
Abstract
Many coastal-dependent species have undergone large-scale population declines due to impacts from habitat loss, including American oystercatchers (Haematopus palliatus). Islands along the Big Bend region of Florida’s Gulf Coast provide important nesting habitat for oystercatchers, but reproductive success here is low and habitat degradation and loss are a major concern. To determine rates and characteristics of habitat loss, we quantified changes in island sizes within two major breeding areas of the Big Bend: the Barge Canal spoil islands and natural islands at Cedar Key. We digitized aerial photographs from the past ~ 40 years, measured area and shoreline retreat of nesting islands, and identified trends over time by fitting linear mixed effects models. The total area of the ten Barge Canal spoil islands decreased by 55% between 1979 and 2016. At Cedar Key, the total area of the six islands measured decreased by 39% between 1974 and 2016, 85% of which occurred after 1995, indicating an increase in erosion rates correlated with oyster reef declines. Changes in available nesting habitat varied between the Barge Canal and Cedar Key islands due to differences in physical attributes; however, all islands significantly decreased in size over time. Given the long life and high site fidelity of American oystercatchers, these islands may currently be acting as an ecological trap for this species. Climate change, sea-level rise, and loss of oyster reefs are likely to continue to drive oystercatcher habitat loss throughout their range; thus, creation and restoration of oyster reefs and nesting islands will become increasingly important.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Coastal environments are highly dynamic systems which are often exposed to rapid and extreme changes resulting in both positive and negative effects on coastal-dependent species. These effects are confounded by significant anthropogenic pressures such as coastline development, recreational use, overfishing, and upland freshwater usage (which impacts discharge to estuaries) and effects from climate change (Leatherman et al. 2000; Kirby 2004; Lotze et al. 2006; Seavey et al. 2011a). Projected effects of climate change globally include sea-level rise and increased intensity and frequency of storms, which erode and degrade coastal habitats (Emanuel 2005; Webster et al. 2005). As the processes of climate change and sea-level rise accelerate, coastal habitats are expected to experience increased levels of flooding and saltwater intrusion, leading to dramatic habitat loss and change (CCSP 2009; Nicholls and Cazenave 2010; Burger 2018). In addition, the impact of these cumulative effects has been apparent in the worldwide decline and degradation of oyster reefs (Beck et al. 2011), which play a vital role in many coastal habitats as ecosystem engineers and function to attenuate wave energy and protect coastlines from erosion (Piazza et al. 2005; Scyphers et al. 2011). The loss of oyster reefs can lead to a negative feedback loop by increasing coastal erosion. The loss and degradation of coastal habitats are the leading causes of extinction risk for habitat specialists including many species that use coastal ecosystems for reproduction, such as shorebirds (Owens and Bennett 2000; Brooks et al. 2002; Franco et al. 2006; Mazaris et al. 2009). As a result, many coastal-dependent shorebirds have undergone large-scale population declines (Erwin et al. 2011; Burger and Niles 2013).
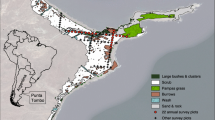
The American oystercatcher (Haematopus palliatus; hereafter oystercatcher) is a large shorebird that breeds along the Atlantic and Gulf coast regions of the USA. The oystercatcher feeds primarily on marine bivalves, depends on coastal areas that support intertidal shellfish beds, and nests near these foraging areas, which makes the oystercatcher especially susceptible to changes in coastal habitats (American Oystercatcher Working Group et al. 2012). The oystercatcher is listed as a species of high conservation concern in the US Shorebird Conservation Plan (Brown et al. 2001) and is listed as Threatened in Florida due to low numbers and limited breeding distribution (Florida Fish and Wildlife Conservation Commission 2011). A statewide analysis conducted in 1994 concluded that there was insufficient suitable habitat available for the long-term stability of oystercatchers in Florida (Cox et al. 1994). Florida’s Big Bend region provides important wintering and nesting habitat for oystercatchers (American Oystercatcher Working Group et al. 2012; Brush et al. 2017; Fig. 1); this region is characterized by a largely undeveloped coastline that is mostly protected from further development. However, over most of the Big Bend region, nesting habitat is limited to a few small (< 3 ha) offshore islands; alternative nesting habitats are not available as they are in other regions. The majority of nesting oystercatchers (47%) in Florida nest on small oyster and shell islands or rakes similar to the Cedar Key islands within the Big Bend (Florida Shorebird Database 2019).

American oystercatcher historical and current nesting islands (black) in the Big Bend region of Florida (gray) at our two study sites in Levy and Citrus counties
The unique underlying hard limestone substrate found along the eastern Gulf of Mexico, in particular in the Big Bend of Florida, reduces subsidence and associated rates of sea-level rise that have been documented in other parts of the Gulf (Swanson and Thurlow 1973; Morton et al. 2004). The Big Bend of Florida is a low wave-energy, shallow, highly karstic region (Johnson and Barbour 1990). Due to the karst nature of this region, there is no significant sediment transport suitable for beach development or maintenance, but shell derived from oyster reefs and offshore shoals line the small offshore islands (Johnson and Barbour 1990). Because it is a sand-starved coastline, this region lacks barrier islands which typically provide protection from ocean and storm forces (Hine et al. 1988) and are commonly used by other breeding populations of oystercatchers (Koczur et al. 2014; Schulte and Simons 2015). In addition, although this region is relatively protected from coastline development, large areas have experienced significant losses in the extent of oyster reefs in the last two decades (Seavey et al. 2011b). Decreased freshwater inputs from rivers have contributed to decreased survival and recruitment of oysters, subsequent erosion of oyster reefs, and ultimately, irreversible loss of substrate critical for oyster resettlement (Seavey et al. 2011b; Frederick et al. 2016). Oyster reefs, directly and indirectly, provide critical foraging, roosting, and nesting habitats for oystercatchers (Brown et al. 2005).
Declines of oyster reefs coupled with sea-level rise in the Big Bend region suggest many small islands used by nesting shorebirds are likely to be impacted. Oystercatchers nest on the ground along the shoreline, making their nests and young susceptible to shoreline erosion, high tides, and storm surges; overwash has been the largest known contributor to nest loss in the region since 2011 (Florida Shorebird Database 2019). Although overwash is frequent at nests placed at low elevations and/or near the high tide line (Jodice et al. 2014), it also occurs at nearly all nesting areas throughout oystercatchers’ range during storm and high wind events (Schulte and Simons 2015; Florida Shorebird Database 2019). In the Cedar Key area, the estimated rate of sea-level rise is 2.08 ± 0.18 mm/year (based on monthly data from 1914 to 2017), which is near the global sea-level rise of 1.70 ± 0.30 mm/year for the twentieth century (NOAA 2009). However, global sea-level rise has an estimated climate change–driven acceleration of 0.08 ± 0.03 mm/year2 and the Big Bend’s extensive low-elevation coastal habitats make it especially susceptible to any change in sea level (Geselbracht et al. 2011; Nerem et al. 2018). The limited current availability of suitable nesting habitat in Florida, coupled with the likely future effects from climate change, raises important conservation issues for oystercatchers and other avian species dependent on these coastal habitats.
The islands in the Big Bend region support the second-largest concentration of nesting oystercatchers in Florida; the next closest nesting concentrations are ~ 168 km to the north and ~ 150 km to the south of our study sites. Because the region has no barrier islands or large shell and marsh rakes, the breeding habitat for oystercatchers here is different and more limited than along the Atlantic and other parts of the Gulf coast. Considering that nesting habitat is potentially limiting for oystercatchers, and that oyster reefs are declining in the Big Bend region, our primary objective was to quantify changes in oystercatcher nesting habitat here over time. We secondarily evaluated changes in oystercatcher reproductive success to determine how they related to habitat loss in the region. Our goal was to quantify breeding habitat loss in the region and lay the foundation for high-priority habitat restoration to benefit American oystercatchers.
Methods
Study Sites
Our study area included the two primary sites currently used by nesting oystercatchers in the southern part of the Big Bend region: the spoil islands of the Barge Canal within the Cross Florida Greenway State Recreation and Conservation Area (hereafter Barge Canal) and natural islands in the Cedar Key area (Fig. 1). These sites support two of only three nesting concentrations of oystercatchers in Florida’s Big Bend.
The Barge Canal spoil islands were created in the mid-1960s during the dredging of the eventually abandoned Cross Florida Barge Canal. At the time of construction, 15 islands were created using dredged limestone rock, 14 of which formed a linear chain perpendicular to the coast (Schreiber and Schreiber 1978). Since the initial construction, these islands have been left untouched. Currently, ten spoil islands are above mean high water, six of which are used by nesting oystercatchers. These spoil islands range from 2 to 5.75 km from the mainland and range in size from 0.1 to 1.5 ha. Surface substrate primarily consists of limestone rock (generally 5–20-cm diameter), shell, and only small amounts of sand. Above the tidal range, most of the islands were vegetated by dense low shrubs, grasses, and woody vegetation, including some trees. Nesting oystercatchers have used six to eight of the spoil islands since at least 1976, supporting approximately 15–28 total pairs annually, representing ~ 11% of the 2019 statewide population (Schreiber and Schreiber 1978; Florida Shorebird Database 2019). Currently on these islands, oystercatchers nest on open substrate and select for nest sites away from vegetation, as is typical for the species (American Oystercatcher Working Group et al. 2012). In addition, these islands are currently used by several other waterbirds, including Wilson’s plovers (Charadrius wilsonia), least terns (Sternula antillarum), and numerous heron species (Ardeidae). Nesting birds have never used four of the islands, so we excluded these from our study.
The second study area consisted of seven naturally occurring islands near Cedar Key. Six islands are characterized by sand and shell substrates with variable amounts of vegetation including ground cover and some tall woody vegetation. The seventh island, Corrigan’s Reef, is a shell rake with minimal vegetation. Sizes of these islands range from 0.35 to 3.0 ha. Historically, all nesting islands in both areas had naturally occurring oysters on them and presumably had high enough elevation for successful nesting. In addition, oyster reefs not directly connected to the islands have historically surrounded many Cedar Key islands. Since 2001, 6–12 pairs of oystercatchers have used the Cedar Key area for nesting annually; there are no prior estimates of breeding populations for this area. Currently, oystercatchers use all nesting habitat above the mean high-water line on these islands. Several of the Cedar Key islands also support populations of breeding Wilson’s plovers and least terns.
Data Acquisition and Analysis
We obtained aerial imagery (resolution = 0.3 m) from the Florida Department of Transportation, Survey and Mapping Office (LABINS 2018) to determine temporal and spatial shoreline changes to nesting islands in our study area. Imagery available for the two sites differed in coverage so there were differences in the years used in our analysis, but they covered similar periods. For Barge Canal, we used imagery from 5 years from 1979 to 2017. For the Cedar Key area, we used imagery from 7 years from 1974 to 2016. For aerial photographs that were not already georeferenced (years before 1995), we manually georeferenced each photograph using control points (e.g., corners of buildings, road intersections). We then hand-digitized the shorelines of all islands used historically by nesting oystercatchers, at a 1:3000 scale along the estimated high-water line (digitized island shapefiles available at: http://ufdc.ufl.edu/IR00010764/00001 and http://ufdc.ufl.edu/IR00010763/00001). The exact height of the tide for each aerial image was unknown; however, when water levels appeared to be below high tide, we used the wet-dry line in the sand or the wrack line as estimates of mean high water. For each island, we used the digitized shorelines to calculate island area in each sample year. We measured linear shoreline retreat (m) on the ocean-side of islands along a single transect located where shoreline loss was the greatest to measure changes in the shoreline from year to year. We also calculated linear shoreline retreat (m) on the mainland side directly across the island from our ocean-side transect. Corrigan’s Reef in Cedar Key is a highly dynamic shell rake and the entire rake shifts over time; therefore, calculating shoreline retreat was not possible. Additionally, two islands were excluded from Barge Canal because they were only above water at all tide levels in the first image used (1979) and were subtidal by the second date in the sequence (1995).
Some measuring errors at individual points are assumed between periods due to varying tide heights, substrate types, image quality, and human error in digitizing and georeferencing. We did not determine an absolute value of the overall predicted horizontal accuracy; however, for our dataset, we assumed an uncertainty of 5 m based on the constraints described by Crowell et al. (1991) which include accounting for distortion of photos, inaccurate photo control points, errors in delineation of mean high water line, and digitizer-related errors. When translating this uncertainty in shoreline position to calculations of island area, the size and shape of the island influence the level of uncertainty in estimates of island area. For example, based on the ± 5-m uncertainty in shoreline position, a perfectly circular island with an area equal to that of an average size island in our study area (20,950 m2) would have an estimated uncertainty of − 2441 m2 to + 2696 m2 in area. We effectively minimized uncertainty by using multiple images that covered a 40-year period, which results in a high level of accuracy of overall trends (Fisher and Overton 1994).
We fit linear mixed effects models using measured changes in island area and shoreline retreat to identify trends over time. In our models, we used year and site as fixed effects and included random effects for intercept and slope to account for differences in individual islands.
We assessed temporal trends of key reproductive metrics to determine whether changes in habitat were associated with changes in reproduction, including the number of nesting pairs for each site per year (for all years available) from 1977 to 2018, overall site productivity (number of fledglings per nesting pair) from 2011 to 2018, and the sources of any known nest failures (Schreiber and Schreiber 1978; Florida Shorebird Database 2019). The Florida Fish and Wildlife Conservation Commission (FWC) began monitoring nesting shorebirds 2–4 times monthly in our study area after 2011. We increased the monitoring effort to twice-weekly visits in 2017 and installed camera traps near the majority of nests to assist with determining nest outcomes and sources of failure. Data were sporadic due to the intermittent nature of surveys during 1977–2011, before the implementation of systematic monitoring of nesting oystercatchers within our study area.
Results
Barge Canal
The total area of the ten Barge Canal islands decreased by 55% between 1979 and 2016 (F1, 31 = 51.58, P < 0.0001). Two of the westernmost spoil islands became subtidal by 1995 and a third was subtidal by 2008 (Fig. 2). The now subtidal spoil islands were originally the smallest islands (< 0.2 ha). Two of the three submerged spoil islands historically supported nesting birds including at least three pairs of oystercatchers and a seabird colony with an estimated 275 birds consisting of laughing gulls (Leucophaeus atricilla) and royal terns (Thalasseus maximus; Schreiber and Schreiber 1978). Least terns are the only seabirds that still nest at Barge Canal. Ocean-side shoreline retreat varied by island and increased over time (likelihood ratio = 44.27, P < 0.0001) at an average rate of 0.89 m/year (F1, 27 = 102.09, P < 0.0001; Fig. 3) Land-side shoreline retreat varied by island and increased over time at an average rate of 0.22 m/year (F1, 27 = 9.65, P = 0.004).

Cumulative change in area of Barge Canal islands in the Big Bend region of Florida, 1979–2017, compared with initial island area in 1979 as determined by aerial imagery. Islands were numbered from nearshore (1) to offshore (10)

Cumulative ocean-side linear shoreline retreat (m) of Barge Canal spoil islands in the Big Bend region of Florida, 1979–2017, as determined by aerial photography

Cumulative change in area of Cedar Key islands in the Big Bend region of Florida, 1974–2016, compared with initial island area in 1974 as determined by aerial photography
In 2016, Hurricane Hermine caused widespread environmental impacts across the Big Bend. We observed changes at Barge Canal as a direct result of the hurricane. Three of the spoil islands increased in area (\( \overline{x} \) = 4% ± 1.8%) following the hurricane. In contrast, erosion occurred on the westernmost spoil island (islands 4, 5, and 7; Fig. 1), lowering elevation and/or decreasing total area (\( \overline{x} \) = 8% ± 0.3%). Additionally, field observations documented reductions in vegetation and increased elevation of three of the spoil islands as a result of wave action and storm surge.
Overall oystercatcher productivity for Barge Canal was low in most years (Table 1). Nest overwash caused the failure of 17% (n = 4) and 8% (n = 3) of oystercatcher nest attempts at Barge Canal in 2017 and 2018 respectively. Observations/data suggest overwash was the primary source of known nest failure from 2011 to 2016, but most causes of nest failure were documented as “unknown” due to inconsistent survey efforts and the challenge of determining nest outcomes in limestone nesting substrate (i.e., absence of predator tracks; Florida Shorebird Database 2019). There were large changes among years but no clear pattern or trend in total number nesting pairs recorded using the site over time (Table 1); however, survey effort was inconsistent.
Cedar Key
The total area of all nesting islands within Cedar Key decreased by 39% between 1974 and 2016, with 85% of the decrease occurring after 1995. Decreases in area varied widely across islands, ranging from a 32% loss of Rattlesnake Key to 100% loss of Derrick Key (Fig. 4). The rate of change in island area was slightly faster in Cedar Key than Barge Canal (F1, 79 = 16.40, P = 0.018). Shorelines were generally stable over time on the mainland sides of the islands (F1, 35 = 0.28, P = 0.599), whereas ocean-side shoreline retreat did increase (likelihood ratio = 54.18, P < 0.0001) at an average rate of 1.28 m/year (F1, 35 = 120.32, P < 0.0001). The three northernmost islands had the highest amount of shoreline retreat (Fig. 5).

Cumulative ocean-side linear shoreline retreat (m) of Cedar Key islands in the Big Bend region of Florida, 1974–2016, as determined by aerial photography
Overall, oystercatcher productivity for Cedar Key was low, producing only four fledglings in 2017 (6 pairs) and 2018 (8 pairs) combined (Table 1). Overwash was a major source of nest failure for oystercatcher nesting in Cedar Key. We documented that 57% (n = 8) and 29% (n = 4) of known nest attempts at Cedar Key failed due to overwash in 2017 and 2018, respectively. In both years, not all overwash events were associated with storm events; some islands experienced frequent near-complete overwash that was associated with spring tides or regular high tides and moderate wind. For example, camera traps on Gomez Key documented overwash of most or the entire island six times during a 6-week period in April 2017. Similarly, oystercatcher nests on Rattlesnake Key were overwashed even more frequently in 2017 and 2018, but we did not have any cameras on the island to document specific events.
Comparable with Barge Canal, there were substantial fluctuations but no pattern or trend in the number of nesting pairs recorded using the Cedar Key site over time (Table 1). However, a clear influence of habitat loss was observed with Derrick Key, which historically supported an average of 5–6 nesting pairs of oystercatchers annually. By 2014, Derrick Key was no longer a suitable nesting habitat because of erosion. In addition, it is unclear where the nesting pairs from Derrick Key relocated once the area became unsuitable nesting habitat.
Discussion
All nesting islands within our study area experienced significant losses in total area in the past 20–40 years. Multiple islands at both sites have become subtidal and several others have become extremely susceptible to overwash due to erosion and an assumed loss in elevation. The two islands in Cedar Key that historically supported the highest numbers of nesting oystercatchers have experienced the highest percentage of area lost. This loss of important nesting habitat, coupled with current rates of erosion and the already limited amount of nesting habitat in the region, makes habitat availability a major limiting factor for oystercatchers nesting in the Big Bend region (Cox et al. 1994). Barge Canal had a lower rate of erosion compared with Cedar Key, which suggests the spoil islands are more stable under current conditions. Unlike typical dredge spoil islands and the islands at Cedar Key, the Barge Canal islands primarily consist of heavy and large-size limestone rock and are more resistant to erosion and movement by wave action and the influences of climate change.
We found that at the natural islands of Cedar Key, the majority of erosion occurred within the last 20 years. The timing of the rapid erosion of Cedar Key islands closely follows that of losses of oyster reefs in the area, suggesting the reefs were stabilizing these islands (Seavey et al. 2011b). In addition, we documented substantial erosion of the Cedar Key islands along shorelines with the greatest wave action and wind energy. While the extent of oyster reefs surrounding the islands in our study area varied, five of the seven Cedar Key islands historically had oyster reef habitat on all sides, but now those reefs are largely reduced (Fig. 6). Oyster reef habitat, as part of a living shoreline, provides a critical role in shoreline protection through wave attenuation (Dame and Patten 1981; Meyer et al. 1997; Piazza et al. 2005; Scyphers et al. 2011; Manis et al. 2015). When oyster reefs decline as they have in the Big Bend region, this important ecosystem service is lost, causing shoreline erosion to increase (Meyer et al. 1997).

Example of habitat change in the form of oyster reef loss (small land masses near two main islands in photos) and erosion of two nesting islands: Derrick Key (100% loss) and McClammory (32% loss) in Cedar Key, FL, from 1982 (left) to 2016 (right). Imagery from LABINS 2018
The reduction of healthy oysters in Cedar Key has led to accelerated habitat loss, but the primary impacts of the loss of oysters at the Barge Canal islands means a loss in critical foraging areas adjacent to nesting islands. In 2017 and 2018, 80% of oystercatcher nests in Cedar Key were located within 100 m of high food abundance (> 50 live oysters per m2), while at the Barge Canal, only 18% of nests fell into this category (Vitale 2019). The loss of foraging areas adjacent to nesting islands could mean breeding adults must fly to distant areas to forage, which in turn can increase predation on chicks because parents are less available to protect them (Nol 1989; Thibault et al. 2010). Oystercatchers tend to nest at higher densities and fledge more chicks when they have direct access to foraging areas (Nol 1989; Ens et al. 1992). Additionally, these adults may deliver less food to their chicks than birds with foraging areas within their nesting territory, resulting in lower growth and fledging rates (Ens et al. 1992; Heg and van der Velde 2001).
The high food abundance available near the Cedar Key nest sites coupled with the limited habitat availability for nesting oystercatchers in the area is the primary reason birds continue to nest at our study sites. Oystercatchers typically exhibit high nest site fidelity (American Oystercatcher Working Group et al. 2012; Murphy et al. 2017) and will return to nest in areas even if habitat is degrading, resulting in a potential ecological trap (Powell and Collier 2000; George et al. 2004; Anteau et al. 2012), in part due to the high cost of changing nesting sites (Switzer 1993). In Cedar Key, we found no relationship between declining habitat availability and nesting population size despite productivity levels too low to support a stable population. Nest site fidelity is common among shorebirds including other species of oystercatchers, but they generally exhibit lower site fidelity to areas that failed to produce chicks. High nest site fidelity at locations that are frequently flooded has been observed elsewhere; however, this behavior is often associated with successful renesting after initial nest loss (Thibault 2008). There is variable productivity at our sites, where some years it is higher than the range-wide average but most years no fledglings are produced (Nol 1989; McGowan et al. 2005; Traut et al. 2006; American Oystercatcher Working Group et al. 2012; Vitale 2019). The cumulative effect of some productive years and the high cost of changing nesting locations contributes to the site fidelity demonstrated by the nesting pairs in our study area. Additionally, it is unclear where breeding oystercatchers go when breeding habitat becomes unusable; for example, it is unknown where the oystercatchers that nested on Derrick Key went after that island became unusable.
In contrast to the natural islands at Cedar Key, the Barge Canal spoil islands were used by more oystercatchers for nesting despite the fact that the birds that nested there had to travel farther to find food. Other studies have shown the importance of managed spoil islands in providing essential habitat for nesting oystercatchers in those areas where beach sites are constrained by numerous threats (McGowan et al. 2005; Virzi et al. 2016). For example, in North Carolina, dredge spoil islands supported higher densities of nesting oystercatchers than naturally occurring barrier islands in the same area (McGowan et al. 2005). In New Jersey, the reproductive success rate on spoil islands was an order of magnitude greater than on barrier beaches (Virzi et al. 2016). However, the spoil island habitat in these studies was associated with lower predation and human disturbance rates in these areas, which was not the case for the Barge Canal during 2017 and 2018 (Vitale 2019). Predation and human disturbance on spoil islands are highly variable, and threats to birds nesting on spoil islands have increased over time. Declines of small coastal islands used by nesting birds have been documented along the Atlantic Coast, therefore limiting the amount of potential habitat available for nesting birds (George et al. 2004; Erwin et al. 2011). As smaller islands continue to erode and disappear, well-managed predator- and disturbance-free spoil island habitats will become increasingly important for breeding shorebirds as climate and anthropogenic pressures increase in coastal areas (Colwell 2010; Burger 2018).
Because oystercatchers are an obligate coastal species that use low-elevation coastal habitats for breeding and nonbreeding, they are particularly vulnerable to the effects of sea-level rise (Clay et al. 2014). Oystercatcher habitat will be impacted as estuarine and open water areas in the region migrate into higher elevations, converting other habitat types in the process (Castaneda and Putz 2007; Geselbracht et al. 2011). These changes driven by rises in sea level include forecasted declines in suitable habitat and carrying capacity of shorebirds in Florida (Aiello-Lammens et al. 2011). The increased severity and frequency of storms as a result of climate change (IPCC 2014) may be as much of a threat as sea-level rise to breeding shorebirds (Seavey et al. 2011a). Additionally, a shift in the timing of storm events earlier in the season could significantly decrease productivity rates (Aiello-Lammens et al. 2011). For example, Tropical Storm Alberto in 2018 caused all of the oystercatcher nests (n = 32) at St. Marks National Wildlife Refuge in the northern Big Bend to get overwashed and as a result, productivity was nearly zero for the area that breeding season (Florida Shorebird Database 2019).
As the processes of climate change and sea-level rise accelerate, the coastal habitats of the Gulf of Mexico are expected to experience increased levels of flooding and saltwater intrusion, leading to accelerated and dramatic habitat loss and change (Burger 2018). The loss of coastal islands and erosion from the cumulative effects of sea-level rise, increased storm frequencies, and worldwide loss and degradation of oyster reefs is likely to decrease oystercatcher abundance and productivity in the Gulf of Mexico and globally (Clay et al. 2014; Burger 2018). Climate change and the associated effects will continue to reshape the coastal landscape and reduce available shorebird habitat (Colwell 2010). In addition, oyster reef declines are not likely to reverse on their own, which will result in continued erosion of important oystercatcher habitat. Shorebirds readily accept and use newly created (such as spoil islands) or restored habitat, which may be an important tool in maintaining populations into the future (Parnell et al. 1986; Scarton et al. 2013; Clay et al. 2014). Potential conservation strategies for oystercatcher nesting habitat may include targeted oyster reef restoration near nesting islands, nesting island management and restoration to reduce nest loss and increase nesting habitat availability, and creation of new nesting habitat. We suggest that managers consider restoration and management strategies that are location-specific and include a combination of tools to ensure resiliency and sustainability of the habitat for productive oystercatcher nesting.
References
Aiello-Lammens, M.E., M.L. Chu-Agor, M. Convertino, R.A. Fischer, I. Linkov, and H.R. Akçakaya. 2011. The impact of sea-level rise on snowy plovers in Florida: Integrating geomorphological, habitat, and metapopulation models. Global Change Biology 17 (12): 3644–3654.
American Oystercatcher Working Group, E. Nol, and R.C. Humphrey. 2012. American oystercatcher (Haematopus palliatus), version 2.0. In Birds of North America, ed. A.F. Poole. Ithaca, New York: Cornell Lab of Ornithology.
Anteau, M.J., T.L. Shaffer, M.H. Sherfy, M.A. Sovada, J.H. Stucker, and M.T. Wiltermuth. 2012. Nest survival of piping plovers at a dynamic reservoir indicates an ecological trap for a threatened population. Oecologia 170 (4): 1167–1179.
Beck, M.W., R.D. Brumbaugh, L. Airoldi, A. Carranza, L.D. Coen, C. Crawford, O. Defeo, G.J. Edgar, B. Hancock, M.C. Kay, H.S. Lenihan, M.W. Luckenbach, C.L. Toropova, G. Zhang, and X. Guo. 2011. Oyster reefs at risk and recommendations for conservation, restoration, and management. Bioscience 61 (2): 107–116.
Brooks, T.M., R.A. Mittermeier, C.G. Mittermeier, G.A.B. Fonseca, A.B. Rylands, W.R. Konstant, P. Flick, J. Pilgrim, S. Oldfield, G. Magin, and C. Hilton-Taylor. 2002. Habitat loss and extinction in the hotspots of biodiversity. Conservation Biology 16 (4): 909–923.
Brown, S.C., C. Hickey, B. Harrington, and R. Gill. 2001. United States shorebird conservation plan. 2nd ed. Manomet, MA: Manomet Center for Conservation Sciences.
Brown, S.C., S. Schulte, B. Harrington, B. Winn, J. Bart, and M. Howe. 2005. Population size and winter distribution of eastern American oystercatchers. Journal of Wildlife Management 69 (4): 1538–1545.
Brush, J.M., A.C. Schwarzer, and P.C. Frederick. 2017. Importance and function of foraging and roost habitat for wintering American oystercatchers. Estuaries and Coasts 40 (1): 286–295.
Burger, J. 2018. Birdlife of the Gulf of Mexico. College Station, Texas: Texas A&M University Press.
Burger, J., and L. Niles. 2013. Shorebirds and stakeholders: Effects of beach closure and human activities on shorebirds at a New Jersey coastal beach. Urban Ecosystem 16 (3): 657–673.
Castaneda, H., and F.E. Putz. 2007. Predicting sea-level rise effects on a nature preserve on the Gulf Coast of Florida: A landscape perspecive. Florida Scientist 70: 166–175.
CCSP. 2009. Coastal sensitivity to sea-level rise: A focus on the mid-Atlantic region. A report by the U.S. Climate Change Science Program and the Subcommittee on Global Change Research. [J. G. Titus, K. E. Anderson, D. R. Cahoon, D. B. Gesch, S. K. Gill, B. T. Gutierrez, E. R.Thieler, and S. J. Williams (Lead Authors)], U.S. Environmental Protection Agency, Washington D.C., USA.
Clay, R.P., A.J. Lesterhuis, S. Schulte, S. Brown, D. Reynolds, and T.R. Simons. 2014. A global assessment of the conservation status of the American oystercatcher Haematopus palliatus. International Wader Studies 20: 62–82.
Colwell, M.A. 2010. Shorebird ecology, conservation, and management. Berkeley, California: University of California Press.
Cox, J.R., M. MacLaughlin, and T. Gilbert. 1994. Closing the gaps in Florida’s wildlife habitat conservation system: Recommendations to meet minimum conservation goals for declining wildlife species and rate plant and animal communities. Florida: Office of Environmental Services, Florida Game and Fresh Water Fish Commission Tallahassee.
Crowell, M., S.P. Leatherman, and M.K. Buckley. 1991. Historical shoreline change: Error analysis and mapping accuracy. Journal of Coastal Research 7: 839–852.
Dame, R.F., and B.C. Patten. 1981. Analysis of energy flows in an intertidal oyster reef. Marine Ecology Progress Series 5: 115–124.
Emanuel, K. 2005. Increasing destructiveness of tropical cyclones over the past 30 years. Nature 436 (7051): 686–688.
Ens, B.J., M. Kersten, A. Brenninkmeijer, and J.B. Hulscher. 1992. Territory quality, parental effort and reproductive success of oystercatchers (Haematopus ostralegus). Journal of Animal Ecology 61 (3): 703–715.
Erwin, R.M., D.F. Brinker, B.D. Watts, G.R. Costanzo, and D.D. Morton. 2011. Islands at bay: Rising seas, eroding islands, and waterbird habitat loss in Chesapeake Bay (USA). Journal of Coastal Conservation 15 (1): 51–60.
Fisher, J., and M. Overton. 1994. Interpretation of shoreline position from aerial photographs. In Coastal engineering, ed. B.L. Edge, 1998–2003. Reston, Virginia: American Society of Civil Engineers.
Florida Fish and Wildlife Conservation Commission. 2011. Biological status review for the American oystercatcher (Haematopus palliates). Tallahassee, Florida: Florida Fish and Wildlife Conservation Commission.
Florida Shorebird Database (FSD) V2.20.0. 2019. https://public.myfwc.com/crossdoi/shorebirds/. Accessed 2019. Online tool for entering and exploring data on Florida’s shorebirds and seabirds, developed and maintained by the Florida Fish and Wildlife Conservation Commission, accessed 1 March 2019.
Franco, A.M.A., J.K. Hill, C. Kitschke, Y.C. Collingham, D.B. Roy, R. Fox, B. Huntley, and C.D. Thomas. 2006. Impacts of climate warming and habitat loss on extinctions at species’ low-latitude range boundaries. Global Change Biology 12 (8): 1545–1553.
Frederick, P., N. Vitale, B. Pine, J. Seavey, and L. Sturmer. 2016. Reversing a rapid decline in oyster reefs: Effects of durable substrate on oyster populations, elevations, and aquatic bird community composition. Journal of Shellfish Research 35 (2): 359–367.
George, R.C., S.H. Schweitzer, and B. Winn. 2004. Reproductive success of American oystercatchers at managed sites in Georgia. Oriole 69: 43–53.
Geselbracht, L., K. Freeman, E. Kelly, D.R. Gordon, and F.E. Putz. 2011. Retrospective and prospective model simulations of sea level rise impacts on Gulf of Mexico coastal marshes and forests in Waccasassa Bay, Florida. Climatic Change 107 (1-2): 35–57.
Heg, D., and M. van der Velde. 2001. Effects of territory quality, food availability and sibling competition on the fledging success of oystercatchers (Haematopus ostralegus). Behavioral Ecology and Sociobiology 49 (2-3): 157–169.
Hine, A.C., D.F. Belknap, J.G. Hutton, E.B. Osking, and M.W. Evans. 1988. Recent geological history and modern sedimentary processes along an incipient, low-energy, epicontinental-sea coastline: Northwest Florida. Journal of Sedimentary Petrology 58: 567–579.
Intergovernmental Panel on Climate Change (IPCC). 2014. Climate change 2014: synthesis report. Contribution of working groups I, II and III to the fifth assessment report of the Intergovernmental Panel on Climate Change. Geneva, Switzerland.
Jodice, P.G.R., J.M. Thibault, S.A. Collins, M.D. Spinks, and F.J. Sanders. 2014. Reproductive ecology of American oystercatchers nesting on shell rakes. Condor 116 (4): 588–598.
Johnson, A.F., and M.G. Barbour. 1990. Dunes and maritime forests. In Ecosystems of Florida, ed. R.L. Myers and J.J. Ewel. Florida: University Presses of Florida, University of Florida.
Kirby, M.X. 2004. Fishing down the coast: Historical expansion and collapse of oyster fisheries along continental margins. Proceedings of the National Academy of Sciences 101 (35): 13096–13099.
Koczur, L.M., A.E. Munters, S.A. Heath, B.M. Ballard, M.C. Green, S.J. Dinsmore, and F. Hernández. 2014. Reproductive success of the American oystercatcher (Haematopus palliatus) in Texas. Waterbirds 37 (4): 371–380.
Land Boundary Information System (LABINS). 2018. On-line survey information and maps for the state of Florida. Florida Department of Environmental Protection. Florida Department of Environmental Protection. http://data.labins.org, accessed 15 Jan 2018.
Leatherman, S.P., K. Zhang, and B.C. Douglas. 2000. Sea level rise shown to drive coastal erosion. Eos, Transactions American Geophysical Union 81 (6): 55–57.
Lotze, H.K., H.S. Lenihan, B.J. Bourque, R.H. Bradbury, R.G. Cooke, M.C. Kay, S.M. Kidwell, M.X. Kirby, C.H. Peterson, J.B.C. Jackson, and M. Bay. 2006. Depletion, degradation, and recovery potential of estuaries and coastal seas. Science 312 (5781): 1806–1809.
Manis, J.E., S.K. Garvis, S.M. Jachec, and L.J. Walters. 2015. Wave attenuation experiments over living shorelines over time: A wave tank study to assess recreational boating pressures. Journal of Coastal Conservation 19 (1): 1–11.
Mazaris, A.D., G. Matsinos, and J.D. Pantis. 2009. Evaluating the impacts of coastal squeeze on sea turtle nesting. Ocean and Coastal Management 52 (2): 139–145.
McGowan, C.P., T.R. Simons, W. Golder, and J. Cordes. 2005. A comparison of American oystercatcher reproductive success on barrier beach and river island habitats in coastal North Carolina. Waterbirds 28 (2): 150–155.
Meyer, D.L., E.C. Townsend, and G.W. Thayer. 1997. Erosion control value of oyster cultch for intertidal marsh. Restoration Ecology 5 (1): 93–99.
Morton, R. A., T. L. Miller, and L. J. Moore. 2004. National assessment of shoreline change: Part 1: Historical shoreline changes and associated coastal land loss along the U.S. Gulf of Mexico: U.S. Geological Survey Open-file Report 2004–1043, 45p.
Murphy, S.P., T. Virzi, and F. Sanders. 2017. Exploring differences in adult survival and site fidelity of migratory and non-migratory American oystercatcher (Haematopus palliatus) populations. Waterbirds 40 (sp1): 32–43.
National Oceanic and Atmospheric Administration (NOAA). 2009. Technical report NOS CO-OPS 53: Sea level variations of the United States 1854–2006. Center for Operational Oceanographic Products and Services, National Ocean Service, National Oceanic and Atmospheric Administration, U.S. Department of Commerce, Silver Spring, Maryland.
Nerem, R.S., B.D. Beckley, J.T. Fasullo, B.D. Hamlington, D. Masters, and G.T. Mitchum. 2018. Climate-change–driven accelerated sea-level rise detected in the altimeter era. Proceedings of the National Academy of Sciences 115 (9): 2022–2025.
Nicholls, R.J., and A. Cazenave. 2010. Sea-level rise and its impact on coastal zones. Science 328 (5985): 1517–1520.
Nol, E. 1989. Food supply and reproductive performance of the American oystercatcher in Virginia. Condor 91 (2): 429–435.
Owens, I.P.F., and P.M. Bennett. 2000. Ecological basis of extinction risk in birds: Habitat loss versus human persecution and introduced predators. Proceedings of the National Academy of Sciences 97 (22): 12144–12148.
Parnell, J.F., R.N. Needham, R.F. Soots Jr., J.O. Fussel III, D.M. Dumond, D.A. McCrimmon Jr., R.D. Bjork, and M.A. Shields. 1986. Use of dredged-material deposition island by birds in coastal North Carolina, USA. Colonial Waterbirds 9: 201–217.
Piazza, B.P., P.D. Banks, and M.K. La Peyre. 2005. The potential for created oyster shell reefs as a sustainable shoreline protection strategy in Louisiana. Restoration Ecology 13 (3): 499–506.
Powell, A.N., and C.L. Collier. 2000. Habitat use and reproductive success of western snowy plovers at new nesting areas created for California. Journal of Wildlife Management 64 (1): 24–33.
Scarton, F., G. Cecconi, and R. Valle. 2013. Use of dredge islands by a declining European shorebird, the Kentish plover Charadrius alexandrinus. Wetlands Ecology and Management 21 (1): 15–27.
Schreiber, R.W., and E.A. Schreiber. 1978. Colonial bird use and plant succession on dredged material islands in Florida. Vol. 1. Washington D.C: United States Army Engineer Waterways Experiment Station.
Schulte, S.A., and T.R. Simons. 2015. Factors affecting the reproductive success of American oystercatchers Haematopus palliatus on the outer banks of North Carolina. Marine Ornithology 43: 37–47.
Scyphers, S.B., S.P. Powers, K.L. Heck, and D. Byron. 2011. Oyster reefs as natural breakwaters mitigate shoreline loss and facilitate fisheries. PLoS One 6 (8): e22396.
Seavey, J.R., B. Gilmer, and K.M. McGarigal. 2011a. Effect of sea-level rise on piping plover (Charadrius melodus) breeding habitat. Biological Conservation 144 (1): 393–401.
Seavey, J.R., W.E. Pine, P. Frederick, L. Sturmer, and M. Berrigan. 2011b. Decadal changes in oyster reefs in the Big Bend of Florida’s Gulf Coast. Ecosphere 2: 114.
Swanson, R.L., and C.I. Thurlow. 1973. Recent subsidence rates along the Texas and Louisiana coasts as determined from tide measurements. Journal of Geophysical Research 78 (15): 2665–2671.
Switzer, P.V. 1993. Site fidelity in predictable and unpredictable habitats. Evolutionary Ecology 7 (6): 533–555.
Thibault, J. M. 2008. Breeding and foraging ecology of the American oystercatcher in the Cape Romain region, South Carolina. M.Sc. Thesis, Clemson University, Clemson, South Carolina.
Thibault, J.M., F.J. Sanders, and P.G. Jodice. 2010. Parental attendance and brood success in American oystercatchers in South Carolina. Waterbirds 33 (4): 511–517.
Traut, A.H., J.M. McCann, and D.F. Brinker. 2006. Breeding status and distribution of American oystercatchers in Maryland. Waterbirds 29 (3): 302–307.
Virzi, T., J.L. Lockwood, D. Drake, S.M. Grodsky, and T. Pover. 2016. Conservation of American oystercatchers Haematopus palliatus within an urbanized barrier island complex. Wader Study 123: 1–11.
Vitale, N. 2019. Habitat change, predators, and disturbance: Factors influencing productivity of American oystercatchers (Haematopus palliatus) nesting in Florida’s Big Bend. M.Sc. Thesis, University of Florida, Gainesville, Florida.
Webster, P.J., G.J. Holland, J.A. Curry, and H.-R. Chang. 2005. Changes in tropical cyclone number, duration, and intensity in a warming environment. Science 309: 1844–1846.
Acknowledgments
We thank the US Fish and Wildlife Service, American Oystercatcher Working Group, Florida Department of Environmental Protection, The Cross Florida Greenway State Recreation and Conservation Area, Lower Suwannee and Cedar Keys National Wildlife Refuge, Florida Shorebird Alliance and the Nature Coast Shorebird Partnership. Special thanks to both Bill Pine and John Jaeger for their guidance in our data analyses and manuscript review, and Joe Marchionno, Andrew Townsend, Julia Magill plus many others who contributed to the success of this project. We thank Patrick Jodice and Andrew Cox for their reviews of this manuscript. Any use of trade, firm, or product names is for descriptive purposes only and does not imply endorsement by the US Government.
Funding
This work was funded by the Florida Fish and Wildlife Conservation Commission and the UF/IFAS Nature Coast Biological Station, with logistical support from the US Geological Survey, Florida Cooperative Fish and Wildlife Research Unit. This study was performed under the auspices of the University of Florida IACUC study #201709798.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Any use of trade, firm, or product names is for descriptive purposes only and does not imply endorsement by the U.S. Government.
Additional information
Communicated by James Lovvorn
Rights and permissions
About this article

Cite this article
Vitale, N., Brush, J. & Powell, A. Loss of Coastal Islands Along Florida’s Big Bend Region: Implications for Breeding American Oystercatchers. Estuaries and Coasts 44, 1173–1182 (2021). https://doi.org/10.1007/s12237-020-00811-3
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s12237-020-00811-3