
Abstract
Cedar Bayou, a natural tidal inlet, was recently dredged to allow for direct water exchange between the Gulf of Mexico and Mesquite Bay, TX, USA. We quantified changes in densities of juvenile nekton (fish, shrimps, and crabs) and community structure in Mesquite Bay after Cedar Bayou was reopened by collecting samples at both control and impact sites using an epibenthic sled 1 year before (October 2013–April 2014) and after (October 2014–April 2015) opening. Significantly higher densities of total nekton were observed at the impact sites after opening using a before-after control-impact design. Red Drum (Sciaenops ocellatus), Atlantic Croaker (Micropogonias undulatus), post-larval penaeid shrimps (Farfantepenaeus aztecus, F. duorarum, and Litopenaeus setiferus), and Blue Crabs (Callinectes sapidus) were significantly more abundant at impact sites after Cedar Bayou was opened. Multivariate analysis showed a significant change in impact site communities after opening and was driven by an increased presence of estuarine-dependent species. Overall, this study demonstrates that opening tidal inlets, such as Cedar Bayou, and reconnecting Mesquite Bay to the Gulf of Mexico increased the presence of numerous estuarine-dependent species, many of which were not present or occurred at very low densities prior to reopening. Thus, reestablishing the historical connectivity between a productive estuary and the open Gulf of Mexico via Cedar Bayou should reinstitute natural nekton recruitment processes important to the Aransas, Mesquite, and San Antonio Bay regions.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Access to estuaries through tidal inlets is essential to the life history of estuarine-dependent organisms. Approximately 75 % of commercially and recreationally valuable species in the Gulf of Mexico are dependent on access to estuaries for at least one critical stage of development (Chambers 1991). Inlets play an important role in providing juvenile estuarine-dependent nekton access to estuarine nursery habitats such as seagrass meadows (King 1971; Reese et al. 2008; Heck et al. 2003), salt marshes (Weinstein 1979; Baltz et al. 1993; Jenkins and Black 1994; Jenkins et al. 1997; Minello 1999; Stunz and Minello 2001; Brown et al. 2004), and oyster reefs (Nevins et al. 2013). These habitats provide juvenile nekton with protection from predation and increased food resources, leading to increased growth rates and recruitment into larger size classes (Rooker et al. 1998a, b; Stunz et al. 2002a; Neahr et al. 2010). Thus, understanding how tidal inlets influence recruitment dynamics is critical to the management of numerous Gulf of Mexico fisheries, such as Red Drum (Sciaenops ocellatus), Blue Crabs (Callinectes sapidus), penaeid shrimps (Farfantepenaeus aztecus, F. duorarum, and Litopenaeus setiferus) (NMFS 2014), and particularly Southern Flounder (Paralichthys lethostigma) as their population has been in decline in Texas for 25 years (TPWD 2003; Nañez-James et al. 2009).
Cedar Bayou is a natural ephemeral tidal inlet that separates Matagorda Island from San Jose Island and historically allowed water exchange between Mesquite Bay and the Gulf of Mexico, Texas, USA (Kraus 2007; Shepsis and Carter 2007). Despite large gaps in quantitative information regarding the inlet’s dynamics throughout time, Cedar Bayou was generally open and flowing prior to 1950 (Ward 2010). The mouth of the inlet was closed in 1979 to prevent contaminants from the Ixtoc oil spill from reaching Texas bays and the mainland (Ward 2010). Despite subsequent coastal storms and coastal engineering-induced changes, the inlet has remained closed or in a reduced state for much of the last 30 years. There was immense interest by the scientific, birding, and fishing communities in how the Mesquite Bay region would respond to the reopening of Cedar Bayou.
Due to Cedar Bayou’s diminished cross-sectional area (≤9.3 m2 from 1980 to 2010), its contemporary role as a major avenue for juvenile nekton recruitment was unclear (Ward 2010), and few studies have examined the effects of opening tidal inlets on estuarine nekton. However, results from the nearby Laguna Madre bay system found that reopening Packery Channel, an isolated tidal inlet near Corpus Christi, TX, resulted in significantly increased densities of many estuarine-dependent species including juvenile Red Drum, Atlantic Croaker (Micropogonias undulatus), penaeid shrimps, and Blue Crabs (Bushon 2006; Reese et al. 2008). Therefore, we hypothesized that even though Cedar Bayou is a relatively small inlet, its reopening would play a substantial role in establishing biological connectivity between the Gulf of Mexico and Mesquite Bay given its great distance from other open flowing inlets —Aransas Pass is located approximately 32 km to the southwest, and Cavallo Pass is located approximately 55 km to the northeast.
In addition to Cedar Bayou’s role in delivering economically important nekton to nursery habitats, the inlet serves as a feeding ground for endangered Whooping Cranes (Grus americana) that overwinter at the Aransas National Wildlife Refuge (Gil-Weir et al. 2012). These cranes rely on Blue Crabs from Cedar Bayou for 41 % of their winter diet (Westwood and Chavez-Ramirez 2005). Periods of lower Blue Crab abundance are correlated to increased winter mortality for Whooping Cranes (Pugesek et al. 2008). While the decline in Blue Crab numbers is driven by a combination of factors, their low abundance is almost certainly impacted by lack of inlet access. Given that reopening a nearby inlet increased juvenile density of Blue Crabs (Reese et al. 2008), reopening Cedar Bayou may play a key role in increasing the food supply for Whooping Cranes overwintering in the region.
Cedar Bayou, when open, is the only connection through Matagorda Island in the northern Aransas Bay and southern San Antonio Bay region that allows estuarine-dependent nekton access to nursery habitats from spawning grounds in the Gulf of Mexico. The restoration of flow to Cedar Bayou provides a rare opportunity to elucidate the direct impacts of tidal inlets on juvenile estuarine-dependent nekton. The overall goals of this project were to (1) determine whether seasonal relative abundance, density, and size of juvenile nekton (fish, shrimps, and crab) species changed after Cedar Bayou was dredged and reopened and (2) assess nekton community changes in Mesquite Bay before and after opening.
Methods
Study Site
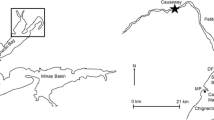
Mesquite Bay is the southernmost component of the larger San Antonio Bay estuary complex (Armstrong 1987; Britton and Morton 1989). It is bordered to the north by San Antonio Bay and to the southwest by the Aransas Bay complex (Fig. 1). Given its position on the landward side of the Matagorda and San Jose barrier islands, Mesquite Bay is isolated from the Gulf of Mexico when Cedar Bayou is closed. Cedar Bayou is approximately 4.8 km in length, and prior to dredging the northern two thirds, Cedar Bayou measured approximately 180 m wide with a mean depth of 2 m below mean sea level. The lower third of the inlet is morphologically dynamic and subject to constant changes in width and depth (Bermudez et al. 2005) with the ebb and flow of tides. Ward’s (2010) comprehensive reconstruction of Cedar Bayou’s history shows that marginal flow was intermittently present up to the 2005 to 2007 time period; however, the inlet was permanently closed from 2007 until the reopening in September 2014.

The Cedar Bayou study area. Both the control sites (triangles) established near the Aransas inlet and impact sites (circles) located near Cedar Bayou were located in seagrass meadows (Halodule wrightii)
Study Design and Sample Site Delineation
This study assessed the effect of reopening Cedar Bayou on abundances, densities, and sizes of nekton using a before-after control-impact (BACI) experimental design (Smith 2002). There are numerous variations of before-after designs (Eberhardt 1976; Green 1979; Schwarz 2012), and we used a BACI design by Stewart-Oaten et al. (1986) that was first used to determine impacts of reopening tidal inlets by Reese et al. (2008) and Milbrandt et al. (2012). Seagreass meadows are the predominant habitat type used by recruiting nekton in this region (Stunz et al. 2002a, b). Four impact sites were established in locations within shallow seagrass meadows (Halodule wrightii) near Cedar Bayou that estuarine-dependent nekton would encounter upon ingressing. Four control sampling sites were also selected in seagrass meadows (H. wrightii) adjacent to Aransas Pass, a functioning tidal inlet. The distance of the control sites from Cedar Bayou (approximately 32 km) was far enough away to be unaltered by its reopening based on previous studies (Bushon 2006). Diversity and abundance of nekton at control sites were considered representative of an estuarine system receiving large supplies of newly recruiting nekton from an open and flowing tidal inlet (Nañez-James et al. 2009; Neahr et al. 2010).
Sampling Procedure
Nekton samples were collected during daylight hours using an epibenthic sled. This device has been well-established in the literature as an efficient gear for sampling small nekton of the size we were targeting in seagrasses and other estuarine habitat types (Stunz et al. 2002b; Reese et al. 2008; Neahr et al. 2010). Briefly, it is composed of a metal frame 0.6 m wide by 0.75 m high, which supports a 1-mm-mesh conical plankton net mounted to skids. Each tow consisted of pulling the sled 16.6 m covering 10 m2 of the seagrass bed. Samples from each individual tow were rough sorted in the field and preserved in 10 % buffered formalin (Reese et al. 2008).
Two sampling events were conducted in each recruitment season (fall, winter, spring) for 2 years (1 year each, before and after opening). Three independent epibenthic sled tows were taken at each of the eight sites (four control and four impact) during each sampling event totaling 288 samples over the entire study period. At each sample site, water temperature (°C), dissolved oxygen (mg L−1), and salinity were recorded using a Hydrolab MS 5 sonde. In the laboratory, fishes and crustaceans in each sample were sorted, counted, identified to the lowest possible taxon, and measured to the nearest 0.1 mm. Fishes were measured using standard length (SL), shrimps were measured using total length (TL) between the tip of rostrum and the telson, and crab species were measured using carapace width (CW). If more than 22 individuals of the same species were collected in a single tow, the largest, smallest, and 20 randomly selected individuals were measured. We assumed that these measurements of randomly sampled individuals were representative of the entire size distribution in the tow. Once a sample was processed, organisms were preserved in 70 % ethanol for long-term storage.
Statistical Analysis
Nekton General Comparisons
Mean density (m−2), mean size (mm), relative abundance (RA %), and change in relative abundance (Δ RA %) estimations were calculated for each species during each recruitment season at the control and impact sites following Reese et al. (2008). Each mean density was calculated from a total of 24 samples collected each season at the control and impact sites. Mean sizes were calculated from the number of individuals of a species measured during each season before and after opening. The RA (%) was calculated by dividing the number of individuals of a species collected by the total number of fishes or crustaceans within a particular season and multiplied by 100. Before opening, RA (%) was subtracted from after opening RA (%) to calculate Δ RA (%). A negative change in relative abundance indicates a decline in relative abundance, while a positive number indicates an increase in relative abundance.
Primary Recruitment Seasons
Recruitment of nekton into estuaries is highly variable; thus, data used to test for significant differences in this BACI design were restricted to peak recruitment seasons for each species group or individual species (Reese et al. 2008). Total nekton, total fish, total crustaceans, and resident shrimp densities were tested over all sample seasons. Resident shrimp density was calculated by summing the densities of Arrow Shrimp (Tozeuma carolinense), Hippolytidae spp., and grass shrimps (Palaemonetes spp.), which comprised over 85 % of the total crustaceans collected during the course of this study. Mean densities and sizes of Red Drum were determined using fall samples only (Holt et al. 1983; Rooker and Holt 1997; Rooker et al. 1998a; Stunz et al. 2002b; Reese et al. 2008), while Southern Flounder and Atlantic Croaker were determined using winter samples only (Haven 1957; Hansen 1969; Rooker et al. 1998b; Searcy et al. 2007; Nañez-James et al. 2009). Mean densities and sizes of post-larval penaeid shrimps and Blue Crabs were calculated by combining fall, winter, and spring samples given that these taxa have complex life histories, disperse widely, and spawn year-around (Pile et al. 1996; Blackmon and Eggleston 2001; Reese et al. 2008).
Nekton Density and Size Comparisons
Density and size data were log (x + 1) transformed to ensure homogeneity of variance and normality of residuals (Zuur et al. 2007; Reese et al. 2008). Transformed data were analyzed with ANOVA in a BACI design using the linear mixed effects (.lme) function in R 3.1.3 (Pinheiro et al. 2015) to identify changes in density and size due to the opening of Cedar Bayou. The main effects model was conducted for every group (e.g., total organisms, fish) and species of interest with a two-way nested ANOVA that tested for a significant interaction between the before-after (BA) and control-impact (CI) main factors using type III sum of squares (α = 0.05). Site was treated as a random factor nested in the CI factor (BA*site(CI)). The .lme function allows for random, nested factors and uses restricted maximum likelihood (REML) to generate a set of contrasts calculated from original data. The REML technique is capable of producing unbiased estimates of variance parameters while ensuring nuisance parameters have no effect. Density data were balanced as a result of experimental design. Since size data were unbalanced given that the number of individuals of a species varied by tow, all ANOVAs used to test these data utilized Helmert contrasts.
To further test for differences within impact locations, BA and CI factors were combined using the “paste” function in R 3.1.3 to create a single factor with four levels: before control (BC), after control (AC), before impact (BI), and after impact (AI). Site remained nested within the new combined BA/CI factors. Then, one-way “post-hoc” ANOVAs (α = 0.05) were used to test for differences within impact and control locations before and after opening for all groups and individual species of interest (BC*AC and BI*AI).
Community Analysis
Multivariate analyses were implemented to test for differences in communities at impact and control locations over the course of the study using statistical procedures from PRIMER (v.7 PERMANOVA+). Mean densities for each species were calculated by sampling event (date) for control and impact sites. All data were fourth root transformed before analysis to reduce the importance of more abundant species and to allow for changes in rarer species to be statistically discernable (Clarke and Green 1988; Reese et al. 2008). A Bray-Curtis resemblance matrix was generated from mean densities for each sampling event for both control and impact sites before and after opening. A permutational multivariate analysis of variance (PERMANOVA) was then calculated using type III sum of squares. This method is widely used to detect differences in locations of multivariate groups (Anderson and Walsh 2013; Ajemian et al. 2015). PERMANOVAs test the null hypothesis that the centroids of the groups are equivalent for all groups as defined in the space of the chosen resemblance measure. Pseudo-F ratios are also generated by partitioning distance matrices for multivariate data and conducting permutations to make the data distribution-free. They are robust to heterogeneity when used in balanced designs (Anderson and Walsh 2013). Pair-wise tests were also performed on significant factors to determine whether control and impact site communities changed between the before and after opening years following Ajemian et al. (2015).
To identify the species most responsible for driving disparity in assemblages, separate Bray-Curtis resemblance matrices were constructed for the impact sites both before and after opening. These matrices were tested for differences using the RELATE package which performs a rank correlation, compares the results to randomly permuted samples, and tests the null hypothesis that no correlation exists between the similarity matrices (Clarke and Gorley 2006; Reese et al. 2008). Impact and control site resemblance matrices were also used to generate a nonmetric multidimensional scaling (nMDS) ordination, which allowed for visual comparison of control site and impact site communities throughout time (Clarke and Warwick 2001; Reese et al. 2008; Ajemian et al. 2015). Bray-Curtis cluster groups were superimposed on the impact site nMDS ordination to facilitate interpretation (Clarke and Warwick 2001; Reese et al. 2008). Finally, a two-way crossed similarity of percentages (SIMPER) analysis was conducted to determine which overall species densities were the most dissimilar between before and after opening samples. All seasons were combined for these comparisons as communities change seasonally (Reese et al. 2008).
Results
Physical Variables
Sample site water temperatures (°C), dissolved oxygen levels (mg L−1), and salinities changed seasonally (Table 1). Dissolved oxygen ranged from 5.9 to 10.4 mg L−1, water temperatures from 14.6 to 24.9 °C, and salinity from 27.7 to 34.4. Differences were minimal within season and likely not biologically significant.
Overall Nekton Relative Abundance and Species Characterization
A total of 126,812 organisms were collected during the course of this study, representing 31 fishes and 7 crustacean species. Any individuals that could not be identified to species were grouped into the lowest possible taxon (e.g., Clupeiformes, Gobiidae, and Gobiosoma for fishes and Xanthidae, Hippolytidae, Penaeidae, and Palaemonetes for crustaceans). As is characteristic of estuarine nekton recruitment patterns, some differences were found among seasons both before and after opening in control sites; however, control sites remained largely the same within each season after opening Cedar Bayou.
The influence of opening Cedar Bayou was most apparent at impact sites. In general, estuarine resident species were abundant both before and after opening. For example, the most abundant fishes during fall before opening were Gobiosoma spp. and Syngnathus spp., and they remained the most abundant after opening. Alternatively, we observed within-season increases in nekton mean density (m−2) and RA (%) for numerous estuarine-dependent species, and many of these species were among the most abundant organisms after opening (Table 2). Red Drum increased from 0 % at the impact sites in the fall before opening to 14.8 % after opening, along with post-larval penaeids which increased in the fall after opening (∆ RA 11.4 %). After opening, winter samples showed increased abundances of Atlantic Croaker (∆ RA 18.8 %) and Blue Crabs (∆ RA 36.1 %). Spring after opening impact samples revealed that estuarine-dependent Pinfish (Lagodon rhomboides) were among the most abundant fishes (RA 16.1 %), and crustacean abundances were led by post-larval penaeids (∆ RA 25.2 %) and adult Farfantepenaeus spp. (∆ RA 13.2 %).
Nekton Density and Size Comparisons
The control sites near the long-established Aransas inlet remained largely unchanged throughout the study, whereas substantial changes were observed in density of nekton at impact sites after Cedar Bayou was opened. Overall, impact sites were characterized by significant increases in mean density of total organisms, total fish, total crustaceans, and resident shrimp following reopening Cedar Bayou (Fig. 2, Tables 3 and 4). Results for total organism density showed a significant BA × CI interaction (Table 3), and a one-way post-hoc ANOVA demonstrated the interaction was due to a significantly higher density of total organisms at impact sites after opening, while there was no significant difference at control sites after opening (Fig. 2, Tables 3 and 4).

Overall mean densities (m−2), and standard error of total organisms (total), fish, crustaceans, and resident shrimp (RS) species in control and impact locations over all seasons before and after opening. A main-effects two-way nested ANOVA, (BA*site(CI)), was used to test each group for significance, *p < 0.05, ***p < 0.001. See Table 4 for sample size
Densities of total fish and crustaceans also changed throughout the study, particularly at impact locations. There was a significant BA × CI interaction (Table 3) of total fish densities, and post hoc ANOVAs showed an increase of fish at impact locations after opening. No significant differences in total fish densities were detected at control sites after opening (Fig. 2, Tables 3 and 4). Density of total crustaceans also increased after opening. The main effects ANOVA for total crustacean density showed a highly significant BA × CI interaction (Table 3), and post hoc tests revealed a significant density increase at impact and control sites after opening (Fig. 2, Tables 3 and 4). The resident shrimp main effects ANOVA did not detect a significant BA × CI interaction; however, post hoc ANOVAs showed significant increases at impact and control sites (Fig. 2, Tables 3 and 4). Although there was a significant result for resident shrimps at control sites, this should be interpreted with caution as it is most likely due to the log transformation of outliers, and likely not truly significant.
Changes in mean density and size were assessed for estuarine-dependent species during their peak recruitment to examine species-specific effects of the Cedar Bayou opening (Fig. 3, Tables 4 and 5). The main effects ANOVA for density of Red Drum detected a significant BA × CI interaction, and post hoc analyses indicated that their density increased significantly at impact sites (Fig. 3, Tables 4 and 5). No significant change in Red Drum density occurred at the control sites between the before and after opening years. We were unable to perform statistical analysis for the size of Red Drum because no individuals were captured at the impact sites before opening. After opening, Red Drum mean size was 9.7 mm ± 0.44 (Fig. 3, Table 4), and mean sizes remained similar at control sites between years (Fig. 3 and Table 4). We did not find a significant BA × CI interaction in density of Southern Flounder (Table 5), and there was no significant change in density at either the impact or control locations. The mean size main effects ANOVA did not show a significant BA × CI interaction (Table 5), and we observed no significant changes in mean size between before and after opening at impact or control locations (Fig. 3, Tables 4 and 5). Densities of Atlantic Croaker had a significant BA × CI interaction (Table 5), and we found a significant decrease in density at the control sites while impact locations showed increased density. Atlantic Croaker mean size did not show a significant interaction; post hoc results showed no change at impact locations (Fig. 3; Tables 4 and 5). We found a significant change in mean size at control sites (Fig. 3, Tables 4 and 5), but few individuals (n = 1) were captured post-opening at control sites.

Mean densities (a) and mean sizes (b) of selected fishes and crustacean species during their season(s) of peak recruitment at impact sites before and after opening the inlet. Mean densities and sizes along with standard errors were calculated from all individuals sampled within the specified recruitment season. Post hoc ANOVA (BI*AI) results are indicated above the bars; **p < 0.01, ***p < 0.001. See Table 4 for sample size
Similar to estuarine-dependent fishes, the density of post-larval penaeid shrimps changed significantly (BA × CI interaction) with the opening of Cedar Bayou (Table 3). Post hoc ANOVAs show that significantly more post-larval penaeids were found at the impact sites after opening while no significant change occurred at the control locations. We also found a significant size interaction where impact sites experienced no change (Fig. 3, Tables 4 and 5), but at control sites penaeid shrimps were significantly larger after opening. Blue Crab densities were significantly different (BA × CI interaction) after opening Cedar Bayou (Table 3). There were significantly more Blue Crabs at impact sites while densities at control sites remained similar. Blue Crab mean sizes followed a similar pattern with a significant interaction (Table 3), decreased sizes at the impact sites (Fig. 3, Tables 4 and 5), but no significant change in mean size at the control sites.
Community Analysis
Multivariate analyses indicated that the composition of the nekton community changed at impact sites following the opening of Cedar Bayou. PERMANOVA results show that a small interaction may have occurred between the BA and CI factors at a community level (998 permutations, df = 1, pseudo-F = 2.19, p = 0.070), although it was not significant. However, pair-wise tests demonstrated that while the control site community did not change before and after opening (393 permutations, df = 10, t = 0.93, p = 0.460), the impact site community did change (418 permutations, df = 10, t = 2.28, p = 0.003). Differences between the control and impact site communities existed before (411 permutations, df = 10, t = 3.17, p = 0.002) and after opening (402 permutations, df = 10, t = 2.03, p = 0.003). The nMDS ordination for the control and impact sites show that the after opening impact site community shifted towards the control site community (Fig. 4a).

Nonmetric multidimensional scaling (nMDS) ordinations for (a) control and impact sites before and after opening, and (b) impact sites only with Bray-Curtis cluster analysis (63 % similarity) superimposed. Densities were averaged for control and impact locations by date for a total of 24 samples
Given that changes to the community at impact sites were so great, additional analyses were performed to determine more fine-scale community changes. The RELATE routine verified that no correlation existed between the abundance matrices for before- and after-opening impact communities (R = 0.281, p = 0.100). Differences between before and after opening nekton densities at the impact sites were also demonstrated with Bray-Curtis cluster analysis and nMDS ordination. The Bray-Curtis cluster revealed four distinct clusters at the 63 % similarity level, represented by two before-opening groups and two after-opening groups (Fig. 4b). The two-way crossed SIMPER analysis showed that increased densities of post-larval penaeid shrimps, Hippolytidae spp., and Blue Crabs at impact sites after opening (Table 6) greatly contributed to the dissimilarity between before- and after-opening communities across all seasons. These same organisms also contributed greatly to within-group similarity (Table 6). Overall, increases in other estuarine-dependent species including Red Drum, Pinfish, and Atlantic Croaker contributed to within-group similarity at the impact sites after opening Cedar Bayou (Table 6).
Discussion
Reopening tidal inlets is often a costly and time-consuming process. Therefore, it is important to document the ecological effects of inlet restorations not only to justify the high cost but also to validate the project as a viable management and conservation tool. The goal of this study was to determine whether reopening a natural tidal inlet would increase recruitment of nekton into estuarine habitats. Dramatic changes to total nekton, fish, and crustacean densities at impact sites after opening indicated that restoration of flow at Cedar Bayou significantly impacted the community as a whole. In addition, we observed elevated recruitment and density for numerous estuarine-dependent species including Red Drum, Blue Crabs, Atlantic Croaker, and post-larval penaeid shrimps. The increases in estuarine-dependent species were large enough to induce an assemblage shift at impact sites after opening, and these results demonstrate the positive effect that tidal inlets can have on nekton density and community.
Physical Parameters
Overall, physical water variables remained similar between control and impact sites, as well as pre- versus post-opening. The reopening of tidal inlets can cause changes in physical water parameters at both a local and regional level (Reese et al. 2008; Milbrandt et al. 2012), and these changes have the potential to impact nekton community structure (Hoff and Ibara 1977; Reese et al. 2008). However, changes in water quality variables observed here were not necessarily attributable to opening Cedar Bayou, and fluctuations were within a range that would be expected; thus, we do not suspect that water quality influenced the nekton recruitment patterns observed in this study.
Nekton Abundance and Density
The effect of opening Cedar Bayou was best characterized by the numerous within-season changes to nekton abundance at impact sites after opening, and the changes we found for numerous species provided a broad perspective of the role Cedar Bayou plays in shaping Mesquite Bay’s ecology. For example, nekton abundance at control sites remained largely unchanged after opening, and most of the abundant species (e.g., Darter Gobies) are common in healthy Texas estuaries (Nelson 1992; Reese et al. 2008). The small abundance changes observed are most likely attributable to normal annual population fluctuations. In contrast, estuarine-dependent species (e.g., Blue Crabs) which were not common at the impact sites before opening became very abundant after opening. The increase in abundance of estuarine-dependent fishes and crustaceans at the impact sites suggests that opening Cedar Bayou allowed estuarine-dependent species to more effectively reach nursery habitats in Mesquite Bay from their offshore spawning grounds. These patterns reinforce the view that inlets play a large role in determining nekton populations and assemblages along the Texas coast.
Additionally, increases in the density of total organisms at the impact sites after Cedar Bayou was opened further suggests there was a large influence of this newly reopened inlet. In contrast, few changes were seen at the control sites, confirming that a greater influx of estuarine-dependent species to impact locations in Mesquite Bay was the cause of the changes we observed. Further evidence from the increase in fish density shows that estuarine-dependent nekton, particularly fishes, are ingressing through Cedar Bayou and settling in nearby seagrass beds (Bell and Westoby 1986). Reese et al. (2008) demonstrated similar patterns for fish densities; however, few other studies have shown a change in fish density after reopening a tidal inlet. For example, inlet restoration studies on Rollover Pass, TX (Reid 1957), and Blind Pass, FL (Milbrandt et al. 2012), reported no significant change in fish density. The primary reason previous studies did not observe a change in total fish density is likely attributable to nekton being able to easily access these areas through other nearby inlets. Thus, the impact of opening geographically isolated tidal inlets such as Cedar Bayou and Packery Channel may be more pronounced as estuarine-dependent nekton are able to recruit to nursery habitats that were previously inaccessible (Reese et al. 2008).
The large increase in density of crustaceans at both control and impact sites after opening was most likely driven by the high abundance of resident shrimps at all sites. Any changes by other crustacean species were masked because they occurred in much lower abundance. The small differences in resident shrimp densities at the control sites were most likely due to environmental variation (Fogarty et al. 1991), while densities at impact sites may have been more directly caused by Cedar Bayou. These resident shrimp species are present in high densities in seagrass beds throughout the year and exhibit peak abundances during different seasons based on environmental conditions. Many of these species, particularly Palaemonetes spp., provide a critical prey base for a variety of marine organisms including Red Drum and Spotted Seatrout Cynoscion nebulosus (Wood 1967; Morgan 1980). This increase in prey resources will be important in sustaining this changed community given the increase of many fish species.
The impact of reopening Cedar Bayou was most clearly demonstrated by two Sciaenid species, Red Drum and Atlantic Croaker. Red Drum did not occur at impact sites before opening, but after opening we found high densities that resembled numbers at control sites. This change in abundance suggests that Red Drum were not able to reach these areas before Cedar Bayou was dredged. Similar findings by Reese et al. (2008) suggest that juvenile Red Drum have difficulty dispersing and accessing nursery habitat farther from tidal inlets. Moreover, other studies have shown that even when habitat is available, juvenile Red Drum have limited dispersal ability and their densities substantially decrease with distance from a tidal inlet (Bushon 2006). Even though densities of Atlantic Croaker decreased at control sites over the course of our study, most likely because of annual variation in recruitment (Fogarty et al. 1991), their densities were higher at impact sites during the peak winter recruitment season. Our data suggest that this increase in density at the impact sites was larger than normal variation and is a function of juveniles having access to Mesquite Bay via Cedar Bayou. Thus, reopening Cedar Bayou allowed Red Drum and Atlantic Croaker to access the extensive seagrass beds of Mesquite Bay, which now provides critical nursery habitat for these economically important species.
Newly settled and juvenile Southern Flounder generally occur at very low densities throughout Texas estuaries (Stokes 1977; Nañez-James et al. 2009; Froeschke et al. 2013), so our result of no change at impact sites is not surprising. Although densities of juvenile Southern Flounder are highest in vegetated areas nearest to open inlets (Nañez-James et al. 2009), our results are consistent with the findings of Bushon (2006), Reese et al. (2008), and Milbrandt et al. (2012), who also observed no change in Southern Flounder populations following the reopening of tidal inlets. Nañez-James et al. (2009) showed that young flounder can reach isolated areas even when nearby inlets are closed. Their naturally low abundance, even near open tidal inlets, may have prevented the detection of any significant impacts indicating that access to tidal inlets may not be the most limiting factor on Southern Flounder recruitment. Southern Flounder populations have been in dramatic decline for the past 25 years (Stunz et al. 2000) and are showing very slow recovery. More research is needed to elucidate Southern Flounder recruitment dynamics and examine in detail the role of tidal inlets in the recruitment process.
The increase in densities of penaeid shrimps and Blue Crabs at impact sites after opening suggests that these species were able to access nursery habitats in Mesquite Bay from the Gulf of Mexico via Cedar Bayou. Blue Crabs in particular seemed able to disperse and settle more effectively with Cedar Bayou open (Heck and Thoman 1984; Patillo et al. 1997; Heck et al. 2001; Bushon 2006; Reese et al. 2008). Penaeid shrimps and Blue Crabs are extremely important to the Texas ecosystem and economy; the Texas shrimping industry alone generates US$159 million annually (NMFS 2014). Moreover, penaeid shrimps and Blue Crabs are key prey items for Spotted Seatrout, Red Drum, Atlantic Croaker, and Southern Flounder (Minello and Zimmerman 1983; Minello et al. 1989; Scharf and Schlicht 2000) and thus represent the forage base supporting the billion dollar recreational sportfish fishery within the state (NMFS 2014). In addition to its economic importance as a fishery, Blue Crabs are a major dietary component of the endangered Whooping Crane. This species’ winter survival directly correlates to Blue Crab abundance (Pugesek et al. 2008), and increased Blue Crab recruitment into Mesquite Bay’s seagrass nurseries has the potential to translate into higher abundance, which could provide an important forage base for the sensitive Whooping Crane population wintering near Cedar Bayou.
The size of most species remained similar at impact and control sites after flow was restored to Cedar Bayou. Other studies of reopening tidal inlets found that the mean size of estuarine-dependent nekton may decrease at impact sites once the inlet is reopened (Bushon 2006; Reese et al. 2008). This size decrease indicates recruitment via the new inlet rather than an extended migration from another inlet farther away (allowing for growth as they are moving). The only two species that showed size differences post-opening were Blue Crabs and penaeid shrimps. Blue Crabs were smaller at the impact sites after opening, while post-larval penaeid shrimps were only slightly larger at control sites, and the biological significance of this change is questionable. The lack of size differences in fishes after opening observed in our study is most likely attributed to the high growth rate of young recruits (Rooker and Holt 1997) and the variability in exact recruitment dates (Rooker et al. 1998b) and also because many species were not available pre-opening to perform size-based assessment. Thus, size may not always be a good indicator of the location of recruitment especially when numerous cohorts of recruits occur simultaneously in the same habitat. Changes in density patterns were a better indicator of recruitment potential via the new tidal inlet and indicated a substantial increase in juvenile nekton after the inlet was opened.
Community Analysis
Flowing tidal inlets can determine nekton community structure in estuarine habitats as was highlighted by the changes we observed in density and abundance of species at impact sites. We found strong seasonal effects on impact site communities indicated by groupings of winter and spring samples and unique fall samples. Seasonal pulses of juvenile estuarine-dependent nekton can cause community to shift and change, which is most likely the cause of changes both seasonally and overall (Reese et al. 2008). Moreover, because most of the species responsible for the shift in community composition are estuarine-dependent (e.g., Blue Crabs and penaeid shrimps) and rely on open inlets for access to seagrass nursery habitat, reopening Cedar Bayou and allowing the ingress of new estuarine-dependent nekton in Mesquite Bay influenced the seagrass communities.
Conclusions
The opening of Cedar Bayou caused positive changes in nekton density and community structure in Mesquite Bay’s previously inaccessible seagrass habitats by providing a means of ingress for estuarine-dependent species. Our findings also show the potential for increasing species abundance, which may translate into increasing population productivity as a direct result of reopening Cedar Bayou. It may take years and additional studies to understand the full effect of restoring Cedar Bayou; nonetheless, the inlet’s positive influence on the Mesquite Bay ecosystem was observed almost immediately after it was reopened, and we anticipate these trends to continue.
References
Ajemian, M.J., J.J. Wetz, B. Shipley-Lozano, J.D. Shively, and G.W. Stunz. 2015. An analysis of artificial reef fishes community structure along the northwestern Gulf of Mexico shelf: potential impacts of “Rigs-to-Reefs” programs. PLoS ONE 10(5): e0126354.
Anderson, M.J., and D.C.I. Walsh. 2013. PERMANOVA, ANOSIM, and the Mantel test in the face of heterogeneous dispersions: what null hypothesis are you testing? Ecological Monographs 83: 557–574.
Armstrong, N.E. 1987. The ecology of open-bay bottoms of Texas: a community profile. U.S. Fishes and Wildlife Service Biological Report 85(7.12) 104 pp.
Baltz, D.M., C. Rakocinski, and J.W. Fleeger. 1993. Microhabitat use by marsh-edge fishes in a Louisiana estuary. Environmental Biology of Fishes 36: 109–126.
Bell, J.D., and M. Westoby. 1986. Variation in seagrass height and density over a wide spatial scale: effects on common fishes and decapods. Journal of Experimental Marine Biology and Ecology 104: 275–295.
Bermudez, H.E., V. Shepsis, and J. Carter. 2005. Restoration of Cedar Bayou and Vinson Slough: Phase 1. Prepared for: Save Cedar Bayou, Inc. by Coast and Harbor Engineering, Austin, TX.
Blackmon, D.C., and D.B. Eggleston. 2001. Factors influencing planktonic, post-settlement dispersal of early juvenile blue crab (Callinectes sapidus Rathbun). Journal of Experimental Marine Biology and Ecology 257: 183–203.
Britton, J.C., and B. Morton. 1989. Shore ecology of the Gulf of Mexico, 3rd ed. Austin: University of Texas Press.
Brown, C.A., S.A. Holt, G.A. Jackson, G.A. Brooks, and G.J. Holt. 2004. Simulating larval supply to estuarine nursery areas: how important are physical processes to the supply of larvae to the Aransas Pass Inlet? Fisheries Oceanography 13: 181–196.
Bushon, A.M. 2006. Recruitment, spatial distribution, and fine-scale movement patterns of estuarine dependent species through tidal inlets in Texas (Master’s Thesis). Texas A&M University Corpus-Christi, Department of Life Sciences.
Chambers, J.R. 1991. Page 45–51. Coastal degradation and fishes population losses. In Stemming the tide of coastal fishes habitat loss, ed. RH Stroud. Savannah: National Coalition for Marine Conservation.
Clarke, K.R., and R.N. Gorley. 2006. PRIMER v6: user manual and tutorial. Plymouth: PRIMER-E.
Clarke, K.R., and R.H. Green. 1988. Statistical design and analysis for a “biological effects” study. Marine Ecology Progress Series 46: 213–226.
Clarke, K.R., and R.M. Warwick. 2001. Change in marine communities: an approach to statistical analysis and interpretation, 2nd ed. Plymouth: PRIMER-E.
Eberhardt, L.L. 1976. Quantitative ecology and impact assessment. Journal of Environmental Management 4: 27–70.
Fogarty, M.J., M.P. Sissenwine, and E.B. Cohen. 1991. Recruitment variability and the dynamics of exploited marine populations. Trends in Ecology and Evolution 6: 241–246.
Froeschke, B.F., P. Tissot, G.W. Stunz, and J.T. Froeschke. 2013. Spatiotemporal predictive models for juvenile southern flounder in Texas estuaries. North American Journal of Fisheries Management 33: 817–828.
Gil-Weir, K.C., W.E. Grant, R.D. Slack, H.H. Wang, and M. Fujiwara. 2012. Demography and population trends of Whooping Cranes. Journal of Field Ornithology 83: 1–10.
Green, R.H. 1979. Sampling design and statistical methods for environmental biologists. Chichester: John Wiley and Sons, Ltd., Printer.Gundlach, E.R., K.J. Finkelstein, and J.L. Sadd. 1981. Impact and persistence of Ixtoc I oil on the South Texas coast. Proceedings of 1981 Oil Spill Conference. Washington, D.C.: American Petroleum Institute Publication Number 4334:477–485.
Hansen, D.J. 1969. Food, growth, migration, reproduction, and abundance of pinfish, Lagodon rhomboides, and Atlantic Croaker, Micropogonias undulatus, near Pensacola, Florida 1963–65. Fishery Bulletin 68: 135–146.
Haven, D.S. 1957. Distribution, growth, and availability of juvenile croaker, M. undulatus, in Virginia. Ecology 38: 88–97.
Heck, K.L., and T.A. Thoman. 1984. The nursery role of seagrass meadows in the upper and lower reaches of the Cheasapeake Bay. Estuaries 7: 70–92.
Heck, K.L., L.D. Coen, and S.G. Morgan. 2001. Pre- and post-settlement factors as determinants of juvenile blue crab abundance; results from the north-central; Gulf of Mexico. Marine Ecology Progress Series 222: 163–176.
Heck, K.L., G. Hays, and R.J. Orth. 2003. Critical evaluation of the nursery role hypothesis for seagrass meadows. Marine Ecology Progress Series 253: 123–136.
Hoff, J.G., and R.M. Ibara. 1977. Factors affecting the seasonal abundance, composition, and diversity of fishes in a southeastern New England Estuary. Estuarine, Coastal and Marine Science 5: 665–678.
Holt, S.A., C.L. Kitting, and C.R. Arnold. 1983. Distribution of young Red Drum among different seagrass meadows. Transactions of the American Fisheries Society 112: 267–271.
Jenkins, G.P., and K.P. Black. 1994. Temporal variability insettlement of coastal fish (Sillaginodes punctata) determined by low-frequency hydrodynamics. Limnology and Oceanography 39: 1744–1754.
Jenkins, G.P., K.P. Black, M.J. Wheatley, and D.N. Hatton. 1997. Temporal and spatial variability in recruitment of a temperate, seagrass-associated fish is largely determined by physical processes in the pre- and post-settlement phases. Marine Ecology Progress Series 148: 23–35.
King, B.D. 1971. Study of migratory patterns of fishes and shellfishes through a natural pass. Technical Report Project 2-55-R, Texas Parks and Wildlife Department, Austin, TX.
Kraus, N.C. 2007. Coastal inlets of Texas. Proceedings coastal sediments 2007, 1475–1488. Reston: ASCE Press.
Milbrandt, E.C., R.D. Bartleson, L.D. Coen, O. Rybak, M.A. Thompson, J.A. DeAngelo, and P.W. Stevens. 2012. Local and regional effects of reopening a tidal inlet on estuarine water quality, seagrass habitat, and fishes assemblages. Continental Shelf Research 41: 1–16.
Minello, T.J. 1999. Nekton densities in shallow estuarine habitats of Texas and Louisiana and the identification of essential fishes habitat. American Fisheries Society Symposium 22: 43–75.
Minello, T.J., and R.J. Zimmerman. 1983. Fishes predation on juvenile brown shrimp, Penaeus aztecus: the effect of simulated Spartina structure on predation rates. Journal of Experimental Marine Biology and Ecology 72: 211–231.
Minello, T.J., R.J. Zimmerman, and E.X. Martinez. 1989. Mortality of young brown shrimp (Penaeus aztecus) in estuarine nurseries. Transactions of the American Fisheries Society 118: 693–708.
Morgan, M.D. 1980. Grazing and predation of the grass shrimp, Palaemonetes pugio. Limnology and Oceanography 25: 896–902.
Nañez-James, S.E., G.W. Stunz, and S.A. Holt. 2009. Habitat use patterns of newly settled southern flounder (Paralichthys lethostigma) in Aransas-Copano Bay, Texas. Estuaries and Coasts 32: 350–359.
National Marine Fisheries Service. 2014. Fisheries economics of the United States, 2012. U.S. Dept. Commerce, NOAA Tech. Memo. NMFS-F/SPO-137, 175p. https://www.st.nmfs.noaa.gov/st5/publication/index.html. Accessed 14 September 2015.
Neahr, T.A., G.W. Stunz, and T.J. Minello. 2010. Habitat use patterns of newly settled spotted seatrout in estuaries of the north-western Gulf of Mexico. Fisheries Management and Ecology 17: 404–413.
Nelson, D.M.. 1992. Distribution and abundance of fishes and invertebrates in Gulf of Mexico estuaries, Volume I: data summaries. ELMR Rep. No. 10. NOANNOS Strategic Environmental Assessments Division, Rockville, MD. 273 pp.
Nevins, J.A., J.B. Pollack, and G.W. Stunz. 2013. Characterizing nekton use of the largest unfished oyster reef in the U.S. in comparison to marsh edge and nonvegetated estuarine habitats. Journal of Shellfishes Research 33: 227–238.
Patillo, M.E., T.E. Czapla, D.M. Nelson, and M.E. Monaco. 1997. Distribution and abundance of fishes and invertebrates in Gulf of Mexico Estuaries Volume II: species life history summaries. Silver Spring: National Oceanic and Atmospheric Administration/National Ocean Service Strategic Environmental Assessments Division of Estuarine living marine resources program. Report No. 11.
Pile, A.J., R.N. Lipcius, J. Van Montfrans, and R.J. Orth. 1996. Density-dependent settler-recruit-juvenile relationships in Blue Crab. Ecological Monographs 66: 277–300.
Pinheiro, J., D. Bates, S. DebRoy, D. Sarkar, and R Core Team. 2015. nlme: Linear and nonlinear mixed effects models. R package version 3.1-121.
Pugesek, B., M. Baldwin, and T.V. Stehn. 2008. A low intensity sampling method for assessing Blue Crab abundance at Aransas National Wildlife Refuge and preliminary results on the relationship of Blue Crab abundance to Whooping Crane winter mortality. Proceedings North American Crane Workshop 10: 13–24.
Reese, M.M., G.W. Stunz, and A.M. Bushon. 2008. Recruitment of estuarine-dependent nekton through a new tidal inlet: the opening of Packery Channel in Corpus Christi, TX, USA. Estuaries and Coasts 31: 1143–1157.
Reid, G.K. 1957. Biologic and hydrographic adjustment in a disturbed Gulf Coast estuary. Limnology and Oceanography 2: 198–212.
Rooker, J.R., and S.A. Holt. 1997. Utilization of subtropical seagrass meadows by newly settled Red Drum (Sciaenops ocellatus): patterns of distribution and growth. Marine Ecology Progress Series 158: 139–149.
Rooker, J.R., G.J. Holt, and S.A. Holt. 1998a. Vulnerability of newly settled red drum (Sciaenops ocellatus) to predatory fish: is early life survival enhanced by seagrass meadows? Marine Biology 131: 141–151.
Rooker, J.R., S.A. Holt, M.A. Soto, and G.J. Holt. 1998b. Post-settlement patterns of habitat use by sciaenid fishes in subtropical seagrass meadows. Estuaries 21: 318–327.
Scharf, F.S., and K.K. Schlicht. 2000. Feeding habits of red drum in Galveston Bay, Texas: seasonal diet variation and predator–prey size relationships. Estuaries 23: 128–139.
Schwarz, C.J. 2012. Design and analysis of BACI experiments. Lecture, Department of statistics and actuarial science, Simon Fraser University, Burnaby, BC, Canada.
Searcy, S.P., D.B. Eggleston, and J.A. Hare. 2007. Environmental influences on the relationship between juvenile and larval growth of Atlantic Croaker (Micropogonias undulatus). Marine Ecology Progress Series 349: 81–88.
Shepsis, V., and J. Carter. 2007. Cedar Bayou – Inlet dynamics and engineering, Proceedings of Coastal Sediments 2007, ASCE Press.
Smith, E.P. 2002. BACI Design. In Encyclopedia of environmetrics, volume 1, ed. AH El-Shaarawi and WW Piegorsch, 141–148. Chichester: John Wiley and Sons, Ltd., Printer.
Stewart-Oaten, A., W.W. Murdoch, and K.R. Parker. 1986. Environmental impact assessment: pseudoreplication in time? Ecology 67: 929–940.
Stokes, G.M. 1977. Life history studies of Southern Flounder (Paralichthys lethostigma) and Gulf Flounder (P. albigutta) in the Aransas Bay area of Texas. Texas Parks and Wildlife Department Technical Series 25: 1–37.
Stunz, G.W., and T.J. Minello. 2001. Habitat-related predation on juvenile wild-caught and hatchery-reared red drum (Sciaenops ocellatus). Journal of Experimental Marine Biology and Ecology 260: 13–25.
Stunz, G.W., T.L. Linton, and R.L. Colura. 2000. Age and growth of Southern Flounder in Texas waters, with emphasis on Matagorda Bay. Transactions of the American Fisheries Society 129: 119–125.
Stunz, G.W., T.J. Minello, and P.S. Levin. 2002a. Growth of newly settled red drum Sciaenops ocellatus in different estuarine habitat types. Marine Ecology Progress Series 238: 227–236.
Stunz, G.W., T.J. Minello, and P.S. Levin. 2002b. A comparison of early juvenile red drum densities among various habitat types in Galveston Bay, Texas. Estuaries 25: 76–85.
Texas Parks and Wildlife Department. 2003. Saltwater finfishes research and management in Texas: a report to the Governor and the 78th Legislature, Austin, TX.
Ward, G.H. 2010. A time line of Cedar Bayou. Interagency contract 0900010973 report to Texas Water Development Board, Center for Research in Water Resources, University of Texas at Austin.
Weinstein, M.P. 1979. Shallow marsh habitats as primary nurseries for fishes and shellfish, Cape Fear River, North Carolina. Fishery Bulletin 77: 339–357.
Westwood, C.M., and F. Chavez-Ramirez. 2005. Patterns of food use of wintering Whooping Cranes in coastal Texas. Proceedings North American Crane Workshop 9: 133–140.
Wood, C. 1967. Physioecology of the grass shrimp, Palaemonetes pugio, in the Galveston Bay estuarine system. Contributions to Marine Science 12: 54–79.
Zuur, A., E.N. Ieno, and G.M. Smith. 2007. Analyzing ecological data. New York: Springer Science and Business Media.
Acknowledgments
We would like to thank Aransas County, Texas, and the Coastal Conservation Association Texas, for funding this research. We would also like to thank the staff at the Harte Research Institute, the Center for Sportfish Science and Conservation, and Texas A&M University-Corpus Christi, for their help in completing this study. A special thanks to all of those who helped process and analyze the samples collected during this study, especially Dr. Judd Curtis, Alex Tompkins, Chas Downey, Ashley Ferguson, David Norris, Meghan Hargis, Joy Brown, Patrick McKinney, and Zach Crain.
Author information
Authors and Affiliations
Corresponding author
Additional information
Communicated by Lawrence P. Rozas
Rights and permissions
About this article

Cite this article
Hall, Q.A., Robillard, M.M.R., Williams, J.A. et al. Reopening of a Remote Tidal Inlet Increases Recruitment of Estuarine-Dependent Nekton. Estuaries and Coasts 39, 1769–1784 (2016). https://doi.org/10.1007/s12237-016-0111-3
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s12237-016-0111-3