
Abstract
Over the past 6 years, the New Hampshire (NH) Department of Environmental Services has shifted its preference for shoreline stabilization from traditional engineered shorelines (e.g., seawalls, concrete armoring) to nature-based living shoreline (LS) solutions. To improve the expectations and outcomes of future projects, we monitored three LS pilot projects in the Great Bay Estuary of NH from 2019 to 2022, estimated short-term recovery of the soil biogeochemistry, plant community, and habitat use by fauna, and documented adaptive management needs. Each LS was paired with a nearby (< 200 m) reference salt marsh and a degraded shoreline. After 4 years, halophyte cover had recovered 25–75% in the low marsh and 26–70% in the high marsh. Creation of salt marsh habitat supported similar mummichog population abundances (10–24 indiv. per trap) similar to reference marshes and substantially greater than no action control shorelines (0–1 indiv. per trap). Aerobic or mildly anaerobic reduction–oxidation potentials in the low marsh (14–302 mV) and high marsh (243–266 V) were attributed to rapid drainage of the marsh in two of the LS projects. A novel planting technique of spreading a turf with pre-installed graminoid plugs across the marsh surface at one of the sites may have jumpstarted development of anerobic soil conditions within 3 years (Low: − 192.7 ± 14.9 mV, High: − 119.0 ± 25.3 mV). Opportunistic foraging waterfowl and burial by wrack led to annual replacement replanting and seasonal wrack removal as adaptive management needs of these and future projects. LS projects in the Great Bay Estuary provide functional salt marsh habitat and improved shoreline resilience that can serve as a valuable strategy for coastal restoration.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Community planners, conservation practitioners, coastal ecologists, and regulators have promoted the construction of nature-based solutions over traditional grey infrastructure since the mid-2000s to improve both shoreline resilience and ecosystem services provided by coastal habitats (National Research Council 2007; Bridges et al. 2014). Living shorelines (LSs) have been promoted as a soft engineering approach that utilizes ecological restoration or creation of wetland systems (e.g., salt marshes, tidal wetlands, oyster reefs, and sand dunes) to stabilize shorelines, connect aquatic and terrestrial systems, and improve ecosystem services (Bilkovic et al. 2017). LSs combine the inherent resiliency of coastal ecosystems with the strength and wave attenuation properties of hardened structures. One widely implemented LS design is the creation of a fringe salt marsh and the placement of a seaward sill composed of root wads, riprap boulders, or coir fiber logs at mean tide level (i.e., low marsh edge) (O’Donnell 2017; Woods Hole Group 2017). The sill prevents immediate loss of fill sediment, reduces initial vegetation propagule mortality, and provides salt marsh grasses time to stabilize the sediment and develop a dense network of peat that is resilient to forces of erosion.
LS projects have been shown to successfully enhance coastal resiliency and abate coastal erosion, one of the main considerations for their implementation. The addition of the structural sill improves upon the fringe marsh’s capacity to attenuate wave energy (Leone and Tahvildari 2023), which has been shown to reduce wave height by at least 50% within the first 10 m (Morgan et al. 2002; Shepard et al. 2011). From a resiliency perspective, LSs recovered both vegetation and surface elevation losses after high-intensity hurricanes better than natural fringe marshes or traditional armoring (Gittman et al. 2014; Smith et al. 2018). Additionally, these projects have maintained pace with long-term sea-level rise through surface sediment accretion and lateral shoreline growth (Polk and Eulie 2018; Palinkas et al. 2023). In North Carolina, Polk et al. (2021) observed stable or expanding marsh width (0.015 m yr−1) with LS projects compared to erosional loss of natural fringe marshes (− 0.31 m yr−1). The construction of several LS projects within an estuary system may have cumulative benefits for shoreline stabilization and ecosystem function and services (McClenachan et al. 2020; Diefenderfer et al. 2021).
The recovery of salt marsh flora, fauna, and soil characteristics as a gauge of restoration success has comprised the bulk of research on LSs. Planted species of Spartina alterniflora and S. patens in the mid-Atlantic and Southeast reached similar densities and canopy heights to local references within 5 years in the Southeast (Bilkovic and Mitchell 2013; Gittman et al. 2016). Other structural measures such as above ground biomass may require up to 10 years to fully recover (Chambers et al. 2021). Nekton take advantage of the new salt marsh habitat even if the vegetation has not fully recovered. Increases in nekton density and richness occur after only several months (Currin et al. 2008), potentially reaching or exceeding density of reference marshes within 10 years (Balouskus and Targett 2016; Isdell et al. 2021). LSs continue to serve as nurseries for nekton as juveniles make up a larger proportion of individuals than at reference fringe marshes (Currin et al. 2008; Gittman et al. 2016). The construction of the sill promotes the colonization of epibenthic filter feeders including barnacles and oysters (Bilkovic and Mitchell 2013); however, limited research observed certain key foundational species like Geukensia demissa (Ribbed mussel) are unlikely to colonize LSs (Bilkovic et al. 2021). Despite immediate high rates of carbon and nitrogen storage, wetland soil characteristics, such as low bulk density, high organic matter content, and consistent saturation, may require decades to develop (Davis et al. 2015; Chambers et al. 2021; Isdell et al. 2021).
LS projects have been gaining traction as a strategy to improve coastal resiliency and abate shoreline erosion. LSs have been widely constructed in the Gulf of Mexico, Southeast, and Mid-Atlantic (i.e., Chesapeake Bay) and the first projects in New England within the past decade (O’Donnell 2017; Smith et al. 2020; NOAA Restoration Center 2020). Published data on the performance of LSs is centered on North Carolina, Delaware, and the Chesapeake Bay region, where the growing season is longer (Kirwan et al. 2009) and ice rafting is not a concern compared to New England (Hardwick-Witman 1985). The New Hampshire Department of Environmental Services (NHDES) has steadily promoted the use of LSs by mapping ideal locations for their implementation (Balasubramanayam and Howard 2019) and shifting the regulatory-preferred shoreline stabilization method from grey infrastructure to LSs (NH Env-WT 609). LS designs may also incorporate adaptation for future acceleration of sea-level rise by elevating the lower seaward edge of the marsh to match sea-level rise projections for certain dates (e.g., 0.52 m above the lower edge of reference marshes for 2060) and re-grading the upper edge to match the slope of the marsh so marsh loss at the low edge is matched by migration gains at the upper edge.
In partnerships with localities and state regulators, since 2016 three LSs were constructed as pilot projects in New Hampshire’s Great Bay Estuary to restore salt marsh habitat and stabilize degraded or eroding shorelines. As some of the first projects in northern New England, questions remain about the efficacy, recovery timelines, and maintenance needs compared to more southern counterparts. With NHDES and municipalities planning for more LSs in the Great Bay Estuary, updated designs and accurate expectations from the lessons of prior projects can improve future success (Clewell and Aronson 2004). Research and documentation on stressors and adaptive management strategies of LSs have been underreported, despite acknowledgement that environmental stressors may dictate the success of projects well after construction. We evaluated the short-term restoration performance (0–4 years) of three LS salt marsh projects over four growing seasons using commonly monitored metrics of salt marsh structure and function including vegetation, nekton use, and intertidal soil development. The colonization of algae and invertebrates on the constructed sills was also monitored to quantify the ecological impact of the structural component. Potential challenges that could impact the efficacy of LSs in northern New England and adaptive management needs are documented.
Methods
Site Description
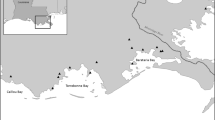
Three LSs were constructed in the Great Bay Estuary (NH) within relatively protected coves: North Mill Pond (NMP; 43.0761, -70.7661) in 2016, Cutts Cove (CC; 43.08442, -70.6568) in 2018, and Wagon Hill Farm (WHF; 43.1249, -70.8721) in 2019 (Table 1, Fig. 1a). Creation of new salt marsh habitat was conducted as mitigation from the impacts of infrastructure upgrades at NMP and CC (Burdick and Moore 2017; Burdick et al. 2021a). CC was constructed along a steep, riprap armored sill whereas NMP was built on top of a degraded salt marsh with relic peat substrate at some high marsh elevations. The WHF project was constructed to restore salt marsh habitat after substantial erosion (0.15–0.3 m yr−1; Burdick et al. 2021b) had converted over 90% of marsh width to mudflat and eroded upland property based on 1993 aerial photography and personal observations by authors (Ballestero, Burdick, and Moore) since 2000. Restoration goals for all three projects included (1) restore or enhance fringe salt marsh habitat and functions (CC, NMP, and WHF), (2) accommodate sea-level rise and marsh migration (CC, WHF), and (3) abate shoreline erosion to protect upland property (CC and WHF). Restoration of salt marsh habitat was evaluated based on vegetation structure, nekton use, and pore water chemistry, which are recommended metrics commonly measured for salt marsh restoration projects in New England (Neckles et al. 2002; Konisky et al. 2006).

a Local site map of living shoreline projects in the Great Bay Estuary of New Hampshire. Study sites included b North Mill Pond in Portsmouth, c Cutts Cove in Portsmouth, and d Wagon Hill Farm in Durham. Each site included a reference salt marsh, living shoreline, and no action control for a Control-Impact experimental design. Drone aerial imagery was collected for each site in 2020
LSs were planned to resemble small natural fringe marshes (< 100 m length) common in the Great Bay Estuary (Jones 2000), sited within a larger continuum of fringe marsh habitat to maximize restoration benefits. Projects were constructed similarly including regrading of slopes, sediment additions within the intertidal zone, construction of a structural sill, and plantings of salt marsh graminoids. At WHF and CC, the toe was protected with a 0.7–1.4 m (2–4 ft) tall riprap sill and plantings included S. alterniflora, S. patens, Distichlis spicata, and Juncus gerardii. NMP was only planted with S. alterniflora and S. patens and built with a coir fiber sill for erosion control at the lower marsh edge which had decomposed by 2019.
A control-impact experimental design was implemented at each site as each possessed a local reference salt marsh and a no action control shoreline (Fig. 1b–d). Restoration success of salt marsh habitat structure and function for LSs was compared to the reference marsh while the no action shoreline provided a gauge of the progress achieved since restoration. The local reference sites and no action control sites were located within 200 m and are influenced by comparable environmental conditions (i.e., fetch, salinity, tidal elevations, etc.) as the LSs. The reference and LS shorelines of CC are in different orientations; however, the difference in fetch is minimal (< 100 m) since both are located in a small, protected cove. Since pre-restoration monitoring was not possible, no action control shorelines were chosen to represent before conditions. No action control shorelines of CC and NMP were mudflats in front of historic riprap armoring of steep berms and WHF’s faced similar erosion loss as the LS shoreline pre-construction and had fully lost low marsh habitat.
Field Methods
The shorelines were divided between low and high marsh zones, based on vegetation patterns, and ten plots were randomly located within each marsh zone. The no action controls of NMP and CC represented only low marsh due to the elevation of the mudflats which could be sampled, while high marsh elevations were steep riprap slopes. Conversely, due to complete erosion loss of the low marsh at the no action control of WHF, it was considered only high marsh. Vegetation and pore water chemistry were sampled at plots during low tide in August–September: 2019–2020 NMP, 2019–2021 CC, and 2019, 2020, 2022 WHF. Monitoring of algae and macroinvertebrate colonization on the riprap sills was expanded to 2022 for CC and 2021 for WHF; however, only algae was monitored in 2022 for WHF.
Pore water (e.g., water held within pore spaces in the root zone of the substrate) was collected using a stainless-steel sipper with a plastic syringe inserted between 5 and 20 cm into the root zone at low tide (Portnoy and Giblin 1997a, b). Pore water was immediately analyzed in the field for reduction–oxidation potential (redox potential, mV) with an Orion 290A+ multimeter and redox probe equipped with a platinum electrode (Thermofisher, Waltham, MA, USA). Collected pore water samples of 10 mL were analyzed in the lab for salinity (psu) with a temperature-corrected optical refractometer and pH with an Orion 5000 multimeter and pH probe, respectively (Thermofisher, Waltham, MA, USA). A subsample of 0.5 mL of pore water was fixed in 2% Zinc Acetate solution in the field and then stored at 4 °C to determine sulfide content (mM) using a colorimetric analysis (Cline 1969) in a spectrophotometer (LaMotte, SmartSpec, Chestertown, MD, USA). Pore water could not be collected from most plots in the LSs due to lack of peat development and rapid drainage in the sandy soils. In these cases, soil redox potential was measured with brightened platinum electrodes inserted to a depth of 10 cm and allowed to equilibrate in situ for 45 min before measuring with a double junction reference electrode and millivolt meter (McKee et al. 1988). A correction of +244.0 mV was added to the half-cell potentiometric readings to arrive at in-situ redox Eh and allow for comparison with pore water redox values (McKee et al. 1988). Due to shifts in monitoring needs, only soil reduction–oxidation potential was monitored for Cutts Cove in 2021.
Since pore water was not present in the majority of plots, soil cores were collected in the LSs to better understand the early development of wetland soil characteristics including water content (% water), bulk density (g cm−1), and organic matter content (%). Soil cores were taken just outside of each 0.5 m2 plot with a 3.6 cm diameter corer (10 cm in length) in late September–early October 2020. The compaction rate of each core was immediately assessed and only the actual 10 cm of the soil column kept. Cores were weighed at the lab and stored at 4 °C. Cores were dried at 50 °C until the weight of the samples remained constant. Loss-on-ignition was conducted on a subsample of each core at 450 °C for four hours to calculate the organic matter content (Craft et al. 1991; Morgan et al. 2009).
All vegetation was identified to species in the field and cover was visually estimated for each species to the nearest 1% within 0.5 m2 quadrats (Neckles et al. 2002) with differentiation between halophyte and non-halophyte species following Tiner (2009). The average halophyte cover and species richness per plot were calculated for the low and high marsh zones. An additional ten random 0.5 m2 plots were sampled on the riprap and coir fiber sills of the LSs to better understand the timeline of algae and macroinvertebrate colonization on the structural component (Pollock 1998; Tiner 2009; Bilkovic and Mitchell 2013). Visual cover of each algae species was estimated. All macroinvertebrates were identified and counted on surfaces and within crevices of the sills within the two-dimensional boundaries of the quadrat.
Use of the salt marsh surface by the nekton community was assessed once over two consecutive spring tide periods in mid-September to early October in 2020 NMP, 2020–2021 CC, and 2020 and 2022 WHF. Nekton sampling at NMP shifted to early October due to Hurricane Teddy (September 23rd, 2020). The early fall sampling was timed to capture a snapshot of the density and size class of resident nekton such as Fundulus heteroclitus (Mummichog) and F. majalis (Striped killifish) (Raposa and Roman 2006). The nekton community was sampled with unbaited Gee’s standard wire mesh minnow traps, a common method of sampling salt marsh nekton community (West and Zedler 2000; Carlisle et al. 2002). Ten minnow traps were staggered along the width and equally spaced along the length of the low and high marsh zones. Traps were left out for 30 min with at least three-quarters of the trap height submerged the entire time (Kneib and Craig 2001). All nekton were identified to species, sex determined, and measured for total length (mm). For traps with high catch rates, a subset of the first ten males, females, and juveniles of each species were measured for total length. Fundulus sp. less than 45 mm were classified as juvenile. The CC and NMP sites were monitored in the low marsh (no high marsh existed in the no action controls). The WHF sites were monitored for both the low and high marsh to account for the no action control shoreline at WHF and for comparison across LSs. It is assumed that the close proximity of the shoreline treatments will not impact nekton catch rates, since resident nekton Fundulus sp. have smaller geographical homeranges during spawning and recruitment (Meyer and Posey 2014) and are highly selective of intertidal habitat at scales of 10–20 m (Crum et al. 2018).
Adaptive Management Surveys
To better understand potential maintenance issues and adaptive management needs, site surveys were completed 3–4 times throughout the year to document the full range of potential stressors on marsh restoration success. Researchers walked throughout each site to document common stressors on salt marshes in New England including intense herbivory (snails, geese, etc.), human and dog trampling, lateral shoreline and stormwater erosion, ice rafting, heavy wrack deposition, and acute vegetation loss. To quantify vegetation loss that was observed near the riprap sills, halophyte cover was calculated between plots within 15 m of the sill and those landward in the low marsh of living shorelines. Adaptive management actions were taken at the sites during the regulatory required monitoring timeframe and are presented including replanting sites with salt marsh graminoids, repairing locations of erosion, and removing wrack off the marsh.
Data Analysis
One-way ANOVAs with post hoc Dunnett’s tests (control = LS treatment) were conducted for the first and last year of monitoring of selected metrics to gauge the individual recovery for each LS site. Redox potential (low and high marsh), halophyte cover (low and high marsh), and F. heteroclitus trap catch rate (site dependent) were chosen to represent indicators of habitat, wetland soil development, and fauna use at the LS sites. For marsh zones without a no action control treatment, paired t-tests or Welch’s t-tests were used to compare between the reference marsh and LS shorelines. Normality and homoscedasticity were verified for each selected metric in each marsh zone with sites aggregated together. Kruskal–Wallis and post-hoc Dunn’s tests were used if assumptions were not met for a selected metric in a marsh zone. A second-degree polynomial regression was conducted on the impact of algae cover on snail and crab density (Site = blocking factor) for the riprap sills of CC and WHF to better understand habitat development of the artificial structures. Combined snail and crab density was square root transformed and outliers were removed to improve normality of the data. Data analyses were conducted with dplyr (Wickham et al. 2019) and stats packages and visualized with ggplot2 (Wickham 2016) and patchwork (Pedersen 2022) packages in R 4.2.1 (R Core Team 2013).
Results
Redox potential in both the low and high marsh generally declined over time at all LS sites yet remained aerobic or mildly anaerobic within the first four years post-construction with values typically greater than +150 mV (Fig. 2, see Table S1 for mean and standard error of metrics). The redox potential was only comparable to the reference at NMP in the high marsh in years 3 and 4 (Fig. 2a–b, Table 2). Highly reduced sediment conditions (mean Eh < 0 mV) in LS marshes were only recorded in three instances: NMP in year 3 (− 42 mV — in low marsh) and WHF in year 3 (− 193 mV — in low marsh, − 119 mV — in high marsh). In year 3, borderline anaerobic conditions at NMP low marsh translated into mean sulfide concentrations of 0.4 mM and WHF had mean sulfide concentrations of 0.9 mM and 0.7 mM in the low and high marsh, respectively. The low marsh of CC remained well-drained and highly aerobic three and four years after installation with the site exhibiting only minor declines in redox potential. Water content of the soil cores within the top 10 cm in the low and high marsh zones across all LSs ranged from 10 to 30% in 2020 (Table 3). The percent organic matter varied between living shoreline sites with WHF greater than 5% in both the low and high marsh in year 1 whereas organic matter was roughly 1% in CC and NMP low marsh in years 2 and 4, respectively, indicating the turnover of belowground biomass produced by plants each year was effectively oxidized.

Comparisons between living shorelines and reference salt marshes for a–b halophyte cover, c–d reduction–oxidation potential, e F. heteroclitus trap catch rate, and f F. heteroclitus adult length over restoration timeline. Asterisks denote where living shoreline is statistically equivalent or greater than the reference marsh for halophyte cover, reduction–oxidation potential, and trap catch rate. Results reported as mean ± standard error. Points have been horizontally shifted by site within each year to avoid overlap
Across the three LSs, halophyte cover increased to 42% and 52% in the low and high marshes, respectively, by the third and fourth years. Marsh cover in the LSs were 23% and 26% less than the reference in the low and high marsh, respectively, by the third or fourth year (Fig. 2c–d, Table S1). Halophyte cover in the high marshes at WHF (year 3) and NMP (year 4) were similar to the reference (p > 0.05, Table 2). No sites had similar cover to the references in the low marsh within the first four years of monitoring. To investigate personal observations of reduced vegetation establishment behind the sills, we divided the low marsh within 15 m of the seaward edge and compared the halophyte cover in the third and fourth years. The seaward width of the LSs possessed 10.0 ± 5.3% less halophyte vegetation cover than the landward plots in the third and fourth years. All LS sites had similar or greater high marsh species richness compared to the references by the third and fourth year (Table S1). The no action control shorelines for CC and NMP had no salt marsh vegetation throughout monitoring and the eroding no action vegetation of WHF was variable yet declining. The increase in high marsh species richness at LSs resulted from natural colonization of halophytes from the local species pool. For example, NMP had the same species composition as the local reference in 2019 including forbs such as Plantago maritima, Limonium nashii, and perennial grasses like Puccinellia maritima (Table S2).
Minnow traps captured 2978 individuals comprising three species: F. heteroclitus (99.6%), Carcinus maenas (0.32%), and F. majalis (0.08%). Catch rates of individual traps were highly variable across sites, seasons, and treatment shorelines (Fig. 2e, Table S1). Trap catch rate of F. heteroclitus at each LS was similar or even greater than the paired references across all sites during monitoring (Table 2). The high marsh of WHF in year 3 was the only instance in which the LS supported a larger population of F. heteroclitus than the reference (p < 0.05). The no action control shorelines consistently supported a smaller F. heteroclitus populations based on mean catches of less than 2 individuals per trap. Adult lengths of F. heteroclitus at the LSs were relatively similar across sites and time (50–59 mm) in the LS and reference marshes (Fig. 3f).

Algae and invertebrate colonization on living shoreline sills throughout monitoring. Wagon Hill Farm and Cutts Cove are riprap sills and North Mill Pond is a decomposed coir fiber sill. a Algae cover (%) of sills, b Semibalanus barnacle density (# m−2) of sills, c Density of gastropods and crabs (# m−2) on the sills, and d Second degree polynomial regression of combined gastropod and crab density (# m−2) and algae cover of plots of Cutts Cove (Years 1–3) and Wagon Hill Farm (Years 0–2) with 95% confidence interval (F3, 65 = 58.01, p < 0.001, R2 = 0.65). Error bars are ± standard error. Sites are distinguished by color and shape: Cutts Cove — orange square, North Mill Pond — yellow diamond, Wagon Hill Farm — blue triangle. Points have been horizontally shifted by site within each year to reduce overlapping in panels a–c
Algae (Fucus and Ascophyllum sp.) and common barnacles (Semibalanus sp.) colonized the riprap sills within several growing seasons with 77.0 ± 3.2% algae cover and 57.0 ± 12.8 barnacle individuals m−2 at CC within four growing seasons (Fig. 4a–b). It should be noted that a large barnacle die-off event occurred between 2020 and 2021 at CC with a 72% decline in live barnacles. Colonization of algae and invertebrates lagged at WHF compared to CC within the same timeframe. For example, algae cover expanded to 45 ± 5.2%, barnacle density to 6.0 ± 2.3 indiv. m−2, and combined crab and snail density to 18.4 ± 4.5 indiv. m−2. The non-filter feeder invertebrate community was composed of Littorina littorea (Common periwinkle), L. saxatilis (Rough periwinkle), and Carcinus maenas (Green crab). No mussels or oysters were found on the sills throughout monitoring. Non-filter feeders heavily colonized the riprap sill of CC by the fourth year (72.6 ± 5.6 indiv. m−2) while WHF had similar densities with CC by the second year (18.4 ± 4.5 indiv. m−2, Fig. 4c). Snail and crab density followed a second-degree polynomial regression with algae cover (F2, 65 = 56.01, p < 0.001, R2 = 0.65, Fig. 4d), suggesting continued colonization by macroinvertebrates until 70–80% algae cover on the riprap sills. Algae, barnacles, and non-filter feeding invertebrates did not colonize the NMP coir fiber log sills, which had decomposed or been buried by 2019.

Examples of the four adaptive management considerations documented at each living shoreline project in the study. a Spartina alterniflora mortality at the seaward edge of North Mill Pond illustrated with the original planted vegetation extent highlighted by dashed lines, b Heavy wrack deposition in the high marsh, in the far corner of example (a), prevents establishment of graminoids, c Erosion of sediment behind the riprap sill of Wagon Hill Farm two years after repairing original erosion with wrack-filled jute ‘pillows’ and sediment, and d Installation of 4 ft. tall snow fencing to prevent geese herbivory in the low marsh after community planting of 8000 S. alterniflora plugs
Discussion
Recovery of Salt Marsh Ecosystem Structure and Functions
The relatively aerobic redox potential, low sulfide concentrations, and rapid drainage throughout monitoring showed limited hydric soil development. High bulk densities and very low organic matter composition of the soil cores, especially at CC, indicated little to no peat development compared to reference marsh values and reported values for natural fringe marshes (Craft et al. 1999; Morgan et al. 2009). Constructed fringe salt marshes typically require decadal time scales to develop peat through deposition and belowground biomass accumulation (Bricker-Urso et al. 1989; Noll et al. 2019; Payne et al. 2019). The lack of time and complete drainage of tidal water contributed to the low organic matter within the first 10 cm in the soil column, especially in CC and NMP. Perhaps the sandy soils used in construction of these pilot projects to support plant establishment also resulted in rapid drainage and decomposition of belowground biomass. Wang et al. (2019) identified increased decomposition and export of dissolved organic carbon through improved drainage of marsh cores.
Individual site histories and construction details may offer additional explanations for the variable development rates of hydric soil conditions. Higher organic matter content at WHF was most likely a result of a novel planting technique. Salt marsh graminoid plugs were cultivated off site and installed in a 5 cm turf, composed of a mixture of organic material and mineral soil, across the entire marsh surface. NMP was constructed on top of a degraded salt marsh for mitigation, so the large swath of relic peat in the high marsh may have contributed to accelerated development of mildly anaerobic conditions and the observed sulfide concentrations. The lack of anaerobic conditions at CC may have resulted from a combination of construction with only sandy soils, steeper marsh slopes, and lower vegetation cover (see Fig. 2a-b). Active foraging of rhizomes from geese (see “Adaptive Management Needs of Living Shorelines in New Hampshire” section) prevented adequate belowground biomass inputs to offset decomposition rates.
Halophyte vegetation, measured in visual cover and species richness, were nearing comparable levels of nearby references within four years even in the shorter growing seasons of northern New England. In the mid-Atlantic coast, vegetation has been shown to reach equivalency in stem density within a few growing seasons (Bilkovic and Mitchell 2013; Gittman et al. 2016) and lag in visual cover by only 11–23% (Currin et al. 2008). Vegetation of the LSs in this study may require longer times to fully develop in New Hampshire as halophyte cover lagged on average 23% and 26% after three or four years compared to local references for low and high marsh zones, respectively. Plant species richness and composition did reach similar or greater levels at all three living shorelines compared to references, a much quicker recovery than observed previously in constructed salt marshes of Great Bay Estuary (Morgan and Short 2002).
Resident salt marsh nekton occupied the LS and reference fringe marshes similarly based on comparable levels of abundance and adult size distribution throughout monitoring. Nekton have been observed to respond to the creation of new salt marsh habitat within several months with increased densities of residents and presence of transient species (Currin et al. 2008). Increased cover of edaphic algae from greater light penetration (less dense S. alterniflora cover) may lead to better grazing habitat for F. heteroclitus (Seliskar et al. 2002); however, nekton catch rates were highly variable over time in the pilot LS projects. The nekton community may require 3–5 years to stabilize to reference levels as the vegetation continues to recover as found for mid-Atlantic LSs (Gittman et al. 2016). The restoration of salt marsh habitat is a substantial enhancement of nekton preferred habitat over the pre-existing mudflats and riprap shorelines, where substantially smaller populations were observed here and by others (Davis et al. 2006; Balouskus and Targett 2016).
Colonization on the Riprap Sills
The riprap sills of the LSs supported greater abundance of macroalgae and invertebrates compared to coir fiber sill as they took advantage of the three-dimensional hard surface and increased habitat heterogeneity. The decomposition of the coir fiber sills within five years of restoration at NMP represents a lost opportunity to reduce wave energy and provide habitat heterogeneity for support of macroalgae, gastropods, and bivalves. The rate of colonization by macroalgae and invertebrates differed between CC and WHF, likely driven by different locations and environmental conditions within the Great Bay Estuary (Jones 2000). For example, Semibalanus are intolerant of lower salinity and therefore may be less abundant at the fresher WHF site (Sundell et al. 2019). The invertebrate community of the sills was dominated at both CC and WHF by Semibalanus barnacles with a smaller yet growing composition of C. maenas, L. littorea, and L. saxatilis. Bilkovic and Mitchell (2013) observed similar trends of the invertebrate community with dominance by epibenthic filter feeders and minor presence of gastropods. It remains to be seen if epibenthic species other than barnacles, like Mytilus edulis (Blue mussel) or G. demissa, will colonize the sills. Inclusion of specific substrates and materials like shell bags may be required to achieve substantial and sustained populations of G. demissa and M. edulis in salt marsh LS projects (see Moody et al. 2022). Natural analog comparisons to oyster reef restorations (three dimensional hard structures in a soft sediment benthos) suggest that the riprap sills may provide similar refugia (Grabowski 2004), providing species-specific habitat characteristics (Hardaway et al. 2007; Zeug et al. 2007). For example, shoreline complexity in the form of a sill, macroalgae colonization, and salt marsh vegetation are preferred by certain species like Menidia menidia (Atlantic silverside) and may lead to higher species richness than reference marshes over time (Balouskus and Targett 2016).
Adaptive Management Needs of Living Shorelines in New Hampshire
LS restoration projects experienced multiple stressors including herbivory, wrack deposition, and erosion of newly placed sediments, which impacted the establishment of marsh vegetation and development of wetland soil characteristics. At CC, aggressive herbivory in the low marsh by Branta canadensis (Canada goose) was documented first in November and December of 2019 and 2020 and then transitioned to a year-round problem in 2021 when local populations became resident rather than migratory (Burdick and Moore 2017; Burdick et al. 2021a). Geese were able to take advantage of the coarse sandy fill and easily exhume S. alterniflora rhizomes for consumption prior to their development of root networks (see Esselink et al. 1997), completely denuding large sections of the low marsh. Herbivory left large holes (0.5 m diameter) of non-stabilized sediments throughout the marsh and dramatically reduced vegetation cover achieved over the first two growing seasons (Smith and Odum 1981; Fig. 4d). Consecutive annual grazing by geese of natural marshes can shift the marsh from a vegetated state to a hypersaline mudflat prone to accelerating erosion (Robert and Robert 2002). To address this concern, we installed fencing with plastic flagging tape around the perimeter of the low marsh to prevent further damage (see Mason 1995; Larson et al. 2018). These initiatives required monthly maintenance throughout the winter from damaging Nor’easter storms. Periodic site visits and wildlife camera deployments confirmed that our placement of perimeter fencing and flagging tape across the marsh was sufficient to stave off further foraging events. The site was replanted in 2023 with 8000 plugs of S. alterniflora and 2000 plugs of D. spicata to offset losses from herbivory.
Vegetation establishment varied across sites and elevation gradient of the LSs. S. alterniflora plugs experienced higher mortality rates and slower growth closer to the seaward edge demonstrated by the lower visual cover (~ 10% less) immediately behind the sills. In addition to herbivory by geese, the plugs at the seaward edge experience greater flooding as well as wave action and erosion of fine sediments compared to more landward plants. The loss of S. alterniflora was especially noticeable at NMP where no S. alterniflora survived within 3–5 m of the seaward edge (Fig. 4a, Burdick and Moore 2017). These seaward areas could be better protected from physical exposure and should be planted at higher densities than landward portions of the low marsh to improve success (Silliman et al. 2015).
One pattern that was observed at both CC and WHF was an initial high establishment of planted graminoids in the high marsh, widespread loss of graminoids in years 1–2, and slowly rebound and spread of cover by S. patens, D. spicata, J. gerardii in years 3–4 (see Fig. 2d). Drought conditions, sandy soils used in LS construction, and plantings at elevations above mean higher high water possibly led to moisture stress in newly planted graminoids at WHF and CC, especially over the winter months (Burdick et al. 2021a, 2021b). The use of finer grain size and increased organic matter in the fill for high marsh could improve soil moisture retention, biogeochemical soil development, and plant survival. Both NMP and WHF experienced heavy wrack deposition which smothered and prevented vegetation establishment in some portions of the high marsh (Fig. 4b). The location of wrack deposition was variable each year, killing salt marsh grasses and hindering development of the high marsh vegetation community (Brewer et al. 1998). In years when wrack deposition was particularly heavy at WHF, it was raked off the marsh in late spring to reduce the scale of impact.
Isolated erosion was observed and addressed at the WHF and CC within the first several growing seasons. At WHF, sandy fill material immediately behind the riprap sill began eroding despite careful anchoring of a jute mat (Fig. 4c), which subsequently required considerable effort to repair to ensure integrity of the sill edge (Burdick et al. 2021b). First, eroding gaps between marsh sediment and sill were stuffed in 2021 with wrack-filled jute tubes and refilled with the eroded sediments that had accumulated below the sill. However, once the jute tubes decomposed within 1–2 years, erosion driven by winter Nor’easter storms continued as before. Gaps behind and within the sill are planned to be filled with 4–6″ stone and recovered with the sediment that accumulated below the sill as the next approach to abate the erosion.
Runoff from the upland created small gullies at all LSs, demonstrating that heavy rainfall in small catchments can erode and degrade new restoration projects (Burdick et al. 2020). At CC the observed erosion was relatively minor before it was addressed by staking tubes of wrack-filled jute (0.2 m diameter by 1.3 m long) across problem gulley areas. In addition at CC, a small swale was constructed post-restoration just above the high marsh, parallel to the shoreline. This swale collected uphill surface runoff and moved it laterally away from the restored marsh surface, greatly reducing erosion from upland runoff. A coir tube was initially placed between low and high marsh areas at WHF; the coir was observed to capture upland eroded sediments after heavy rains and interrupt upland gully formation from continuing to the low marsh. If unchecked, erosion behind the sills or creation of gullies from stormwater runoff may wash out large amounts of sediment and lead to marsh loss or restoration failure.
Considerations for Restoration Success of Living Shorelines
The development of the pilot LS projects in New Hampshire can be defined by considerable gains of salt marsh habitat, functions, and ecosystem services in a relatively short timeframe. The results of the monitoring are even more encouraging when considering only riprap armoring was present at CC and very little salt marsh remained at WHF before restoration efforts, resulting in a net increase of salt marsh habitat for the system. The short-term restoration progress in this study reflected similar recovery timelines of previously documented LSs and restored marshes: biotic factors recovered relatively quickly (0–10 years) whereas biogeochemical processes will require decades (Craft et al. 2003; Noll et al. 2019; Isdell et al. 2021). The relatively quick recovery of vegetation and nekton could be a short-term incremental step within a longer, more gradual recovery as the pore water chemistry of wetland soils and remaining gaps in vegetation cover slowly develop over decades. However, persistent below-reference conditions over long timeframes of biotic factors and biogeochemical processes of created and restored wetlands, resembling asymptotic trajectories, have been well-documented in specific case studies (Zedler and Callaway 2000; Morgan and Short 2002) and meta-analyses (Moreno-Mateos et al. 2012; Jones et al. 2018). The restoration trend of certain factors may be less dependent on time than on other biotic and abiotic factors (Raposa et al. 2018; Lilian et al. 2021) such as the development of the invertebrate community on the riprap sills or herbivory by geese. Long-term monitoring should be continued during the first ten years to support adaptive management so that the LSs do not enter an alternative state (Folke et al. 2004) and provide less ecosystem functions than references (Borja et al. 2010; Moreno-Mateos et al. 2017).
Conclusion
Pilot projects remain an essential tool in coastal restoration for state and local stakeholders to better refine expectations for initial costs, recovery rates, and adaptive management needs. The pilot LSs in New Hampshire quickly recovered valuable ecosystem structure and functions within an initial phase (0–4 years), primarily driven by vegetation and nekton. Installation of three similar projects allowed us to partially attribute the varying recovery of the vegetation and soil biogeochemistry to unique site histories and unique external stressors. Significant impediments to restoration progress were identified such as waterfowl herbivory, high plant mortality, winter ice damage, and excessive wrack deposition. The small-scale nature of pilot projects provided the opportunity to innovate and attempt new adaptive management techniques without substantial costs (relative to initial construction). The first LS projects in New Hampshire demonstrate the potential for the tool to restore salt marsh habitat, improve resilience to climate change, and abate shoreline erosion in northern New England as have been widely observed in the Southeast and Mid-Atlantic.
Data Availability
Data and R code for the project are public and available at: https://doi.org/10.6084/m9.figshare.23957979. Data includes metadata on site locations and descriptions and all monitoring data from 2019 - 2022.
References
Balasubramanayam, V., and K. Howard. 2019. New Hampshire living shoreline site suitability assessment. Portsmouth, New Hampshire: New Hampshire Department of Environmental Services. https://www.des.nh.gov/sites/g/files/ehbemt341/files/documents/2020-01/r-wd-19-19.pdf. Accessed 15 Mar 2023.
Balouskus, R.G., and T.E. Targett. 2016. Fish and blue crab density along a riprap-sill-hardened shoreline: Comparisons with Spartina marsh and riprap. Transactions of the American Fisheries Society 145: 766–773.
Bilkovic, D.M., and M.M. Mitchell. 2013. Ecological tradeoffs of stabilized salt marshes as a shoreline protection strategy: Effects of artificial structures on macrobenthic assemblages. Ecological Engineering 61: 469–481.
Bilkovic, D. M., M.M. Mitchell, J.D. Toft, and M.K. La Peyre. 2017. A primer to living shorelines. In Living Shorelines, ed. D. M. Bilkovic, M. M. Mitchell, J. D. Toft, and M. K, 3–9. La Peyre, editors. Living Shorelines. Boca Raton, Florida: CRC Press.
Bilkovic, D.M., R.E. Isdell, A.G. Guthrie, M.M. Mitchell, and R.M. Chambers. 2021. Ribbed mussel Geukensia demissa population response to living shoreline design and ecosystem development. Ecosphere. https://doi.org/10.1002/ecs2.3402.
Borja, Á., D.M. Dauer, M. Elliott, and C.A. Simenstad. 2010. Medium- and long-term recovery of estuarine and coastal ecosystems: Patterns, rates and restoration effectiveness. Estuaries and Coasts 33: 1249–1260.
Brewer, J., J. Levine, and M. Bertness. 1998. Interactive effects of elevation and burial with wrack on plant community structure in some Rhode Island salt marshes. Journal of Ecology 86: 125–136.
Bricker-Urso, S., S.W. Nixon, J.K. Cochran, D.J. Hirschberg, and C. Hunt. 1989. Accretion rates and sediment accumulation in Rhode Island salt marshes. Estuaries and Coasts 12:300–317.
Bridges, T. S., P.W. Wagner, K. Burks-Copes, M.E. Bates, Z. Collier, C.J. Fischenich, J.Z. Gailani, D.L. Leuck, C.D. Piercy, J.D. Rosati, E.J. Russo, D.J. Shafer, B.C. Suedel, E.A. Vuxton, and T.V. Wamsley. 2014. Use of natural and nature-based features (NNBF) for coastal resilience. Vicksburg, Mississippi: Engineer Research and Development Center, United States Army Corps of Engineers. https://usace.contentdm.oclc.org/digital/collection/p266001coll1/id/3442/. Accessed 15 Mar 2023.
Burdick, D.M. and G.E. Moore. 2017. Salt marsh mitigation at Brewster Street, Portsmouth, New Hampshire. Year 2 monitoring report 2017. Auburn, New Hampshire: Stantec Consulting Services, Inc.
Burdick, D.M., G.E. Moore, T.P. Ballestero, R. Grizzle, K. Ward, J.G. McKown, and A.R. Payne. 2020. A multi-habitat restoration in Cutts Cove, Portsmouth, NH: year three final report. Concord, New Hampshire: New Hampshire Department of Environmental Services.
Burdick, D.M., G.E. Moore, T.P. Ballestero, R. Grizzle, K. Ward, J.G. McKown, and A.R. Payne. 2021a. A multi-habitat restoration in Cutts Cove, Portsmouth, NH: year four final technical report. Concord, New Hampshire: New Hampshire Department of Environmental Services.
Burdick, D.M., T.P. Ballestero, J. Ballestero, J.G. McKown, and G.E. Moore. 2021b. A living shoreline for wagon hill farm: report of activities and monitoring 2021. Concord, New Hampshire: New Hampshire Department of Environmental Services.
Carlisle, B.K., A.M. Donovan, A.L. Hicks, V.S. Kooken, J.P. Smith, and A.R. Wilbur. 2002. A volunteer’s handbook for monitoring New England salt marshes. Boston, Massachusetts: State of Massachusetts Office of Coastal Zone Management. https://www.mass.gov/files/documents/2016/08/oe/vols-guide-salt-marsh-2002.pdf. Accessed 15 Mar 2023.
Chambers, R.M., A.L. Gorsky, R.E. Isdell, M.M. Mitchell, and D.M. Bilkovic. 2021. Comparison of nutrient accrual in constructed living shoreline and natural fringing marshes. Ocean and Coastal Management 199: 964–5691.
Clewell, A.F. and J. Aronson. 2004. Ecological restoration: principles, values, and structure of an emerging profession. Washington D.C.: Island Press.
Cline, J.D. 1969. Spectrophotometric determination of hydrogen sulfide in natural waters. Limnology and Oceanography 14: 454–458.
Craft, C.B., E.D. Seneca, and S.W. Broome. 1991. Loss on ignition and kjeldahl digestion for estimating organic carbon and total nitrogen in estuarine marsh soils: Calibration with dry combustion. Estuaries and Coasts 14: 175–179.
Craft, C., J. Reader, J.N. Sacco, and S.W. Broome. 1999. Twenty-five years of ecosystem development of constructed Spartina alterniflora (Loisel) marshes. Ecological Applications 9: 1405–1419.
Craft, C., P. Megonigal, S. Broome, J. Stevenson, R. Freese, J. Cornell, L. Zheng, and J. Sacco. 2003. The pace of ecosystem development of constructed Spartina alterniflora marshes. Ecological Applications 13: 1417–1432.
Crum K.P., R.G. Balouskus, and T.E. 2018. Targett. Growth and movements of mummichogs (Fundulus heteroclitus) along armored and vegetated estuarine shorelines. Estuaries and Coasts 41: 131–143.
Currin, C.A., P.C. Delano, and L.M. Valdes-Weaver. 2008. Utilization of a citizen monitoring protocol to assess the structure and function of natural and stabilized fringing salt marshes in North Carolina. Wetlands Ecology and Management 16: 97–118.
Davis, J.L., C.A. Currin, C. O’Brien, C. Raffenburg, and A. Davis. 2015. Living shorelines: Coastal resilience with a blue carbon benefit. PLoS ONE. https://doi.org/10.1371/journal.pone.014259.
Davis, J.L.D., R.L. Takacs, and R. Schnabel. 2006. Evaluating ecological impacts of living shorelines and shoreline habitat elements. In Management, policy, science, and engineering of nonstructural erosion control in Chesapeake Bay: Proceedings of the 2006 Living Shoreline Summit, ed S. Y. Erdle, J. L. D Davis, and K.G. Sellner, 71–77. Boca Raton, FL: CRC Press.
Diefenderfer, H.L., G.D. Steyer, M.C. Harwell, A.J. LoSchiavo, H.A. Neckles, D.M. Burdick, G.E. Johnson, K.E. Buenau, E. Trujillo, J.C. Callaway, R.M. Thom, N.K. Ganju, and R.R. Twilley. 2021. Applying cumulative effects to strategically advance large-scale ecosystem restoration. Frontiers in Ecology and the Environment 19: 108–117.
Esselink, P., G.J.F. Helder, B.A. Aerts, and K. Gerdes. 1997. The impact of grubbing by Greylag Geese (Anser anser) on the vegetation dynamics of a tidal marsh. Aquatic Botany 55: 261–279.
Folke, C., S. Carpenter, B. Walker, M. Scheffer, T. Elmqvist, L. Gunderson, and C.S. Holling. 2004. Regime shifts, resilience, and biodiversity in ecosystem management. Annual Review of Ecology, Evolution, and Systematics 35: 557–581.
Gittman, R.K., A.M. Popowich, J.F. Bruno, and C.H. Peterson. 2014. Marshes with and without sills protect estuarine shorelines from erosion better than bulkheads during a category 1 hurricane. Ocean and Coastal Management 102: 94–102.
Gittman, R.K., C.H. Peterson, C.A. Currin, F. Joel Fodrie, M.F. Piehler, and J.F. Bruno. 2016. Living shorelines can enhance the nursery role of threatened estuarine habitats. Ecological Applications 26: 249–263.
Grabowski, J.H. 2004. Habitat complexity disrupts predator-prey interactions but not the trophic cascade on oyster reefs. Ecology 85: 995–1004.
Hardaway, S., W.G. Reay, J. Shen, S. Lerberg, D. Milligan, C. Wilcox, and K. O’Brien. 2007. Performance of Sills: St. Mary’s City, St. Mary’s River, Maryland. St. Mary’s City, Maryland: Virginia Institute of Marine Sciences. https://scholarworks.wm.edu/cgi/viewcontent.cgi?article=1560&context=reports. Accessed 15 Mar 2023.
Hardwick-Witman, M.N. 1985. Biological consequences of ice rafting in a New England salt marsh community. Journal of Experimental Marine Biology and Ecology 87: 283–298.
Isdell, R.E., D.M. Bilkovic, A.G. Guthrie, M.M. Mitchell, R.M. Chambers, M. Leu, and C. Hershner. 2021. Living shorelines achieve functional equivalence to natural fringe marshes across multiple ecological metrics. PeerJ Life & Environment. https://doi.org/10.7717/peerj.11815.
Jones, H.P., P.C. Jones, E.B. Barbier, R.C. Blackburn, J.M. Rey Benayas, K.D. Holl, M. McCrackin, P. Meli, D. Montoya, and D.M. Mateos. 2018. Restoration and repair of Earth’s damaged ecosystems. Proceedings of the Royal Society B: Biological Sciences. https://doi.org/10.1098/rspb.2017.2577.
Jones, H.J. 2000. A technical characterization of estuarine and coastal New Hampshire. Durham, New Hampshire: PREP Publications. https://scholars.unh.edu/prep/294. Accessed 15 Mar 2023.
Kirwan, M.L., G.R. Guntenspergen, and J.T. Morris. 2009. Latitudinal trends in Spartina alterniflora productivity and the response of coastal marshes to global change. Global Change Biology 15: 1982–1989.
Kneib, R.T., and A.H. Craig. 2001. Efficacy of minnow traps for sampling mummichogs in tidal marshes. Estuaries and Coasts 24: 884–893.
Konisky, R.A., D.M. Burdick, M. Dionne, and H.A. Neckles. 2006. A regional assessment of salt marsh restoration and monitoring in the Gulf of Maine. Restoration Ecology 14: 516–525.
Larson, M., R. Swadek, J. Elizabeth, K. Alexander, and C. Haight. 2018. Salt marsh restoration design guidelines. New York City, New York: City of New York Parks and Recreation. https://naturalareasnyc.org/media/pages/in-print/partner-publications/9d5be9f959-1601408905/nycparks_saltmarshrestorationdesignguidelines.pdf. Accessed 15 Mar 2023.
Leone, A., and N. Tahvildari. 2023. Comparison of spectral wave dissipation by two living shoreline features in a sheltered tidal bay. Estuaries and Coasts 46: 323–335.
Lilian, M., C. Bastien, J. Juan José, J.M. Rey Benayas, B. Marie-Lise, M.R. Carolina, J.G. Alday, J. Renaud, D. Thierry, B. Elise, M. Michel, A. Didier, C. Emmanuel, and A.C. Francisco. 2021. Conceptual and methodological issues in estimating the success of ecological restoration. Ecological Indicators. https://doi.org/10.1016/j.ecolind.2021.107362.
Mason, J.R. 1995. Evaluation of white plastic flags as visual repellents for snow geese on coastal salt marshes. International Journal of Pest Management 41: 19–21.
McClenachan, G.M., M.J. Donnelly, M.N. Shaffer, P.E. Sacks, and L.J. Walters. 2020. Does size matter? Quantifying the cumulative impact of small-scale living shoreline and oyster reef restoration projects on shoreline erosion. Restoration Ecology 28: 1365–1371. https://doi.org/10.1111/rec.13235.
McKee, K.L., I.A. Mendelssohn, and M.W. Hester. 1988. Reexamination of pore water sulfide concentrations and redox potentials near the aerial roots of Rhizophora mangle and Avicennia germinans. American Journal of Botany 75: 1352–1359.
Meyer, D.L., and M.H. Posey. 2014. Isolated salt marsh colonization by a resident species, mummichog (Fundulus heteroclitus), and a transient species, pinfish (Lagodon rhomboides). Journal of Experimental Marine Biology and Ecology 460: 197–210.
Moody, J.A., S.A. Bouboulis, L. Haaf, E.R. Rothermel, and D.A. Kreeger. 2022. The spatiotemporal development of two shellfish populations and their associated filtration capacity on a living shoreline near Milford, Delaware, USA. Ecological Engineering 180: 106661. https://doi.org/10.1016/j.ecoleng.2022.10661.
Moreno-Mateos, D., M.E. Power, F.A. Comín, and R. Yockteng. 2012. Structural and functional loss in restored wetland ecosystems. PloS Biology. https://doi.org/10.1371/journal.pbio.1001247.
Moreno-Mateos, D., E.B. Barbier, P.C. Jones, H.P. Jones, J. Aronson, J.A. López-López, M.L. McCrackin, P. Meli, D. Montoya, and J.M. Rey Benayas. 2017. Anthropogenic ecosystem disturbance and the recovery debt. Nature Communications. https://doi.org/10.1038/ncomms14163.
Morgan, P.A., and F.T. Short. 2002. Using functional trajectories to track constructed salt marsh development in the Great Bay Estuary, Maine/New Hampshire, U.S.A. Restoration Ecology 10: 461–473.
Morgan, P.A., D.M. Burdick, and F.T. Short. 2009. The functions and values of fringing salt marshes in northern New England, USA. Estuaries and Coasts 32: 483–495.
National Research Council. 2007. Mitigating shore erosion along sheltered coasts. Washington D.C: National Academy of Sciences Press. https://www.nationalacademies.org/our-work/mitigating-shore-erosion-along-sheltered-coasts. Accessed 15 Mar 2023.
Neckles, H.A., M. Dionne, D.M. Burdick, C.T. Roman, R. Buchsbaum, and E. Hutchins. 2002. A monitoring protocol to assess tidal restoration of salt marshes on local and regional scales. Restoration Ecology 10: 556–563.
NOAA Restoration Center. 2020. NOAA restoration atlas database. Washington D.C. https://www.fisheries.noaa.gov/resource/map/restoration-atlas. Accessed 15 Mar 2023.
Noll, A., C. Mobilian, and C. Craft. 2019. Five decades of wetland soil development of a constructed tidal salt marsh, North Carolina, USA. Ecological Restoration 37: 163–170.
O’Donnell, J.E.D. 2017. Living shorelines: A review of literature relevant to New England coasts. Journal of Coastal Research 332: 435–451.
Palinkas, C.M., M.C. Bolton, and L.W. Staver. 2023. Long-term performance and impacts of living shorelines in mesohaline Chesapeake Bay. 2021. Ecological Engineering 290: 106944. https://doi.org/10.1016/j.ecoleng.2023.106944.
Payne, A.R., D.M. Burdick, and G.E. Moore. 2019. Potential effects of sea-level rise on salt marsh elevation dynamics in a New Hampshire estuary. Estuaries and Coasts. 42: 1405–1418. https://doi.org/10.1007/s12237-019-00589-z.
Pederson T. 2022. Patchwork: the composer of plots. https://github.com/thomasp85/patchwork. Accessed 15 Mar 2023.
Polk, M.A., and D.O. Eulie. 2018. Effectiveness of living shorelines as an erosion control method in North Carolina. Estuaries and Coasts 41: 2212–2222.
Polk, M.A., R.K. Gittman, C.S. Smith, and D.O. Eulie. 2021. Coastal resilience surges as living shorelines reduce lateral erosion of salt marshes. Integrated Environmental Assessment and Management 18: 82–98. https://doi.org/10.1002/ieam.4447.
Pollock, L.W. 1998. A practical guide to the marine animals of Northeastern North America. Brunswick, New Jersey: Rutgers University Press.
Portnoy, J.W., and A.E. Giblin. 1997a. Effects of historic tidal restrictions on salt marsh sediment chemistry. Biogeochemistry 36: 275–303.
Portnoy, J.W., and A.E. Giblin. 1997b. Biogeochemical effects of seawater restoration to diked salt marshes. Ecological Applications 7: 1054–1063.
R Core Team. 2013. R: a language and environment for statistical computing. R Foundation for Statistical Computing. Vienna, Austria. http://www.r-project.org/. Accessed 15 Mar 2023.
Raposa, K.B., and C.T. Roman. 2006. Seasonal habitat-use patterns of nekton in a tide-restricted and unrestricted New England salt marsh. Wetlands 21: 451–461.
Raposa, K.B., S. Lerberg, C. Cornu, J. Fear, N. Garfield, C. Peter, R.L.J. Weber, G. Moore, D. Burdick, and M. Dionne. 2018. Evaluating tidal wetland restoration performance using National Estuarine Research Reserve System reference sites and the Restoration Performance Index (RPI). Estuaries and Coasts 41: 36–51.
Robert, J.L., and R.F. Robert. 2002. Foraging geese, vegetation loss and soil degradation in an Arctic salt marsh. Applied Vegetation Science 5: 7–16.
Seliskar, D.M., J.L. Gallagher, D.M. Burdick, and L.A. Mutz. 2002. The regulation of ecosystem functions by ecotypic variation in the dominant plant: A Spartina alterniflora salt-marsh case study. Journal of Ecology 90: 1–11.
Shepard, C.C., C.M. Crain, and M.W. Beck. 2011. The protective role of coastal marshes: A systematic review and meta-analysis. PLoS ONE. https://doi.org/10.1371/journal.pone.0027374.
Silliman, B.R., E. Schrack, Q. He, R. Cope, A. Santoni, T. van der Heide, R. Jacobi, M. Jacobi, and J. van de Koppel. 2015. Facilitation shifts paradigms and can amplify coastal restoration efforts. Proceeding of the National Academy of Sciences 112: 14295–14300.
Smith, C.S., B. Puckett, R.K. Gittman, and C.H. Peterson. 2018. Living shorelines enhanced the resilience of saltmarshes to Hurricane Matthew (2016). Ecological Applications 28: 871–877.
Smith, C.S., M.E. Rud, R.K. Gittman, E.C. Melvin, V.S. Patterson, J.J. Renzi, E.H. Wellman, and B.R. Silliman. 2020. Coming to terms with living shorelines: A scoping review of novel restoration strategies for shoreline protection. Frontiers in Marine Science. https://doi.org/10.3389/fmars.2020.00434.
Smith, T.J. III., and W.E. Odum. 1981. The effects of grazing by snow geese on coastal salt marshes. Ecology 62: 98–106.
Sundell, K., A.L. Wrange, P.R. Jonsson, and A. Blomberg. 2019. Osmoregulation in barnacles: An evolutionary perspective of potential mechanisms and future research directions. Frontiers in Physiology 10: 877. https://doi.org/10.3389/fphys.2019.00877.
Tiner, R.W. 2009. Field guide to tidal wetland plants of the northeastern United States and neighboring Canada. Amherst, Massachusetts: University of Massachusetts Press.
Wang, F.K.D., M.E. Koreger, J.W. Gonneea, and Phlman, and J. Tang. 2019. Water salinity and inundation control soil carbon decomposition during salt marsh restoration: An incubation experiment. Ecology and Evolution 9: 1911–1921.
West, J.M., and J.B. Zedler. 2000. Marsh-creek connectivity: Fish use of a tidal salt marsh in southern California. Estuaries and Coasts 23: 699–710.
Wickham, H. 2016. ggplot2: Elegant graphics for data analysis. Springer – Verlag, New York City, New York. https://ggplot2.tidyverse.org/. Accessed 15 Mar 2023.
Wickham, H., M. Averick, J. Bryan, W. Chang, L.D. McGowan, R. Francois, G. Grolemund, A. Hayes, L. Henry, J. Hester, M. Kuhn, T.L. Pdersen, E. Miller, S.M. Bache, K. Muller, J. Ooms, D. Robinson, D.P. Seidel, V. Spinu, K. Takahashi, D. Vaughan, C. Wilke, K. Woo, and H. Yutani. 2019. Welcome to the tidyverse. Journal of Open Source Software 4: 1686. https://doi.org/10.21105/joss.01686.
Woods Hole Group. 2017. Living shorelines in New England: state of the Practice. Boston, Massachusetts: The Nature Conservancy. https://www.conservationgateway.org/ConservationPractices/Marine/crr/_Documents/Final_StateofthePractice_7.2017.pdf. Accessed 15 Mar 2023.
Zedler, J.B., and J.C. Callaway. 2000. Evaluating the progress of engineered tidal wetlands. Ecological Engineering 15: 211–225.
Zeug, S.C., V.R. Shervette, D.J. Hoeinghaus, and S.E. Davis. 2007. Nekton assemblage structure in natural and created marsh-edge habitats of the Guadalupe Estuary, Texas, USA. Estuarine, Coastal and Shelf Science 71: 457–466.
Acknowledgements
We thank Chris Peter for the advice and guidance on the Restoration Performance Index, Andy Payne for the help with field work, Nate Hermann, Jenny Gibson, and Easton White for advice on statistical analysis, Tom Brightman and Joel Ballestero for planning and assisting with living shoreline maintenance, Robert Isdell for the valuable insight in the manuscript review, and the research assistants that made the study possible: Chloe Brownlie, Nate Hermann, Lars Hammer, Maranda Keller, and Hanna Mogensen.
Funding
The Aquatic Resource Mitigation Program and Coastal Program of the New Hampshire Department of Environmental Services and the New Hampshire Department of Transportation provided funding for the design, construction, and monitoring of the living shoreline projects. Funding for this research was provided by the University of New Hampshire Graduate School, University of New Hampshire Marine Biology program, the William Spaulding Endowment in support of Marine Biology Research at the Jackson Estuarine Laboratory, and the Society for Ecological Restoration Laderman Student Grant. JEL contribution number 588.
Author information
Authors and Affiliations
Corresponding author
Additional information
Communicated by John C. Callaway
Supplementary Information
Below is the link to the electronic supplementary material.
Rights and permissions
Springer Nature or its licensor (e.g. a society or other partner) holds exclusive rights to this article under a publishing agreement with the author(s) or other rightsholder(s); author self-archiving of the accepted manuscript version of this article is solely governed by the terms of such publishing agreement and applicable law.
About this article

Cite this article
McKown, J.G., Moore, G.E., Burdick, D.M. et al. Short-Term Recovery of Pilot Living Shoreline Projects for Salt Marsh Habitat in New Hampshire. Estuaries and Coasts 47, 315–329 (2024). https://doi.org/10.1007/s12237-023-01284-w
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s12237-023-01284-w