
Abstract
Optogenetics, a technique that employs light for neuromodulation, has revolutionized the study of neural mechanisms and the treatment of neurological disorders due to its high spatiotemporal resolution and cell-type specificity. However, visible light, particularly blue and green light, commonly used in conventional optogenetics, has limited penetration in biological tissue. This limitation necessitates the implantation of optical fibers for light delivery, especially in deep brain regions, leading to tissue damage and experimental constraints. To overcome these challenges, the use of orange-red and infrared light with greater tissue penetration has emerged as a promising approach for tetherless optical neuromodulation. In this review, we provide an overview of the development and applications of tetherless optical neuromodulation methods with long wavelengths. We first discuss the exploration of orange-red wavelength-responsive rhodopsins and their performance in tetherless optical neuromodulation. Then, we summarize two novel tetherless neuromodulation methods using near-infrared light: upconversion nanoparticle-mediated optogenetics and photothermal neuromodulation. In addition, we discuss recent advances in mid-infrared optical neuromodulation.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Neuromodulation encompasses various stimulation modalities, e.g. electricity, chemical agents, magnetic fields, or light, to modulate neural activity in the central and peripheral nervous systems [1]. As brain mapping, neuronal disorder research, and brain simulation continue to advance, neuromodulation plays a crucial role in the field of neuroscience by uncovering underlying neural mechanisms and offering potential therapeutic strategies. Electrical stimulation techniques provide valuable insights into the behavior and cognitive functions of specific brain regions and neural circuits, but they also face two key challenges: spatially confining the electric field to a single neuron and targeting specific neuron types with distinct molecular signatures [2]. Chemogenetics, designed for neuromodulation with slow kinetics, can selectively activate ligand receptors using peripherally or orally administered chemical actuators [3, 4]. This technique is currently widely applied in the laboratory for understanding molecular interactions in the nervous system, and its translation from the lab to clinical neurosurgery is expected to yield significant potential benefits [5]. Magnetogenetics, which utilizes magnetic nanoparticle heating of thermosensitive ion channels [6,7,8] or nanoscale magnetic torque forcing of mechanosensitive ion channels [9,10,11], enables the control of neural activity with high spatial resolution and response times of seconds to sub-seconds. Nevertheless, the confinement of experimental animals within magnetic coil systems, limiting their freedom of movement and potentially affecting the study's natural conditions, is coupled with the use of strong magnetic fields for activation, which can be challenging to precisely control and may lead to unintended effects or interference with surrounding tissues.
Optogenetics, an optical technique that manipulates neural activity using visible light, has gained significant attention [12,13,14]. It combines optics and genetics, offering not only the advantage of the high spatiotemporal resolution of optics owing to the illumination with high propagation speed, which can be temporally modulated as high-frequency pulses and spatially confined to the diffraction limit but also gene targeting to confine regulation to functionally relevant neurons, achieving cell-type specificity. This powerful optogenetic technique with exceptional spatiotemporal precision and sensitivity has become an indispensable tool for thousands of neuroscience groups worldwide [15]. In recent years, the use of optogenetics has not only led to abundant discoveries on functional connectivity in neural circuits but also has provided therapeutic avenues and prosthetics for treating neuropsychiatric disorders owing to optogenetics’ capability of improving, restoring and substituting motor, sensory, and cognitive functions [16,17,18,19,20,21,22].
However, the delivery of light through brain tissue poses a significant challenge for optical neuromodulation due to the intrinsic interaction between light and biological substances [2]. Light-tissue interaction is influenced by absorption and scattering, which determine the penetration depth of light. Brain tissue, being highly scattering and absorbing, presents a poor penetration of short-wavelength visible light [23,24,25]. Consequently, conventional optogenetics heavily relies on the insertion of optical fibers, especially in deep brain regions. These tethered systems impose constraints on experimental design and interpretation, leading to tissue damage, acute inflammation, and the risk of infection [26, 27]. In addition, tethering restricts the three-dimensional movement of animals during behavioral experiments, limiting their usefulness for long-term behavior tests [28]. Furthermore, frequent movement of the fiber can result in fiber displacement, leading to potential signal instability or even interruptions [29].
To address these limitations, researchers have turned their attention to optical neuromodulation with long-wavelength light, such as orange-red and infrared light, which exhibit deeper tissue penetration and lower photo-toxicity compared to visible light (Fig. 1). Moreover, the secondary effects of light, such as photothermal [30], photoacoustic [31], and photoelectrochemical [32] effects have also been leveraged for neuromodulation, and the advantages of these second modalities have been exploited while maintaining the advantages of light stimulation. With broad multidisciplinary integration, neuromodulation by the secondary effects of light is making great strides toward achieving less invasive, more efficient, and safer approaches. Nanomaterials have also been incorporated as transducers in neuromodulation, offering higher spatiotemporal resolution and minimizing tissue damage [1].

Optical neuromodulation: Ion channels are opened to regulate neural activity by various stimulation modalities including light and the secondary effects of light.
In this paper, we provide an overview of tetherless optical neuromodulation methods, focusing on the use of long-wavelength light. These methods can be broadly categorized into two groups based on their operation mechanisms: (1) the extension of conventional optogenetics using red-shifted photosensitive proteins responsive to orange-red light and (2) alternative methods utilizing near-infrared to mid-infrared light stimulation. We discuss the discovery and recent advances of these representative tetherless optical neuromodulation methods, starting with orange-red to near-infrared wavelengths, followed by mid-infrared. Finally, we present a summary and discussion of the progress in the field.
Orange-red Light Optogenetics
In 2005, a groundbreaking study by Karl Deisseroth and his colleagues marked a pivotal moment in the field of optogenetics. They recorded action potentials in cultured hippocampal neurons upon blue light illumination, achieved through the expression of channelrhodopsin2 (ChR2) [12]. Fig. 2A illustrates the key components of in vivo optogenetics implementation, which involve carefully selecting photosensitive proteins capable of modulating specific cellular functions, delivering these proteins to targeted cellular populations using virus vector transfection, and precisely delivering light with tailored modulation patterns [22].

Copyright 2022 Elsevier.
Optogenetics and opsins. A The schedule of optogenetic implementation in living animals. B A summary of photosensitive proteins. Opsins used in conventional optogenetics are typically most sensitive in the blue-to-yellow spectral range. Adapted with permission from [33].
Microbial rhodopsins, known for their structural simplicity and rapid kinetics, are the most commonly used photosensitive proteins in optogenetics. However, opsins used in conventional optogenetics are typically most sensitive in the blue-to-yellow spectral range (Fig. 2B) [33], while these wavelengths show poor penetration in biological tissue. In the pursuit of enhancing optogenetics performance, researchers have dedicated significant efforts to the development of new rhodopsins with long-wavelength responsive properties. These innovations hold the promise of enabling tetherless optogenetics, representing a significant step forward in the field. Achieving this goal involves identifying previously unknown microbial channelrhodopsins (ChRs) through genome mining and applying molecular engineering techniques to generate variants that are sensitive to orange-red light. In this section, we delve into the development and application of long-wavelength-responsive microbial rhodopsins, highlighting two distinct types that have shown great promise.
Exploring Orange-red Wavelength Excitatory Rhodopsins
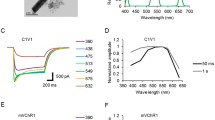
In 2008, Zhang et al. [34] embarked on a quest to explore new opsins. They screened U.S. Joint Genome Institute databases and discovered a cation-conducting channelrhodopsin (VChR1) from Volvox carteri that can drive spiking at 589 nm. However, the expression level and photocurrents of VChR1 were found to be significantly lower than ChR2 in most host cells [34]. A chimeric opsin variant, C1V1, composed of sequences from both ChR1 and VChR1, retained the red-shifted spectrum but also exhibited improved membrane trafficking and enhanced photocurrents [32]. In particular, the E122T mutation (C1V1E122T) showed reliable spiking with 630 nm light [35]. In the same year, Govorunova et al. [36] identified one new channelrhodopsin from Mesostigma viride (MChR1) with the same red-shifted spectrum as VChR1.
With comprehensive consideration of the properties of different ChR variants, a new variant was engineered by Lin et al. [37]. The new red-activatable ReaChR is optimally excited with orange to red light (590–630 nm) and offers improved membrane trafficking, higher photocurrents, and faster kinetics compared to VChR1 and C1V1 [37]. ReaChR expressed in the vibrissa motor cortex can drive spiking and vibrissa motion in awake mice when excited with red light through the intact skull [37]. Klapoetke et al. [38] de novo sequenced 127 species of algae and they found that Chrimson's response spectrum is red-shifted by 45 nm compared to ReaChR. Further, they engineered Chrimson with the K176R mutant, denoted ChrimsonR, which sped up the off-kinetics without altering the red-shifted action spectrum [38]. Reliable spiking in cortical slices with ChrimsonR expression can be elicited even under 735 nm illumination [38].
Continuing the quest for novel opsins, Deisseroth's group turned their attention to marine microbial organisms. Their efforts led to the identification of a promising new marine opsin named ChRmine by screening the genomes of over 600 marine microbes [39]. Reliable photostimulation (spike probability >0.8) in ChRmine-expressing neurons at a depth of up to 7 mm was recorded [40]. Mouse behavior in place preference was modulated with transcranial light delivery to activate ChRmine-expressing neurons in the ventral tegmental area [40]. By leveraging the structural information of ChRmine, they designed the variant rsChRmine with red-shifted properties [41]. rsChRmine-expressing neurons in freely moving mice showed a response to 720 and 750 nm stimulation [41]. This may be the first rhodopsin so far to be directly activated even under near-infrared irradiation (>740 nm) [41].
Exploring Orange-red Wavelength Inhibitory Rhodopsins
Inhibiting cellular electrical activity requires the expression of inhibitory rhodopsins. In the early stages, Natronomonas halorhodopsin (NpHR), an ion pump rhodopsin, was chosen as an optogenetic tool for silencing neuronal activity due to its stable photocurrents with step-like characteristics [42]. Subsequently, the developed eNpHR3.0 exhibited photocurrents that were 20 times stronger than those reported for NpHR in response to 589 nm light [43].
In parallel with the exploration of red-shifted excitatory rhodopsins, researchers also worked on the development of red-shifted inhibitory rhodopsins. Chuong et al. [44] identified Jaws from H. salinarum (strain Shark) following some engineering processes. Photocurrents on Jaws induced by 632 nm red light were significantly higher than eNpHR3.0 or ArchT [44]. The inhibition of in vivo neural activity by Jaws was also much higher than eNpHR3.0 when illuminated with 637 nm red light [44]. Moreover, researchers successfully inhibited neurons in the awake mouse cortex by delivering 635 nm red light through the intact skull [44].
In a groundbreaking study, Govorunova et al. [45] reported that four ACRs derived from the Labyrinthulea exhibited red-shifted properties with maximum action spectra at 590–600 nm. Furthermore, they stated that RubyACRs can generate larger photocurrents and offer more efficient optogenetic inhibition for the long-wavelength spectral range compared to Jaws, as Jaws’ efficiency is limited by the translocation of only one ion per captured photon across the membrane [45].
Section Discussion
Spectral tuning of rhodopsins into the deep-red and near-infrared (NIR) spectral regions is of great demand in optogenetics because more bathochromic light into the NIR range penetrates deeper into living tissue. It has been extraordinarily difficult to achieve action spectrum peak shifts of more than 10–20 nm (while maintaining high functionality in ChRs) by molecular engineering alone [46]. In recent years, machine learning (ML) has shown unique advantages in studying rhodopsin structure, predicting function, and analyzing properties [47, 48]. Arnold and colleagues designed ChRger2, which was demonstrated to excite neuronal activity and control locomotion directionally in freely moving mice without fiber-optic implantation [49]. Thus, integrating ML-based approaches with genome mining and molecular engineering holds promise for enhancing the efficiency of discovering novel photosensitive proteins.
In addition to spectral properties, other properties of photosensitive proteins are crucial for their application in tetherless optogenetics, such as light sensitivity and expression level [50]. Enhancing photosensitivity can somewhat alleviate the requirement for light penetration, enabling minimally invasive or noninvasive neuromodulation. Feng’s group engineered a novel photosensitive protein with ultrahigh light sensitivity (SOUL) [51]. They demonstrated that SOUL can be used to modulate neural activity in deep brain regions and change behavioral performance via transcranial illumination with orange light (589 nm) [51]. Therefore, future advances could integrate improvements in wavelength with other properties to achieve more effective noninvasive neuromodulation.
Near-infrared Light Upconversion-mediated Optogenetics
Due to the limited absorption of NIR light from 700 nm to 1500 nm by biological tissues, NIR light possesses the unique characteristic of deep tissue penetration and is often referred to as the "optical window of biological tissues" [52]. In addition, NIR light propagates through air with minimal loss, allowing researchers to position the light source at varying distances above the experimental subject’s head. Unfortunately, infrared light-responsive opsins may not exist in nature. This is because the energy barriers of the opsins must be very low to absorb and detect low-energy infrared photons. Too low energy barriers will make thermal energy more likely to spontaneously excite the opsins, resulting in unendurable high thermal noise [53,54,55]. Moreover, we can confirm that it will be a high cost in both time and money to obtain an ideal opsin responsive to NIR wavelengths, based on the experience of exploration of long-wavelength opsins as discussed in the previous section. Hence, it is difficult to replace visible light with NIR light directly in conventional optogenetics. In recent years, some alternative neuromodulation methods based on NIR light have been developed and shown great promise. Among various neuromodulation schemes based on NIR light, upconversion-mediated optogenetics and photothermal neuromodulation are the two most frequently applied novel strategies, as shown in Fig. 3A and B, respectively. Hence, we present an overview of these representative strategies in the following two sections.

A The mechanism of NIR UCNP-mediated optogenetics. B The mechanism of NIR photothermal neuromodulation targeting TRPV1.
The principle of upconversion is based on anti-Stokes’s law. In brief, the upconversion material is excited by low-energy light and emits high-energy light, i.e., the material emits light of short wavelength and high frequency after excitation by light of long wavelength and low frequency [56]. Unlike the conventional luminescence process (involving only one ground state and one excited state), the upconversion process requires many intermediate states to accumulate the energy of excited photons at low frequencies. There are three main luminescence mechanisms: excited state absorption, energy conversion processes, and photon avalanches [56]. The highly efficient upconversion process can be achieved by doping with trivalent rare-earth ions, due to their long sub-stable energy lifetimes [57]. Taking energy transfer upconversion as an example, it requires a lanthanide pair, such as ytterbium/erbium (Yb3+/Er3+), ytterbium/thulium (Yb3+/Tm3+), and ytterbium/holmium (Yb3+/Ho3+), to serve as a sensitizer for photon absorption and an activator for photon emission, respectively [58]. Some innovative technologies such as super-resolution microscopy, optogenetics, nanolasing, and optical anticounterfeiting have been developed based on upconversion nanoparticles (UCNPs) in recent decades [59,60,61,62,63,64,65,66,67].
UCNP-mediated optogenetics was first proposed in 2011 by Karl Deisseroth and Polina Anikeeva [68]. About four years thereafter, Shah et al. [69] took upconversion nanomaterial as relay media to transfer infrared light (980 nm) to blue luminescence (475 nm) and they further activated neural cells in vitro based on UCNP-mediated optogenetics, with efficiency comparable to that of 470 nm blue light. Since then, this technology has made breakthroughs every year (Fig. 4). In 2016, Bansal et al. [70] developed a novel quasi-continuous wave excitation method to enhance the emission intensity of UCNPs. By using this excitation regime, in-vivo C. elegans models expressing ChR2 in their mechanosensory neurons were activated with low-power NIR illumination, showing a reversal response akin to being touched [70]. Immediately afterward, Ai et al. [71] demonstrated that UCNP-mediated optogenetics can noninvasively activate ChR2 under 808 nm illumination and effectively manipulate cation influx. Moreover, the relay media used in their experiments, lanthanide-doped upconversion nanocrystals, could be site-specifically conjugated onto the cell surface based on native metabolic glycan biosynthesis [71]. Although C. elegans and zebrafish are small and optically transparent with simple nervous systems, the successful implementation of upconversion-mediated optogenetics on them paved the way for validation of the noninvasive application of NIR light-enabled optogenetics in mammals.

The development of UCNP-mediated optogenetics: Proposed in 2011; Application in cultured cells, C. elegans, and zebrafish; Application in mouse with an implantable microdevice; Deep brain neuronal stimulation in rodents without implants; Establishment of near-infrared image vision in mammals; Bidirectional photoactivation with switchable upconversion emissions; Near-infrared manipulation of multiple neuronal populations via trichromatic upconversion. Adapted with permission from: ref [69], copyright 2015 The Royal Society of Chemistry; ref [70], copyright 2016 Wiley-VCH; ref [71], copyright 2017 Wiley-VCH; ref [72], copyright 2017 Elsevier; ref [74], copyright 2018 The American Association for the Advancement of Science; ref [53], copyright 2019 Elsevier; ref [76], copyright 2019 Springer Nature; ref [77], copyright 2021 Springer Nature.
In 2017, Shi and colleagues successfully applied UCNP-mediated optogenetics to wireless regulation of the central nervous system in rodents [72]. They designed an implantable microdevice comprising UCNPs [72]. In combination with a robotic tetherless NIR projection system, the brain activities of the mouse in various regions at different depths were modulated, including the cortical striatum (3 mm), visual cortex (1 mm), and ventral tegmental area (4.5 mm) [72]. About three years later, they reported another flexible and fully implantable upconversion device using thermoplastic polypropylene as the backbone, with long-term biocompatibility in live animals for more than four months [73]. The neural activities and behaviors in vivo of mice with the UCNP device implanted in the spinal cord could be modulated under remote NIR irradiation [73]. However, using implantable microdevices not only complicates the procedure and poses the risk of infection but also requires a second surgery to remove the microdevices after neuromodulation is completed.
In an attempt to achieve better biocompatibility and deeper penetration to avoid the use of an implanted device, Chen et al. [74] synthesized core-shell UCNPs (NaYF4:Yb/Tm@SiO2) in 2018. Good performance of this kind of material was shown based on their measurements and calculations, including ~2.5% conversion yield of NIR to blue light, good biocompatibility, low cytotoxicity, long-term stability, and low dispersion in tissue [74]. In line with this, they achieved the first truly UCNP-mediated optogenetics without implants to stimulate deep brain neurons in rodents [74]. For example, they delivered ChR2 to ventral tegmental area dopamine neurons of TH-Cre mice via adeno-associated virus followed by UCNP injection [74]. Then the release of dopamine could be evoked by transcranial NIR irradiation [74]. Moreover, the results of in-vivo experiments were consistent with their findings on the performance of their UCNPs [74]. Subsequently, Yamanaka’s group from Japan also demonstrated that UCNP-mediated optogenetics without fiber could manipulate the locomotor behavior of mice by modulation of neurons in the dorsal striatum (~ 2 mm in depth) [75].
Notably, UCNP-mediated optogenetics not only addresses the issue of deep tissue penetration but also offers another noteworthy advantage. Hybrid UCNPs with different excitation and emission wavelengths can enable simultaneous neuronal excitation and inhibition within a single region or across multiple deep brain regions. This capability allows for the manipulation of multiple neuronal populations using a single set of particles. Regarding the spectral tunability of UCNPs, Mei et al. [76] reported cross-relaxation-mediated switchable UCNPs which could emit green and red light, simply by adjusting the excitation wavelength from 980 nm to 808 nm. These UCNPs could activate VChR1 and Jaws in the same cell programmable to manipulate membrane polarization [76]. Liu et al. [77] developed an excitable trichromatic UCNP to emit blue, green, and red emissions, by adjusting the excitation wavelength to 980 nm, 808 nm, and 1532 nm, respectively. Correspondingly, ChR2, C1V1, and ChrimsonR could be activated to selectively modulate three distinct neuronal populations, offering the ability for sophisticated brain functions and behavior dissection [77]. Recently, Shi’s group [78] reported a hierarchical device with a multi-arrayed optrode format containing engineered UCNPs with up to three modulation wavelengths matched with different channelrhodopsins. Spatially selective and combinatory optogenetics in behaving mice was achieved with this device [78].
Several noteworthy applications of UCNP-mediated optogenetics deserve mention due to their significant impact. One particularly promising area is visual restoration, leveraging the eye's natural ability for efficient light transmission. An intriguing development by Ma et al. [53] involved the creation of ocular injectable photoreceptor-binding UCNPs. They demonstrated that mice with these nanoantennae could perceive NIR light as well as see NIR light patterns [53]. Moreover, the NIR pattern vision is compatible with native vision, offering options for mammalian vision repair and enhancement [53]. Regenerating peripheral nerves after severe injury remains a formidable challenge. In this context, Yan et al. [79] devised an innovative approach involving the development of acetylcholine-modified UCNPs. These nanoparticles possess the unique ability to anchor themselves to acetylcholine receptors on the postsynaptic membrane, facilitating membrane depolarization of cholinergic motor neurons through UCNP-mediated optogenetics [79]. This, in turn, triggers a cascade of events that play a crucial role in guiding the reattachment of completely transected tibial nerves [79]. The results of electrophysiological and behavioral experiments further verified the functional recovery of transected peripheral nerves by this approach [79].
Indeed, the superiority of UCNP-mediated optogenetics, including advantages such as excellent penetration depth without the requirement for optical fiber implantation and spectral tunability under various excitation conditions, holds promise for the emergence of even more intriguing applications. However, there are still some limitations in UCNP-mediated optogenetics, and the corresponding improvements are underway and will continue. For example, improved biocompatibility of UCNPs via novel methods for synthesis and surface engineering will greatly raise the safety of neuromodulation. The low conversion efficiencies inevitably require high power irradiation or a high concentration of UCNPs to produce an effective stimulation strength. This approach may lead to concerns such as tissue overheating, photodamage, or cytotoxicity. The enhanced efficiency of triplet-triplet annihilation upconversion may provide a potential solution to the challenges associated with high-power illumination or the need for a high concentration of UCNPs [62]. Furthermore, the combination of highly photosensitive opsins, such as ChRmine, with opsin-bound UCNPs may enable the manipulation of neuronal activity with low-power NIR irradiation [80].
Photothermal Neuromodulation by Near-infrared Light
Biological tissues rich in water absorb NIR light and convert it into heat. Furthermore, the conversion efficiency and focus of this photothermal effect can be significantly enhanced by various photothermal transducers, including gold nanomaterials [81], carbon nanomaterials [82], and semiconducting polymer nanobioconjugates [83]. This highly efficient and spatially precise photothermal effect in biological tissues is extensively used in current biomedical applications, such as tumor ablation, biosensing, drug-targeted delivery, and bioimaging [84,85,86,87,88,89,90]. Specifically, two photothermal-based mechanisms are applied for neuromodulation: the optocapacitive effect and photothermal activation of transient receptor potential (TRP) cation channels.
The optocapacitive effect operates by inducing a transient temperature increase, on the order of kilokelvins per second, upon light irradiation. This temperature increase alters the capacitance of the cell membrane, subsequently generating a capacitive current across the membrane. As a result, electrogenic cells, like neurons, are depolarized [91]. In 2012, Shapiro et al. [92] proposed this mechanism and conducted voltage and current clamp measurements in three models (oocytes, cultured human embryonic kidney cells, and an artificial bilayer) under NIR illumination to verify it. Their results demonstrated that the change in membrane capacitance of cells was induced by the transient local temperature increase. Moreover, the change in membrane capacitance depends on the temporal gradient of heating and not on the absolute temperature [92]. It is challenging to achieve a steep slope of temperature increase in brain tissue with high scattering properties using the NIR laser alone. Hence, Carvalho-de-Souza’s group [93] and Tian’s group [94] developed gold and silicon nanoparticles, respectively, as efficient photothermal transducers to achieve rapid localized brain tissue warming. Then efficient and highly localized optocapacitive modulation of neurons was achieved. However, even though the thermal effect of NIR light can be focused on a specific brain region to achieve spatial specificity with the assistance of photothermal transducers, differentiating the effect on various types of neurons within the target brain region remains a challenge.
As an alternative to the non-specific optocapacitive mechanism, photothermal neuromodulation targeting thermosensitive ion channels has been extensively studied. These thermosensitive transmembrane proteins, similar to photosensitive transmembrane proteins, can act as switches, forming the second type of photothermal-based neuromodulation method: photothermal activation of TRP cation channels. Currently, a total of 11 thermoTRPs, including thermoreceptors TRPV1-TRPV4 and TRPM2-TRPM5, as well as cold receptors TRPM8, TRPC5, and TRPA1, have been confirmed to exhibit temperature-dependent behavior, ranging from responding to painful heat to comforting warmth, coolness, and bone-chilling cold [95]. High-resolution structures of almost all TRP channels have been revealed in recent years. They share the structural features of four subunits, each of which contains six transmembrane segments (TM1−TM6), with TM1−TM4 and TM5-TM6 serving as the voltage-sensitive and pore domains, respectively [63, 96].
One well-characterized member of the TRP family is the TRPV1 receptor, which plays crucial roles in thermoregulation, pain perception, maintenance of bone homeostasis, and gastrointestinal function [97]. TRPV1 was first successfully cloned from rat dorsal root in 1997 [98], and the structures of the full-length TRPV1 closed and open states at 3 Å resolution were revealed in 2013 [99, 100]. In recent years, the mechanisms of how TRPV1 senses temperature and nociception, and thus adapts to its surroundings, have also been extensively revealed [101,102,103]. In mammals, TRPV1 is widely distributed in the peripheral and central nervous systems. TRPV1 is a non-selective cation channel but differs in its permeability to divalent cations, Ca2+ being the most permeable [104]. Activation of the ion channel can be achieved by binding to different sites on the transmembrane protein TRPV1 by various noxious stimuli, such as capsaicin, noxious heat, and acidic pH values. Notably, in neural cells expressing TRPV1, when the temperature rises to 43℃, the temperature-gated TRPV1 located on the plasma membrane opens, allowing extracellular Ca2+ to flow into cells and eliciting action potentials. Fig. 3B illustrates the principle of NIR photothermal neuromodulation targeting TRPV1.
Proof-of-concept demonstrations of photothermal neuromodulation targeting TRPV1 were initially performed on cells in vitro. In 2015, Nakatsuji et al. [105] demonstrated that gold nanorods (GNRs) coated with cationized high-density lipoproteins target cell membranes. Through the conversion of 780 nm NIR illumination into heat using GNRs as photothermal transducers, TRPV1 ion channels are subsequently activated, resulting in intracellular Ca2+ influx in cocultured dorsal root ganglion neurons [105]. In 2016, Pu’s group [106] developed organic semiconducting polymer nanoparticles (SPNs) with higher photothermal conversion efficiency, faster heating capability, and better photothermal stability at 808 nm illumination as compared with GNRs. For the specific activation of thermosensitive ion channels on neurons, SPNs were conjugated with an anti-TRPV1 antibody, referred to as SPNsbc [106]. Upon remote 808 nm NIR light illumination, SPNsbc acted as photothermal nanotransducers to activate TRPV1 rapidly and specifically and elicit Ca2+ influx of cultured neuron hybrid ND7/23 cells that intrinsically express TRPV1 on their plasma membrane (Fig. 5A) [106]. One interesting application of NIR photothermal neuromodulation targeting TRP channels in vivo was reported by Nelidova et al. [107] in an article in Science. In their study, thermoTRPs (TRPV1 or TRPA1) were expressed in light-insensitive retinal cones in a mouse model with regional retinal degeneration [107]. Gold nanorods conjugated with thermoTRP antibodies were delivered to the extracellular domains of retinal cones as well [107]. Upon NIR stimulation, activity was increased in retinal cones, as well as downstream neurons in the ganglion cell layer and cortical area [107]. Moreover, the mouse was able to perform a learned light-driven behavior (Fig. 5B) [107]. However, further investigation is needed to explore the full potential and effectiveness of the techniques mentioned in these papers when applied to relatively deep neuro tissue areas of living animals.

Photothermal Neuromodulation with two NIR windows. NIR-I (750-1000 nm): A Photothermal activation of the TRPV1 ion channels in cells in vitro. B Restoring light sensitivity using tunable near-infrared sensors. NIR-II (1000–1350 nm): C Antibody-conjugated gold nanoparticles as nanotransducers for photothermal stimulation of neurons in rats. D Tetherless photothermal deep-brain stimulation with MINDs in freely behaving mice. Adapted with permission from: ref [106], copyright 2016 American Chemical Society; ref [107], copyright 2020 The American Association for the Advancement of Science; ref [111], copyright 2022 Springer Nature; ref [30], copyright 2022 Springer Nature.
In recent years, it has been well demonstrated that the second NIR window (NIR-II, 1000–1350 nm) has a stronger penetrating capability and higher power safety limit compared to the first NIR window (NIR-I, 750-1000 nm), making it more suitable for biological applications [52, 108,109,110]. In 2022, Liu et al. [111] reported one novel NIR-II light-mediated neuromodulation strategy. In their study, PEGylated polydopamine (PDA)-coated Au nanoparticles conjugated with anti-TRPV1 antibody (noted as Au@PDA-PEG-Ab) were constructed. The encapsulated dopamine improved the surface chemical tunability, thermal stability, and biocompatibility as well as biodegradability of the nano transducers, while the attached antibodies improved their targetability to the target neurons [111]. In vivo, mouse experiments demonstrated the ability of this strategy to remotely modulate the activity of hippocampus neurons located 5 mm beneath the cortex, where endogenous TRPV1 is abundantly expressed (Fig. 5C) [111]. Another recent study by Hong’s group [30] reported photothermal neuromodulation with wide-field NIR-II illumination through the scalp and skull of freely behaving mice without any fiber implantations or head tethering. They used a semiconducting polymer (pBBTV) as the core and an amphiphilic polymer (PLGA-PEG) as the shell to synthesize the photothermal nano transducer, referred to as MINDS [30]. MINDS have an average diameter of ~40 nm and a photothermal conversion efficiency of up to 71% at 1064 nm [30]. The conditioned place preference results, as well as the electrophysiological and tissue sectioning results presented in their article, demonstrate that this novel strategy can successfully use NIR-II light to excite midbrain ventral tegmental area (VTA) neurons through the intact scalp and skull (Fig. 5D) [30]. This strategy, with the same but extended steps of exogenous gene transfection and light illumination as traditional optogenetics, offers the possibility of neuromodulation in behavioral experiments of multiple-subject social interactions.
NIR photothermal neuromodulation, which converts NIR light into thermal energy through photothermal nanotransducers, can either alter the electrical capacitance or activate thermosensitive TRP cation channels, resulting in reversible modulation of nerve cell excitability, as demonstrated earlier. However, some contradictory results regarding NIR photothermal neuromodulation have been reported. For instance, Nam’s group [112] reported that the spike rates of cultured hippocampal neurons with GNRs attached to cellular membranes could be inhibited through membrane-localized photothermal effects upon 785 nm NIR illumination. The different results of GNR-mediated photothermal stimulation with and without the TREK-1 channel blocker fluoxetine suggested that the TREK-1 channel, one of the thermosensitive K+ channels expressed intrinsically throughout the whole brain area, is involved in the inhibition of action potentials [112]. Moreover, they developed a closed-loop system by combining a thermoplastic simulation platform based on GNRs and 808 nm NIR irradiation with a proportional-integral controller, which can precisely modulate the spike rates of cultured neurons to remain constant or change in a sinusoidal form [113]. Recently, a collaborative study conducted by Singamaneni’s group and Raman’s group [114] has further shown that combining PDA nanoparticles with NIR light can inhibit the activity of neurons in the mouse brain using the photothermal effect, and can inhibit their firing by controlling the laser intensity in a graded manner. Once the light stimulation is stopped, the neurons return to full normalcy without cell damage.
NIR photothermal neuromodulation, characterized by tetherless properties, can complement conventional optogenetics as a modulatory approach. Compared to conventional optogenetics, which requires exogenous gene transfection into target neurons, photothermal neuromodulation does not require prior transgenesis in most cases. Even in a photothermal neuromodulation approach targeting TRP channels, where TRP gene transfection is performed in order to allow more uniform expression of TRP on neurons or where one wants to target a specific type of neuron for modulation because TRP channels are endogenous proteins widely expressed in mammals, their low immune rejection and high safety profile will be far superior to that of exogenous photosensitive proteins that must be transfected into experimental animals in conventional optogenetics. This feature will facilitate the development of photothermal approaches to modulate neurons in large mammals and even clinically. Although photothermal neuromodulation operates by multiple mechanisms and is able to achieve excitatory or inhibitory modulation of neurons, challenges also exist to ensure that set-ups are precise across experiments to generate consistent and reproducible results.
Mid-infrared Optical Neuromodulation
To avoid the detrimental effects of excessive photothermal reactions, precise parameter settings are crucial in both UCNP-mediated and photothermal-based neuromodulation techniques. Seeking to bypass the thermal effects associated with NIR approaches, researchers have recently turned their attention to the mid-infrared (MIR) range, specifically between 3.5 μm and 5.7 μm, which exhibits minimal water absorption. In a recent collaboration [115] involving Shu’s group, Song’s group, Wen’s group, and Chang’s group, they discovered that the carbonyl group (–C=O) enriched in the functional domain responsible for K+ selectivity in the K+ channel resonantly absorbed the photon energy of MIR stimulation. This absorption led to increased K+ permeability and a stronger K+ current. However, all the resonance peaks of the –COO group, responsible for screening Na+ ions in the Na+ channel, did not align with MIR stimulation, missing the chance to accumulate energy. At the cellular level, the augmented K+ current influences neuronal output by narrowing signal-action potential waveforms and introducing a shunting effect. This delicate interplay results in weakened responses to subtle stimuli and heightened responses to stronger stimuli, demonstrating gain modulation [115]. In animal studies, zebrafish subjected to various intensities of MIR stimulation displayed similar gain modulation effects [115].
Building on these remarkable findings, Chang and colleagues [116] further demonstrated the potential of mid-infrared modulation (MIM) techniques. They delivered mid-infrared light energy to brain neurons through open or thinned intact skulls, resulting in a significant increase in neuronal firing rates and the activation of neurons in targeted brain regions of rodents. They unveiled the capability of MIM to penetrate up to 0.3 mm into brain tissue, activating over 10% of the cortical neuronal population in vivo [116]. Furthermore, the number of activated neurons correlated with the duration of irradiation [116]. To explore the practical applications of MIM, the researchers devised a sound-licking associative learning task. During training, MIM stimulation did not expedite the shortening of water-licking reaction time or improve the success rate. However, it significantly enhanced the learning speed, enabling them to outperform control mice by ~50% and achieve high success rates within a remarkably short period [116].
While these findings highlight the potential of MIR as a nonthermal physical neuromodulation method, it is important to acknowledge its limitations. With or without skull removal, its penetration depth into brain tissue is limited to ~0.3 mm, confining its effectiveness on cortical areas. Furthermore, the absence of exogenous gene transfection limits the precision of neuromodulation in targeting specific cell types. In addition, MIR neuromodulation relies solely on K+ resonance for its functionality, necessitating the presence of K+ channels in target neurons and restricting its application to neuronal activation.
Discussion and Summary
In this paper, various strategies for tetherless neuromodulation using light ranging from orange-red to mid-infrared have been discussed, alongside their effectiveness and noninvasive performance in neuroscience experiments on living animals. These techniques hold great promise for studying intact brain circuits and treating neurological disorders through artificial neuromodulation without the need for optical implantation. Each method has its advantages and disadvantages, which can be examined through four factors: protein expression, light penetration depth, photothermal effect, and mediating nanomaterials.
In mammalian systems, viral delivery is commonly used to achieve specificity in gene expression within targeted cells. However, delivering exogenous photosensitive proteins to neural cells in large mammals, especially primates presents challenges. While studies have shown that local injection of adeno-associated virus expressing ChR2 in retinal neurons of rats does not evoke immunogenic responses [117, 118], there are concerns about using proteins from other species, such as bacteria and algae, which may be perceived as foreign by the human immune system or be toxic to human cells [119, 120]. Alternatively, temperature-sensitive proteins like TRP cation channels have garnered attention as targets for remote activation due to their low or no immune response. TRP ion channels, such as TRPV1, naturally exist in the mammalian nervous system, and achieving sustained and uniform levels of TRPV1 expression through transgene delivery can enhance the efficiency of photothermal neuromodulation. Some strategies for near-infrared and mid-infrared neuromodulation do not require protein transfection, thereby improving biosafety. However, achieving cellular modulation specificity without transfection of target neurons is challenging. A promising direction for protein expression in targeted neurons is systemic viral delivery via intravenous injection, which reduces invasiveness compared to intracranial injection [40, 49, 121]. High light-sensitive and high photocurrent opsins are needed for systemic viral delivery due to the low multiplicity of infection [40, 49].
The brain, characterized by high scattering and absorbing properties, hinders the penetration of short-wavelength light [24, 25]. In the wavelength range from orange-red to mid-infrared, orange-red light has the smallest penetration depth in brain tissue, followed by mid-infrared and then near-infrared. Consequently, the reviewed strategies can be ranked in terms of penetration depth as follows: upconversion-mediated optogenetics and photothermal neuromodulation > mid-infrared light neuromodulation > orange-red light neuromodulation. Future development of photosensitive proteins should aim to red-shift their action spectra to longer wavelengths. Calculations suggest that the maximum penetration depth in the brain is ~1070 nm [2, 30]. Photothermal neuromodulation using a 1064 nm neodymium-doped yttrium aluminum garnet laser can reach the deepest brain tissue without the need for optical implantation. However, the use of upconversion nanoparticles (UCNPs) for neuromodulation with an excitation wavelength of 1064 nm has not been reported.
When light is absorbed by biological tissue, a significant portion is converted to heat, leading to an increase in local brain temperature. Studies have reported temperature elevations under different wavelengths of light, ranging from 0.3–5.4 °C for UCNP-mediated optogenetics [74], 2℃ for NIR photothermal neuromodulation [30], and <0.69 °C for mid-infrared [116]. The average brain temperature in healthy humans is ~38.5 °C C, with deeper brain regions often exceeding 40 °C C [122]. Elevated temperature not only raises concerns about thermal damage to brain tissue and function but also introduces potential inconsistencies in neuromodulation outcomes. Studies of NIR photothermal neuromodulation have shown that the photothermal effect causes neuronal inhibition and that a possible mechanism is related to the opening of K+ channels resulting in K+ efflux [112,113,114]. Another research paper pointed out that the intensity of light inherited from conventional optogenetics used light also inevitably heats brain tissue, causing a localized brain tissue warming of 0.2–2 °C [123]. This photothermal effect may inhibit neuronal firing, even if no photosensitive proteins are expressed in neurons [123], where the photothermal effect causes activation of neuronal K+ channels, which in turn causes outflow of membrane currents and eventually inhibits neuronal firing [123]. Careful selection of light stimulation parameters, such as power intensity and pulse frequency, is crucial, considering the potential inhibitory effects of the photothermal response when interpreting experimental results.
The incorporation of nanomaterials with light offers several advantages for optical neuromodulation, including higher spatiotemporal resolution and the ability to use lower stimulus intensities. The conversion efficiency of nanomaterials has been a focal point for improvement, with photothermal conversion typically being more efficient than photon upconversion, which only has a 1%–10% conversion yield of NIR to visible light [124, 125]. The ability to optically evoke spikes necessitates that opsins possess not only photocurrents sufficient to depolarize neurons above the spike threshold but also on-, off- and recovery kinetics fast enough to precisely control spike timing and fidelity [50, 126]. Conventional optogenetics with direct activation of photosensitive proteins by light has a kinetic time of milliseconds or even sub-milliseconds. When nano-mediated materials are introduced, the kinetic process is extended. Electrophysiological measurements revealed a latency of 2.9 s and an offset time of 8.6 s, and a similar latency time of 5.0 s and offset time of 10.7 s was reported from behavioral experiments when NIR photothermal neuromodulation targeting TRPV1 was used [30]. This second-level response delay hampers the precise tracking of the true relationship between neuronal activity and animal behavior. Moreover, some functional modifications of nanomaterials have to be considered carefully, such as biocompatibility and targeting. The nano transducers labeled with an anti-TRPV1 antibody on the surface enable targeted delivery to the TRPV1 channels to the cell membrane. Enhanced targetability might enable the intravenous delivery of nanotransducers, reducing invasiveness.
The reviewed work has provided valuable insights into tetherless optical neuromodulation tools and strategies for controlling neural activity in vivo, particularly in rats. A notable related study even demonstrated minimally invasive optogenetic stimulation of macaque cortical neurons via transdural illumination. Taking advantage of the eye's exceptional light transmission properties, the photosensitive protein ChrimsonR was transfected into the foveal retinal ganglion cells of a patient with retinitis pigmentosa in 2021, marking a groundbreaking milestone [127]. This pioneering application of optogenetic treatment for the disease stands as the first-in-human report to date [128].
Looking ahead to future clinical applications, the potential applications of tetherless optical neuromodulation in deep brain regions are promising. These include modulating dopamine release from the VTA, and improving movement disorders in Parkinson's disease. In addition, the special anatomical features of the spinal cord and spine may make the spinal cord a prominent target for wireless neuromodulation. Moreover, the treatment of various non-neurological diseases may also benefit from these emerging technologies, including atherosclerosis attenuation [129] and specific cancer therapies [130].
Nonetheless, applying these techniques to larger mammals, especially primates with their larger brain volumes and deeper brain tissues, presents formidable challenges. Overcoming these hurdles necessitates extensive interdisciplinary collaborations and the exploration of innovative approaches to optical tool development. In particular, there is a pressing need to enhance the efficiency and safety of photosensitive protein transfection, along with improvements in the conversion efficiency, biocompatibility, and precision targeting of mediating nanomaterials. To broaden the horizons of potential solutions, the exploration of different modal stimuli converted into light, such as X-ray to light [131], or the conversion of long wavelengths of light into different modalities, even embracing multimodal stimuli [132], can provide a wealth of opportunities to surmount these challenges. Moreover, it is anticipated that future advancements in these techniques will synergistically complement one another.
References
Yang C, Park S. Nanomaterials-assisted thermally induced neuromodulation. Biomed Eng Lett 2021, 11: 163–170.
Jiang S, Wu X, Rommelfanger NJ, Ou Z, Hong G. Shedding light on neurons: Optical approaches for neuromodulation. Natl Sci Rev 2022, 9: nwac007.
Sternson SM, Roth BL. Chemogenetic tools to interrogate brain functions. Annu Rev Neurosci 2014, 37: 387–407.
Armbruster BN, Li X, Pausch MH, Herlitze S, Roth BL. Evolving the lock to fit the key to create a family of G protein-coupled receptors potently activated by an inert ligand. Proc Natl Acad Sci U S A 2007, 104: 5163–5168.
Poth KM, Texakalidis P, Boulis NM. Chemogenetics: Beyond lesions and electrodes. Neurosurgery 2021, 89: 185–195.
Sebesta C, Torres Hinojosa D, Wang B, Asfouri J, Li Z, Duret G. Subsecond multichannel magnetic control of select neural circuits in freely moving flies. Nat Mater 2022, 21: 951–958.
Hescham SA, Chiang PH, Gregurec D, Moon J, Christiansen MG, Jahanshahi A, et al. Magnetothermal nanoparticle technology alleviates parkinsonian-like symptoms in mice. Nat Commun 2021, 12: 5569.
Chen R, Romero G, Christiansen MG, Mohr A, Anikeeva P. Wireless magnetothermal deep brain stimulation. Science 2015, 347: 1477–1480.
Wu S, Li H, Wang D, Zhao L, Qiao X, Zhang X, et al. Genetically magnetic control of neural system via TRPV4 activation with magnetic nanoparticles. Nano Today 2021, 39: 101187.
Su CL, Cheng CC, Yen PH, Huang JX, Ting YJ, Chiang PH. Wireless neuromodulation in vitro and in vivo by intrinsic TRPC-mediated magnetomechanical stimulation. Commun Biol 2022, 5: 1166.
Lee JU, Shin W, Lim Y, Kim J, Kim WR, Kim H, et al. Non-contact long-range magnetic stimulation of mechanosensitive ion channels in freely moving animals. Nat Mater 2021, 20: 1029–1036.
Fenno L, Yizhar O, Deisseroth K. The development and application of optogenetics. Annu Rev Neurosci 2011, 34: 389–412.
Karl D. Optogenetics: 10 years of microbial opsins in neuroscience. Nat Neurosci 2015, 18: 1213–1225.
Boyden ES, Zhang F, Bamberg E, Nagel G, Deisseroth K. Millisecond-timescale, genetically targeted optical control of neural activity. Nat Neurosci 2005, 8: 1263–1268.
Häusser M. Optogenetics - the might of light. N Engl J Med 2021, 385: 1623–1626.
Keshmiri Neghab H, Soheilifar MH, Grusch M, Ortega MM, Esmaeeli Djavid G, Saboury AA, et al. The state of the art of biomedical applications of optogenetics. Lasers Surg Med 2022, 54: 202–216.
Guo F, Du Y, Qu FH, Lin SD, Chen Z, Zhang SH. Dissecting the neural circuitry for pain modulation and chronic pain: Insights from optogenetics. Neurosci Bull 2022, 38: 440–452.
Fougère M, van der Zouwen CI, Boutin J, Neszvecsko K, Sarret P, Ryczko D. Optogenetic stimulation of glutamatergic neurons in the cuneiform nucleus controls locomotion in a mouse model of Parkinson’s disease. Proc Natl Acad Sci U S A 2021, 118: e2110934118.
Etter G, van der Veldt S, Manseau F, Zarrinkoub I, Trillaud-Doppia E, Williams S. Optogenetic gamma stimulation rescues memory impairments in an Alzheimer’s disease mouse model. Nat Commun 2019, 10: 5322.
Choi S, Hachisuka J, Brett MA, Magee AR, Omori Y, Iqbal NUA, et al. Parallel ascending spinal pathways for affective touch and pain. Nature 2020, 587: 258–263.
Chen G, Lai S, Bao G, Ke J, Meng X, Lu S, et al. Distinct reward processing by subregions of the nucleus accumbens. Cell Rep 2023, 42: 112069.
Bansal A, Shikha S, Zhang Y. Towards translational optogenetics. Nat. Biomed Eng 2023, 7: 349–369.
Yaroslavsky AN, Schulze PC, Yaroslavsky IV, Schober R, Ulrich F, Schwarzmaier HJ. Optical properties of selected native and coagulated human brain tissues in vitro in the visible and near infrared spectral range. Phys Med Biol 2002, 47: 2059–2073.
Tedford CE, DeLapp S, Jacques S, Anders J. Quantitative analysis of transcranial and intraparenchymal light penetration in human cadaver brain tissue. Lasers Surg Med 2015, 47: 312–322.
Li T, Xue C, Wang P, Li Y, Wu L. Photon penetration depth in human brain for light stimulation and treatment: A realistic Monte Carlo simulation study. J Innov Opt Health Sci 2017, 10: 1743002.
Xu HT, Pan F, Yang G, Gan WB. Choice of cranial window type for in vivo imaging affects dendritic spine turnover in the cortex. Nat Neurosci 2007, 10: 549–551.
Polikov VS, Tresco PA, Reichert WM. Response of brain tissue to chronically implanted neural electrodes. J Neurosci Methods 2005, 148: 1–18.
Montgomery KL, Yeh AJ, Ho JS, Tsao V, Mohan Iyer S, Grosenick L, et al. Wirelessly powered, fully internal optogenetics for brain, spinal and peripheral circuits in mice. Nat Methods 2015, 12: 969–974.
All AH, Zeng X, Teh DBL, Yi Z, Prasad A, Ishizuka T, et al. Expanding the toolbox of upconversion nanoparticles for in vivo optogenetics and neuromodulation. Adv Mater 2019, 31: e1803474.
Wu X, Jiang Y, Rommelfanger NJ, Yang F, Zhou Q, Yin R, et al. Tether-free photothermal deep-brain stimulation in freely behaving mice via wide-field illumination in the near-infrared-II window. Nat Biomed Eng 2022, 6: 754–770.
Jiang Y, Huang Y, Luo X, Wu J, Zong H, Shi L, et al. Neural stimulation in vitro and in vivo by photoacoustic nanotransducers. Matter 2021, 4: 654–674.
Parameswaran R, Carvalho-de-Souza JL, Jiang Y, Burke MJ, Zimmerman JF, Koehler K, et al. Photoelectrochemical modulation of neuronal activity with free-standing coaxial silicon nanowires. Nat Nanotechnol 2018, 13: 260–266.
Chen B, Cui M, Wang Y, Shi P, Wang H, Wang F. Recent advances in cellular optogenetics for photomedicine. Adv Drug Deliv Rev 2022, 188: 114457.
Zhang F, Prigge M, Beyrière F, Tsunoda SP, Mattis J, Yizhar O, et al. Red-shifted optogenetic excitation: A tool for fast neural control derived from Volvox carteri. Nat Neurosci 2008, 11: 631–633.
Yizhar O, Fenno LE, Prigge M, Schneider F, Davidson TJ, O’Shea DJ, et al. Neocortical excitation/inhibition balance in information processing and social dysfunction. Nature 2011, 477: 171–178.
Govorunova EG, Spudich EN, Lane CE, Sineshchekov OA, Spudich JL. New channelrhodopsin with a red-shifted spectrum and rapid kinetics from Mesostigma viride. mBio 2011, 2: e00115-e111.
Lin JY, Knutsen PM, Muller A, Kleinfeld D, Tsien RY. ReaChR: A red-shifted variant of channelrhodopsin enables deep transcranial optogenetic excitation. Nat Neurosci 2013, 16: 1499–1508.
Klapoetke NC, Murata Y, Kim SS, Pulver SR, Birdsey-Benson A, Cho YK, et al. Independent optical excitation of distinct neural populations. Nat Methods 2014, 11: 338–346.
Marshel JH, Kim YS, Machado TA, Quirin S, Benson B, Kadmon J, et al. Cortical layer-specific critical dynamics triggering perception. Science 2019, 365: eaaw5202.
Chen R, Gore F, Nguyen QA, Ramakrishnan C, Patel S, Kim SH, et al. Deep brain optogenetics without intracranial surgery. Nat Biotechnol 2021, 39: 161–164.
Kishi KE, Kim YS, Fukuda M, Inoue M, Kusakizako T, Wang PY, et al. Structural basis for channel conduction in the pump-like channelrhodopsin ChRmine. Cell 2022, 185: 672-689.e23.
Zhang F, Aravanis AM, Adamantidis A, de Lecea L, Deisseroth K. Circuit-breakers: Optical technologies for probing neural signals and systems. Nat Rev Neurosci 2007, 8: 577–581.
Gradinaru V, Zhang F, Ramakrishnan C, Mattis J, Prakash R, Diester I, et al. Molecular and cellular approaches for diversifying and extending optogenetics. Cell 2010, 141: 154–165.
Chuong AS, Miri ML, Busskamp V, Matthews GAC, Acker LC, Sørensen AT, et al. Noninvasive optical inhibition with a red-shifted microbial rhodopsin. Nat Neurosci 2014, 17: 1123–1129.
Govorunova EG, Sineshchekov OA, Li H, Wang Y, Brown LS, Spudich JL. RubyACRs, nonalgal anion channelrhodopsins with highly red-shifted absorption. Proc Natl Acad Sci U S A 2020, 117: 22833–22840.
Zhang F, Vierock J, Yizhar O, Fenno LE, Tsunoda S, Kianianmomeni A, et al. The microbial opsin family of optogenetic tools. Cell 2011, 147: 1446–1457.
Karasuyama M, Inoue K, Nakamura R, Kandori H, Takeuchi I. Understanding colour tuning rules and predicting absorption wavelengths of microbial rhodopsins by data-driven machine-learning approach. Sci Rep 2018, 8: 15580.
Inoue K, Karasuyama M, Nakamura R, Konno M, Yamada D, Mannen K, et al. Exploration of natural red-shifted rhodopsins using a machine learning-based Bayesian experimental design. Commun Biol 2021, 4: 362.
Bedbrook CN, Yang KK, Robinson JE, MacKey ED, Gradinaru V, Arnold FH. Machine learning-guided channelrhodopsin engineering enables minimally invasive optogenetics. Nat Methods 2019, 16: 1176–1184.
Lin JY. A user’s guide to channelrhodopsin variants: Features, limitations and future developments. Exp Physiol 2011, 96: 19–25.
Gong X, Mendoza-Halliday D, Ting JT, Kaiser T, Sun X, Bastos AM, et al. An ultra-sensitive step-function opsin for minimally invasive optogenetic stimulation in mice and macaques. Neuron 2020, 107: 197.
Chan MH, Huang WT, Chen KC, Su TY, Chan YC, Hsiao M, et al. The optical research progress of nanophosphors composed of transition elements in the fourth period of near-infrared windows I and II for deep-tissue theranostics. Nanoscale 2022, 14: 7123–7136.
Ma Y, Bao J, Zhang Y, Li Z, Zhou X, Wan C, et al. Mammalian near-infrared image vision through injectable and self-powered retinal nanoantennae. Cell 2019, 177: 243-255.e15.
Luo DG, Yue WWS, Ala-Laurila P, Yau KW. Activation of visual pigments by light and heat. Science 2011, 332: 1307–1312.
Ala-Laurila P, Albert RJ, Saarinen P, Koskelainen A, Donner K. The thermal contribution to photoactivation in A2 visual pigments studied by temperature effects on spectral properties. Vis Neurosci 2003, 20: 411–419.
Auzel F. Upconversion and anti-Stokes processes with f and d ions in solids. Chem Rev 2004, 104: 139–173.
Wang HQ, Batentschuk M, Osvet A, Pinna L, Brabec CJ. Rare-earth ion doped up-conversion materials for photovoltaic applications. Adv Mater 2011, 23: 2675–2680.
Wang F, Liu X. Recent advances in the chemistry of lanthanide-doped upconversion nanocrystals. Chem Soc Rev 2009, 38: 976–989.
Yi Z, All AH, Liu X. Upconversion nanoparticle-mediated optogenetics. Adv Exp Med Biol 2021, 1293: 641–657.
Wen S, Zhou J, Zheng K, Bednarkiewicz A, Liu X, Jin D. Advances in highly doped upconversion nanoparticles. Nat Commun 2018, 9: 2415.
Qin X, Xu J, Wu Y, Liu X. Energy-transfer editing in lanthanide-activated upconversion nanocrystals: A toolbox for emerging applications. ACS Cent Sci 2019, 5: 29–42.
Meir R, Hirschhorn T, Kim S, Fallon KJ, Churchill EM, Wu D, et al. Photon upconversion hydrogels for 3D optogenetics. Adv Funct Materials 2021, 31: 2010907.
Liu Y, Yi Z, Yao Y, Guo B, Liu X. Noninvasive manipulation of ion channels for neuromodulation and theranostics. Acc Mater Res 2022, 3: 247–258.
Liu Y, Lu Y, Yang X, Zheng X, Wen S, Wang F, et al. Amplified stimulated emission in upconversion nanoparticles for super-resolution nanoscopy. Nature 2017, 543: 229–233.
Fu X, Fu S, Lu Q, Zhang J, Wan P, Liu J, et al. Excitation energy mediated cross-relaxation for tunable upconversion luminescence from a single lanthanide ion. Nat Commun 2022, 13: 4741.
Fan Q, Sun C, Hu B, Wang Q. Recent advances of lanthanide nanomaterials in Tumor NIR fluorescence detection and treatment. Mater Today Bio 2023, 20: 100646.
Dong H, Sun LD, Yan CH. Lanthanide-doped upconversion nanoparticles for super-resolution microscopy. Front Chem 2021, 8: 619377.
Deisseroth K, Anikeeva P. Upconversion of light for use in optogenetic methods: U.S. Patent 9,522,288[P]. 2016-12-20.
Shah S, Liu JJ, Pasquale N, Lai J, McGowan H, Pang ZP, et al. Hybrid upconversion nanomaterials for optogenetic neuronal control. Nanoscale 2015, 7: 16571–16577.
Bansal A, Liu H, Jayakumar MKG, Andersson-Engels S, Zhang Y. Quasi-continuous wave near-infrared excitation of upconversion nanoparticles for optogenetic manipulation of C. elegans. Small 2016, 12: 1732–1743.
Ai X, Lyu L, Zhang Y, Tang Y, Mu J, Liu F, et al. Remote regulation of membrane channel activity by site-specific localization of lanthanide-doped upconversion nanocrystals. Angew Chem Int Ed Engl 2017, 56: 3031–3035.
Wang Y, Lin X, Chen X, Chen X, Xu Z, Zhang W, et al. Tetherless near-infrared control of brain activity in behaving animals using fully implantable upconversion microdevices. Biomaterials 2017, 142: 136–148.
Wang Y, Xie K, Yue H, Chen X, Luo X, Liao Q, et al. Flexible and fully implantable upconversion device for wireless optogenetic stimulation of the spinal cord in behaving animals. Nanoscale 2020, 12: 2406–2414.
Chen S, Weitemier AZ, Zeng X, He L, Wang X, Tao Y, et al. Near-infrared deep brain stimulation via upconversion nanoparticle-mediated optogenetics. Science 2018, 359: 679–684.
Miyazaki T, Chowdhury S, Yamashita T, Matsubara T, Yawo H, Yuasa H, et al. Large timescale interrogation of neuronal function by fiberless optogenetics using lanthanide micro-particles. Cell Rep 2019, 26: 1033-1043.e5.
Mei Q, Bansal A, Jayakumar MKG, Zhang Z, Zhang J, Huang H, et al. Manipulating energy migration within single lanthanide activator for switchable upconversion emissions towards bidirectional photoactivation. Nat Commun 2019, 10: 4416.
Liu X, Chen H, Wang Y, Si Y, Zhang H, Li X, et al. Near-infrared manipulation of multiple neuronal populations via trichromatic upconversion. Nat Commun 2021, 12: 5662.
Lin X, Sun T, Tang M, Yang A, Yan-Do R, Chen D, et al. 3D upconversion barcodes for combinatory wireless neuromodulation in behaving animals. Adv Healthc Mater 2022, 11: e2200304.
Yan J, Wan Y, Ji Z, Li C, Tao C, Tang Y, et al. Motor neuron-specific membrane depolarization of transected peripheral nerves by upconversion nanoparticle-mediated optogenetics. Adv Funct Materials 2023, 33: 2303992.
Matsubara T, Yamashita T. Remote optogenetics using up/down-conversion phosphors. Front Mol Biosci 2021, 8: 771717.
Chen J, Gong M, Fan Y, Feng J, Han L, Xin HL, et al. Collective plasmon coupling in gold nanoparticle clusters for highly efficient photothermal therapy. ACS Nano 2022, 16: 910–920.
Kokalari I, Keshavan S, Rahman M, Gazzano E, Barzan G, Mandrile L, et al. Efficacy, biocompatibility and degradability of carbon nanoparticles for photothermal therapy of lung cancer. Nanomedicine 2021, 16: 689–707.
Wang W, Zhang X, Ni X, Zhou W, Xie C, Huang W, et al. Semiconducting polymer nanoparticles for NIR-II fluorescence imaging-guided photothermal/thermodynamic combination therapy. Biomater Sci 2022, 10: 846–853.
Attia ABE, Balasundaram G, Moothanchery M, Dinish US, Bi R, Ntziachristos V, et al. A review of clinical photoacoustic imaging: Current and future trends. Photoacoustics 2019, 16: 100144.
Fan R, Chen C, Hou H, Chuan D, Mu M, Liu Z, et al. Tumor acidity and near-infrared light responsive dual drug delivery polydopamine-based nanoparticles for chemo-photothermal therapy. Adv Funct Materials 2021, 31: 2009733.
Molaei MJ. Two-dimensional (2D) materials beyond graphene in cancer drug delivery, photothermal and photodynamic therapy, recent advances and challenges ahead: A review. J Drug Deliv Sci Tec 2021, 61: 101830.
O’Neal DP, Hirsch LR, Halas NJ, Payne JD, West JL. Photo-thermal tumor ablation in mice using near infrared-absorbing nanoparticles. Cancer Lett 2004, 209: 171–176.
Xu C, Pu K. Second near-infrared photothermal materials for combinational nanotheranostics. Chem Soc Rev 2021, 50: 1111–1137.
Yin C, Li X, Wang Y, Liang Y, Zhou S, Zhao P, et al. Organic semiconducting macromolecular dyes for NIR-II photoacoustic imaging and photothermal therapy. Adv Funct Materials 2021, 31: 2104650.
Zhen X, Pu K, Jiang X. Photoacoustic imaging and photothermal therapy of semiconducting polymer nanoparticles: Signal amplification and second near-infrared construction. Small 2021, 17: e2004723.
Carvalho-de-Souza JL, Pinto BI, Pepperberg DR, Bezanilla F. Optocapacitive generation of action potentials by microsecond laser pulses of nanojoule energy. Biophys J 2018, 114: 283–288.
Shapiro MG, Homma K, Villarreal S, Richter CP, Bezanilla F. Infrared light excites cells by changing their electrical capacitance. Nat Commun 2012, 3: 736.
Carvalho-de-Souza JL, Treger JS, Dang B, Kent SBH, Pepperberg DR, Bezanilla F. Photosensitivity of neurons enabled by cell-targeted gold nanoparticles. Neuron 2015, 86: 207–217.
Jiang Y, Li X, Liu B, Yi J, Fang Y, Shi F, et al. Rational design of silicon structures for optically controlled multiscale biointerfaces. Nat Biomed Eng 2018, 2: 508–521.
Castillo K, Diaz-Franulic I, Canan J, Gonzalez-Nilo F, Latorre R. Thermally activated TRP channels: Molecular sensors for temperature detection. Phys Biol 2018, 15: 021001.
Hilton JK, Kim M, Van Horn WD. Structural and evolutionary insights point to allosteric regulation of TRP ion channels. Acc Chem Res 2019, 52: 1643–1652.
Randhawa PK, Jaggi AS. TRPV1 and TRPV4 channels: Potential therapeutic targets for ischemic conditioning-induced cardioprotection. Eur J Pharmacol 2015, 746: 180–185.
Caterina MJ, Schumacher MA, Tominaga M, Rosen TA, Levine JD, Julius D. The capsaicin receptor: A heat-activated ion channel in the pain pathway. Nature 1997, 389: 816–824.
Cao E, Liao M, Cheng Y, Julius D. TRPV1 structures in distinct conformations reveal activation mechanisms. Nature 2013, 504: 113–118.
Liao M, Cao E, Julius D, Cheng Y. Structure of the TRPV1 ion channel determined by electron cryo-microscopy. Nature 2013, 504: 107–112.
Laursen WJ, Schneider ER, Merriman DK, Bagriantsev SN, Gracheva EO. Low-cost functional plasticity of TRPV1 supports heat tolerance in squirrels and camels. Proc Natl Acad Sci U S A 2016, 113: 11342–11347.
Luo L, Wang Y, Li B, Xu L, Kamau PM, Zheng J, et al. Molecular basis for heat desensitization of TRPV1 ion channels. Nat Commun 2019, 10: 2134.
Yao J, Liu B, Qin F. Modular thermal sensors in temperature-gated transient receptor potential (TRP) channels. Proc Natl Acad Sci U S A 2011, 108: 11109–11114.
Kauer JA, Gibson HE. Hot flash: TRPV channels in the brain. Trends Neurosci 2009, 32: 215–224.
Nakatsuji H, Numata T, Morone N, Kaneko S, Mori Y, Imahori H, et al. Thermosensitive ion channel activation in single neuronal cells by using surface-engineered plasmonic nanoparticles. Angew Chem Int Ed Engl 2015, 54: 11725–11729.
Lyu Y, Xie C, Chechetka SA, Miyako E, Pu K. Semiconducting polymer nanobioconjugates for targeted photothermal activation of neurons. J Am Chem Soc 2016, 138: 9049–9052.
Nelidova D, Morikawa RK, Cowan CS, Raics Z, Goldblum D, Scholl HPN, et al. Restoring light sensitivity using tunable near-infrared sensors. Science 2020, 368: 1108–1113.
Ge X, Fu Q, Bai L, Chen B, Wang R, Gao S, et al. Photoacoustic imaging and photothermal therapy in the second near-infrared window. New J Chem 2019, 43: 8835–8851.
Lei Z, Zhang W, Li B, Guan G, Huang X, Peng X, et al. A full-spectrum-absorption from nickel sulphide nanoparticles for efficient NIR-II window photothermal therapy. Nanoscale 2019, 11: 20161–20170.
Wang C, Dai C, Hu Z, Li H, Yu L, Lin H, et al. Photonic cancer nanomedicine using the near infrared-II biowindow enabled by biocompatible titanium nitride nanoplatforms. Nanoscale Horiz 2019, 4: 415–425.
Liu J, Li J, Zhang S, Ding M, Yu N, Li J, et al. Antibody-conjugated gold nanoparticles as nanotransducers for second near-infrared photo-stimulation of neurons in rats. Nano Converg 2022, 9: 13.
Yoo S, Hong S, Choi Y, Park JH, Nam Y. Photothermal inhibition of neural activity with near-infrared-sensitive nanotransducers. ACS Nano 2014, 8: 8040–8049.
An Y, Nam Y. Closed-loop control of neural spike rate of cultured neurons using a thermoplasmonics-based photothermal neural stimulation. J Neural Eng 2021, 18. doi: https://doi.org/10.1088/1741-2552/ac3265.
Gholami Derami H, Gupta P, Weng KC, Seth A, Gupta R, Silva JR, et al. Reversible Photothermal Modulation of Electrical Activity of Excitable Cells using Polydopamine Nanoparticles. Adv Mater 2021, 33: e2008809.
Liu X, Qiao Z, Chai Y, Zhu Z, Wu K, Ji W, et al. Nonthermal and reversible control of neuronal signaling and behavior by midinfrared stimulation. Proc Natl Acad Sci U S A 2021, 118: e2015685118.
Zhang J, He Y, Liang S, Liao X, Li T, Qiao Z, et al. Non-invasive, opsin-free mid-infrared modulation activates cortical neurons and accelerates associative learning. Nat Commun 2021, 12: 2730.
Doroudchi MM, Greenberg KP, Liu J, Silka KA, Boyden ES, Lockridge JA, et al. Virally delivered Channelrhodopsin-2 Safely and Effectively Restores Visual Function in Multiple Mouse Models of Blindness. Mol Ther 2011, 19: 1220–1229.
Sugano E, Isago H, Wang Z, Murayama N, Tamai M, Tomita H. Immune responses to adeno-associated virus type 2 encoding channelrhodopsin-2 in a genetically blind rat model for gene therapy. Gene Ther 2011, 18: 266–274.
Chow BY, Boyden ES. Optogenetics and translational medicine. Sci Transl Med 2013, 5: 177ps5.
Gundelach LA, Hüser MA, Beutner D, Ruther P, Bruegmann T. Towards the clinical translation of optogenetic skeletal muscle stimulation. Pflugers Arch 2020, 472: 527–545.
Chan KY, Jang MJ, Yoo BB, Greenbaum A, Ravi N, Wu WL, et al. Engineered AAVs for efficient noninvasive gene delivery to the central and peripheral nervous systems. Nat Neurosci 2017, 20: 1172–1179.
Rzechorzek NM, Thrippleton MJ, Chappell FM, Mair G, Ercole A, Cabeleira M, et al. A daily temperature rhythm in the human brain predicts survival after brain injury. Brain 2022, 145: 2031–2048.
Owen SF, Liu MH, Kreitzer AC. Thermal constraints on in vivo optogenetic manipulations. Nat Neurosci 2019, 22: 1061–1065.
Kaiser M, Würth C, Kraft M, Hyppänen I, Soukka T, Resch-Genger U. Power-dependent upconversion quantum yield of NaYF4: Yb3+, Er3+ nano- and micrometer-sized particles - measurements and simulations. Nanoscale 2017, 9: 10051–10058.
Zheng W, Huang P, Tu D, Ma E, Zhu H, Chen X. Lanthanide-doped upconversion nano-bioprobes: Electronic structures, optical properties, and biodetection. Chem Soc Rev 2015, 44: 1379–1415.
Mattis J, Tye KM, Ferenczi EA, Ramakrishnan C, O’Shea DJ, Prakash R, et al. Principles for applying optogenetic tools derived from direct comparative analysis of microbial opsins. Nat Methods 2011, 9: 159–172.
Sahel JA, Boulanger-Scemama E, Pagot C, Arleo A, Galluppi F, Martel JN, et al. Partial recovery of visual function in a blind patient after optogenetic therapy. Nat Med 2021, 27: 1223–1229.
Harris AR, Gilbert F. Restoring vision using optogenetics without being blind to the risks. Graefes Arch Clin Exp Ophthalmol 2022, 260: 41–45.
Gao W, Sun Y, Cai M, Zhao Y, Cao W, Liu Z, et al. Copper sulfide nanoparticles as a photothermal switch for TRPV1 signaling to attenuate atherosclerosis. Nat Commun 2018, 9: 231.
Ma Z, Zhang J, Zhang W, Foda MF, Zhang Y, Ge L, et al. Intracellular Ca2+ cascade guided by NIR-II photothermal switch for specific tumor therapy. iScience 2020, 23: 101049.
Bartley AF, Fischer M, Bagley ME, Barnes JA, Burdette MK, Cannon KE, et al. Feasibility of cerium-doped LSO particles as a scintillator for X-ray induced optogenetics. J Neural Eng 2021, 18: https://doi.org/10.1088/1741. https://doi.org/10.1088/2552/abef89.
Sun F, Shen H, Yang Q, Yuan Z, Chen Y, Guo W, et al. Dual behavior regulation: Tether-free deep-brain stimulation by photothermal and upconversion hybrid nanoparticles. Adv Mater 2023, 35: e2210018.
Acknowledgments
This review was supported by China Postdoctoral Science Foundation (2022M723356), "From 0 to 1" Original Innovation Project of the Basic Frontier Scientific Research Program of the Chinese Academy of Sciences (29J20-015-III), Chinese Academy of Sciences 100 Talents Project: Research on Task oriented Functional Brain Development of Infants (29J20-052-III) and Natural Science Basic Research Plan in Shaanxi Province of China (2022JQ544).
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
The authors declare that there are no conflicts of interest.
Rights and permissions
Springer Nature or its licensor (e.g. a society or other partner) holds exclusive rights to this article under a publishing agreement with the author(s) or other rightsholder(s); author self-archiving of the accepted manuscript version of this article is solely governed by the terms of such publishing agreement and applicable law.
About this article

Cite this article
Sun, C., Fan, Q., Xie, R. et al. Tetherless Optical Neuromodulation: Wavelength from Orange-red to Mid-infrared. Neurosci. Bull. 40, 1173–1188 (2024). https://doi.org/10.1007/s12264-024-01179-1
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s12264-024-01179-1