
Abstract
Large wood influences river geomorphology and ecology, so similar function is presumed in estuarine tidal channels. Consequently, restoration ecologists and engineers often recommend large wood supplementation for tidal marsh restoration, but there is no guidance on how much large wood is appropriate or where it should be located. GIS analysis of high-resolution aerial photos was used to map the distribution of individual tree logs greater than 2-m length in reference tidal marshes of eight Puget Sound river deltas. Statistical analysis showed that distributary networks, channel size, marsh size, fetch, topography, and woody vegetation affect large wood distribution on the marsh surface and in tidal channels. Large wood densities were 28 to 50 times lower in Puget Sound tidal channels than in Western Washington streams. These results provide an initial foundation for further studies on the ecological and geomorphological significance of large wood in tidal marshes, and some initial guidance to engineers and planners for large wood placement in marsh restoration projects.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
There is an extensive literature on the geomorphological and ecological importance of large wood in fluvial ecosystems (reviewed in Lester and Boulton 2008; Jones et al. 2014; Roni et al. 2015). This literature has dealt with wood distributions in fluvial systems; recruitment rates from stream bank erosion and debris flows; mobility rates; effects on fluvial geomorphology; effects on stream ecology, particularly on aquatic insects and fish; and the effectiveness of large wood placement in stream habitat restoration for threatened and endangered fish. In contrast, the literature on large wood in tidal marshes amounts to only a handful of papers. One review remarked, “…there is little substantive documentation but considerable speculation on the role of large wood in estuaries. Most of this conjecture is based on extrapolating knowledge about the functional role of wood in rivers to assumptions about potential roles of wood in estuaries…” (Simenstad et al. 2003). Little has changed since this review. There is only one study on the geomorphological role of large wood in tidal wetlands, which showed that in a tidal forested (Sitka spruce) wetland, large wood could control pool spacing in tidal channels and force otherwise featureless plane-bed reaches into step-pool type channels (Diefenderfer and Montgomery 2009). However, the tidal forced step-pool channels differed from those in fluvial systems in that flood tide dominance caused pools to be located upstream of log jams in the tidal channels, rather than downstream as in fluvial channels. There is also only one study on the role of large wood in structuring tidal marsh vegetation, which showed that large wood serves as nurse logs for shrubs and trees, elevating seedlings above a critical tidal inundation threshold (Hood 2007a).
There are very few studies examining potential interactions between fish and estuarine large wood. In Oregon and Washington (USA) tidal marshes, there are only three studies on interactions between tidal channel large wood and juvenile salmonids (Oncorhynchus spp.), with conflicting results. Two, using underwater videography and snorkeling observations, found an association between large wood and juvenile salmonid aggregations (McMahon and Holtby 1992, Van de Wetering 2001), while the third, relying on seining, found no difference between tidal channel reaches with and without large wood (Wick 2002). The third study also found no effect of large wood on the local abundance of benthic or epibenthic invertebrates, or on sediment deposition rates, grain size, or organic carbon content. In two Australian estuaries, telemetry was used to evaluate the effect of large wood on black bream (Acanthopagrus butcheri) distributions (Hindell 2007). The results showed inconsistent effects across estuarine regions, diel periods, and seasons for each estuary. In a mesohaline region of Chesapeake Bay (Maryland, USA), sampling with a dropped box trap showed that four species of fish (Fundulus heteroclitus, F. majalis, Gobiosoma bosc, Gobiesox strumosus), two species of crabs (Callinectes sapidus, Rhithropanopeus harrisii), and grass shrimp (Palaemonetes pugio) were an order of magnitude more abundant in the presence of woody debris (i.e., small wood) than in its absence; furthermore, field and laboratory experiments showed that grass shrimp survivorship of fish predation was doubled in the presence of woody debris (Everett and Ruiz 1993).
Due to the cultural importance and threatened/endangered status of salmon, and their dependence on critical rearing habitat in tidal marshes and tidal channels (Macdonald et al. 1988; Magnusson and Hilborn 2003; David et al. 2016), there are extensive efforts to restore these habitats to support salmon population recovery throughout Washington, Oregon, and northern California. Habitat restoration planners and engineers often recommend addition of large wood to tidal channels in marsh restoration sites, applying fluvial paradigms to estuarine systems (Simenstad et al. 2003) despite the limited and conflicting evidence for large wood benefitting estuarine fish. Large wood placement on tidal marsh surfaces to affect vegetation or other ecological functions (e.g., as possible raptor perches) is less common. Unfortunately, there is no guidance for how much large wood should be added to restoration sites, or where it should be placed. As a result, some restoration sites appear to have unnatural and perhaps dysfunctional amounts of wood incorporated into their designs (Fig. S1).
In response to the evident need for design guidance, this paper aims to describe the distribution of large wood in reference tidal marshes of the major river deltas in Puget Sound to inform efforts to restore tidal marsh ecosystems to reference conditions and to provide an initial foundation for further studies on the ecological and geomorphological significance of tidal marsh wood. In particular, the principal questions of interest are:
-
How much large wood is typically found in Puget Sound river delta tidal marshes?
-
Is the distribution and abundance of large wood consistent with various potential mechanisms of wood delivery to the marshes, such as association with the banks of river distributaries (river branches that flow away from the mainstem river to the ocean) or blind tidal channels (tidal channels with only one connection to another water body, i.e., to a distributary or the ocean), and areas of high fetch—or is large wood randomly distributed on marsh surfaces and in blind tidal channels?
-
Are there any other tidal marsh characteristics that might affect large wood distributions (e.g., marsh surface elevation, tidal channel size)?
Characteristics of contributing drainage basins (basin size, logging history, land use intensity, location and size of dams, etc.) were not considered, except briefly, though this is an issue deserving further investigation. Historical management actions, such as snag removal and levee construction, were also not considered. While these actions have clearly impacted rivers and their deltas (Gonor et al. 1988; Collins et al. 2002), their impact has been greatest on mainstem and distributary river channels, and likely only indirect on blind tidal channels and marsh surfaces, which are the focus of this paper.
Methods
Study Sites
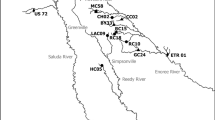
This study focused on tidal marshes in five of the largest river deltas in Puget Sound and two much smaller deltas in Hood Canal (western Washington; Fig. 1). The Skagit River drains an 8300-km2 watershed and has the largest Puget Sound delta and the largest extant marsh. The two principal distributaries of the Skagit River, the North Fork (NF) and South Fork (SF), form two active and distinct sub-deltas. Winter storms in Puget Sound generally come from the south, so the 380-ha NF delta tidal marsh experiences significant southerly storm fetch (11 km) across Skagit Bay, while the 1090-ha SF delta tidal marsh is relatively sheltered (6-km westerly fetch). Both deltas experience mixed semi-diurnal tides with a mean range of 3.2 m. More detail on Skagit Delta ecology and geomorphology can be found in Hood et al. (2016).

Tidal marsh study site locations (black polygons)
The Stillaguamish River drains a 1800-km2 basin and has 500 ha of tidal marsh at its mouth. Its mean tidal range is 3.3 m, and the modern delta has a 12-km southerly storm fetch.
The Nooksack River drains a 2200-km2 watershed. The 490-ha Nooksack delta marshes are located along the north margin of Bellingham Bay and thus have an extensive, 22-km, southerly storm fetch. This is the northern-most Puget Sound delta, experiencing the smallest mean tide range in Puget Sound, 2.6 m.
The Nisqually River drains a 2000-km2 watershed and has the southern-most delta with the largest mean tide range, 4.1 m, as well as the least storm fetch (3-km northerly fetch), because it is located on a southern shore. Recent habitat restoration has increased the Nisqually tidal marshes from 100 to 460 ha (Ellings et al. 2016), but the newly restored marsh is far from an equilibrium, so only the pre-restoration marshes were evaluated.
The Snohomish River drains a 4700-km2 basin and has the second largest delta in Puget Sound. Its extant marshes amount to 640 ha, of which 325 ha were historical or recent dike breach sites (Yang et al. 2010; Hood 2014), leaving 315 ha of reference marsh for consideration by this study. Most of the emergent zone Snohomish reference marshes are located in an area of the delta known as the Quilceda Marsh, and these were the focus for this study. The mean tide range for the lower Snohomish delta is 3.3 m.
All of the large river deltas are oligohaline and have generally similar vegetation communities, dominated primarily by sedges (Carex lyngbyei, Schoenoplectus pungens, S. tabernaemontani, and Bolboschoenus maritimus). The Nooksack has the greatest proportion of shrub and forested tidal wetlands, followed by the Skagit, Snohomish, and the Nisqually; the Stillaguamish has the lowest proportion.
The tidal marshes of the Union and Dosewallips deltas, draining into Hood Canal, are the smallest in this study at 118 ha and 38 ha, respectively, with mean tide ranges of 3.7 m and 3.5 m, respectively. Because of small drainage basin size (310 km2 for the Dosewallips; 62 km2 for the Union River), river discharge is comparatively low and the tidal marshes in the deltas are thus mesohaline to polyhaline. Consequently, the marsh vegetation is dominated by salt-tolerant species such as Distichlis spicata and Sarcocornia pacifica. The Union River is relatively low gradient, so as one moves upstream, the oligohaline tidal marshes transition into a relatively broad Sitka spruce and Western red cedar floodplain swamp. In contrast, the Dosewallips River has a steep gradient, because it drains the Olympic Mountains, so its vegetation transitions quickly from mesohaline intertidal marsh to forest. The Union Delta has a 2-km southerly storm fetch, the Dosewallips Delta a 7-km easterly fetch.
The Skagit, Stillaguamish, Snohomish, and Nisqually deltas host extensive agricultural land use, as well as several small, but growing towns. Consequently, river levees and sea dikes protect this land use from floods and storms. The Nooksack, Union, and Dosewallips deltas have less intensive land use in their deltas, but there is significant land use upstream. The Nooksack and Union deltas have abundant tidal forest habitat that gradually transitions to forested floodplain, so river levees are not present in these uninhabited areas. The Dosewallips delta is very steep, and is itself located at the end of a steep and narrow valley so river levees are not present and only low, remnant sea dikes are present in areas of abandoned farmland.
GIS Analysis
Individual pieces of wood were identified from true color aerial photos in an ArcMap 10.6 geographic information system (GIS), and each piece was manually digitized in a polygon shape file to create a census of all large wood (> 2-m length) in each river delta (Fig. S2). Aerial photos were selected that had relatively high resolution (15–30-cm pixels; Table S1), were flown at low tide, and were taken early in the growing season when vegetation cover was low. Photos were ground-truthed to the extent possible, not by recovering specific pieces of large wood in the field, because some photos were several years old during which time wood could move, but by confirming the general frequency of occurrence and detectability of wood on the marsh surface and within blind tidal channels. Field visits also confirmed that wood was not being missed in blind tidal channels by being covered by water. Even the largest channels rarely had residual water depths more than 0.5 m at low tide, and water was generally sufficiently clear to see even smaller pieces of wood.
Large wood was only identified in herbaceous tidal marsh; tidal shrub and tidal forest wetlands were ignored because canopy cover could obscure wood. Large wood was also only identified in blind tidal channels (channels that drain tidal marshes and generally only have one outlet), not river distributaries, because distributaries are generally much deeper than blind tidal channels, so submerged wood could be easily missed in distributaries, but not in blind tidal channels. Additionally, historical clearing of river snags (large wood extending from river banks or lodged within the channel) has occurred in river distributaries to facilitate river traffic (Gonor et al. 1988; Collins et al. 2002), but it is unlikely to have occurred in blind tidal channels, because they are unimportant to navigation. Thus, blind tidal channels are less likely than distributaries to have been impacted by anthropogenic legacies and may better represent reference conditions. Finally, most wood placement in tidal marsh restoration projects occurs in blind tidal channels, rarely in distributaries, so this is where restoration guidance is most needed.
Each digitized wood polygon was identified as being on the marsh surface or in a tidal channel. Large wood that was partially in a channel and partially on the marsh surface was classified as being channel wood, not marsh surface wood. Large wood that spanned the tops of tidal channels, but was not in the channel, i.e., not touching the channel bottom, was classified as being marsh surface wood. Only wood longer than 2 m was included in the census. Wood lengths were approximated as half of the polygon perimeter. Wood widths were not estimated because measurement error would be high, but minimum widths were approximately twice the resolution of the air photos used for GIS digitization, i.e., widths > 30–60 cm. Large accumulations of wood were often encountered in which individual pieces of wood were sometimes hard to distinguish, because logs could be on top of other logs. While a best estimate of the extent of each log was made, the total number and length of logs in these accumulations was likely underestimated. Photo resolution was insufficient to confidently distinguish logs with root wads from logs without, so this effort was abandoned.
To determine the elevations at which wood was located in the Skagit Delta, a digital elevation model (DEM) derived from 2012 lidar data (1-m spacing, 15-cm vertical accuracy, referenced to NAVD-88) was converted from raster form to polygon form using the ArcMap Conversion tool. This change in data formatting allowed the lidar polygon file to be intersected with the large wood polygon file using the Geoprocessing-Intersect tool, thereby associating the lidar elevations with each large wood polygon.
To determine the degree of large wood association with distributary reaches or blind tidal channels, the Select by Location tool was used to select large wood polygons within various distances (10 m, 20 m, etc.) of distributary or blind tidal channel polygons, and the number of selected polygons was then noted from the associated attribute table. Large wood count was graphed against each distance interval. Distributary reach size was indexed by the mean width of the first 100 m of the distributary downstream of a bifurcation. Mean width was calculated as the area of the polygon encompassing the 100-m length, divided by 100. This measure was chosen because distributary reach widths are irregular, but tend to flare in a downstream direction. Reach length was defined as the distance from one bifurcation till the next. Counts of large wood within a reach began 40 m downstream from a bifurcation to avoid the influence of the upstream, paternal reach.
Statistical Analysis
Simple linear regression was used to examine relationships between channel size and the count and total length of wood found in tidal channels and also between marsh island area and marsh surface wood count and total length. This was done to explore possible scaling of wood with channel or marsh island size. Marsh islands are bounded by distributaries and marine waters and are an important geomorphic unit in tidal marshes (e.g., Hood 2006, 2014). Because ecological patterns and processes are influenced by landforms and related physical processes, such as hydrodynamics and sediment transport, a parallel landscape-scale allometry in ecological patterns and processes has also been suggested (reviewed in Hood 2007b). Scaling relationships are described by power functions, so dependent and independent variables were log transformed to fit power functions and to equalize variance in the residuals. The slope of the fitted, log-transformed, regression lines is equal to the exponent of the power functions, i.e., the scaling exponent. Model I regression was used, because [1] measurement error for the independent variable (marsh island area or channel surface area) was low compared to the dependent variables (wood count or total length), i.e., marsh island boundaries were easy to distinguish while large wood was relatively more difficult to distinguish depending on vegetation cover, sun angle (shadow and glare), and water depth; and [2] prediction was desired to provide guidance for restoration planning and design (Sokal and Rohlf, 1995). Scaling relationships were compared between the river deltas using analysis of covariance (ANCOVA). When regression slopes were not significantly different between deltas, a common regression slope was calculated, followed by testing for differences in regression intercepts (Zar 1984). The criterion for statistical significance was p < 0.05.
The role of channel size on the distribution of large wood within channels was also analyzed more directly, by comparing wood frequency distributions vs. channel location measured by proportion of a channel length, i.e., standardized channel distances. These standardized distances were logit-transformed for regression against channel size as indexed by channel outlet widths, which were measured from air photos in a GIS. Logit transformation is necessary for regression of proportions bounded by 0 and 1. To more intuitively visualize differences in frequency distributions, channel widths were binned by apparent clusters in the data, and wood frequency distributions along untransformed standardized channel distances were compared between adjacent pairs of binned width clusters using the Kolmogorov–Smirnov two-sample test. When wood distributions were not significantly different, the channel width bins were grouped; when significantly different (p < 0.01), they were kept distinct. This resulted in three size categories of channels: small, medium, and large for each of three river delta systems evaluated: the NF Skagit, SF Skagit, and Snohomish. Other, smaller, river deltas had too few channels with too little wood for reliable analysis.
Results
The study deltas were chosen to represent diverse conditions of fetch, tide range, and spatial extent. However, the smaller deltas had few tidal channels (Table S2), so small sample size in the small deltas sometimes limited the ability to use inferential statistics. Thus, some results are necessarily focused on the larger deltas.
Marsh Surface Wood
The number of logs > 2-m length on the marsh surface scaled similarly with the size of the marsh island for all river delta marshes (Fig. 2). There was no significant difference in the slopes of the log-transformed regressions (F7,69 = 0.961), so that a common regression slope of 1.10 could be calculated, but there was a significant difference in the regression intercepts (F7,76 = 17.76, p < < 0.00001). Likewise, total length of logs > 2-m length showed similar patterns and had no significant difference in the slopes of the log-transformed regressions (F7,69 = 2.084), a common regression slope of 1.24, and significant differences in regression intercepts (F7,76 = 16.14, p < < 0.00001). The scaling relationships for all logs > 2-m length were paralleled by similar scaling relationships for logs > 5-m length as well as for logs > 10-m length (Fig. 3), indicating relevance of the scaling phenomenon regardless of the lower size limit of wood included in analysis.

Marsh surface log abundance and size relative to marsh area. Regression equations for log abundance (top frame) are: Nooksack, y = 61x1.13, R2 = 0.95; NF Skagit, y = 10.4x1.17, R2 = 0.76; SF Skagit, y = 0.96x1.29, R2 = 0.65; Stillaguamish, y = 17.0x0.84, R2 = 0.67; Snohomish (Quilceda marsh), y = 8.6x1.20, R2 = 0.97; Nisqually, y = 0.68x1.13, R2 = 0.69; Union, y = 7.5x1.20, R2 = 0.75; Dosewallips, y = 12.7x1.15, R2 = 0.68. Regression equations for total log length (bottom frame) are: Nooksack, y = 364x1.12, R2 = 0.98; NF Skagit, y = 87x1.14, R2 = 0.76; SF Skagit, y = 6.7x1.34, R2 = 0.62; Stillaguamish, y = 224x0.69, R2 = 0.67; Snohomish, y = 59x1.20, R2 = 0.96; Nisqually, y = 3.0x1.39, R2 = 0.70; Union, y = 48x1.24, R2 = 0.90; Dosewallips, y = 99x1.02, R.2 = 0.64

Scaling of marsh surface wood count and total length with marsh island area, for logs with lengths greater than 2 m, 5 m, and 10 m in four Puget Sound river deltas
Large, southerly, storm fetch was hypothesized to increase the retention of logs in a delta. The regression intercepts of the wood abundance scaling functions represent the relative density of wood in each delta when marsh island area is held constant; these intercepts were highly correlated with southerly storm fetch (Fig. 4).

Relationship to fetch of the y-intercepts of the large wood (LW) abundance vs. marsh area regressions. The Dosewallips Delta is the outlier to the regression. Omitting the outlier produces a regression equation of y = 0.75e0.22× with R.2 = 0.97
Channel Wood
For the NF Skagit, SF Skagit, Snohomish, and Nisqually deltas wood density within blind tidal channels scaled negatively with channel size (i.e., surface area), as did total wood length density (Fig. 5), while maximum wood length scaled positively with channel size. There were too few channels with large wood in the other deltas for reliable analysis. Scaling exponents appeared to be heterogeneous, ranging from −0.38 to −0.85 for wood density, from −0.34 to −0.64 for total wood length density, and from 0.12 to 0.26 for maximum wood length. However, ANCOVA could not detect significant differences in scaling exponents for any relationship (F3,97 = 1.008 for wood density, F3,97 = 0.355 for total length density, F3,97 = 0.122 for maximum length). Thus, a common scaling exponent was calculated for each relationship (−0.50 for density, −0.43 for total length density, 0.23 for maximum length). ANCOVA found significant differences in regression intercepts only for wood density (F3,100 = 3.670, p < 0.05 for wood density, F3,100 = 2.765, for total length density, F3,100 = 1.181 for maximum length). Post hoc tests found differences in the y-intercepts for wood density between the Snohomish and all other deltas and between the SF Skagit and the Nisqually. There was no detectable difference between the Nisqually and NF Skagit, but this was likely due to the small sample size for both deltas. Just as for marsh surface wood, the regression intercepts for blind channel wood density were correlated with southerly storm fetch (Fig. 6). While the small number of data points suggests this correlation should be treated with caution, the parallel with marsh surface wood suggests the effect of fetch is real.

Abundance and size of channel wood relative to channel area. Regression equations for wood density (top frame) are: NF Skagit, y = 5.4x−0.73, R2 = 0.42; SF Skagit, y = 8.3x−0.50, R2 = 0.42; Snohomish (Quilceda marsh), y = 23x−0.38, R2 = 0.28; Nisqually, y = 1.73x−0.85, R2 = 0.92. Regression equations for total length density (middle frame) are: NF Skagit, y = 37x−0.63, R2 = 0.29; SF Skagit, y = 67x−0.43, R2 = 0.31; Snohomish, y = 132x−0.34, R2 = 0.18; Nisqually, y = 15x−0.61, R2 = 0.87. Regression equations for maximum wood length (bottom frame) are: NF Skagit, y = 11x0.12, R2 = 0.05; SF Skagit, y = 15x0.24, R2 = 0.34; Snohomish, y = 15x0.23, R2 = 0.26; Nisqually, y = 9.3x0.26, R2 = 0.84; Union, y = 8.6x0.75, R2 = 0.69. The Dosewallips Delta had too few channels with logs for evaluation; the Union Delta had channels within a narrow range of sizes and are plotted only to evaluate their general consistency with other sites

Relationship to fetch of the y-intercepts of the large wood (LW) density vs. channel area regressions. The solid line is the fitted regression. Bubble area indicates relative sample size in each underlying regression. Greek letters are shared for those intercept values that were not significantly different according to post hoc tests following the ANCOVA
Negative scaling of wood density suggests the smallest channels have the highest wood density. However, this is a little misleading. The smallest channels actually had no large wood at all and thus could not be plotted on the log-scale graphs in Fig. 5. The smallest channels were too small for large wood to fit in the channels; instead large wood spanned their bank tops on the marsh surface. Thus, the negative scaling applies for channels above a size threshold size of about 0.008 ha, which typically have outlet widths of approximately 1.1 m.
Large wood is much more abundant on the marsh surface than within tidal channels, but the relative density of large wood in these two areas is variable in Puget Sound river delta marshes. Large wood density appears to be generally higher on marsh surfaces than in tidal channels for those deltas exposed to high winter storm fetch, and higher in channels than on marsh surfaces for those with low storm fetch (Fig. 7). To allow comparison with the fluvial literature, large wood density was also computed per unit length of channel. The NF Skagit, SF Skagit, and Snohomish (Quilceda) marshes had the most channels with large wood, 28, 56, and 45, respectively, which comprised 72%, 65%, and 82% of their blind tidal channels, respectively. Within those channels, the NF and SF Skagit channels had similar wood density, 0.0084 m−1 and 0.0080 m−1, respectively, while the Quilceda channel wood density was 0.0179 m−1. The other deltas had far fewer channels containing large wood (range = 2 to 8) and their average wood density was 0.0047 m−1 (range = 0.0022 to 0.0156).

Comparison of wood density over marsh island surfaces versus within blind tidal channels, without regard for spatial heterogeneity within these two categories
Distribution Heterogeneity-Marsh Surface Wood
Large wood was heterogeneously distributed across the marsh surface and within blind tidal channels in each river delta. Marsh surface distribution was controlled by topography (elevation gradients, discontinuities, and distributary planform), vegetation, and fetch. For example, in the SF Skagit Delta, the gradient in marsh surface elevation serves as topographic sieve that allows large wood to pass over low elevation marsh, but traps most wood at elevations between mean high water (MHW, the average level of all daily high tides over a 19-yr tidal epoch) and mean higher high water (MHHW, the average level of the higher of the two semi-diurnal tides). The frequency distribution curve of wood abundance versus marsh elevation (insert, Fig. 8) has an excess Kurtosis value of 2.45, indicating excess tail values compared to a normal distribution, and a skewness value of −1.13, indicating excess tail on the left of the distribution. Together these confirm visual inspection of wood distribution over the marsh surface (Fig. 8), i.e., that there is little wood at low marsh elevations, and much gets trapped at mid-range marsh elevations while gradually declining as one moves inland to higher marsh elevations.

Distribution of wood (black polygons) relative to marsh surface elevation in the South Fork Skagit Delta. The lower elevation limit of the vegetated marsh is located where yellowish elevations in the digital elevation model (DEM) border solid blue. Mean high water (MHW) is located where yellowish elevations change to orange. The inset graph shows wood frequency (y-axis) relative to marsh surface elevation at 10-cm bin intervals (x-axis). The three largest bins are located between MHW and mean higher high water (MHHW)
In the Snohomish Delta, wood accumulations were coincident with a low, 0.5-m scarp that distinguished old and young marsh (Fig. 9). Aerial photographs show that the lower-elevation, young marsh has developed through sedimentation and accretion between 1938 and 2003, as has been described previously for the nearby Skagit Delta (cf. Hood 2006). Similarly, wood accumulates against dikes, which act as tall artificial scarps (Fig. S3). Tidal shrub vegetation can also trap large accumulations of wood (Fig. 10). In this case, there is interaction between topography and vegetation, because the shrubs are often growing on natural distributary levees. The higher elevation and often coarser and better drained sediments of the natural levees, compared to adjacent marsh, facilitate shrub establishment. In these examples, topographic trapping is especially effective when storm fetch is directed toward the scarps, dikes, or shrub thickets.

Detail of the Quilceda marsh in the Snohomish Delta showing [A] accumulation of logs along topographic discontinuities (scarps) that distinguish older, higher marsh (dark green) from younger, lower marsh (light green). Marsh surface and channel logs are depicted by yellow and red polygons, respectively. Inset [B] shows detail of undigitized logs (gray) accumulated along a scarp. The lidar image of inset B shows the abrupt, approximately 50 cm, change in elevation associated with wood accumulation. Vegetation seaward of the scarp consists of intertidal sedges; landward vegetation consists of high intertidal shrubs and trees, which facilitate log trapping

An example of high intertidal shrubs and trees (black polygons) trapping marsh surface wood (yellow polygons). Channel wood is represented by red polygons. The shrubs and trees are growing on natural river and distributary levees that gently grade to the general marsh elevation; no scarps are present. Arrows indicate direction of ebb tide river flow. Bare areas of marsh near shrubs and trees occur because the wood has been intercepted by windward shrub thickets. Storm fetch is southerly
Large wood was generally most abundant within 20 m of a river distributary (Fig. 11), with abundance typically decreasing with distance from a distributary in a negative exponential fashion. Fetch appears to redistribute wood in the Union Delta, whose river has low discharge and gradient (so low stream power), and where there is significant southerly storm fetch. Here large wood is broadly distributed in the higher reaches of the delta, near adjacent uplands, with no tight association with river distributaries. Another slight exception is the North Fork Skagit Delta, where wood distribution is also strongly affected by fetch and tidal shrub vegetation, and the tidal shrub vegetation is itself associated with natural distributary levees. Here wood is most abundant within 80 m of distributary banks, before showing negative exponential decay with distance.

Marsh surface wood abundance per 20-m distance interval from the distributary network of a delta
Distributary reach width was positively related to wood density within 40 m of the distributary margins (Fig. 12), which is consistent with the idea that larger distributaries should carry more river flow and convey more wood from upstream sources. However, only three deltas could be analyzed statistically, because sample sizes were limited by either a paucity of distributaries in small deltas, a paucity of wood near the distributaries, or because very high cover of tidal forest and shrubs on natural distributary levees prevent direct river delivery of wood to distributary banks (Nooksack Delta).

Log density per meter of channel length within 40 m of a distributary versus distributary reach width
Similar to distributary channels, large wood was generally most abundant within 10 m of a blind tidal channel bank (Fig. 13), again with abundance usually decreasing in a negative exponential fashion with distance from the channel bank. However, in this case, there was no relationship between blind tidal channel size (outlet width) and wood density within 10 m of a blind tidal channel bank (R2 = 0.01).

Marsh surface wood abundance per 10-m distance interval from blind tidal channels in the South Fork Skagit delta. Channel size is indexed by outlet width (W)
Distribution Heterogeneity—Channel Wood
Only the NF Skagit, SF Skagit, and Snohomish deltas were examined for patterns in channel wood distributions, because the other, smaller, deltas had too few channels or too little channel wood for reliable analysis. Wood distribution within tidal channels varied with channel size. Small channels generally accumulated wood near their outlets, large channels near their heads (Fig. 14). These distribution patterns hold whether wood is > 2-m, > 5-m, or > 10-m length (data not shown). As tidal channels increased in size, there was a headwards shift in wood frequency distributions. The rate at which wood distributions moved headwards with increasing channel size was similar for the three deltas examined, with regression slopes ranging from 0.02 to 0.06, all of which were significantly different from 0 (p < 0.0001 for all), but despite their similarity they were nevertheless statistically different from each other (ANCOVA: F2,1050 = 17.08; p < 0.0001), with the 95% confidence limits for the slope estimates not overlapping for any of the deltas.

(Left frames) Bar graphs depicting wood distribution, for logs > 2-m length, along channels from outlet to head for small (gray bars), medium (white bars), and large (black bars) channels, where size is indicated by their outlet widths (see legend). Small channels generally accumulated wood near their outlets, large channels near their heads. (Right frames) Scatter plots depicting the same data as a continuous function of channel size, showing a general headwards shift in wood frequency distributions as channels increase in size. The Quilceda marsh was the only area in the Snohomish Delta examined, due to its relative lack of anthropogenic disturbance
These distribution patterns are likely controlled by channel width, which narrows with increasing distance from channel outlets. For all three river deltas examined, large wood was generally trapped in channel reaches < 4-m wide, and most frequently in channels 1–2-m wide (Fig. 15). In small tidal channels, these narrow channel widths are near the channel outlets. For increasingly larger channels, these critically narrow, wood-trapping, channel widths become increasingly more distant from the channel outlets. Additionally, the length of the wood trapped in a channel reach was correlated with channel reach width (Fig. 16; p < 0.001 for the NF Skagit delta and i < < 0.0001 for the SF Skagit and Snohomish deltas), though the amount of variance explained by these regressions was low, ranging from 8 to 12%. The low R2 values are likely due to the channels trapping many smaller pieces of wood in addition to the largest that match the channel capacity. These regression lines indicate similar scaling of wood length with channel width for all three deltas (ANCOVA: F2,904 = 0.537; p > 0.50, i.e., no significant differences in slopes); trapped wood length is a power function of channel width, with exponents (= slopes of the log-transformed data) of 0.20 for the SF delta and 0.25 for the NF and Snohomish deltas. Regression lines for data values along the upper edge of the data cloud would be representative of the longest wood trapped at various channel widths, i.e., the maximum capacity of a channel width. These regression lines are also similar for all three deltas, though no statistical tests were made because the data were visually selected post hoc to represent a boundary of the data cloud; they are presented here as an exploratory analysis. For these exploratory regressions, wood length is a power function of channel width with exponents of 0.55 for the SF, 0.56 for the NF, and 0.64 for the Snohomish deltas. For the SF, NF, and Snohomish deltas, respectively, the longest wood trapped in a channel was 43 m in a 3.5-m-wide channel, 31 m in a 2.6-m-wide channel, and 26 m in a 4.4-m-wide channel. In comparison, the longest wood found on the marsh surface of the SF, NF, and Snohomish deltas, was 42 m, 47 m, and 41 m, respectively. Thus, it is likely that significantly larger wood is simply not available for trapping in channels that are many 10 s of meters wide, so that the maximum capacity of very large channels is never reached by individual logs.

Frequency distributions of the channel widths where large wood was observed. Wood was generally trapped in channel reaches < 4-m wide, and most frequently in channels 1–2-m wide. The Quilceda marsh was the only area in the Snohomish Delta examined, due to its relative lack of anthropogenic disturbance

Correlation of wood length with width of the tidal channel reach where the wood was observed. Least squares regressions of log-transformed data are depicted by lower regression lines; all three regressions are significant, with p < 0.001 for the NF Skagit delta and p < < 0.0001 for the SF Skagit and Snohomish deltas. The upper envelopes of the data clouds are delimited by the upper regression lines. Similar regression slopes for all thee deltas are found for both groups of regression lines, suggesting similar processes are responsible for wood trapping in each delta
Discussion
Marsh Surface Wood
The processes that control large wood abundance and distribution in river delta tidal marshes can be assigned to three principle categories: wood delivery, redistribution, and trapping. Additionally, wood can be lost from a system through export (i.e., the inverse of trapping), through decay, and through burial. Decay and burial were not examined in this study. However, C14 dating of large-diameter (> 1 m) tidal marsh logs indicates decay may take several centuries (Tonnes 2008). Burial of marsh surface wood is likely also a relatively slow process, because marsh accretion when sediment supply is high is typically similar to rates of sea level rise (Kirwan et al. 2016), which in the vicinity of the Skagit Delta has been 2 mm per year over the past century (Hood et al. 2016). Sea level rise is likely also the principle control on the rate of wood burial in tidal channels (Allen 1997, 2000).
Large wood is delivered to delta marshes primarily by their rivers, as indicated by the high abundance of wood on distributary margins, and an exponential decline in wood abundance with distance from the distributaries (Fig. 11). Watershed land use should also logically affect wood abundance in the delta and some indication of this is shown by the Dosewallips having higher relative density of marsh surface wood, when marsh area and fetch are statistically controlled, than the other river deltas with higher intensity land use in their watersheds (Fig. 4; Studentized residual = 4.157, df = 5, p < 0.01). Inspection of aerial photos shows that the Dosewallips river system has the least amount of anthropogenic impact, suggesting more intact riparian zones and greater recruitment of large wood to the river and estuary. The upper 60% of the basin is in the Olympic National Park, the next 30% is in the Olympic National Forest, and the lowest 10% is comparatively lightly impacted by small farms and rural residences. In contrast farms and residential development impact 33% of the mainstem Nisqually River corridor (along with two dams), 50% of the mainstem Skagit River corridor (along with three dams), 50% of the mainstem Stillaguamish River corridor, 60% of the Union River corridor (along with one dam), 70% of the mainstem Snohomish River corridor, and 80% of the Nooksack River corridor (with two dams). These impacts are focused on the lower elevations of the rivers and primarily affect their floodplains. On the other hand, river basin size (surface area) was completely unrelated to the regression intercepts of the wood abundance scaling functions, with R2 = 0.02 for best fits of linear, power function, and exponential forms. Thus, greater recruitment of large wood from intact riparian zones may be responsible for the exceptionalism of the Dosewallips Delta.
Large wood could also be delivered to river deltas from nearby erosional coastlines. The importance of this alternate source likely varies with shoreline geometry, ocean currents, geology (e.g., rocky vs. sedimentary shorelines), and coastal land use. Additionally, large river deltas with correspondingly large rivers and river basins are more likely to have a correspondingly large proportion of their wood being of river origin.
Wood redistribution in river deltas seems likely to be most influenced by storm fetch and flood tides. The effect of fetch on wood redistribution was particularly evident in the Union Delta, where wood was not much associated with distributary channels, as it typically was with the other river deltas, but instead was abundant 60 to 340 m from the distributaries. Flood tide movement of wood was indicated by the association of large wood with the banks of blind tidal channels, which is likely the result of flood tides carrying wood into blind tidal channels and depositing it along the banks as the tide rises over the marsh surface.
The results suggest that as large wood is redistributed by tides and storm winds and waves, it becomes trapped in various parts of the delta. Storm fetch was highly correlated with the relative density of wood on the marsh surface, which indicates that at a coarse scale storm wind and waves are important in trapping large wood in deltas with large storm fetch, and preventing export to the ocean. When storm fetch is negligible, wood density is higher in blind tidal channels than the marsh surface, which suggests flood tides are the primary retention process, pushing wood into the channels and onto their banks. But when storm fetch is large, wood density can be higher on the marsh surface than in tidal channels, which suggests wood brought into tidal channels by the tides is energetically distributed out of the channels at high tide by storm winds and waves and onto the marsh surface.
At a finer scale, wood trapping is associated with topographic variation, such as small scarps, dikes, and elevation gradients that prevent wood from being pushed further landward by flood tides, wind, and waves. The Quilceda marsh in the Snohomish Delta is a clear example of wood accumulating against a small, 0.5-m scarp, that demarcated the boundary between old marsh and newly (between 1938 and 2003) prograded marsh. A similar example of wood accumulations against a progradational marsh scarp has been documented in the Nehalem Bay marshes on the Oregon coast (Johannessen 1964; Eilers 1975).
Dikes are tall artificial scarps, and those with significant storm fetch can trap so much wood that it smothers marshes and causes extensive damage to marsh vegetation (MacLennan 2005). This is an example of dikes having seaward impacts on marsh habitat, in addition to facilitating landward conversion of historical marshes to agricultural and urban use (e.g., Hood 2004).
Tidal shrub and forest vegetation also trap wood, in part because they grow at higher elevation, sometimes on natural levees or progradational scarps, but also because their woody stems form a barrier to landward wood movement. Because wood can serve as nurse logs for tidal shrub and forest vegetation (Hood 2007a), there is potential for seaward vegetation succession where a topographic discontinuity allows large wood to accumulate, which then provides nurse logs for woody vegetation establishment, which then traps additional wood seaward of the woody vegetation that also serves as nurse logs for the development of still more woody vegetation that traps more wood. This recursive process of seaward woody vegetation movement is probably kept in check by physical stresses such as storm waves and salinity. A similar process of waterward vegetation succession has been described for the Great Slave Lake (Canada) where large wood interacts with the lake shoreline, lake currents, and fetch to form large accretions of wood that provide nurse logs for trees, which in turn trap more wood, contributing to shoreline evolution (Kramer and Wohl 2015). However, in the Great Slave Lake, this successional process has been facilitated by falling water levels over the past 8000 years, while Puget Sound river deltas have experienced sea level rise during this time.
Where there are no abrupt topographic scarps, a gradual increase in marsh elevation from the sea to land can result in a winnowing of wood along the elevation gradient, with a few, perhaps larger logs, trapped at lower elevations, at least until a large storm pushes them further landward, and many more logs trapped between mean high water (MHW) and mean higher high water (MHHW). Wood that reaches elevations above MHHW is likely either dumped there during river floods or pushed there on higher high tides during energetic storms.
Finally, wood trapping efficiency was affected by marsh island size. Log count scaled with marsh area to a power of 1.10, and total log length to a power of 1.24. Scaling exponents > 1 indicate wood abundance and total length increased faster than did marsh island size, with total length increasing faster than abundance, i.e., large islands have disproportionately more and longer logs than small islands. Why was log density not uniform relative to marsh island area? One explanation is that large wood spends more time traveling across larger islands and therefore has more opportunity to get hung up on a topographic discontinuity, other logs, or shrub vegetation, and this is perhaps more likely as log length increases. On smaller islands, wood may float onto the island at high tide or in a storm, and then relatively quickly float off on the next high tide or storm. Another possibility is that because marsh-surface large wood is associated with channel banks, scaling of large wood with marsh island area could be influenced by scaling of tidal channel length with marsh island area (Hood 2015), given similar scaling exponents of 1.10 vs. 1.24, respectively.
All of the processes of wood delivery, redistribution, and trapping as a result of storms, fetch, tides, distributary geometry, marsh topography, and vegetation create distribution patterns for marsh surface wood ranging from simple to complicated depending on the relative strength and interaction of each process. Patterns are likely to vary between deltas as a result of differences in watershed size (which affects river discharge, delta gradient, distributary count, marsh vegetation type, etc., [Hood 2007b]) and watershed land use, and their interaction with coastal variation in fetch and tide range.
Blind Tidal Channel Wood
In fluvial channels, wood is recruited from river banks and carried downstream from narrower reaches to wider reaches (Wipfli et al. 2007; Kramer and Wohl 2017). This upstream to downstream movement of wood in fluvial channels is fundamentally different from blind tidal channels in herbaceous marsh, where tides or storms push wood from downstream to upstream, i.e., from wide distributary channels into increasingly narrower blind tidal channels, and this necessarily affects the distribution of wood in blind tidal channels. Except for the smallest tidal channels, which are too small to contain large wood, the larger the tidal channel the lower the log density in the channel, because wood is pushed into blind tidal channels by flood tides or storms until it is lodged within the narrowing channel and can go no farther. Wide channel reaches are mostly areas of wood transit, with generally only momentary (likely on the scale of days to years) accumulations of wood. Narrow channel reaches are likely long-term repositories of trapped wood, with the size of the wood trapped correlated with the width of the channel reach. In small (short) tidal channels whose outlet widths are 2–4 m, most wood is trapped near the outlets. In larger (longer) channels, most wood is trapped further and further from the outlet as channel size increases.
Scaling of wood length, density, and distribution with channel size likely has geomorphic and ecological consequences that are also associated with channel size. If invertebrate production is associated with wood in estuarine systems (e.g., Everett and Ruiz 1993) as it is in freshwater (Benke and Wallace 2003) and if fish abundance and productivity are generally associated with wood in estuarine systems (e.g., Everett and Ruiz 1993) as they are in freshwater (reviewed in Roni et al. 2015), then these ecological functions could also scale with tidal channel size in tandem with the scaling of wood (cf. Hood 2002, 2007b).
Implications for Tidal Marsh Restoration
Many restoration biologists and engineers uncritically apply the fluvial large wood paradigm to estuarine habitat restoration, but this is probably unwise given the significant differences between both systems, including bidirectional tidal vs. unidirectional fluvial flow, much lower topographic gradients in marshes, and the importance of storm fetch in estuarine systems, all of which affect estuarine wood distribution, and which may affect geomorphic interaction with large wood (e.g., Diefenderfer and Montgomery 2009).
Large wood supply or retention is probably also lower in estuarine than riverine systems, because the density of large wood, greater than 2-m length, in natural and unmanaged Western Washington lowland stream channels 0–6-m wide and 6–30-m wide was 0.29 m−1 and 0.52 m−1, respectively (Fox and Bolton 2007), which was 28 and 50 times more than in Puget Sound blind tidal channels. One possible reason for this difference is that the reference lowland streams were in nearly pristine habitat, while this study’s river deltas are likely impacted by extensive land use in their drainage basins that diminishes wood supply. Another possible reason for the difference is a legacy of log removal from navigable rivers and beaches (Gonor et al. 1988), which may have depleted wood supply to river deltas. On the other hand, a study of unusually large accumulations of wood that was smothering vegetation in two small coastal Puget Sound saltmarshes found that 46% of the logs were anthropogenic (e.g., had sawed ends, creosote, or embedded metal hardware used in towing log rafts), 10% were biogenic (e.g., had rootwads), and 43% were of indeterminate origin (MacLennan 2005), so at least in some areas natural wood supply may have been replaced and even surpassed by accidental anthropogenic supply (e.g., loss from towed log rafts). Thus, it is unclear what a historical pre-settlement baseline wood abundance was in blind tidal channels and on marsh surfaces, but in any case, the difference in large wood density between blind tidal channels and fluvial channels is so dramatic that it should caution engineers and restoration ecologists to carefully consider how much large wood they import to a restoration site. Finally, while the minimum log lengths were the same for my study and that of Fox and Bolton (2007), minimum log diameter was 10 cm in Fox and Bolton (2007), while the minimum detectable limit for log diameter in my air photo analysis was likely 30–60 cm, so that thinner large wood was omitted from consideration in Puget Sound tidal marshes.
These difference should inspire further investigation of the ecological and geomorphological consequences of blind tidal channel large wood and how it compares with fluvial wood function. The lower abundance of large wood in blind tidal channels compared to fluvial channels does not necessarily mean that large wood is less important in blind tidal channels, but it may further suggest that its ecological and geomorphological roles differ between tidal and fluvial systems. Additionally, the diameter of large wood likely also has significant influence on its geomorphological and ecological function (e.g., Gurnell et al. 2002; Montgomery et al. 2003), with particularly large contrast expected between 10- and 100-cm diameter wood. For example, the probability that the dominant shrub (Myrica gale) in the Skagit Delta tidal marshes was found growing on a nurse log increased steadily with log diameter, with none found on wood less than 20-cm diameter; similarly, the species of plants on tidal marsh nurse logs depended on nurse log diameter, with Sitka spruce (Picea sitchensis) growing on nurse logs averaging 120-cm diameter, while willows and other shrubs grew on logs averaging 60–80 cm diameter (Hood 2007a).
Tidal marsh restoration in the Pacific Northwest is generally done to provide critical rearing habitat for threatened Chinook salmon, so large wood is generally placed almost exclusively in tidal channel habitat to provide cover for fish or scour low-tide pools. However, the ecological role of large wood on marsh surfaces is likely underappreciated. In addition to providing nurse logs that support shrubs and trees (Hood 2007a), large marsh surface wood frequently provides perches for raptors such as bald eagles (Haliaeetus leucocephalus) and occasionally provides nesting platforms for Canada geese (Branta canadensis) (personal observations). More speculatively, rodent trails in high-elevation tidal marshes (personal observations) suggest nearby large wood may provide supra-tidal nest sites for rodents, similar to large wood on river banks and cobble bars (Steel et al. 1999). Indeed, two Washington Department of Fish and Wildlife biologists have independently observed hunting dogs try to dig presumed rodents out of rotting logs (Art Kendal [retired] and Curran Cosgrove, personal communication). The rodents, in turn, may be an important resource for local Northern harriers (Circus hudsonius) that commonly patrol the marsh.
Whatever ecological role is played by large wood in tidal marshes, a complete understanding of this role and how it varies across the landscape requires understanding how large wood is itself distributed and what processes control that distribution. Thus, the work presented here is only an initial step in better understanding the ecological effects of large wood in tidal marshes.
Tidal Marsh Restoration Guidance
Habitat restoration designers should consider how the processes described here—riverine delivery, tidal and storm redistribution, and topographic and vegetation trapping—apply to their particular site, so that they only add large wood to their restoration sites in appropriate quantities and at appropriate locations. For example, are upstream sources of large wood limited by dams, levees, or deforestation? If so, then large wood supplementation of a restoration site may be appropriate. Where wood supply is not constrained, sites with high fetch may require little if any large wood supplementation to match reference conditions, because wood may easily recruit to the site and be retained. Is the restoration site adjacent to or far from large river distributaries? If nearby, then wood supplementation may be unnecessary, assuming unconstrained wood supply; if far from a distributary, then it may be inappropriate, because large wood would not naturally accumulate in such areas. It is probably inappropriate to place wood in low-elevation marshes (relative to the tidal frame), because these are not natural locations for wood trapping. Likewise, within blind tidal channels, wood is more appropriately placed in channels 2–4-m wide than in wider reaches.
All of this, of course, assumes that large wood has an important ecological role in tidal marshes. This seems likely (e.g., Hood 2007a), but there is considerable need to conclusively demonstrate and quantify its ecological role, particularly with regard to estuarine fish, and particularly for species of management concern.
References
Allen, J.R.L. 2000. Late Flandrian (Holocene) tidal palaeochannels, Gwent Levels (Severn Estuary), SW Britain: Character, evolution and relation to shore. Marine Geology 162: 353–380.
Allen, J.R.L. 1997. Simulation models of salt-marsh morphodynamics: Some implications for high-intertidal sediment couplets related to sea-level change. Sedimentary Geology 113: 211–223.
Benke, A.C., and B. Wallace. 2003. Influence of wood on invertebrate communities in streams and rivers. In The Ecology and Management of Wood in World Rivers, ed. S.V. Gregory, K.L. Boyer, and A.M. Gurnell, 149–177. Bethesda, Maryland: American Fisheries Society.
Collins, B.D., D.R. Montgomery, and A.D. Haas. 2002. Historical changes in the distribution and functions of large wood in Puget lowland rivers. Canadian Journal of Fisheries and Aquatic Science 59: 66–76.
David, A.T., C.A. Simenstad, J.R. Cordell, J.D. Toft, C.S. Ellings, A. Gray, and H.B. Berge. 2016. Wetland loss, juvenile salmon foraging performance, and density dependence in Pacific Northwest estuaries. Estuaries and Coasts 39: 767–780.
Diefenderfer, H.L., and D.R. Montgomery. 2009. Pool spacing, channel morphology, and the restoration of tidal forested wetlands of the Columbia River, U.S.A. Restoration Ecology 17: 158–168.
Eilers, H.P. 1975. Plants, plant communities, net production and tide levels: the ecological biogeography of the Nehalem Salt Marshes, Tillamook County, Oregon. Ph.D. thesis. Corvallis, Oregon: Oregon State University.
Ellings, C.S., M.J. Davis, E.E. Grossman, I. Woo, S. Hodgson, K.L. Turner, G. Nakai, J.E. Takekawa, and J.Y. Takekawa. 2016. Changes in habitat availability for outmigrating juvenile salmon (Oncorhynchus spp.) following estuary restoration. Restoration Ecology 24: 415–427.
Everett, R.A., and G.M. Ruiz. 1993. Coarse woody debris as a refuge from predation in aquatic communities. Oecologia 93: 475–486.
Fox, M., and S. Bolton. 2007. A regional and geomorphic reference for quantities and volumes of instream wood in unmanaged forested basins of Washington State. North American Journal of Fisheries Management 27: 342–359.
Gonor, J.J., J.R. Sedell, and P.A. Benner. 1988. What we know about large trees in estuaries, in the sea, and on coastal beaches. In From the forest to the sea, a story of fallen trees, eds. C. Maser, R.F. Tarrant, J.M. Trappe, and J.F. Franklin, 83–112. U.S. Forest Service General Technical Report GTR-PNW-229. Portland, Oregon: Pacific Northwest Research Station.
Gurnell, A.M., H. Piegay, F.J. Swanson, and S.V. Gregory. 2002. Large wood and fluvial processes. Freshwater Biology 47: 601–619.
Hindell, J.S. 2007. Determining patterns of use by black bream Acanthopagrus butcheri (Munro, 1949) of re-established habitat in a south-eastern Australian estuary. Fish Biology 71: 1331–1346.
Hood, W.G. 2015. Geographic variation in Puget Sound tidal channel planform geometry. Geomorphology 230: 98–108.
Hood, W.G. 2014. Differences in tidal channel network geometry between reference marshes and marshes restored by historical dike breaching. Ecological Engineering 71: 563–573.
Hood, W.G. 2007a. Large woody debris influences vegetation zonation in an oligohaline tidal marsh. Estuaries and Coasts 30: 441–450.
Hood, W.G. 2007b. Landscape allometry and prediction in estuarine ecology: Linking landform scaling to ecological patterns and processes. Estuaries and Coasts 30: 895–900.
Hood, W.G. 2006. A conceptual model of depositional, rather than erosional, tidal channel development in the rapidly prograding Skagit River Delta (Washington, USA). Earth Surface Processes and Landforms 31: 1824–1838.
Hood, W.G. 2004. Indirect environmental effects of dikes on estuarine tidal channels: Thinking outside of the dike for habitat restoration and monitoring. Estuaries 27: 273–282.
Hood, W.G. 2002. Landscape allometry: From tidal channel hydraulic geometry to benthic ecology. Canadian Journal of Fisheries and Aquatic Sciences 59: 1418–1427.
Hood, W.G., E.E. Grossman, and C. Veldhuisen. 2016. Assessing tidal marsh vulnerability to sea-level rise in the Skagit River Delta. Northwest Science 90: 79–93.
Johannessen, C.L. 1964. Marshes prograding in Oregon: Aerial photographs. Science 146: 1575–1578.
Jones, K.K., K. Anlauf-Dunn, P.S. Jacobsen, M. Strickland, L. Tennant, and S.E. Tippery. 2014. Effectiveness of instream wood treatments to restore stream complexity and winter rearing habitat for juvenile coho salmon. Transactions of the American Fisheries Society 143: 334–345.
Kirwan, M.L., S. Temmerman, E.E. Skeehan, G.R. Guntenspergen, and S. Fagherazzi. 2016. Overestimation of marsh vulnerability to sea level rise. Nature Climate Change 6: 253–260.
Kramer, N., and E. Wohl. 2017. Rules of the road: A qualitative and quantitative synthesis of large wood transport through drainage networks. Geomorphology 279: 74–97.
Kramer, N., and E. Wohl. 2015. Driftcretions: The legacy impacts of driftwood on shoreline morphology. Geophysical Research Letters 42: 5855–5864. https://doi.org/10.1002/2015GL064441.
Lester, R.E., and A.J. Boulton. 2008. Rehabilitating agricultural streams in Australia with wood: A review. Environmental Management 42: 310–326.
Macdonald, J.S., C.D. Levings, C.D. McAllister, U.H.M. Fagerlund, and J.R. McBride. 1988. A field experiment to test the importance of estuaries for Chinook salmon (Oncorhynchus tshawytscha) survival: Short-term results. Canadian Journal of Fisheries and Aquatic Sciences 45: 1366–1377.
MacLennan, A. 2005. An analysis of large woody debris in two Puget Sound salt marshes; Elger Bay, Camano Island, and Sullivan Minor Marsh, Padilla Bay. MS thesis. Bellingham, Washington: Western Washington University.
Magnusson, A., and R. Hilborn. 2003. Estuarine influence on survival rates of Coho (Oncorhynchus kisutch) and Chinook salmon (Oncorhynchus tshawytscha) released from hatcheries on the U.S. Pacific Coast. Estuaries 26: 1094–1103.
McMahon, T.E., and L.B. Holtby. 1992. Behaviour, habitat use, and movements of coho salmon (Oncorhynchus kisutch) smolts during seaward migration. Canadian Journal of Fisheries and Aquatic Sciences 49: 1478–1485.
Montgomery, D.R., B.D. Collins, J.M. Buffington, and T.B. Abbe. 2003. Geomorphic effects of wood in rivers. In The Ecology and Management of Wood in World Rivers, ed. S.V. Gregory, K.L. Boyer, and A.M. Gurnell, 21–47. Bethesda, Maryland: American Fisheries Society.
Roni, P., T. Beechie, G. Pess, and K. Hanson. 2015. Wood placement in river restoration: Fact, fiction, and future direction. Canadian Journal of Fisheries and Aquatic Science 72: 466–478.
Simenstad, C.A., A. Wick, S. Van de Wetering, and D.L. Bottom. 2003. Dynamics and ecological functions of wood in estuarine and coastal marine ecosystems. In The Ecology and Management of Wood in World Rivers, ed. S.V. Gregory, K.L. Boyer, and A.M. Gurnell, 265–277. Bethesda, Maryland: American Fisheries Society.
Sokal, R.R., and F.J. Rohlf. 1995. Biometry. New York: W. H. Freeman.
Steel, E.A., R.J. Naiman, and S.D. West. 1999. Use of woody debris piles by birds and small mammals in a riparian corridor. Northwest Science 73: 19–26.
Tonnes, D.M. 2008. Ecological functions of marine riparian areas and driftwood along north Puget Sound shorelines. Master’s Thesis. School of Marine and Environmental Affairs, University of Washington, Seattle.
Van de Wetering, S. 2001. Juvenile salmonid densities across variable habitats within the Siletz River estuary. Technical Report 2001E3. Siletz, Oregon: Confederated Tribes of the Siletz Indians.
Wick, A.J. 2002. Ecological function and spatial dynamics of large woody debris in oligohaline brackish estuarine sloughs for juvenile Pacific salmon. MS thesis. Seattle: University of Washington.
Wipfli, M.S., J.S. Richardson, and R.J. Naiman. 2007. Ecological linkages between headwaters and downstream ecosystems: Transport of organic matter, invertebrates, and wood down headwater channels. Journal of the American Water Resources Association 43: 72–85.
Yang, Z., T. Khangaonkar, M. Calvi, and K. Nelson. 2010. Simulation of cumulative effects of nearshore restoration projects on estuarine hydrodynamics. Ecological Modelling 221: 969–977.
Zar, J.H. 1984. Biostatistical Analysis. Englewood Cliffs, New Jersey: Prentice Hall.
Acknowledgements
Thanks to Curt Veldhuisen, Mike LeMoine, Kim Jones, and Tish Conway-Cranos for reviewing the draft manuscript. This work was funded by the US Environmental Protection Agency (grant no. PA-00J322-01) and the Washington Department of Fish and Wildlife Estuary and Salmon Recovery Program (project no. RCO 18-2253).
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Competing Interests
The author declares no competing interests.
Additional information
Communicated by Stijn Temmerman
Supplementary Information
Below is the link to the electronic supplementary material.
Rights and permissions
Springer Nature or its licensor holds exclusive rights to this article under a publishing agreement with the author(s) or other rightsholder(s); author self-archiving of the accepted manuscript version of this article is solely governed by the terms of such publishing agreement and applicable law.
About this article

Cite this article
Hood, W.G. Distribution of Large Wood in River Delta Tidal Marshes: Implications for Habitat Restoration. Estuaries and Coasts 46, 109–127 (2023). https://doi.org/10.1007/s12237-022-01122-5
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s12237-022-01122-5