
Abstract
Holoparasite Orobanche gracilis Sm. and O.fuliginosa Reut. ex Jord. were micromorphologically and anatomically examined. In anatomical studies, the transverse and surface section of the root, stem and leaf of species were examined to determine interspecific variation. Studied species have non-photosynthetic scale leaves. There are glandular and eglandular trichomes in the leaf and stem of studied species. The glandular trichomes are capitate type in O. gracilis and capitate and peltate types in O. fuliginosa. The seed surface types are reticulate. The depth of anticlinal walls are medium or remarkable in the seed of O. gracilis and remarkable in the seed of O. fuliginosa.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
1 Introduction
Orobanchaceae consists of 90 genera and 1800 species all over the world (Nickrent 2010). While the genus Orobanche is represented worldwide by approximately 170 species (Uhlich et al. 1995), the genus is represented by 39 species in Turkey (Özhatay and Kültür 2006; Yıldırımlı 2008; Zare et al. 2009). Orobanche species are obligate parasitic flowering plants. These species are parasitic on a wide range of plant families such as Asteraceae, Fabaceae, Solanaceae, Apiaceae, Cucurbitaceae (Linke et al. 1989; Goldwasser et al. 2000). Orobanche is divided into four sections: Trionychon Wallr, Orobanche Wallr, Gymnocaulis Nutt. and Myzorrhiza (Philippi) Beck. (Beck von Mannagetta 1930; Gilli 1971; Foley 2001; Pujadas 2002). Trionychon and Orobanche are the big sections. The most studied species belong to section Orobanche (Gilli 1971). Some authors have recognised these sections as separate genera, but the classification of this genus is still under discussion (Schneeweiss 2001, 2007). According to Pamphilis and Palmer (1990), the species belonging to Orobanche are devoid from photosynthetic leaves. Orobanche is a genus widely investigated in agricultural studies (Antonova 1996; Matusova et al. 2004; Skoric et al. 2010) because, as obligate root holoparasite, it causes damages to the cultures.
It is known that species with similar morphological aspects, can reveal significant differences in the anatomical structures of the vegetative organs such as root, stems and leaf. Accordingly, in recent years, anatomical characters have been used in taxonomy (Kharazian 2007). Concerning the anatomical structure of the parasitic plants, a weak-developed phloem and a well-developed xylem can be observed in the vascular bundles (Khan et al. 2009). Moreover the anatomical study of root is very important to understand the host-parasite relationship (Goldwasser et al. 2000). For the Orobanche the root structure is well developed because the species are full parasitic plant. Features of seed morphology and micromorphology have been also widely used to distinguish the different taxa (Plaza et al. 2004). In particular the structure of periclinal and anticlinal walls of seed coat represents an important taxonomic character (Barthlott 1976) which strongly contributes to the characterization of species (Plaza et al. 2004).
The aim of this study is to evaluate comparatively the anatomical and micromorphological features of the fully parasitic plants O. fuliginosa and O. gracilis species.
2 Materials and methods
Materials were collected from the following localities of Black Sea Region in Turkey. Samples for anatomical studies were fixed in 70 % alcohol. The transverse and surface sections of root, stem and leaves were excised by hand and they were covered with glycerin-gelatin (Vardar 1987). Anatomic measurements were made using imaging software. For morphometric analysis more than 20 measurements were performed for all anatomical parts and the samples collected from different localities were evaluated separately. Photographs were taken with Nicon Eclipse Ni-U Microscope. Stomatal index was calculated according to the method described by Meidner and Mansfield (1968). For scanning electron microscopy (SEM), dried leaves and seeds (herbarium specimens) were mounted on stubs using double-sided carbon tape and coated with 12.5–15 nm gold in a sputter coater. Coated leaves and seeds were examined and photographed with JMS-6060 LV SEM. Statistical analyses were performed using package software SPSS 22. The terminology of seed surface sculpturing basically follows Stearn (1992) and Plaza et al. (2004).
- Orobanche fuliginosa :
-
Turkey, Samsun, Çarşamba, 17 m, Akbulut, 13.
Turkey, Samsun, Bafra, Cernek Lake, 1 m, Akbulut,14.
- Orobanche gracilis :
-
Turkey, Tokat, Borabay Lake, 1050 m, Akbulut, 11.
Turkey, Ordu, Unye, Sarıhalil village, 500 m, Akbulut, 12.
3 Results
3.1 The anatomical properties
O. fuliginosa: In the transverse section of the root, remnant of the epidermis was visible, the cortex was multilayered and the xylem extended to the centre of the stele, covering 48 % of the section (Fig. 1a) The xylem was composed of cells with thick walls (Fig. 1a).

Transverse and surface section of O. fuliginosa (a–e), a root, b stem, c cross-section of the leaf, d leaf surface (upper epidermis), e leaf surface (lower epidermis). Transverse and surface section of O. gracilis (f–j), f root, g stem, h cross-section of the leaf, i leaf surface (upper epidermis), j leaf surface (lower epidermis). pe periderm, c cortex, ph phloem, x xylem, e epidermis, p pith, ue upper epidermis, le lower epidermis, vb vascular bundle, h hairs, s stomata (scale bar 100 µm)
In the transverse sections of the stem the layer of the epidermis with oval- rectangular cells was observed. (Fig. 1b). The cortex appeared multi-layered and consisted of oval or polygonal cells (Fig. 1b). The pith covered 74 % of the section and consisted of large and cylindrical parenchymatic cells (Fig. 1b). The xylem and phloem elements were clear. Trichomes were observed on the stem (Fig. 2a–d).

Glandular hairs in the leaf and stem surfaces. O. fuliginosa (a–d), a four cells, b three cells, c multicellular, d unicellular (peltate). O. gracilis (e–g), e multicellular, f three cells, g multicellular
In the transverse section of the leaf a 1–2 μ cuticle was present on leaf surfaces. Epidermis was single layered (Fig. 1c). The upper and lower epidermal cells were different in size and rectangular and orbicular in shape, respectively (Fig. 1c–e; Table 1). Species has non-photosynthetic scale leaves. Stomata index was 17.24 for upper surface of the leaf (Table 2) while the abaxial surface did not exhibit any stomata. Capitate (3–6 cells) and unicellular glandular trichomes were present on the adaxial surface of the leaf (Fig. 2a–d).
O. gracilis: In the transverse section of the root the periderm appeared multi-layered and the phelloderm was clearly detectable. The cortex was 4–5 layered and parenchymatic and the xylem extended to the centre of the stele covering 61 % of transverse section (Fig. 1f).
In the transverse section taken from the middle part of the stem epidermis was composed of a single row oval cells (Fig. 1g; Table 1). The cortex was multilayered and consisted of polygonal cells whereas the pith consisted of large parenchymatic cells and covered 34 % of the cross section of the stem. (Fig. 1g). The vascular bundles were arranged in a ring. There were multicellular glandular trichomes on the epidermis (Fig. 2e–g).
O. gracilis have non-photosynthetic scale leave and in the transverse section epidermis was formed by a single cell row on both lower and upper surface (Fig. 1h–j). The vascular bundles were collateral. There were very few stomata on abaxial surface of the leaf (Fig. 3h). Stomata index was 11.11 for the upper epidermis (Table 2). There were capitate trichomes with 3–6 cells. The anatomical measurements of both O. fuliginosa and O. gracilis are reported in Table 1.

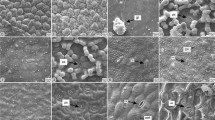
SEM micrographs of seed and leaf. O. fuliginosa (a–e), a, b upper epidermis, c lower epidermis, d, e seed surface. O. gracilis (f–j), f, g upper epidermis, h lower epidermis, i, j seed surface. gh glandular hairs, s stomata, e epidermis, aw anticlinal wall, pw periclinal wall, nt narrow trough (scale bar 100 µm)
3.2 The micromorphological properties
O. fuliginosa: As previously described the cells of leaf epidermis were rectangular or irregular. The upper epidermal cells were more sinuate in comparison to the lower epidermis and the anticlinal walls were sinuate to curved. The walls were more sunken than the other parts of cell. The stomata were present on the adagia surface (Fig. 3c). SEM images showed that dense glandular and eglandular trichomes were present especially on upper surface (Fig. 3a, b).
The seed of O. fuliginosa had ovoid to ellipsoid shape (Fig. 3d; Table 3) and its surface type was reticulate (Fig. 3e). The anticlinal and periclinal cell walls of the seed coat were clearly visible and significantly higher than other parts of the cell. The depth of anticlinal walls was remarkable (Fig. 3e; Table 3). Narrow trough was on all the anticlinal wall (Fig. 3e).
O. gracilis: The epidermal cells of upper and lower leaf surfaces, generally rectangular, showed straight or curved anticlinal walls. The stomata, generally present on the adaxial surface were on same level or upper than epidermal cells and exhibited a long pore (Fig. 3h). The cuticula layer had plication. Capitate glandular trichomes were present on the upper surface of the leaf (Fig. 3f, g).
O. gracilis had ellipsoid to oblongoid seed shape (Fig. 3i; Table 3) and its surface type was reticulate (Fig. 3j). The anticlinal and periclinal cell walls of the the seed coat were clearly visible. The walls was above from the other parts of cell. Depth of the anticlinal walls was medium or remarkable (Table 3; Fig. 3j). Narrow trough was only remarkable on edges of anticlinal wall (Fig. 3j).
4 Discussion
This paper presents a comparative anatomical and micromorphological study of holoparasite O. gracilis and O. fuliginosa species. Orobanche are obligate root holoparasites and they damage the crops. Holoparasitic plants lack chlorophyll and cannot complete their life-cycle without exploiting a suitable host (Khan et al. 2009).
According to Watson and Dallwitz (1991) primary vascular tissue comprises a ring of bundles, or two or more rings of bundles. In studied species, the vascular bundles were arranged in a ring. The cortex was larger in O. fuliginosa than in O. gracilis. According to Pamphilis and Palmer (1990), Orobanche species are devoid of the leaves, while Watson and Dallwitz (1991) reported that some of them have reduced leaves. In this research, both the studied species have non-photosynthetic scale leaves. In agreement with Khan et al. (2009) which reported that in all parasitic plants, vascular bundles are scarcely developed, in the vascular bundles of investigated species the phloem was very reduced but the xylem was slightly more developed.
Watson and Dallwitz (1991) stated that stomata are usually present even if sometimes incompletely developed. According to Holm et al. (1997), the functions of stomata are reduced or almost lost in species of Orobanche. Stomata were very reduced in the abaxial and adaxial surface of the studied species. Metcalfe and Chalk (1979) noted that there are anomocytic stomata in the Orobanche species. Our findings were similar with Metcalfe and Chalk (1979). There are glandular trichomes in leaf and stem of studied species. The glandular trichomes were capitate type with 3–6 cells in O. gracilis and capitate (3–6 cells) and peltate (unicellular) types in O. fuliginosa.
Seed of Orobanche are very small. The use of SEM allows the observation of microstructures and many authors pointed out the importance of SEM for investigating the seed coats and fruit surface (Plaza et al. 2004; Akçin 2007; 2008). Plaza et al. (2004) evidenced that the seed characters are useful for the identification of most of the studied Orobanche species. They determined four morphological seed types in Orobanche genus and reported that the seed shape of O. gracilis was ovoid-subglobose (Plaza et al. 2004). In our study, the seed shape of O. gracilis was ellipsoid to oblongoid and the seed sizes were 0.17–0.39 × 0.30–0.40 mm. Any information was not found about seed of O. fuliginosa. In the present study, we found that O. fuliginosa have ovoid to ellipsoid seed shape and the seed sizes were 0.20–0.42 × 0.21–0.33 mm, therefore bigger than those of O. gracilis. Morewer the narrow trough of O. fuliginosa was on all the anticlinal wall while it was only remarkable on edges of anticlinal wall in O. gracilis. Both species had reticulate seed surfaces and the depth of anticlinal walls in O. fuliginosa were remarkable than O. gracilis.
In conclusion both anatomical and above all micromorphological features revealed a clear distinction between the two investigated species.
References
Akçin ÖE (2007) Nutlets micromorphology of some Onosma L. (Boraginaceae) species from Turkey. Biologia 62(6):684–689
Akçin ÖE (2008) Seed coat and fruit surface micromorphology of some Cynoglossum L. (Boraginaceae) species. Bangladesh J Bot 37(2):115–119
Antonova TS (1996) The role of peroxidase in the resistance of sunflower against Orobanche cumana in Russia. Weed Res 36:113–121
Barthlott W (1976) Morphologie der Samen von Orchideen im Hinblick auf taxonomische und funktionelle Aspekte. In: Senghas K (ed) Proceedings of the 8th World Orchid Conference. Frankfurt, Hamburg, Parey, pp 444–455
Beck von Mannagetta G (1930) Orobanchaceae. In: Engler A (ed) Das Pflanzenreich, vol IV. Verlag von Wilhelm Engelmann, Leipzig, pp 1–348
Foley MJY (2001) Orobanchaceae in the “Flora iberica” area: new taxa, excluded taxa, and typifîcation. Anales Jard Bot Madr 58(2):223–233
Gilli A (1971) Die Orobanchaceen der Türkei. Feddes Rep 82(6):384–406
Goldwasser Y, Plakhıne D, Kleıfeld Y, Zamskı E, Rubın B (2000) The differential susceptibility of vetch (Vicia spp.) to Orobanche aegyptiaca. Anatomical studies. Ann Bot 85:257–262
Holm L, Doll J, Holm E, Pancho J, Herberger J (1997) World weeds-natural histories and distribution. Wiley, pp 511–530
Khan MA, Sharıf T, Ahmad M, Zafar M, Tareen RB (2009) Anatomıcal characterization of parasitic plants of Pakıstan. Pak J Bot 41(6):2661–2669
Kharazian N (2007) The taxonomy and variation of leaf anatomical characters in the genus Aegilops L. (Poaceae) in Iran. Turk J Bot 31:1–9
Linke KH, Sauerborn J, Saxena MC (1989) Orobanche field guide. International center of Agricultural Research in the Dry Areas. Food Legume Improvement Program, p 42
Matusova R, Van Mourık T, Bouwmeester J (2004) Changes in the sensitivity of parasitic seeds to germination stimulants. Seed Sci Res 14:335–344
Meidner H, Mansfield TA (1968) Physiology of stomata. McGraw-Hill, London
Metcalfe CR, Chalk L (1979) Anatomy of dicotyledonsi. Oxford University Press, London, p 276
Nickrent DL (2010) The parasitic plant connection. http://www.parasiticplants.siu.edu. Accessed 1 June 2010
Özhatay N, Kültür Ş (2006) Check-list of additional taxa to the supplement flora of Turkey III. Turk J Bot 30:281–316
Pamphılıs CW, Palmer JD (1990) Loss of photosynthetic and chlororespiratory genes from the plastid genome of a parasitic flowering plant. Nature 348:337–339
Plaza L, Fernandez I, Juan R, Pastor J, Pujadas A (2004) Micromorphological studies on seeds of Orobanche species from the Iberian Peninsula and the Balearic Islands, and their systematic significance. Ann Bot 94:167–178
Pujadas A (2002) Orobanche L. In: Lopez JA, Catalaan P, Saez LI (eds) Plantas parasitas de la Peninsula Iberica y Baleares. Mundi-Prensa, Madrid
Schneeweiss GM (2001) Relationships within Orobanche sect. Tryonychon: insights from its sequences markers. In: Fer A, Thalouarn P, Joel DM, Musselman LJ, Parker C, Verkleij JAC (eds). Proceeding of the 7th International Parasitic Weed Symposium, Nantes, France, 49–52
Schneeweiss GM (2007) Correlated evolution of life history and host range in the nonphotosynthetic parasitic flowering plants Orobanche and Phelipanche (Orobanchaceae). J Evol Biol 20:471–478
Skorıc D, Joıta MP, Sava E (2010) Sunflower breeding for resistance to broomrape (Orobanche cumana Wallr.). An. I.N.C.D.A. Fundulea, vol. LXXVIII, (1): 63–79
Stearn WT (1992) Botanical latin. David and Charles, London
Uhlich H, Pusch J, Barthel KJ (1995) Die Sommerwurzarten Europas
Vardar Y (1987) Botanikte Preparasyon Teknikleri. Ege Üniversitesi Fen Fakültesi Baskı İşleri, İzmir
Watson L, Dallwitz MJ (1991) The families of flovering plants. http://delta-intkey.com/angio/www/orobanch.htm. Accessed 07 June 2011
Yıldırımlı Ş (2008) The chorology of the Turkish species of Oleaceae, Onagraceae, Orobanchaceae and Oxalidaceae families. Ot Sistematik Botanik Dergisi 15(2):151–166
Zare G, Dönmez AA, Koca AD (2009) A new record for the flora of Turkey: Orobanche palaestina Reut. (Orobanchaceae). Hacet J Biol Chem 38(2):149–154
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article

Cite this article
Akbulut, M.K., Akçin, Ö.E. & Şenel, G. Anatomical and micromorphological properties of holoparasite Orobanche gracilis and Orobanche fuliginosa (Orobanchaceae) species. Rend. Fis. Acc. Lincei 27, 565–571 (2016). https://doi.org/10.1007/s12210-016-0522-x
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s12210-016-0522-x