 and -23 blockade mitigates elastase-induced abdominal aortic aneurysm. Sci Rep 9(1):10447. https://doi.org/10.1038/s41598-019-46909-y
Yu J, Ahn S, Kim HJ, Lee M, Ahn S, Kim J, Jin SH, Lee E, Kim G, Cheong JH, Jacobson KA, Jeong LS, Noh M (2017) Polypharmacology of N6-(3-Iodobenzyl)adenosine-5'-N-methyluronamide (IB-MECA) and related A3 adenosine receptor ligands: peroxisome proliferator activated receptor (PPAR) γ partial agonist and PPARδ antagonist activity suggests their antidiabetic potential. J Med Chem 60(17):7459–7475. https://doi.org/10.1021/acs.jmedchem.7b00805
Zhou Y, Schneider DJ, Blackburn MR (2009) Adenosine signaling and the regulation of chronic lung disease. Pharmacol Ther 123(1):105–116. https://doi.org/10.1016/j.pharmthera.2009.04.003
Acknowledgements
This study was supported by a grant from the National Research Foundation of Korea (NRF), the Ministry of Science and ICT (MSIT), Republic of Korea (No. NRF-2016R1A3B1908631).
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
The authors declare that they have no conflict of interest.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
About this article

Cite this article
Boo, HJ., Park, S., Noh, M. et al. LJ-529, a partial peroxisome proliferator-activated receptor gamma (PPARγ) agonist and adenosine A3 receptor agonist, ameliorates elastase-induced pulmonary emphysema in mice. Arch. Pharm. Res. 43, 540–552 (2020). https://doi.org/10.1007/s12272-020-01235-0
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s12272-020-01235-0