
Abstract
Transformation of coppices to high forests has caused fundamental changes in site conditions and a decline of many species across Central Europe. Nevertheless, some formerly coppiced forests still harbour a number of the declining species and have become biodiversity hotspots in the changing landscape. We focused on the best-preserved remnant of formerly grazed and coppiced subcontinental oak forest in the Czech Republic – the Dúbrava forest near the town of Hodonín. To improve our understanding of the ecology of declining species, we studied local habitat requirements of vascular plants most endangered at the national level. We recorded vegetation composition and sampled important site variables in plots with the largest populations of endangered species and in additional plots placed randomly across all major forest habitats. We demonstrated that sites with endangered species have a highly uneven distribution in ecological space and that their species composition is often similar to open-canopy oak forests. Within this habitat, the endangered species are concentrated in places with a high light availability and high soil pH. Light-demanding species characteristic of subcontinental oak forests are the best indicators of these sites, while broadly distributed shade-tolerant and nutrient-demanding species avoid them. These results support the view that the occurrence of many endangered species in the Dúbrava forest is a legacy of the long history of traditional management that kept the canopies open. Light-demanding species are now threatened by ongoing successional changes. Therefore, active conservation measures are recommended, including opening up the canopies, early thinning of young stands, control of expansive and invasive species and understorey grazing or mowing.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Historical forms of management such as coppicing, grazing or litter raking have shaped the appearance of European forests and the composition of their biotic communities for centuries (Rackham 2003). Since the eighteenth century these management forms have been abandoned across Europe, and recently they have practically vanished, particularly in Western and Central Europe (Müllerová et al. 2014). Transformation of most forest stands to high forests or plantations of economically important species has brought fundamental changes in their site conditions and caused a decline of many species (Baeten et al. 2009; Naaf and Wulf 2010; Müllerová et al. 2015). Nevertheless, some formerly coppiced forests – particularly lowland oak forests – still harbour populations of declining species and have thus become biodiversity hotspots and the subject of conservation efforts in the changing landscape.
Even in protected areas and at other sites designated for biodiversity conservation, successional changes are taking place, either as a result of management abandonment or other recent issues such as increased nitrogen deposition and high ungulate densities (Hofmeister et al. 2004; Ward 2005; Milner et al. 2006; Verheyen et al. 2012, Brunet et al. 2016). To facilitate survival of the remaining populations of endangered species, targeted management measures have to be designed and incorporated into management plans of particular forest sites. However, any effective management plan requires sufficient knowledge of the current state of the endangered populations and their habitat requirements.
Our study focused on one of the largest and best-preserved remnants of formerly coppiced lowland oak forests in the Czech Republic – the Dúbrava forest near the town of Hodonín. Regarding the number and population sizes of the endangered vascular plant species, it is probably the most valuable single oak forest in the country. To improve our understanding of the ecology of endangered vascular plant species and to design an effective management plan, we studied the habitat requirements of these species and compared them to the site conditions in a broad spectrum of habitats in the forest. Specifically, we hypothesized that (1) endangered species are not dispersed evenly across the vegetation continuum but concentrated in particular habitat types, mostly those with a species composition resembling formerly coppiced oak forests, and (2) habitats with endangered species differ from other habitats regarding environmental characteristics.
Material and methods
Study area
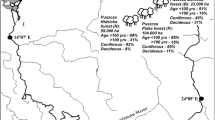
The study area is situated within the large lowland Dúbrava forest (100 km2). It is located north-west of the town of Hodonín in the south-easternmost part of the Czech Republic (48.9° N, 18.1° E) at an altitude between 160–240 m a.s.l. (Fig. 1 ). The climate of the region is subcontinental, with mean July temperatures 19.8 °C, mean January temperatures −1.5 °C and a mean annual rainfall 585 mm (Vesecký 1961). The terrain is flat, mildly undulating, with low dunes and shallow depressions. It is formed by base- and nutrient-poor Pleistocene eolian sands overlaying Tertiary impermeable base-rich sediments. This combination supports a fine-scale mosaic of dry and wet, base-rich and base-poor sites (Novák and Pelíšek 1943; Šmarda 1961). The actual study area is approximately 15 km2 in size.

Position of vegetation plots sampled in the study area. The forest is segmented into management units of different age, structure and tree species composition (grey lines). Empty circles (N = 47) indicate context plots placed randomly across eight major forest habitats. Full circles (N = 35) indicate endangered species plots, i.e. plots with the largest populations of the endangered species.
The past use of the forest was diverse and included wood pasture, hay making and coppicing (Šmarda 1961; Řepka 2009; Szabó 2013). These practices have been gradually abandoned until the first half of the twentieth century, when clear-cutting and extensive planting of pine (Pinus sylvestris) started to prevail. However, patches of overgrown and largely singled-out oak coppices have been preserved in many places. They are mostly dominated by Quercus robur and form a rare type of subcontinental oak forest that is classified as the Carici fritschii-Quercetum roboris association (Chytrý and Horák 1997; Roleček 2007, 2013) and listed as priority habitat 91I0 (Euro-Siberian steppic woods with Quercus spp.) in Annex 1 of Council Directive 92/43/EEC. Locally shade-tolerant broad-leaved trees are admixed, mainly Tilia cordata. Wet depressions are often treeless or covered with stands of Salix aurita and Betula spp. Stands dominated by Alnus glutinosa occur along streams.
Out of more than 650 plant species identified in the study area, 9 are listed as critically endangered and 20 as strongly endangered in the Czech Red List (Grulich 2012). The site is protected under EU legislation as a Site of Community Importance (CZ0624070 – Hodonínská doubrava) and under Czech legislation as a National Nature Monument (NPP Hodonínská Dúbrava).
Sampling design
The analysed dataset consisted of two parts. The first one included 35 plots from sites with endangered plant species. We mapped distribution of 20 species most endangered from the national perspective. The species were selected based on the current version of the Czech Red List (Grulich 2012) and consultation with its author (V. Grulich pers. comm.). The selection included 9 critically endangered species and 11 strongly endangered species (Table 1 ). We sampled two sites with the largest populations of each of the 20 species (one site was sampled in case only a single population of a species was found). We assumed that the largest populations indicate habitats with site conditions most suitable for a particular species. At each site, the sampling plot (these plots are further referred to as ‘endangered species plots’) was positioned in the place with the highest population density or close to the centre of the local population.
The other dataset was sampled to provide context for the endangered species plots and to cover the variation in vegetation composition and site conditions across the study area. We used Natura 2000 habitat maps (NCA CR 2011) to stratify vegetation, and randomly placed a total number of 47 plots (further called ‘context plots’) among the strata, proportionally to the common logarithm of the total area of each habitat and with a minimum number of two plots per habitat (Table 2 ).
Vegetation sampling
Plant species composition was recorded in circular plots of one meter radius in both endangered species plots and context plots. All vascular plant species in the herb layer were recorded and the cover-abundance of each species was visually estimated on a 9-point Braun-Blanquet scale (Dengler et al. 2008). Vegetation samples were recorded in June to August 2012–2014. Species nomenclature follows Kubát et al. (2002), with the exception of Hieracium onegense (Norrl.) Norrl.
Sampling and measuring of site variables
Soil samples were taken from the upper part (5 cm) of the mineral soil horizon in each plot. Samples were dried at room temperature and sieved. Total carbon and inorganic nitrogen were measured in a 0.1 mm fraction using Carlo Erba NC 2500 elemental analyser. Available phosphorus was determined colorimetrically in mixed samples prepared with a 1-M solution of sodium bicarbonate at pH 8.5. Contents of Ca2+, Mg2+ and K+ cations were determined from mixed samples in a Mehlich III extraction. Soil acidity was assessed by shaking a suspension of 20 g of soil and 50 ml of distilled water and subsequently measuring the pH using a glass electrode.
Light availability in the understorey was estimated as canopy openness measured by means of hemispherical photography using a Nikon FC-E8 fish-eye lens in the centre of each plot. Canopy openness values were calculated in the Sidelook programme. Here, the automatic thresholding algorithm (Nobis and Hunziker 2005) was used to distinguish black and white pixels.
To assess soil moisture, soil water content was measured in all plots in August 2014 during two days without rain. We used a Delta-T HH2 device with a ThetaProbe ML2x probe, which determines the volumetric soil water content by measuring the impedance of the probe inserted in upper 5 cm of soil (Gaskin and Miller 1996).
To estimate long-term soil wetness driven by topography, we used the convergence index as implemented in SAGA GIS (Conrad et al. 2015). This index assesses terrain convexity in the surroundings of the focal site; negative values indicate terrain depressions whereas positive values indicate elevations. For its calculation we used a detailed Digital Elevation Model (DEM) created by LIDAR technology with 5-m pixel resolution (DEM 4G provided by the Czech Office for Surveying, Mapping and Cadastre). Slope orientations of DEM pixels up to 25 m from the focal site were considered and we used inverse distance weighting of pixel values to give local topography a higher importance. We employed the convergence index instead of the more widely used topographic wetness index (Kopecký and Čížková 2010) because the topography of the study area is generally flat. Soil moisture patterns are therefore probably driven rather by local terrain shape than by regional flow accumulation.
To determine stand age class (10-years span) corresponding to each vegetation plot, we used forestry maps.
Data analysis
To show the position of endangered species plots in a broad vegetation context, non-metric multidimensional scaling (NMDS) in R (R core team 2014) operated from the JUICE 7.0 programme (Tichý 2002) was performed on the merged dataset of endangered species plots and context plots. Logarithmically transformed percentage cover data was used and rare species were downweighted in the analysis. Only plots with at least three species were included (76 plots in total).
To assess habitat requirements of the endangered species, we restricted the dataset to vegetation plots sampled in open-canopy oak forests, their margins and undisturbed clearings, i.e. the community that prevails in the study area and is protected here. We performed a two-sample Mann-Whitney test from the coin package (Hothorn et al. 2006) in R to test whether endangered species plots significantly differ from context plots in the observed values. Statistical significance was evaluated using a randomization test based on 999 permutations. Characteristic species of the endangered species plots and other habitats were determined using the Φ coefficient of fidelity with group size standardized to 10% of the dataset (Tichý and Chytrý 2006). Species with a Φ value ≥ 0.25 were considered characteristic.
Results
According to the NMDS results, endangered species plots are clearly concentrated in the part of the ordination space associated with the vegetation of open-canopy oak forests (Fig. 2 ). Several endangered species plots are located near the centroids of wetland vegetation (reed and tall-sedge beds, alder carrs, alluvial forests) whereas none is associated with shady oak-hornbeam forests, disturbed clearings and broad-leaved or coniferous plantations.

NMDS plot showing the position of vegetation plots with the largest populations of the endangered species in the context of all major vegetation types in the study area. Endangered species plots are represented by full circles, context plots by empty circles. Triangles represent centroids of plots belonging to one vegetation type. All open-canopy oak forest types (Pannonian thermophilous oak forests on sand, wet acidophilous oak forests and acidophilous oak forests on sand; see also Table 2 ) were included into oak forests. The plot shows that the largest populations of the most endangered species are concentrated in open-canopy oak forests.
Regarding site conditions within the open-canopy oak forest, endangered species plots are characterized by higher light availability in the understorey and higher soil pH (Table 3 ). Besides, these plots have a higher herb layer cover, are more species-rich and have higher numbers of other endangered species. However, when numbers of endangered species are relativized by the total number of species or herb layer cover in the analysed plots, the significance of the differences between the endangered species plots and context plots decreases (Z = 2.233, P = 0.023 or Z = 1.3739, P = 0.1768, respectively).
Endangered species plots have higher numbers of characteristic species than context plots (Table 4 ). Many of these species are also characteristic of the endangered species plots in open-canopy oak forests (Table 5 ).
Discussion
Habitat requirements of the endangered species
We showed that the largest populations of the most endangered species in the former coppice under study are conspicuously concentrated in open-canopy oak stands on relatively base-rich soils. Even within open-canopy oak stands, endangered species prefer sites with above-average light availability such as canopy gaps, road margins and undisturbed clearings. These findings are in agreement with the knowledge of light requirements of the species under study, whose mean Ellenberg indicator value for light is 6.8 whereas none of the species has a value lower than 5 (Table 1 ). On this scale, value 7 is attributed to plants of well-lit places which also occur in partial shade, while value 5 represents semi-shade plants (Ellenberg et al. 1992).
Also species characteristic of the endangered species plots (e.g. Potentilla alba, Carex fritschii, Geranium sanguineum, Galium boreale and Anthericum ramosum) are light-demanding to semi-shade plants, many of which are diagnostic species of subcontinental oak forests of the Carici fritschii-Quercetum roboris association (Chytrý and Horák 1997; Roleček 2007). On the contrary, broadly distributed shade-tolerant and nutrient-demanding forest species such as Poa nemoralis, Mycelis muralis and Moehringia trinervia tend to avoid vegetation with the most endangered species. The results of the characteristic species analysis within open-canopy oak forests reflect the preference of the most endangered species for base-rich soils (Table 5 ). While endangered species plots are characterized by the presence of diagnostic species of subcontinental oak forests (i.e. a relatively basophilous vegetation type; Roleček 2013), the common acidotolerant grasses Anthoxanthum odoratum and Agrostis capillaris show preference for sites without endangered species, often classified as acidophilous oak forests.
However, our interpretation of the higher number of other endangered species (excluding the target species for which the plots were sampled) in endangered species plots remains cautious, as detailed analysis showed that the observed relationship may simply result from the higher number of plant individuals in these plots.
Generally, our results suggest that species endangered at the national scale may be considered endangered also in Dúbrava forest, because they are concentrated at favourable sites (high light availability, base-rich soils), which are prone to successional changes, including spread of shading trees and invasive herb species (see below for details).
Our results are in agreement with the research conducted in Swedish forests (Berg et al. 1994, 1995), where the majority of endangered vascular plant species occupies semi-shady (52 %) and open (25 %) habitats, in spite of the prevalence of shady habitats in the study area. In the same study, abiotic factors were found generally more important for endangered plants than for other organism groups compared (cryptogams, invertebrates, vertebrates), with high soil calcium content being the most important factor. This also agrees with our results. The habitat requirements of endangered species observed in our study further correspond to the ecological traits characteristic of critically endangered species in the Czech Republic in general (Gabrielová et al. 2013): They are mostly competitively inferior species confined to open, less productive habitats on base-rich soils.
Historical roots of the endangered diversity
Our results support the view that the abundant occurrence of endangered species is a legacy of the long history of traditional management in Dúbrava forest. This management kept the canopies open and created site conditions suitable for light-demanding species. Besides indirect evidence from written sources describing past forest use (Šmarda 1961; Szabó 2013), we find support for this hypothesis also in old botanical literature. Formánek (1887) characterized the occurrence of the now critically endangered Gladiolus palustris at Dúbrava as ‘rather abundant in open oak scrub and on small forest meadows’. Hruby (1926) documented the open forest-steppe character of some parts of the forest with a photograph of a park-like stand with scattered oak standards, low scrub and open grasslands with the light-demanding grass Stipa capillata. The strikingly open character of many oak stands is moreover visible on aerial photographs of the area from the 1950s (http://kontaminace.cenia.cz; last accessed 29 July 2015).
It is notable that some of the studied endangered species (e.g. Carex buxbaumii, C. fritschii, Festuca amethystina, Gladiolus palustris and Thalictrum simplex subsp. galioides) are widely scattered in large parts of their distributional ranges. This pattern may result from the dynamics of their distribution during the Holocene, which should also be considered when interpreting their occurrence at Dúbrava. It is generally accepted that many ancient light- and base-demanding species, broadly distributed in Central Europe during the early Holocene, retreated from this region later on for different reasons, including (i) the mid-Holocene forest optimum acting as a bottleneck for light-demanding species (Lang 1994; Ložek 2007; Pokorný et al. 2015); (ii) natural acidification of the environment (Iversen 1958; Pokorný and Kuneš 2005) and (iii) various human-induced changes including direct habitat destruction and alteration of site conditions (Wood et al. 2000). However, some of the retreating species have survived at some sites and regions, thus their recent occurrences may be considered relicts of a former broader distribution (Grandcolas et al. 2014). It has been suggested that the Dúbrava forest and its surroundings are such a long-term refugium for some of the endangered species studied (Roleček et al. 2015). This may particularly apply to species belonging to ancient species pools of central European forest-steppe (Chytrý 2012) and oligotrophic calcareous wetlands (Sádlo 2000). This hypothesis is supported by studies providing multi-proxy evidence for the persistence of open forest-steppe habitats during the mid-Holocene forest maximum in the south-easternmost Czech Republic (Hájková et al. 2011; Kuneš et al. 2015). Similarly, open wetlands in this area show a long-term continuity (Rybníček 1983). A historical explanation may also apply to the observed preference of the endangered species for a high soil pH, as the ancient central European species pools have their origin in base-rich Pleistocene environments (Pärtel 2002; Chytrý et al. 2003; Ewald 2003).
It may seem paradoxical that a forest could be a refugium for light-demanding species. However, as presented above, the history of European woods is variegated (Vera 2000; Rackham 2003; Kirby and Watkins 2015). We believe that natural and human-induced disturbances kept spaces open for light-demanding species from different taxonomic groups for centuries or even millennia, even if the composition of the tree layer fluctuated substantially (as evidenced for Dúbrava by pollen analysis, see Jamrichová et al. 2013). Only in the last centuries the situation changed, with decline of traditional forest management and broad-scale environmental change being the major causes (Kwiatkowska and Wyszomirski 1988; Jakubowska-Gabara 1996; Müllerová et al. 2015). The Dúbrava forest has also been affected by this development (Konvicka et al. 2008; Řepka 2009), but, probably thanks to its extent, diversity of site conditions and hardly changed tree layer structure in some parts, it has preserved the extraordinary richness in declining species. After all, continuity may be the key to its unique natural value.
Recent vegetation changes and recommended conservation measures
As already mentioned above, the oak forest vegetation in Dúbrava forest has experienced substantial changes in the past decades and this development seems to have accelerated recently. The observed changes are mainly connected to intensive forest management in some stands and absence of management in others, but also to broad-scale phenomena such as plant invasions and high game density. Based on the results of our research and relevant literature data (Konvička et al. 2004; Konvicka et al. 2008; Řepka 2009; Hédl et al. 2010; Jamrichová et al. 2013; Vild et al. 2013; Müllerová et al. 2015), we have attempted to estimate the main threats to the endangered species populations at Dúbrava and to recommend measures to preserve them (Table 6 ). Although further studies are needed to assess the role of different threats and the effect of different conservation measures, we believe that our recommendations may aid the nature conservation authorities to create an effective management plan for this valuable site, as well as other sites of lowland open-canopy oak forests.
References
Baeten L, Bauwens B, De Schrijver A, De Keersmaeker L, Van Calster H, Vandekerkhove K, Roelandt B, Beeckman H, Verheyen K (2009) Herb layer changes (1954–2000) related to the conversion of coppice-with-standards forest and soil acidification. Appl Veg Sci 12:187–197
Berg Å, Ehnström B, Gustafsson L, Hallingbäck T, Jonsell M, Weslien J (1994) Threatened plant, animal, and fungus species in Swedish forests: distribution and habitat associations. Conservation Biol 8:718–731
Berg Å, Ehnström B, Gustafsson L, Hallingbäck T, Jonsell M, Weslien J (1995) Threat levels and threats to red-listed species in Swedish forests. Conservation Biol 9:1629–1633
Brunet J, Hedwall P-O, Holmström E, Wahlgren E (2016) Disturbance of the herbaceous layer after invasion of an eutrophic temperate forest by wild boar. Nordic J Bot 34:120–128
Chytrý M (2012) Vegetation of the Czech Republic: diversity, ecology, history and dynamics. Preslia 84:427–504
Chytrý M, Horák J (1997) Plant communities of the thermophilous oak forests in Moravia. Preslia 68:193–233
Chytrý M, Kučera T, Kočí M, Grulich V & Lustyk P (eds) (2010) Katalog biotopů České republiky. Ed. 2 (Habitat catalogue of the Czech Republic. Ed. 2). AOPK ČR, Prague
Chytrý M, Tichý L, Roleček J (2003) Local and regional patterns of species richness in Central European vegetation types along the pH/calcium gradient. Folia Geobot 38:429–442
Conrad O, Bechtel B, Bock M, Dietrich H, Fischer E, Gerlitz L, Wehberg J, Wichmann V, Böhner J (2015) System for Automated Geoscientific Analyses (SAGA) v. 2.1.4. Geosci Model Developm 8:1991–2007
Dengler J, Chytrý M, Ewald J (2008) Phytosociology. In Jørgensen SE, Fath BD (eds) Encyclopedia of ecology. Elsevier, Oxford, pp 2767–2779
Ellenberg H, Weber HE, Düll R, Wirth V, Werner W, Paulissen D (1992) Zeigerwerte von Pflanzen in Mitteleuropa. Scripta Geobot 18:1–248
Ewald J (2003) The calcareous riddle: Why are there so many calciphilous species in the Central European flora? Folia Geobot 38:357–366
Formánek E (1887) Květena Moravy a rakouského Slezska (Flora of Moravia and Austrian Silesia). Edvard Formánek, Brno
Gabrielová J, Münzbergová Z, Tackenberg O, Chrtek J (2013) Can we distinguish plant species that are rare and endangered from other plants using their biological traits? Folia Geobot 48:449–466
Gaskin GJ, Miller JD (1996) Measurement of soil water content using a simplified impedance measuring technique. J Agric Eng Res 63:153–159
Grandcolas P, Nattier R, Trewick S (2014) Relict species: a relict concept? Trends Ecol Evol 29:655–663
Grulich V (2012) Red List of vascular plants of the Czech Republic, 3rd edition. Preslia 84:631–645
Hájková P, Roleček J, Hájek M, Horsák M, Fajmon K, Polák M, Jamrichová E (2011) Prehistoric origin of the extremely species-rich semi-dry grasslands in the Bílé Karpaty Mts (Czech Republic and Slovakia). Preslia 83:185–204
Hédl R, Kopecký M, Komárek J (2010) Half a century of succession in a temperate oakwood: from species-rich community to mesic forest. Diversity & Distrib 16:267–276.
Hofmeister J, Mihaljevič M, Hošek J (2004) The spread of ash (Fraxinus excelsior) in some European oak forests: an effect of nitrogen deposition or successional change? Forest Ecol Managem 203:35–47
Hothorn T, Hornik K, van de Wiel MA, Zeileis A (2006) A Lego system for conditional inference. Amer Statistician 60:257–263
Hruby J (1926) Göding in Mähren und seine Umgebung. Čas Morav Zemsk Mus Brno 24:60–97
Iversen J (1958) The bearing of glacial and interglacial epochs on the formation and extinction of plant taxa. Uppsala Univ Årsskr 6:210–215
Jakubowska-Gabara J (1996) Decline of Potentillo albae-Quercetum Libb. 1933 phytocoenoses in Poland. Vegetatio 124:45–59
Jamrichová E, Szabó P, Hédl R, Kuneš P, Bobek P, Pelánková B (2013) Continuity and change in the vegetation of a Central European oakwood. The Holocene 23:46–56
Kirby K, Watkins C (eds) (2015) Europe’s changing woods and forests: from wildwood to managed landscapes. CABI, London
Konvička M, Čížek L, Beneš J (2004) Ohrožený hmyz nížinných lesů: ochrana a management (Endangered insects of lowland forests: conservation and management). Sagittaria, Olomouc
Konvicka M, Novak J, Benes J, Fric Z, Bradley J, Keil P, Hrcek J, Chobot K, Marhoul P (2008) The last population of the Woodland Brown butterfly (Lopinga achine) in the Czech Republic: habitat use, demography and site management. J Insect Conserv 12:549–560
Kopecký M, Čížková Š (2010) Using topographic wetness index in vegetation ecology: Does the algorithm matter? Appl Veg Sci 13:450–459
Kubát K, Hrouda L, Chrtek J Jun., Kaplan Z, Kirschner J, Štěpánek J (2002) Klíč ke květeně České republiky (Key to the flora of the Czech Republic). Academia, Praha
Kuneš P, Svobodová-Svitavská H, Kolář J, Hajnalová M, Abraham V, Macek M, Tkáč P, Szabó P (2015) The origin of grasslands in the temperate forest zone of east-central Europe: long-term legacy of climate and human impact. Quatern Sci Rev 116:15–27
Kwiatkowska AJ, Wyszomirski T (1988) Decline of Potentillo albae-Quercetum phytocoenoses associated with the invasion of Carpinus betulus. Vegetatio 75:49–55
Lang G (1994) Quartäre Vegetationsgeschichte Europas. Gustav Fischer Verlag, Jena
Ložek V (2007) Zrcadlo minulosti: česká a slovenská krajina v kvartéru (Mirror of the past: Czech and Slovak landscape in the Quaternary). Dokořán, Praha
Milner JM, Bonenfant C, Mysterud A, Gaillard JM, Csanyi S, Stenseth NC (2006) Temporal and spatial development of red deer harvesting in Europe: biological and cultural factors. J Appl Ecol 43:721–734
Müllerová J, Szabó P, Hédl R (2014) The rise and fall of traditional forest management in southern Moravia: a history of the past 700 years. Forest Ecol Managem 331:104–115
Müllerová J, Hédl R, Szabó P (2015) Coppice abandonment and its implications for species diversity in forest vegetation. Forest Ecol Managem 343:88–100
Naaf T, Wulf M (2010) Habitat specialists and generalists drive homogenization and differentiation of temperate forest plant communities at the regional scale. Biol Conservation 143:848–855
NCA CR (2011) Habitat mapping layer [electronic georeferenced database]. Version 2011. Nature Conservation Agency of the Czech Republic, Prague, Accessed 5 Sept 2011
Nobis M, Hunziker U (2005) Automatic thresholding for hemispherical canopy-photographs based on edge detection. Agric Forest Meteorol 128:243–250
Novák V, Pelíšek J (1943) Stručná charakteristika půd na přesypových pískách v lesní oblasti Dubrava u Hodonína (Brief characteristics of soils on eolian sand in the forest region of Dubrava near Hodonín). Lesn Práce 8:225–235
Pärtel M (2002) Local plant diversity patterns and evolutionary history at the regional scale. Ecology 83:2361–2366
Pokorný P, Kuneš P (2005) Holocene acidification process recorded in three pollen profiles from Czech sandstone and river terrace environments. Ferrantia 44:101–107
Pokorný P, Chytrý M, Juřičková L, Sádlo J, Novák J, Ložek V (2015) Mid-Holocene bottleneck for central European dry grasslands: Did steppe survive the forest optimum in northern Bohemia, Czech Republic? The Holocene 25:716–726
R Core Team (2014) R: a language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria. Available at http://www.R-project.org
Rackham O (2003) Ancient woodland: its history, vegetation and uses in England. Castlepoint Press, Dalbeattie
Řepka R (2009) Druhová diverzita vyšších rostlin versus lesnický management v evropsky významné lokalitě (EVL) Hodonínská Doubrava (Vascular plant species richness versus forestry management in the SCI Hodonínská Doubrava). Zprávy Čes Bot Společn Mater 24:111–120
Roleček J (2007) Formalized classification of thermophilous oak forests in the Czech Republic: What brings the Cocktail method? Preslia 79:1–21
Roleček J (2013) Thermophilous oak forests. Quercetea pubescentis. In Chytrý M (ed) Vegetation of the Czech Republic 4. Forest and scrub vegetation. Academia, Praha, pp 296–337
Roleček J, Hájek M, Karlík P, Novák J (2015) Reliktní vegetace na mezických stanovištích (Relict vegetation on mesic sites). Zprávy Čes Bot Společn 50:201–245
Rybníček K (1983) The environmental evolution and infilling process of a former lake near Vracov (Czechoslovakia). Hydrobiologia 103:247–250
Sádlo J (2000) Původ travinné vegetace slatin v Čechách: sukcese kontra cenogeneze. Preslia 72:495–506
Šmarda F (1961) Rostlinná společenstva území přesypových písků lesa Doubravy u Hodonína (Plant communities of the eolian sand area in Doubrava forest near Hodonín). Práce Brněnské základny Českoslov Akad věd 413:1–56
Szabó P (2013) The end of common uses and traditional management in a Central European wood. In Rotherham ID (ed) Cultural severance and the environment. The ending of traditional and customary practice on commons and landscapes managed in common. Springer Netherlands, Dordrecht, pp 205–213
Tichý L (2002) JUICE, software for vegetation classification. J Veg Sci 13:451–453
Vera FWM (2000) Grazing ecology and forest history. CABI, New York
Verheyen K, Baeten L, De Frenne P, Bernhardt-Römermann M, Brunet J, Cornelis J, Decocq G, Dierschke H, Eriksson O, Hédl R, Heinken T, Hermy M, Hommel P, Kirby K, Naaf T, Peterken G, Petřík P, Pfadenhauer J, Van Calster H, Walther G-R, Wulf M, Verstraeten G (2012) Driving factors behind the eutrophication signal in understorey plant communities of deciduous temperate forests J Ecol 100:352–365
Vesecký A (1961) Podnebí Československé socialistické republiky. Tabulky (Climate of the Czechoslovak Socialist Republic. Tables). Hydrometeorologický ústav, Praha
Vild O, Roleček J, Hédl R, Kopecký M, Utinek D (2013) Experimental restoration of coppice-with-standards: response of understorey vegetation from the conservation perspective. Forest Ecol Managem 310:234–241
Ward AI (2005) Expanding ranges of wild and feral deer in Great Britain. Mammal Rev 35:165–173
Wood A, Stedman-Edwards P, Mang J (eds) (2000) The root causes of biodiversity loss. Earthscan, London
Acknowledgements
Our thanks go out to Martin Kopecký for calculation of the convergence index, Martina Fabšičová, Pavel Unar, Vladan Riedl and Martin Kopecký for help with vegetation sampling within project IAA600050812, Radim Hédl for kindly providing soil chemistry data acquired within project IAA600050812, David Zelený for the discussion regarding ordination methods and Jan W. Jongepier for improving our English. This paper was produced as part of the project ‘Coppice forests as the production and biological alternative for the future’ (No. CZ.1.07/2.3.00/20.0267) with financial contribution of the EC and the state budget of the Czech Republic. Besides, the research leading to these results received funding from the European Research Council under the European Union’s Seventh Framework Programme (FP7/2007–2013) / ERC Grant agreement No. 278065, and the long-term research project RVO 67985939 from the Czech Academy of Sciences.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article

Cite this article
Roleček, J., Vild, O., Sladký, J. et al. Habitat requirements of endangered species in a former coppice of high conservation value. Folia Geobot 52, 59–69 (2017). https://doi.org/10.1007/s12224-016-9276-6
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s12224-016-9276-6