
Abstract
The charophytic green alga Nitellopsis obtusa was first reported in the New World in the St. Lawrence River in 1978. Since that time, N. obtusa has been reported from inland lakes throughout Michigan, northern Indiana, and western New York and has been listed as an aggressive invasive species by the United States Geological Survey. This study addresses the distribution of N. obtusa by surveying 390 waterbodies throughout New York. Previous reports are confirmed and new localities are presented, including new reports from five counties (Franklin, Ontario, Seneca, Wayne, and Yates). In total, N. obtusa was found in 17 counties at 31 sites, including 16 inland lakes, seven sites in the St. Lawrence River, and eight sites in Lake Ontario.
Similar content being viewed by others
Avoid common mistakes on your manuscript.
Nitellopsis obtusa (Desv. in Loisel.) J. Groves (Charophyta: Characeae) is the only extant species of the Early Quaternary genus Nitellopsis (Soulié-Märsche et al., 2002; Feist et al., 2005). Nitellopsis obtusa (Figs. 1, 2) is a dioecious, robust species, that is capable of growing more than 2 m tall in dense beds at depths to 10 m (Simons & Nat, 1996; John, 2002; Pullman & Crawford, 2010). Wood and Imahori (1965) placed N. obtusa in the tribe Chareae based on the 5-celled coronula and branchlets that do not furcate. Nitellopsis obtusa differs from other members in the Chareae by lacking stipulodes and possessing specialized star-shaped bulbils (Fig. 1B, C). These bulbils are unique among the Characeae in their complex morphology, role in vegetative reproduction, and star-like shape (Bharathan, 1987; John, 2002). This shape gives rise to the common name Starry Stonewort.

Nitellopsis obtusa. A. Habit. Bulbils develop at all nodes on the thallus including main axis nodes (arrows), branchlet nodes (arrowheads) and rhizoid nodes (not shown). B. Ventral bulbil view C. Dorsal bulbil view.

Developing Nitellopsis obtusa antheridium (male reproductive structure) shown at branchlet node.
Nitellopsis obtusa has a widespread, yet rare distribution in its native range of Europe and Asia (Simons & Nat, 1996; Soulié-Märsche et al., 2002; Kato et al., 2014). Previously unknown in the New World, the first record of N. obtusa in North America was in the St. Lawrence River in 1978, where it was hypothesized to be introduced by ballast water from trans-oceanic shipping (Geis et al., 1981). In 1983, it was recorded in the St. Clair-Detroit River system in Michigan, and is currently distributed throughout Michigan’s inland lakes, northern Indiana, and western New York (Schloesser et al., 1986; Eichler, 2010; Pullman & Crawford, 2010).
Initial studies in North America were cursory and suggested Nitellopsis obtusa outcompetes native aquatic vegetation and inhibits fish spawning (Pullman & Crawford, 2010; Kipp et al., 2014). This species is a major nuisance in lakes that hold recreational value: it fouls boat motors and impedes swimming and fishing. Nitellopsis obtusa fragments easily and may be spread as debris on boats and trailers. Control methods have been attempted, but the United States Geological Survey has stated that successful eradication is expensive (Kipp et al., 2014). Reports from lake associations and state agencies indicate that the range of N. obtusa is increasing in Michigan and New York (Pullman & Crawford, 2010; Eichler, 2010). Eichler (2010) reported the distribution of N. obtusa in New York, adding 13 sites to the 10 reported by Geis et al. (1981). Voucher specimens are important for the collection information they provide and to confirm determinations. Attempts were made to locate voucher specimens of N. obtusa summarized in Eichler (2010) without success. In this study we confirm, with voucher specimens, 13 of 23 previously reported localities (Geis et al., 1981; Eichler, 2010) and report 18 new records of N. obtusa, for a total of 31 sites in New York.
Materials and methods
Waterbodies (canals, lakes, ponds, rivers, streams, etc.) were selected using the Spatial Survey Design and Analysis package in R (Kincaid & Olsen, 2013) to place 400 points throughout New York. Satellite imagery was then used to locate the body of water with an area greater than 0.05 sq km closest to each point. Of these, 390 sites were accessible and were sampled from June to September 2014 (Fig. 3). Near shore regions were surveyed by wading and the use of a dredge fashioned after Allen (1887). The New York Department of Environmental Conservation’s guidelines for preventing the spread of invasive species were followed by soaking all collecting materials in Formula 409® for at least 10 minutes after each use. Water conductivity and pH measurements were collected with an In Situ SmarTroll MP (Ft. Collins, CO, U.S.A). Living material was collected in triplicate in Whirl-Paks® (Nasco, Fort Atkinson, WI, U.S.A.) and returned to The New York Botanical Garden for identification and vouchering. Voucher specimens were deposited in The William and Lynda Steere Herbarium (NY) and when possible, duplicates were distributed to the Academy of Natural Sciences of Drexel University (PH) and the Norton-Brown Herbarium, University of Maryland-College Park (MARY). Additionally, herbarium collections (NY) were examined for other New York Nitellopsis obtusa records.

Map of New York, U.S.A. showing county boundaries and the 390 sites sampled (dots).
Discussion
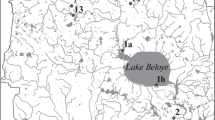
Nitellopsis obtusa localities are listed in Table I. By combining an herbarium survey with 2014 field work we were able to confirm N. obtusa in 13 of 23 previously reported sites (Geis et al., 1981; Eichler, 2010). Geis et al. (1981) reported 10 sites in close proximity in Jefferson and St. Lawrence counties. We confirmed sites in both counties but did not re-survey all (Geis et al., 1981) sites. The two previously reported sites we were unable to confirm were Waneta Lake and Lake Ontario at Wilson Tuscarora State Park (Eichler, 2010). Because both of these sites include large areas, it is possible that the exact location was missed by our sampling strategy. We found N. obtusa for the first time in seven inland lakes, five sites in the St. Lawrence River, and six sites in Lake Ontario (Fig. 4). These include new reports in five counties: Franklin, Ontario, Seneca, Wayne, and Yates. Nitellopsis obtusa is distributed throughout the St. Lawrence River corridor and across the central Finger Lakes region of New York (Fig. 4). Notably, N. obtusa was not found in other areas of New York (e.g., The Adirondack Park, The Catskill Park, Long Island).

Map of New York, U.S.A. showing county boundaries and Nitellopsis obtusa localities listed in Table I. Gray boxes indicate the initial reports by Geis et al. (1981). Green circles indicate records from Eichler (2010) that were confirmed in this study. Orange pentagons indicate records from Eichler (2010) that were not confirmed. Red diamonds indicate new records. For the purposes of display, nearby records were aggregated (see Table I).
Geis et al. (1981) reported water chemistry parameters including pH and conductivity for 19 sites in a 120 km stretch of the St. Lawrence River. Table II presents water chemistry measurements for the St. Lawrence River, as well as parameter ranges for all year 2014 sites where Nitellopsis obtusa was found in New York. The pH and conductivity values in the St. Lawrence River were similar between Geis et al. (1981) and our year 2014 measurements. However, the values across all N. obtusa year 2014 sites show a broader range, indicating that N. obtusa inhabits a wider range of pH and conductivity than previously known in North America. Simons and Nat (1996) reported water chemistry parameters for N. obtusa habitats in The Netherlands. The geographic coverage (Simons & Nat, 1996) is small when compared to year 2014 sites, but there are differences that are worth noting. The minimum pH of The Netherlands habitats is lower than the measurements from year 2014 sites, while the maximum pH is similar (Table II). This indicates that native N. obtusa occupies a broader range of habitats than North American N. obtusa. The minimum conductivity recorded from year 2014 sites is 160 μS/cm lower than the conductivity measured from The Netherlands. It is unclear whether this difference is the result of a broader tolerance in North American N. obtusa or a lack of sampling lakes with low conductivity in The Netherlands. The maximum conductivity of 2250 μS/cm in its native range suggests that N. obtusa is able to inhabit slightly brackish water, which is consistent with the ballast water hypothesis of introduction. It remains unknown whether North American N. obtusa has a similar salinity tolerance to native populations, or if the introduced N. obtusa has a restricted physiological tolerance.
Mann et al. (1999) reported that only male sexually reproductive Nitellopsis obtusa plants occur in North America and that N. obtusa could be moved clonally from lake to lake by human activity. Our work supports these statements; of the specimens that were reproductive, only male specimens were found in New York (Fig. 2). Pullman and Crawford (2010) reported female N. obtusa plants in Michigan. However, Figure 3 of Pullman and Crawford (2010) describes “orange-colored oocytes,” but antheridia are pictured. A relationship between latitude and gender has been suggested for N. obtusa in its native range, with only male plants observed at higher latitudes (Soulié-Märsche et al., 2002). It is possible that female plants exist in North America but have avoided detection.
Endozoochory, dispersal via vertebrate digestive tract (predominately waterfowl), has been proposed as the primary dispersal pathway for Characeae (Proctor, 1962, 1968; Charalambidou & Santamaría, 2002). This mechanism relies on mature oospores, which survive passing through the digestive tract of waterfowl. The apparent absence of reproductive female Nitellopsis obtusa in North America makes endozoochory an unlikely form of dispersal. Epizoochory, dispersal on feathers or feet of waterfowl, could disperse bulbils or vegetative propagules to new sites and has been discussed as a method of transport in Michigan (Pullman & Crawford, 2010). In New York, all lakes where N. obtusa was found had substantial human development, including boat launches. Our survey included many undeveloped lakes, and N. obtusa was not found in these low human impact areas. If waterfowl were indeed responsible for dispersing N. obtusa we would expect a higher frequency of N. obtusa statewide as well as occurrence in low human impact lakes. Consequently, in New York, N. obtusa is not likely transported by endozoochory or epizoochory and is most likely transported by humans.
The new county and site records we report indicate two patterns. The range of Nitellopsis obtusa in New York has not expanded beyond the boundary described by the localities reported in Eichler (2010). In areas where N. obtusa has been established, there is increasing density, with more local lakes becoming infested. This is particularly evident in Ontario and Yates counties in the Oswego River/Finger Lakes Watershed and in the Jefferson county region of the Lake Ontario Tributaries Watershed (Table I; Fig. 4). Further work is needed to understand how environmental variables and human activities contribute to the current distribution of N. obtusa in North America. Proactive management and public awareness will be necessary to limit and control the invasion of this enigmatic macroalgae.
Literature cited
Allen, T. F. 1887. Dredge for Chara. Botanical Gazette 12: 297.
Bharathan, S. 1987. Bulbils of some charophytes. Proceedings: Plant Sciences 97: 257–263.
Charalambidou, I. & L. Santamaría. 2002. Waterbirds as endozoochorous dispersers of aquatic organisms: a review of experimental evidence. Acta Oecologica 23: 165-176.
Eichler, L. W. 2010. Annual Report–2010 Darrin Fresh Water Institute Aquatic Plant Identification Program. DFWI Technical Report 2010-11
Feist, M., N. Grambast-Fessard, M. Guerlesquin, K. Karol, L. Huinan, R. M. McCourt, W. Qifei & Z. Shenzen. 2005. Treatise on Invertebrate Paleontology. PartB. Volume 1: Charophyta. The Geological Society of America, Inc., Boulder CO
Geis, J. W., G. J. Schumacher, D. J. Raynal & N. P. Hyduke. 1981. Distribution of Nitellopsis obtusa (Charophyceae, Characeae) in the St Lawrence River: a new record for North America. Phycologia 20: 211–214.
John, D. M. 2002. The Freshwater Algal Flora of the British Isles: An Identification Guide to Freshwater and Terrestrial Algae. Cambridge University Press, Cambridge.
Kato, S., H. Kawai, M. Takimoto, H. Suga, K. Yohda, K. Horiya, S. Higuchi & H. Sakayama. 2014. Occurrence of the endangered species Nitellopsis obtusa (Charales, Charophyceae) in western Japan and the genetic differences within and among Japanese populations: Nitellopsis obtusa from Lake Biwa. Phycological Research 62: 222-227.
Kincaid, T. M. & A. R. Olsen. 2013. spsurvey: spatial survey design and analysis. R package version 2.6. URL: http://www.epa.gov/nheerl/arm/
Kipp, R. M., M. McCarthy & A. Fusaro. 2014. Nitellopsis obtusa. USGS Nonindigenous Aquatic Species Database, Gainesville, FL.
Mann, H., V. W. Proctor & A. S. Taylor. 1999. Towards a Biogeography of North American Charophytes. Australian Journal of Botany 47: 445-458.
Pullman, G. D. & G. Crawford. 2010. A decade of starry stonewort in Michigan. LakeLine.
Proctor, V. W. 1962. Viability of Chara oospores taken from migratory water birds. Ecology 43: 528-529.
———. 1968. Long-distance dispersal of seeds by retention in digestive tract of birds. Science 160: 321-322.
Schloesser, D. W., P. L. Hudson & S. J. Nichols. 1986. Distribution and habitat of Nitellopsis obtusa (Characeae) in the Laurentian Great Lakes. Hydrobiologia 133: 91–96.
Simons, J. & E. Nat. 1996. Past and present distribution of stoneworts (Characeae) in The Netherlands. Hydrobiologia 340: 127–135.
Soulié-Märsche, I., M. Benammi & P. Gemayel. 2002. Biogeography of living and fossil Nitellopsis (Charophyta) in relationship to new finds from Morocco. Journal of Biogeography 29: 1703–1711.
Wood, R. D. & K. Imahori. 1965. A revision of the Characeae. Monograph of the Characeae. J. Cramer, Weinheim.
Acknowledgments
We would like to thank all of the landowners who allowed access to their property. We thank John D. Hall, Hilary A. McManus, Heather M. Meyer, William Pérez, Paul M. Skawinski and Norman Trigoboff for their contribution to field collections. Dario J. Cavaliere, Stephen D. Gottschalk, William Pérez and two reviewers for helpful suggestions on the manuscript. Dario J. Cavaliere provided assistance with micrographs. This work was supported by the Sarah K. de Coizart Article TENTH Perpetual Charitable Trust and this material is also based upon work supported by the National Science Foundation under Grant Numbers DEB-1020660 and DEB-1036466.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article

Cite this article
Sleith, R.S., Havens, A.J., Stewart, R.A. et al. Distribution of Nitellopsis obtusa (Characeae) in New York, U.S.A.. Brittonia 67, 166–172 (2015). https://doi.org/10.1007/s12228-015-9372-6
Published:
Issue Date:
DOI: https://doi.org/10.1007/s12228-015-9372-6