
Abstract
Coastal dunes in Northwestern Europe were subject to substantial scrub expansion during the past decades, a process often considered as a major problem for conservation management. This paper explores this process in De Westhoek nature reserve (Belgium), focussing on the three major constituents of the early scrub: Salix repens, Hippophae rhamnoides and Ligustrum vulgare. We study changes in dominance pattern between 1953 and 2010 based on vegetation maps and the demography of these species based on growth ring counting. We relate this data to the edaphic factors soil moisture and soil age.
Early scrub development occurred in three phases: (1) rapid, mainly secondary scrub expansion after release of grazing pressure, (2) a main shift in relative abundance of the three main species, and (3) further succession into tall scrub and woodland along with local scrub decay.
Edaphic factors play a major role in vegetation dynamics, but their relative importance diminishes along with landscape ageing. Simultaneously, the outcome of competitive interaction between scrub species, in which population demography plays a key role, gains importance. Primary and secondary scrub development seem to converge in terms of dominance patterns of S. repens, H. rhamnoides and L. vulgare. Spontaneous decay of mainly L. vulgare offers an opportunity for dune managers for the restoration of herbaceous dune vegetation with high conservation value.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
In Northwestern Europe, the cover of scrub in coastal dune vegetation substantially increased during the last century (Van der Maarel et al. 1985a; Provoost et al. 2011; Hodgkin 1984; Petit-Berghem 2004; Decuyper et al. 2020). Scrub has an important nature conservation value, mainly as a structural element in animal habitats (Mortimer et al. 2000). This value is appreciated within the European Habitats Directive, where two different types of coastal dune scrub are recognized and protected: type 2160 – ‘Dunes with Hippophae rhamnoides’ and type 2170 – ‘Dunes with Salix repens ssp. argentea’ (European Commission 2007).
From a management perspective, however, dune scrub is often seen as a problem rather than a valuable habitat. Scrub encroaches herbaceous vegetation, resulting in the decline of habitats with high conservation value (Richards and Burningham 2011; Binggeli et al. 1992; Isermann et al. 2007). Therefore, preservation of the priority habitat type 2130 – fixed coastal dunes with herbaceous vegetation or 2190 – humid dune slack is mostly given priority at the expense of scrub habitat types 2160 and 2170 (Houston 2008a, 2008b), also within De Westhoek nature reserve (Cosyns et al. 2013). The focus on removal may explain why relatively little is published on the ecology and vegetation dynamics of coastal scrub in Northwestern Europe (Provoost et al. 2011). However, this information is important for a better understanding of the entire dune vegetation dynamics and can contribute to a more efficient management (Leten et al. 2005). The main challenge in this respect is attaining an optimal alignment of mechanical scrub removal with natural vegetation dynamics in order to minimize management costs and optimize biodiversity gains.
Vegetation dynamics are typically related to climate, edaphic factors and internal dynamics (Glenn-Lewin et al. 1992). Considering the limited time frame, we only consider the latter two in this study. First, the spatial extent of different vegetation types is related to the distribution of the main edaphic factors. However, soil and vegetation patterns also depend on temporal dynamics; the spatial and temporal components of vegetation dynamics are not independent of each other. In this respect, the historical context of the landscape is crucial (Benjamin et al. 2005; Dutoit and Alard 1995). Scrub expansion on coastal dunes in Northwestern Europe is a typical example of vegetation development after abandonment of low-intensity grazing in semi natural habitats and a strong decrease in rabbit density caused by an outbreak of myxomatosis followed by VHR (Prévosto et al. 2011; Poschlod et al. 2005). However, in most dune systems, scrub expansion cannot be regarded as merely secondary succession (Glenn-Lewin et al. 1992; Mortimer et al. 2000). On former mobile dunes and in pioneer dune slacks, scrub develop starts from bare, mineral soil and follows a primary succession series (Olff et al. 1993; Lichter 2000). This means that scrub succession must be viewed in the context of both succession types and associated successional pathways and changes in soil properties.
A second aspect of vegetation dynamics, the internal vegetation dynamics, is strongly related to the demography of dominant species and lifespan of the entire scrub community (Oliver and Larson 1996; Curt et al. 2003; Magda et al. 2009; Good et al. 1990; Hudjetz et al. 2014). Van der Maarel et al. (1985b) investigated the demography of a number of woody species on the dunes of Voorne (the Netherlands), focussing on late-successional long-lived scrub and trees. However, similar information on the short-lived (roughly < 50 years) protagonists of early scrub development, namely Hippophae rhamnoides, Salix repens and Ligustrum vulgare, is lacking, although these species are currently major constituents of calcareous dune scrub in Northwestern Europe. Moreover, the early stages are important because they may determine vegetation development, vegetation pattern and species composition of mid- and late-successional stages.
This study wants to contribute to the knowledge on early scrub development on Northwestern European coastal dunes by tackling the following questions: (1) Can different phases of scrub development be distinguished according to the dominance pattern of the three main short-lived shrub species considered? (2) Can these patterns be related to soil development and moisture on the one hand and demography of the dominant species on the other? and (3) Is there a difference between primary and secondary scrub development?
We hypothesize that both edaphic and demographical factors play an important part in scrub dynamics, with different mechanisms prevailing in primary versus secondary development.
Material and methods
Study area
The field work for this study was carried out in De Westhoek nature reserve, the westernmost dune site along the Belgian coast, adjacent to the French border (2°35′ E – 51°5′ N, Fig. 1). The site got protected as a nature reserve in 1957 and was designated as a Habitats Directive Special Area of Conservation in 1996. Its surface area is 342 ha. Geologically it belongs to the Flemish dunes, which stretch from the Aa estuary in northern France to the IJzer estuary in Belgium. Most of this dune belt is young and still highly calcareous.

Location of De Westhoek Nature reserve (2°35′ E – 51°5′ N)
Most of the Belgian dunes were used for livestock grazing until the beginning of the twentieth century. In De Westhoek, grazing largely stopped around World War 2. Only a small part in the southwest was grazed until the 1960s (Provoost and Leten 2012). For several decades, site management concentrated on restriction of accessibility. Between 1997 and 2000, however, rather large-scale nature restoration measures were taken, including the removal of 26 ha of scrub and woodland in favour of dune grassland (EU habitat type 2130) and herbaceous dune slack vegetation (2190). Domestic grazers were re-introduced to preserve these habitats. Mainly Shetland ponies, Konik horses and Highland cattle are used, and grazing density fluctuates around one animal per five hectares (Leten et al. 2005; Galatowitsch 2012). Management targets for scrub mainly focus on preservation of the surface area.
Vegetation mapping
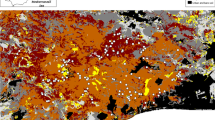
A detailed vegetation map based on field work was made in 2010. A true colour orthophoto (2008) with 0.3 meter pixels was used as a geographical reference. Maps of historical vegetation structure were derived from stereoscopic interpretations of aerial photographs. Three series of images on scale 1/5,000 were used: (1) panchromatic photos from 15 September 1953, (2) panchromatic photos from 20 August 1974 (both from the archives of the Flemish Ministry of Public Works), and (3) false colour-near infrared diapositives from 14 July 1989 (Eurosense, commissioned by the Flemish Government). All analogue imagery was scanned and georeferenced. Although the image quality differs considerably, four basic vegetation structural classes can be distinguished accurately on all photographs: bare sand, herbaceous vegetation, scrub and woodland. On the 1953 images, only low scrub appears. The image quality does not allow further discrimination of this low vegetation into different dominance types. On the more recent images taller scrub appears. It can be subdivided according to Salix repens, Hippophae rhamnoides and/or Ligustrum vulgare dominance, based on differences in vegetation height (through stereoscopic view), image tone, texture and on the 1989 image also colour. The vegetation map of D'Hondt (1981) and aerial images of 1917 (Royal Air Force, Documentation Centre for World War 1, Brussels), 1948 and 1968 (Belgian National Geographical Institute) were used as auxiliary information for vegetation mapping and determination of soil age. ArcGIS 10 was used for image georeferencing, digitizing of vegetation polygons and analysis of vegetation maps.
Soil/scrub age and groundwater level
Soil age, scrub age and soil moisture data were added as layers in a GIS environment. The soil and scrub age maps were derived from an overlay of the vegetation maps, resulting in five discrete classes. Soil age is defined here as the timespan since the onset of vegetation development, as deduced from the aerial photographs. Similarly, scrub age is expressed as the timespan since the first scrub appeared on a certain location. Phreatic groundwater levels are measured within the entire site since 1999. We created a map of the average spring (15 March – 15 April) groundwater level through interpolation of average levels in a 101 piezometer installed at the site. We used the mean of the spring levels within the period 2014–2018 because this was an average period in hydrological terms for which a maximum of data was available (Provoost et al. 2020).
Focal species
The focal species in this study are Salix repens, Hippophae rhamnoides and Ligustrum vulgare because the early stages of scrub in the area were almost exclusively composed of these species (Provoost and Leten 2012).
Salix repens growing in Western European dune slacks is treated as S. repens L. var. argentea (Sm.) Wimm. and Grab., synonymous with S. repens subsp. argentea, S. repens subsp. arenaria and S. arenaria (Stace et al. 2005). It is a low willow, attaining about 2 m in height at most, but its height highly depends on the landscape position. At the study site, the characteristic height of S. repens-dominated scrub is 1.1–1.2 m. Individuals growing between taller scrub species such as H. rhamnoides attain heights up to 2.2 m, while more exposed S. repens patches only reach 30 cm to 1 m (personal observations). Salix repens is found in cold and temperate parts of Eurasia and grows in nutrient-poor, moist habitats on different soil types (Tutin et al. 1964; Weeda et al. 1985). On Belgian coastal dunes, germination is largely restricted to newly formed dune slacks where S. repens is amongst the first colonizers. Very rarely, plants newly establish on north slopes (personal observations). Abundant arbuscular mycorrhiza as well as ectomycorrhiza are an aid to survive in nutrient-poor environments (Van der Heijden 2001). Once established, S. repens vegetatively propagates with subterranean or creeping stems and through this mechanism it is well adapted to sand burial. This explains why it can also be found on dune ridges high above the groundwater table or in dune grassland. Salix repens is wind- (or insect-) pollinated and dispersed, and is therefore a good colonizer (Vroege and Stelleman 1990). As a pioneer species it is only moderately shade-tolerant (Weeda et al. 1985).
Hippophae rhamnoides L. is a thorny, deciduous and dioecous shrub, naturally occurring from the Himalayas, through the Black Sea area to the coasts of Western Europe but widely naturalized outside this area. Several genetically distinct subspecies can be distinguished, revealing an Asiatic origin followed by a westward expansion and further speciation (Jia et al. 2012). Within Central and Western Europe, three subspecies are recognized: fluviatilis and carpatica, occurring along mountain rivers, and the coastal subspecies rhamnoides. The latter is not found south of the 10°C January isotherm (northwest Spain) because low temperatures are required prior to seed germination (Pearson and Rogers 1962). To the north, it is found up to 68°N in Norway (Stewart and Pearson 1967). The temperature-dependent limitation of seed germination, combined with its preference for calcareous soils, results in an actual core distribution of subspecies Hippophae from Normandy in northern France up to Bergen in the Netherlands, north of which soil lime content is significantly lower (Rozema et al. 1985) and abundance of Hippophae is lower. During the late glacial and early postglacial, H. rhamnoides was widespread on the well buffered mineral soils, but it retreated in mountainous habitats and along the coasts during the Holocene reforestation (Bartish et al. 2006).
According to Stewart and Pearson (1967), the height of H. rhamnoides ranges between one and nine metres. At the study site, however, the characteristic height is one to three metres, with occasionally larger plants of up to about four metres (personal observations). Rogers (1961) reports a rapid vegetative growth with rhizome like roots growing up to two metres per year. Hippophae rhamnoides is also successfully reproducing in a generative way. Its fleshy berries are consumed and transported by birds and seed viability reaches 95 to 100% (Stewart and Pearson 1967).
Zoon (1995) reported that on Dutch dunes individual shoots were rarely older than 25 years. However, on the Wadden Sea Island of Ameland, Decuyper et al. (2020) occasionally sampled individual ramets older than 40 with a maximum of 56 years. Skogen (1972) even found 80 year old ramets along the fjord of Trondheim in Norway, suggesting maximum age strongly depends on habitat type or climate.
Hippophae is found in a relatively wide range of dune landscape types (Weeda et al. 1987; Meltzer 1941). The capacity of H. rhamnoides to grow in nutrient-poor soils is attributed to its nitrogen fixating ability (Stewart and Pearson 1967; Troelstra et al. 1987) and vesicular-arbuscular mycorrhiza (Gardner et al. 1984). Mature Hippophae is shade-intolerant and soon dies if overgrown by trees or large shrubs. Also seedling growth is suppressed when shaded (Pearson and Rogers 1962).
Ligustrum vulgare L. is a deciduous to semi-evergreen shrub, native to Central and Southern Europe and North Africa but widely naturalized in other temperate parts of the world. According to Tutin et al. (1964), Ligustrum can attain a height of up to 5 m. However, in our study area within the scrub it reaches a typical height of 1 to 3 m (personal observations). Ligustrum grows on a variety of mostly calcareous soil types. It is found in scrub but also in the understory of deciduous woodland (Tutin et al. 1964–1980, Zhao et al. 2013). Vegetative spread occurs through root suckers and stem layers but is relatively limited. The species is easily distributed over long distances by fructivorous birds and pollination is entomophilous. Ligustrum is moderately tolerant to sand burial (Weeda et al. 1988).
Autecological and edaphic data
Field measurements were carried out in De Westhoek nature reserve in order to determine the lifespan, inundation tolerance and preference for soil age of the three focal species. Lifespan, regardless of recruitment date, was obtained through growth ring counting on basal stem fragments of dead ramets (= a single stem shoot of an individual). Randomly selected stems of H. rhamnoides (n = 134), L. vulgare (n = 57) and S. repens (n = 56) were cut and collected in 2002 and 2014. They were taken to the lab and polished before counting the growth rings.
Furthermore, 1,213 randomly distributed growing locations of the focal species were measured in 2014 using cm-accurate GPS. The points were overlaid with the GIS layers in order to obtain figures on soil age and groundwater level below the surface for each location.
The preference of a species for a certain soil age class is expressed as the relative number of individuals of that species within a certain age class, divided by the relative portion of the surface area of that age class within the entire site.
Scrub age plots
Age composition of the different species within actual scrub stands was investigated in 14 plots of 8 by 8 m2, scattered throughout the site (Fig. 2, 2010 map). The plots were stratified according to soil age and avoided site margins as well as woodland areas. Within these plots, every single shrub ramet was cut and a basal stem sample was taken to the lab for age determination through visual growth ring counting. The rare missing age values due to partial decay of Hippophae or Ligustrum stem cores were estimated using a stem diameter versus age linear regression. The relative representation of age classes per species gives an idea of the potential outcome of interspecific competition. However, in a synchronic approach the demographic composition is biased by lifespan because of the disappearance of short lived individuals.

Above – vegetation maps of De Westhoek Nature reserve in 1953, 1974, 1989 and 2010. The 2010 map also shows the location of the scrub demography plots. Below – surface area of the different vegetation types derived from the vegetation maps above
The position of the plots was measured with a cm-accurate GPS device. Soil and scrub age were determined through GIS overlay of the coordinates with the soil and scrub age maps. Field work for this part of the study was carried out in the summer of 2002. Statistical tests were carried out in RStudio version 1.3.1073.
Results
Overall vegetation change
Aerial photo interpretation reveals two main landscape trends at the site during the second half of the twentieth century (Fig. 2). The first trend is dune fixation (decrease of bare sand), and the second one is the expansion of scrub and woodland. The most substantial increase in scrub cover, from 25 to 48% of the area, occurred within the first time interval 1953–1974. Between 1974 and 1989, there was little change in the balance between bare sand, herbaceous vegetation, scrub and woodland. However, there was a marked change in the composition of the scrub, with a clear increase of Hippophae and Ligustrum at the expense of Salix repens. Between 1989 and 2010 scrub cover only increased with 0.9%. The increase would have been 5.1% without the artificial scrub clearing. Between 1997 and 2000, 26 ha of tree plantations and (largely Ligustrum) scrub were removed as a nature restoration measure. The cleared area mostly developed into herbaceous vegetation and low S. repens scrub, resulting in a 14-ha net decrease of the scrub area. Also within the last phase, a substantial portion of the early scrub, dominated by S. repens, Hippophae and Ligustrum got replaced by other scrub types.
General scrub dynamics
Vegetation transition schemes based on interpretations of aerial photographs are shown in Tables 1, 2 and 3. In 1953, substantial parts of the dune slacks were covered with low Hippophae and S. repens scrub (Herbauts 1971). This vegetation is recognizable on the image, but the photo quality does not allow differentiation between the two dominant species. Between 1953 and 1974, this scrub expanded, grew taller and differentiated into different (distinguishable) vegetation types dominated by S. repens (25%), Hippophae (50%) and/or Ligustrum (25%, Table 1). Young slacks were colonized by S. repens but soon evolved into Hippophae/S. repens mixed vegetation. Hippophae and Ligustrum mainly showed an expansion at the expense of formerly grazed herbaceous vegetation, indicating that both species can act as pioneers in secondary scrub development. Secondary scrub accounts for 62.5 ha or 67% of the newly developed scrub.
Between 1974 and 1989 (Table 2), S. repens showed a general decline. An area of 24 ha or 82.5% of the initial S. repens-dominated vegetation succeeded into other scrub types dominated by Hippophae (roughly 75%) and Ligustrum (25%). Only 1 ha of S. repens patches (3.8%) remained unchanged between 1974 and 1989.
Hippophae-dominated scrub increased by nearly 50%, at the expense of S. repens, herbaceous vegetation as well as newly fixed dunes. Hippophae/S. repens codominated vegetation appeared to be a transitional type, where Hippophae gradually took over dominance.
Ligustrum-dominated scrub expanded with nearly 90%, mainly at the expense of Hippophae (68%). However, also the reverse transition of Ligustrum into Hippophae dominant scrub occured, albeit less frequently. Succession from S. repens vegetation accounted for 25% of the newly formed Ligustrum scrub. This process was unambiguous, as no transition of Ligustrum into S. repens occurred. Also the spatial patterns of both successional pathways towards Ligustrum dominance differed markedly. Detailed inspection of the vegetation maps in GIS revealed that the transition of S. repens into Ligustrum scrub largely occured in large distinct patches, while the transition starting from Hippophae showed a pattern of scattered small patches.
Transitions of scrub into bare sand and herbaceous vegetation are mainly related to eastward transgression of the central mobile dune, covering all vegetation on its path. Scrub regression into herbaceous vegetation only occurs at small scale. It is mainly the result of Ligustrum decay (about 1 ha or 5% of the initial area), decay of Hippophae scrub only accounts for about 0.5 ha (0.9% of the initial Hippophae-dominated area).
The vegetation changes between 1989 and 2010 (Table 3) were substantially influenced by artificial scrub clearing in 1997–2000 (coloured columns). Still, a number of distinct trends can be observed within the area unaffected by this clearing. First, scrub dominated by S. repens spontaneously increased from 6.8 to 12.1 ha, with an additional 5.4 ha formed within the cut areas. The natural expansion mostly occured as pioneer vegetation development in newly formed dune slacks (53%) but is also due to decay of Hippophae in mixed scrub with S. repens (37%). Again, S. repens scrub was highly ephemeral, with only 15% of the 1989 area still appearing as S. repens-dominated vegetation in 2010.
Second, Hippophae scrub was subject to large changes between 1989 and 2010. About 16% succeeded into ‘other’ scrub types (dominated by Crataegus monogyna, Prunus spinosa, Salix cinerea and others) or woodland, and another 16% of the original area disappeared due to scrub clearing, the movement of mobile dunes or spontaneous decay. On the other hand, about 30 ha of it was newly formed, mainly as pioneer scrub on the stabilized central mobile dune. Only a small area of Hippophae succeeded into Ligustrum-dominated or mixed scrub (2.2 ha). By contrast, a substantial transition of Ligustrum into Hippophae did occur (8.6 ha).
Finally, between 1989 and 2010, Ligustrum-dominated scrub showed a major decline from 42.7 to 12.9 ha (−70%). Scrub clearing decreased the Ligustrum scrub by 16.5%, succession into woodland and mature scrub accounted for 16%, and dune mobility (sand covering) accounted for another 12%. About 7 ha or 16% of the Ligustrum scrub regressed into herbaceous vegetation. In reality, Ligustrum decay was larger, but part of the open habitat was already colonized by Hippophae in 2010. This likely explains the large area of Ligustrum succeeding into Hippophae (26%).
Overall, the total scrub cover in De Westhoek increased from 88 ha (26% of the area) in 1953 to 166 ha (49%) in 2010. This area is the result of substantial internal landscape dynamics involving vegetation succession and regression, large-scale dune mobility and artificial scrub removal. The actual scrub developed through primary succession from bare sand (46%), secondary succession from formerly grazed herbaceous vegetation (28%) or was already scrub in 1953 (26%).
Species autecology
Figure 3 shows that S. repens shows a clear preference for moist conditions with a median average spring groundwater level of 21 cm below the surface level and a narrow optimum. However, the species tolerates a wider moisture range, from very wet (spring flooding) to dry conditions (groundwater table > 5 m deep). Hippophae rhamnoides and Ligustrum vulgare show a similar response to groundwater levels. Both species do not tolerate spring flooding and prefer significantly dryer soil conditions than S. repens, with median average spring groundwater level values around 1,05 m below the ground surface (one-way ANOVA, d.f. = 2, F = 40.41, P << 0.01).

Comparison of spring groundwater depth in the period 2014–2018 across the three study species Salix repens (n = 185), Hippophae rhamnoides (n = 560) and Ligustrum vulgare (n = 468). One-way ANOVA: d.f. = 2, F = 40.41, P << 0.01. A post-hoc Tukey HSD test reveals that groundwater levels for S. repens significantly differ from those for both L. vulgare and H. rhamnoides (P << 0.01)
Salix repens grows within a wide range of soil age (Fig. 4). It is one of the first colonizers in newly formed dune slacks but can also be found in old wet to dry dune grassland on soils fixed for many decades or probably even centuries. Also Hippophae is present in a relatively wide soil age range but with a clear shift in preference towards the higher age classes (Fig. 4). By contrast, Ligustrum is hardly found in young, recently fixed dunes. The species has the most pronounced preference for old soils at the site, fixed for at least several decades.

Preference of different life stages of Salix repens (n = 185), Hippophae rhamnoides (n = 560) and Ligustrum vulgare (n = 468) for fixation age of the landscape within De Westhoek nature reserve. Preference is defined in the methods section
Lifespan patterns based on ring counts of dead stems are shown in Fig. 5. Salix repens and H. rhamnoides show a clearly peaked distribution with a median age of nine and thirteen years, respectively. Maximum ages are nineteen for Salix and 39 for Hippophae. The age distribution of dead Ligustrum ramets at the site is not clearly peaked. The median age is 23 years, the oldest individual found was 36. A summary of the species’ characteristics from the literature and own observations is presented in Table 4.

Age distribution of individual dead ramets of Salix repens (n = 56), Hippophae rhamnoides (n = 134) and Ligustrum vulgare (n = 57) in De Westhoek nature reserve
Age composition of scrub plots
Table 5 summarizes the characteristics of the forteen plots in which scrub was cut to determine ramet age. The age composition of the different species is presented in Fig. 6 as cumulative curves of ramet establishment. Three groups of plots can be distinguished in function of soil moisture (depth of groundwater table). Plots a–d (Table 5) represent scrub on dry ridges, where the average spring groundwater level is more than 3.5 m below the surface and has a negligible impact on vegetation development. In these parts of the dune, fixation and scrub development started relatively recently (after 1953). Scrub expansion is slow, only ten to twenty new ramets are formed within the first ten years. Hippophae expansion generally shows an s-shaped curve, indicating a phase of increasing establishment followed by a decrease. The latter phase had not yet been reached in plot a, which still showed vigorous rejuvenation at the time of cutting. After more than thirty years, scrub composition is still poorly differentiated with Hippophae clearly dominating. Although Ligustrum individuals of more than twenty years old are present in the plots, this species is still sparse at the time of cutting.

Cumulative ramet establishment of Hippophae rhamnoides, Salix repens and Ligustrum vulgare in scrub demography plots
The second and largest group of plots (Table 5, e through k) is situated in moist dune slacks, with average spring groundwater levels roughly between 55 cm and 1.75 m below the surface. This implies that vegetation and soil development are at least seasonally influenced by groundwater but that the surface is never inundated. In all plots, the soil is fixed for more than fifty years, so they have formerly been grazed and vegetation can be considered secondary scrub. Ligustrum and Hippophae are both abundant, but the plots show a marked variability in their abundance ratio. Plots e and j are dominated by Hippophae whereas plots g, h and i are dominated by Ligustrum, while f and k hold an intermediate position. Most demographic curves in plots e-k show a linear or concave down shape, indicating there was no phase of acceleration of rejuvenation or it has already disappeared out of the demographic image.
In plot e, tall scrub development is relatively recent. Hippophae appears in the late 1970s, gradually displaces Salix and still dominates the vegetation in 2002. Although one Ligustrum stem of over 20 years old is found, the species is hardly present in the plot at the time of cutting. The other plots f to k (Table 5) consist of old scrub which was already developing in 1953. The aerial photo interpretation reveals that, with the exception of plot k, all locations went through a phase of Ligustrum dominance. Plot k shows a shift in the balance between Hippophae and Ligustrum over time, without one of both completely prevailing. In plots g, h and i, Ligustrum was still dominating at time of cutting, with vigorously rejuvenation of the species in plot g. In plots f and j, the Ligustrum phase peaked in the 1970s and 80s and since then, Hippophae predominated. Plot j clearly shows Hippophae rejuvenation at the time of cutting.
The last group of plots (Table 5, l, m and n) consists of scrub in wet dune slacks, where the average spring groundwater table is higher than 50 cm below the surface level. Scrub expansion occurs much faster here compared to the dry dunes. In plots l and m, forty to one hundred new ramets established within ten years.
The slack plots are characterized by the presence or abundance of S. repens. Hippophae is dominant and still rejuvenating in the youngest plot (l), only fixed for about fifteen years at the time of cutting. As in plots a–d, Hippophae shows a clearly s-shaped curve here, with peak expansion in the early 1990s. In plot m, fixed about ten years earlier, Hippophae is still abundant but no longer rejuvenating. In plot n, situated in a dune slack formed about fifty years ago, Hippophae nearly disappeared and Ligustrum gradually took over.
Discussion
Early scrub expansion
Several scrub development phases with distinct dominance patterns can be distinguished in De Westhoek dunes. The site was characterized by rapid scrub development between 1953 and 1974, with an average expansion of 1.1% of the site’s total area per year. This phase lasted until the 1970s, when, ultimately, most of the slacks were covered with scrub and only the most dynamic locations, such as the central mobile dune and the foredunes, remained devoid of vegetation. The onset of the scrub encroachment is not clearly documented. High-quality terrain photographs and descriptions from the beginning of the twentieth century sketch a largely grazed, open landscape dominated by mobile dunes and herbaceous vegetation (Massart 1912). The slacks were covered by damp grassland and low scrub with S. repens, H. rhamnoides and, to a lesser extent, L. vulgare (De Bruyne 1905; Massart 1912; Rahir 1932). According to the descriptions of the site from in the 1950s and 1960s, scrub vegetation was clearly taller and denser, indicating the expansion was already well on its way (Depuydt 1967; Delaunois 1952).
A similar process of rapid scrub development is described by Van der Maarel et al. (1985a) for the dunes of Voorne (the Netherlands). These authors denominate it with the term ‘revolutionary succession’ and link it to the release from cattle grazing in 1910. This link with grazing abandonment applies to many of the coastal dunes (Provoost et al. 2011) and other semi-natural landscapes in Northwestern Europe (Prévosto et al. 2011; Poschlod et al. 2005) and is also the most probabe explanation for the onset of scrub expansion in De Westhoek dunes.
In the early 1950s, also the collapse of rabbit populations caused by myxomatosis (Fenner and Radcliffe 1965) stimulated scrub encroachment on dunes. This certainly holds for species with palatable juvenile shoots such as Crataegus monogyna or Prunus spinosa (Hodgkin 1984; Smit et al. 2010). In De Westhoek as well as in Oostvoorne, however, scrub expansion started before the myxomatosis outbreak, so it can only have played a secondary role.
The early phase of scrub expansion in De Westhoek can largely be considered a secondary vegetation development because tall scrub mainly invaded formerly grazed herbaceous vegetation or low scrub (Mortimer et al. 2000). Only 20% of the scrub area in 1974 resulted from primary development, that is, colonization of locations not vegetated in 1953. It consisted of Hippophae and S. repens, which, in contrast to Ligustrum, are able to colonize in very early, nutrient poor stages of (primary) vegetation succession (Weeda et al. 1988). Both species are adapted to nutrient-poor pioneer conditions through intense symbiotic relationships with fungi and nitrogen-fixing bacteria (Tadych and Blaszkowski 2000).
Secondary scrub, encroaching existing herbaceous vegetation, mainly consisted of Hippophae (55%), but also Ligustrum (24%) and S. repens (21%), were well represented. Salix repens was already present in the formerly grazed herbaceous vegetation in 1953 and likely it merely grew taller after the removal of livestock. The rapid expansion of Hippophae and Ligustrum in former grassland probably largely involved establishment through seed germination, as both species appeared much more scattered throughout the landscape.
Changes in dominance pattern
A second phase in scrub development is characterized by a shift in the relative abundance of the dominant species S. repens, H. rhamnoides and L. vulgare (Fig. 7). Our results describe these shifts and link them to species characteristics in a hypothetical way. Unravelling the mechanisms of competition would require experiments. At our study site, this phase of shifting dominance patterns largely coincides with the evolution observed between 1974 and 1989. Within this period, S. repens-dominated vegetation is largely displaced by Hippophae and Ligustrum. Both species are taller and on average live five to ten years longer than S. repens (Table 4), which most likely explains this evolution. Apparently, more than half of the larger Ligustrum-dominated patches have a S. repens history. This is probably related to the shade tolerance of Ligustrum, enabling the species to establishment in dense, relatively tall and therefore shaded S. repens vegetation. Once developed, Ligustrum prevents Hippophae colonization by its shading effect (Kollmann and Reiner 1996), resulting in a monospecific vegetation. It seems that the dense S. repens-dominated vegetation which developed on a large scale soon after the decline of the domestic grazing pressure, exerted a priority effect (Temperton et al. 2004), inhibiting the development of Hippophae but enabling the development of large Ligustrum-dominated scrub patches.

Transitions in dominance of Hippophae rhamnoides, Salix repens and Ligustrum vulgare during early scrub succession in De Westhoek dunes
Between 1989 and 2010, some patches show a transition of Hippophae- to S. repens-dominated scrub. This is due to Hippohae decay during long spring inundation in wet dune slacks. Salix repens is well adapted to this inundation and regains dominant if it is still present in the Hippophae-dominated vegetation. It is indicated as a regressional pathway in Fig. 7.
The competition balance between Hippophae and Ligustrum is more subtle. The aerial photograph interpretation mostly shows transitions from Hippophae to Ligustrum dominance, but, to a lesser extent, also the reverse transition occurs.
Hippophae is favoured in early stages of primary succession due to its ability to colonize mineral soils and its strong vegetative propagation (Rogers 1961). In well developed, organic soils, however, Ligustrum seems to gradually outcompete Hippophae, although both species can coexist in the scrub for more than thirty years. This is longer than the average life span of both species. The gradual replacement of Hippophae is confirmed by the demography plots in moist dune slacks where scrub development started at least fifty years ago. Nearly all these plots have been through a phase of Ligustrum dominance and most plots were still Ligustrum-(co)dominated at the time of cutting. As both species attain a similar height within the scrub, likely the longer lifespan and shade tolerance of Ligustrum vulgare explain its modest but decisive competitive superiority to Hippophae rhamnoides.
The reverse transition of Ligustrum into Hippophae-dominated scrub mainly occurred between 1989 and 2010 and at least partly goes along with the regression of Ligustrum scrub into herbaceous vegetation. We consider this regression a third phase in early scrub development, probably related with scrub age. At an age of at least 30 to 40 years, many Ligustrum stands seem to collapse on a small to relatively large scale. Subsequently, Hippophae can fill in this gap and regain dominance. This process is also seen in some of our scrub demography plots which contain remnants of old Ligustrum along with rejuvenating Hippophae.
The decreasing vitality of mature scrub is well documented for Hippophae (Sloet van Oldenruitenborgh 1976; Van der Maarel et al. 1985a). It is caused by plant-specific, soil-borne nematodes (Oremus and Otten 1981; Maas et al. 1982), possibly synergistically enforced by fungi (Zoon et al. 1993). Although no information was found for Ligustrum, the observed large-scale decay of the species suggests similar involvement of soil-borne pathogens. Scrub degeneration might also be caused by nutrient shortage, especially phosphorous (Zoon 1995). It is not clear whether nutrients or other chemical factors play a role in the scrub regression observed in De Westhoek dunes.
Edaphic versus demographical factors
In this study, two edaphic factors were taken into account, soil moisture and soil development expressed as the timespan since the onset of vegetation development. The importance of soil moisture is firstly reflected in differences in inundation tolerance between our three studied species. Hippophae rhamnoides and L. vulgare are clearly intolerant to prolonged flooding, in contrast to S. repens. Consequently, we can see that in very wet slacks S. repens can maintain or regain dominance when Hippophae is eliminated. This evolution is clearly observed between 1989 and 2010 and can, at least partly, be explained by Hippophae decay during the exceptionally wet year 2001 (Provoost et al. 2020). However, wet slacks with average spring groundwater tables exceeding 20 cm below the ground level only account for 4.6% of the area (Provoost et al. 2020), so quantitatively this process is of minor importance at the site. Second, soil moisture largely determines the speed of soil organic matter accumulation and, inextricably linked, vegetation development (Sevink 1991; Jones et al. 2008). This moisture effect is exerted with average groundwater tables up to about 2 m below the surface level (Provoost et al. 2004), which is the case in about 60% of the site. The effects of drought stress, causing this slowing down of vegetation development on dry dunes, is clearly noticeable in demography plots a–d, which show a much slower scrub expansion than the wet and moist plots.
Also, as soil development is slowed down on dry dunes, it takes much longer until Ligustrum can establish, so vegetation remains Hippophae-dominated much longer. Ligustrum indeed demands higher nutrient levels and only establishes in landscapes with a certain level of soil development (Weeda et al. 1988).
As the regional precipitation surplus shows strong temporal fluctuations (Provoost et al. 2020), a more extensive exploration of the relationship between groundwater level (or precipitation balance) and scrub dynamics requires a much higher temporal resolution of vegetation data and is therefore not an issue in this study. For the Westhoek dunes, a general increase in precipitation surplus was observed between 1953 and 1974, but it remained relatively stable between 1974 and 2010. However, we do not see a convincing relationship between these trends and our findings on scrub development.
The importance of soil development is mostly reflected in the gradual replacement of the pioneer species S. repens and H. rhamnoides by L. vulgare, which is clearly related to older, more organic soils. This perfectly fits within a facilitation driven, autogenic, primary succession, typically associated with coastal dunes (Olson 1958; Olff et al. 1993). However, the first phase of strong scrub expansion in De Westhoek, between 1953 and 1974, largely consisted of secondary scrub development in formerly grazed herbaceous vegetation. Here we see that Ligustrum colonizes habitats together with Hippophae and Salix; the facilitation phase of primary succession seems to be skipped. Based on the three species considered in this study, the composition of older stages of primary succession seems to converge with secondary developed scrub (Fig. 7). This might be expected because the changes in soil characteristics related to autogenic succession appear to diminish over time (Sevink 1991; Jones et al. 2008), together with the quantitative importance of soil factors as differentiating drivers of vegetation succession (Tilman 1990). Hence, this would result in a shift towards vegetation dynamics characterized by a more subtle competitional interaction between Hippophae and Ligustrum, in which, according to our results, population demography plays an important role. In future succession towards tall scrub and woodland, this shift towards vegetation dynamics associated with differences in plant life histories, growth rates and other competitive qualities is likely to reinforce (Lichter 2000; Huston and Smith 1987). However, as primary scrub succession at the site is at most about 65 years old and has only very locally resulted in Ligustrum dominance, the degree of convergence between primary and secondary scrub development remains unclear. The large-scale Ligustrum decay, for example, might be typically associated with secondary scrub.
Management implications
Scrub encroachment poses a main challenge for the conservation of species-rich herbaceous dune habitats. Figures on landscape changes in De Westhoek dunes show that scrub develops in different vegetation types over a wide soil moisture and soil age range. Only mobile dunes and frequently inundated slacks remain devoid of it. Within a highly scrub-encroached landscape, low- to moderate-pressure grazing management seems unable to reduce the scrub area or even substantially limit its expansion (Hoffmann et al. 2005; van der Hagen et al. 2020). Therefore, mechanical scrub clearing is applied in order to preserve the surface area of herbaceous vegetation. The phases of natural scrub decay, probably related to scrub age, can play an important role in achieving maximum synergy between natural vegetation dynamics and management targets. Our results show that the area of natural scrub decay was within the same order of magnitude as the artificial scrub clearing. This process offers a possibility to change scrub into herbaceous vegetation communities without having to deploy heavy machinery and thus substantially reduce costs and terrain damage.
Finally, special attention should be paid to EU-habitat type 2170 ‘Salix repens scrub’. It is an important coastal dune element with high conservation value. Salix repens is a niche specialist, highly tolerant to inundation, sand burial and nutrient poor soils. However, it is clearly an inferior competitor because of its smaller proportions and shorter life span, and it gets overgrown by tall scrub (and trees). At least in De Westhoek dunes, active management (removal of tall scrub and trees) is required in order to preserve this habitat type.
Nomenclature
Lambinon et al. 2012
References
Bartish IV, Kadereit JW, Comes HP (2006) Late Quaternary history of Hippophae rhamnoides L. (Elaeagnaceae) inferred from chalcone synthase intron (Chsi) sequences and chloroplast DNA variation. Molec Ecol 15:4065–4083
Benjamin K, Domon G, Bouchard A (2005) Vegetation composition and succession of abandoned farmland: effects of ecological, historical and spatial factors. Landscape Ecol 20:627–647
Binggeli P, Eakin M, Macfadyen A, Power J, McConnel (1992) Impact of the alien sea buckthorn (Hippophae rhamnoides L.) on sand dune ecosystems in Ireland. In Carter RWG, Curtis TGF, Sheehy-Skeffington MJ (eds) Coastal dunes; geomorphology, ecology and management for conservation. Balkema, Rotterdam, pp. 325–337
Cosyns E, Provoost S, Zwaenepoel A (2013) Evaluatie van het gevoerde beheer en opmaak van een beheerplan voor het VNR De Duinen en Bossen van De Panne. Wvi & INBO, Brugge
Curt T, Prévosto B, Klesczewski M, Lepart J (2003) Post-grazing Scots pine colonization of mid-elevation heathlands: population structure, impact on vegetation composition and diversity. Ann Forest Sci 60:711–724
De Bruyne C (1905) Over onze duinflora. Handelingen van het 9de Vlaamsch Natuur- en Geneeskundig Congres, Aalst
Decuyper M, Van den Dool R, Slim PA, Kuiters AT, Jansen JM, Sass-Klaassen U (2020) Population dynamics of Hippophae rhamnoides shrub in response of sea-level rise and insect outbreaks. PLOS One 15: e0233011
Delaunois H (1952) De Zeeduinen van de Westhoek. Nat Stedenschoon 25:91–110
Depuydt F (1967) Bijdrage tot de geomorfologische en fytogeografische studie van het domaniaal reservaat De Westhoek. Ministerie van landbouw, Bestuur van waters en bossen, Dienst domaniale natuurreservaten en natuurbescherming, werken nr 3, Brussel
D'Hondt A (1981) De vegetatie van de Westhoek, een fytosociologische studie van het staatsnatuurreservaat ten behoeve van het beheer, met vegetatiekaart. Laboratorium voor Morfologie, Systematiek en Ecologie van de Planten, Ghent University
Dutoit T, Alard D (1995) Mécanisme d’une succession végétale secondaire en pelouse calcicole: une approche historique. Compt Rend Acad Sci Sér 3 Sci Vie 318:897–907
European Commission (2007) The interpretation manual of European Union habitats – EUR27. European Commission DG Environment, Brussels
Fenner F, Radcliffe FN (1965) Myxomatosis. Cambridge University Press, Cambridge
Galatowitsch S (2012) Ecological restoration. Sinauer, Sunderland
Gardner IC, Clelland DM, Scott A (1984) Mycorrhizal improvement in non-leguminous nitrogen-fixing associations with particular reference to Hippophae rhamnoides L. Pl & Soil 78:189–199
Glenn-Lewin DC, Peet RK, Veblen TT eds (1992) Plant succession: theory and prediction. Chapman & Hall, London
Good JEG, Bryant R, Carlill P (1990) Distribution, longevity and survival of upland hawthorn (Crataegus monogyna) scrub in North Wales in relation to sheep grazing. J Appl Ecol 27:272–283
Herbauts J (1971) Flore et végétation des dunes de la Réserve Naturelle domaniale du Westhoek. Ministerie van Landbouw, Bestuur van Waters en Bossen, Werken nr 5, Brussel
Hodgkin SE (1984) Scrub encroachment and its effects on soil fertility on Newborough Warren, Anglesey, Wales. Biol Conservation 29:99–119
Hoffmann M, Cosyns E, Lamoot I (2005) Large herbivores in coastal dune management: do grazers do what they are supposed to do? In Herrier J-L, Mees J, Salman A, et al. (eds) Proceedings ‘Dunes and Estuaries 2005’ – International Conference on Nature Restoration Practices in European Coastal Habitats. VLIZ Special Publication 19, pp. 249–267
Houston J (2008a) Management of Natura 2000 habitats: 2130* Fixed coastal dunes with herbaceous vegetation (‘grey dunes’). European Commission, Brussels
Houston J (2008b) Management of Natura 2000 habitats: 2190 Humid dune slacks. European Commission, Brussels
Hudjetz S, Lennartz G, Krämer K, Roß-Nickoll M, Gergs A, Preuss TG (2014) Modeling wood encroachment in abandoned grasslands in the Eifel National Park, model description and testing. PLOS One. 9:e113827
Huston M, Smith T (1987) Plant succession- life history and competition. Amer Naturalist 130:168–198
Isermann M, Diekmann M, Heemann S (2007) Effects of the expansion by Hippophae rhamnoides on plant species richness in coastal dunes. Appl Veg Sci 10:33–42
Jia D-R, Abbott RJ, Liu T-L, Mao K-S, Bartish IV, Liu J-Q (2012) Out of the Qinghai-Tibet Plateau: evidence for the origin and dispersal of Eurasian temperate plants from a phylogeographic study of Hippophae rhamnoides (Elaeagnaceae). New Phytol 194:1123–1133
Jones MLM, Sowerby A, Williams DL, Jones RE (2008) Factors controlling soil development in sand dunes: evidence from a coastal dune soil chronosequence. Pl & Soil 307:219–234
Kollmann J, Reiner SA (1996) Light demands of shrub seedlings and their establishment within scrublands. Flora 191:191–200
Lambinon J, De Langhe J, Delvosalle L, Duvigneaud J (2012) Flora van België, het Groothertogdom Luxemburg, Noord-Frankrijk en de aangrenzende gebieden. Botanic Garden Meise, Brussels
Leten M, Van Nieuwenhuyse H, Herrier J-L (2005) Invasive scrub and trees in the coastal dunes of Flanders (Belgium): an overview of management goals, actions and results. In Herrier J-L, Mees J, Salman A, Seys J, Van Nieuwenhuyse H, Dobbelaere I (eds) Proceedings Dunes and Estuaries 2005. International conference on nature restoration practices in European coastal habitats. VLIZ Special Publication 19, Oostende, pp 111–127
Lichter J (2000) Colonization constraints during primary succession on coastal Lake Michigan sand dunes. J Ecol 88:825–839
Maas PWT, Oremus PAI, Otten H (1982) Longidorus sp and Tylenchorhynchus microphasmis on Hippophae rhamnoides in a coastal sand dune area. Nematologica 28:158–158
Magda D, Chambon-Dubreuil E, Agreil C, Gleizes B, Jarry M (2009) Demographic analysis of a dominant shrub (Cytisus scoparius): prospects for encroachment control. Basic Appl Ecol 10:631–639
Massart J (1912) La cinquantième herborisation générale de la Société Royale de Botanique de Belgique sur le littoral Belge. Bull Soc Roy Bot Belgique 51:1–185
Meltzer JA (1941) Die Sanddorn-Liguster Assoziation. Ned Kruidk Arch 51:385–395
Mortimer SR et al. (2000) The nature conservation value of scrub in Britain. Report 308, Joint Nature Conservation Committee, Peterborough
Olff H, Huisman J, van Tooren B (1993) Species dynamics and nutrient accumulation during early succession in coastal sand dunes. J Ecol 81:693–706
Oliver CD, Larson BC (1996) Forest stands dynamics, Update edition. FES Other Publications 1, 520 pp
Olson JS (1958) Rates of succession and soil changes on southern Lake Michigan sand dunes. Bot Gaz 119:125–170
Oremus PAI, Otten H (1981) Factors affecting growth and nodulation of Hippophae rhamnoides L ssp. rhamnoides in soils from 2 successional stages of dune formation. Pl & Soil 63:317–331
Pearson MC, Rogers JA (1962) Hippophae rhamnoides L. J Ecol 50:501–513
Petit-Berghem Y (2004) La gestion conservatoire des dépressions tourbeuses intra-dunales: l’exemple des dunes du Nord de la France. Géocarrefour 79:277–284
Poschlod P, Bakker JP, Kahmen S (2005) Changing land use and its impact on biodiversity Basic Appl Ecol 6:93–98
Prévosto B, Kuiters L, Bernhardt-Römermann M, Dölle M, Schmidt W, Hoffmann M, Van Uytvanck J, Bohner A, Kreiner D, Stadler J, Klotz S, Brandl R (2011) Impacts of land abandonment on vegetation: successional pathways in European habitats. Folia Geobot 46:303–325
Provoost S, Ampe C, Bonte D, Cosyns E, Hoffmann M (2004) Ecology, management and monitoring of grey dunes in Flanders. J Coastal Conservation 10:33–42
Provoost S, Jones MLM, Edmondson SE (2011) Changes in landscape and vegetation of coastal dunes in northwest Europe: a review. J Coastal Conservation 15:207–226
Provoost S, Leten M (2012) Het Westhoekreservaat: historisch en landschapsecologisch kader. In Berquin H (ed) In het zand geschreven. De duinen van de Westhoek: een geschiedenis. Lier, pp 259–292
Provoost S, Van Gompel W, Vercruysse E (2020) Beheerevaluatie kust. Eindrapport 2015–2019. Report 2020.18, Research Institute Instituut for Nature and Forest, Brussels
Rahir E (1932) Dunes et bois de La Panne, parc national. La fédération nationale pour la défense de la nature, Bruxelles
Richards EG, Burningham H (2011) Hippophae rhamnoides on a coastal dune system: a thorny issue? J Coastal Conservation 15:73–85
Rogers JA (1961) The autecology of Hippophae rhamnoides L. Unpublished PhD thesis, University of Nottingham
Rozema J, Laan P, Broekman R, Ernst WHO, Appelo CAJ (1985) On the lime transition and decalcification in the coastal dunes of the province of North Holland and the island of Schiermonnikoog. Acta Bot Neerl 34:393–411
Sevink J (1991) Soil development in the coastal dunes and its relation to climate. Landscape Ecol 6:49–56
Skogen A (1972) The Hippophae rhamnoides alluvial forest at Leinora, central Norway. Kongel Norske Vidensk Selsk Foerh 4:1–115
Sloet van Oldenruitenborgh CLM (1976) Duinstruwelen in het deltagebied. Veenman and zonen, Wageningen
Smit C, Bakker ES, Apol MEF, Olff H (2010) Effects of cattle and rabbit grazing on clonal expansion of spiny shrubs in wood-pastures. Basic Appl Ecol 11:685–692
Stace CA, van den Mejiden R, de Kort I (2005) Interactive flora of NW Europe. ETI Bioinformatics, Amsterdam
Stewart WDP, Pearson MC (1967) Nodulation and nitrogen-fixation by Hippophae rhamnoides L. in the field. Pl & Soil 26:348–360
Tadych M., Blaszkowski J (2000) Arbuscular fungi and mycorrhizae (Glomales) of the Slowinski National Park, Poland. Mycotaxon 74:463–482
Temperton VM, Hobbs RJ, Nuttle T, Halle S (2004) Assembly rules and restoration ecology: bridging the gap between theory and practice. Island Press, Washington DC
Tilman D (1990) Constraints and tradeoffs: toward a predictive theory of competition and succession. Oikos 58:3–15
Troelstra SR, Blacquiere T, Wagenaar R, Vandijk C (1987) Ionic balance, proton efflux, nitrate reductase-activity and growth of Hippophae rhamnoides L ssp. rhamnoides as influenced by combined N-nutrition or N2-fixation. Pl & Soil 103:169–183
Tutin TG et al. (1964) Flora Europea. Cambridge University Press, Cambridge
Van der Hagen HGJM, Assendorp D, Calame W, van der Meulen F, Sýkora KV, Schaminée JHJ (2020) Is livestock grazing a key factor for changing vegetation patterns in lime rich coastal dunes in the Netherlands? J Coastal Conservation 24:15
Van der Heijden EW (2001) Differential benefits of arbuscular mycorrhizal and ectomycorrhizal infection of Salix repens. Mycorrhiza 10:185–193
Van der Maarel E, Boot R, van Dorp D, Rijntjes J (1985a) Vegetation succession on the dunes near Oostvoorne, the Netherlands – a comparison of the vegetation in 1959 and 1980. Vegetatio 58:137–187
Van der Maarel E, De Cock N, De Wildt E (1985b) Population dynamics of some major woody species in relation to long-term succession on the dunes of Voorne. Vegetatio 61:209–219
Vroege PW, Stelleman P (1990) Insect and wind pollination in Salix repens L and Salix caprea L. Israel J Bot 39:125–132
Weeda EJ, Westra R, Westra C, Westra T (1985) Nederlandse oecologische Flora. Wilde planten en hun relaties 1. IVN, Amsterdam
Weeda EJ, Westra R, Westra C, Westra T (1987) Nederlandse oecologische Flora. Wilde planten en hun relaties 2. IVN, Amsterdam
Weeda EJ, Westra R, Westra C, Westra T (1988) Nederlandse oecologische Flora. Wilde planten en hun relaties 3. IVN, Amsterdam
Zhao W, Goebel C, Cardina J (2013) Temporal and spatial pattern of a privet (Ligustrum vulgare) invasion. Invasive Pl Sci Managem 6:310–319
Zoon FC (1995) Biotic and abiotic soil factors in the succession of sea buckthorn, Hippophaë rhamnoides L. in coastal sand dunes. Unpublished PhD thesis, Univerity of Wageningen
Zoon FC, Troelstra SR, Maas PWT (1993) Ecology of the plant-feeding nematode fauna associated with Sea Buckthorn (Hippophae-rhamnoides L-ssp. rhamnoides) in different stages of dune succession. Fundam Appl Nematol 16:247–258
Acknowledgements
We thank the Flemish Agency for Nature and Forests, the site manager for the research permission and the support. Thanks also go to Jan Cuvelier for help with the heavy fieldwork and to Maurice Hoffmann and Jan Van Uytvanck for their constructive comments on the manuscript.
Author information
Authors and Affiliations
Contributions
The author(s) read and approved the final manuscript.
Corresponding author
Ethics declarations
Competing interests
The authors declare that they have no competing interests.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
About this article

Cite this article
Provoost, S., Declerck, L. Early scrub development in De Westhoek coastal dunes (Belgium). Folia Geobot 55, 315–332 (2020). https://doi.org/10.1007/s12224-020-09385-1
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s12224-020-09385-1