
Abstract
This study investigated the safety, efficacy, and immunological influence of allogeneic umbilical cord-derived mesenchymal stromal cells (IMSUT-CORD) processed in serum-free medium and cryoprotectant, for treating steroid-resistant acute graft-versus-host disease (aGVHD). In a phase I dose-escalation trial, IMSUT-CORD were infused intravenously twice weekly over two cycles with up to two additional cycles. Four patients received a dose of 1 × 106 cells/kg, while three received 2 × 106/kg. Of 76 total adverse events, fourteen associated or possibly associated adverse events included 2 cases of a hot flash, headache, and peripheral neuropathy, 1 each of upper abdominal pain, hypoxia, increased γ-GTP, somnolence, peripheral vascular pain at the injection site, thrombocytopenia, hypertension, and decreased fibrinogen. At 16 weeks after the initial IMSUT-CORD infusion, three patients showed complete response (CR), two partial response (PR), one mixed response, and one no response. The overall response rate was 71.4%, and the continuous CR/PR rate was 100% for over 28 days after CR/PR. NK cell count significantly increased and correlated with treatment response, whereas IL-12, IL-17, and IL-33 levels decreased, but did not correlate with treatment response. CCL2 and CCL11 levels increased during IMSUT-CORD therapy. IMSUT-CORD are usable in patients with steroid-resistant aGVHD (UMIN000032819: https://www.umin.ac.jp/ctr).
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Acute graft-versus-host disease (aGVHD) occurs in 30–70% of patients undergoing allogeneic hematopoietic stem cell transplantation (HSCT) [1, 2], even after intensive prophylaxis with immunosuppressive reagents. Although severe aGVHD is first treated with steroids, steroid-resistant aGVHD represents a major cause of mortality after allogeneic HSCT; therefore, effective therapies remain warranted [3].
Although bone marrow (BM)-derived mesenchymal stromal cells (BM-MSCs) are beneficial as a salvage treatment against treatment-resistant severe aGVHD following allogeneic HSCT [4,5,6,7,8], BM-MSC harvesting is an invasive procedure that may cause hemorrhage, infection, or chronic pain. Furthermore, BM-MSCs exhibit accelerated senescence with increased donor age [9]. To overcome these limitations, perinatal tissue-derived MSCs, including those from cord blood (CB), placenta, and umbilical cord (UC) [10], have been investigated. We have previously examined the utilization of UC as an abundant source of MSCs for immunotherapy [11, 12]. We developed an efficient method to cryopreserve the whole UC tissue [13] and obtain master cells from frozen-thawed UC tissue using an improved explant method [14] and expanded the UC-MSCs (IMSUT-CORD) from master cells in GMP-grade processing. Processing and cryopreservation were conducted using reagents including serum-free medium and serum-free cryoprotectant. We provide a proof of concept using the serum-free manufactured UC-MSCs with an anti-inflammatory property illustrated using an allogeneic mixed lymphocyte reaction (MLR) and xenogeneic GVHD model [15]. UC-MSCs showed a rapid and high proliferative ability and HLA-DR negativity even in the presence of Interferon (IFN)-γ compared with BM-MSCs [12, 16].
Herein, we report the results of a clinical trial of serum-free manufactured IMSUT-CORD for patients with first-line treatment-resistant severe aGVHD following allogeneic HSCT, while considering safety, efficacy, and immunological influences.
Materials and methods
Manufacturing UC-MSCs (IMSUT-CORD)
UC collection, storage, and release including the safety and quality management were approved by the Ethics Committee of The Institute of Medical Science, The University of Tokyo (IMSUT) hospital (IRB#33-2, REC#29-28) and the individual collection hospitals (NTT Medical Center Tokyo IRB#16-14 and Yamaguchi hospital IRB#33-2). After obtaining informed consent from the donor’s mother, the UC was collected at birth, transferred in an in-house cool box to the Cord Blood and Cord Bank of the Research hospital, The Institute of Medical Science at The University of Tokyo (the bank name is IMSUT CORD) [10]. Infectious disease tests were conducted using the mother’s blood and cord blood. The health assessment of both the infant and mother was followed for more than 6 months after birth to ensure the safety of UC. UC was manufactured into the product cells, designated IMSUT-CORD in accordance with the GMP standard [10]. Briefly, UC tissue was cut into 1 cm long and frozen in serum-free cryoprotectant (Stem-cellbanker; ZENOAQ Resource Co. Ltd., Fukushima, Japan) within 48 h after birth [13]. After comparative assessments of four donors derived UC, a UC derived from one donor with a relatively high growth rate and good immunosuppressive potency was selected as a master cell candidate. Then, we performed a viral safety evaluation of the master cells; the selected donor-derived master cells were sufficient to be expanded into the product cells (IMSUT-CORD) for implementation. Master cells were obtained from frozen-thawed UC tissues using an improved explant method [14], and cryopreserved again. The qualified master cells were thawed and further expanded in a large-scale Corning CellSTACK culture chambers (Corning Inc. NY) using a GMP-compatible in-house serum-free medium (Rohto Pharmaceutical Co. Ltd., Japan) [15]. Expanded product cells were suspended in an in-house, serum-free cryoprotectant comprising 10% DMSO (Cryoserv, Mylan GmbH, Switzerland), dextran 40 (Low molecular dextran D injection; Otsuka Pharmaceutical factory, Inc. (Otsuka), Japan), and bicarbonate solution (Bicanate injection; Otsuka, Japan), and were frozen using a controlled-rate freezer (Thermo Fisher Scientific, 7473, MA, USA) to suppress latent heat, and cryopreserved in a liquid nitrogen tank until use. The product cells (IMSUT-CORD), were packaged in 16 mL, which contained at least 64 × 106 viable MSCs after thawing. More than 2 weeks after the completion of the cryopreservation process, quality tests of frozen-thawed IMSUT-CORD were performed.
Analysis of surface markers in IMSUT-CORD
Flow cytometry was performed as described previously[15]. The cells were labeled with monoclonal antibodies listed in the Supplementary Table 1. The cells were acquired using BD™FACSCanto II flow cytometer (BD) and analyzed using FlowJo software (BD). To assess the influence of IFN-γ stimulation on IMSUT-CORD, the expression of HLA-DR, CD80, CD86, PD-L1, and PD-L2 of IMSUT-CORD were examined using flow cytometry in the absence or presence of IFN-γ at the dose of 100 ng/mL.
Analysis of inhibitory effect of allogeneic MLR and phytohemagglutinin-L (PHA-L) stimulation by IMSUT-CORD
Inhibitory effect of allogeneic MLR [12, 15] and PHA-L stimulation was analyzed by co-culture of UC-MSCs. In MLR, 5-(and -6)-carboxyfluorescein diacetate succinimidyl ester (CFSE) (Vybrant CFDA SE Cell Tracer Kit; Invitrogen)-labeled peripheral blood-mononuclear cells (MNCs) were used as responder (R) cells. PMDC05 cells were kindly provided by Dr. Narita at the Faculty of Medicine, Niigata University [17] and used as stimulator (S) cells following irradiation. The principal of the CFSE-stained R cells in MLR assay and 10 μg/mL of PHA-L stimulation were conducted as previously described [12, 15]. On the day of MLR, R and S cells co-cultured with or without IMSUT-CORD were mixed. After 4 days of culture, the CFSE fluorescence intensities of R, T cells in the CD4+ and CD8+ gate were analyzed using FACS Canto II and FlowJo software (Supplementary table 1). The immunosuppressive effects of UC-MSCs were compared based on the blockade of the daughter cell peaks. Briefly, the parental intensity of the un-proliferated CFSE histogram gate was the standard line, which is indicated as M1, and subsequent gates (M2, M3, M3, ….) of the proliferated daughter cells were manually set to achieve approximately two-fold progressive reductions in CFSE intensity (Fig. 1C). The inhibition ratio (%) was indicated as the ratio of each division index; R + S + UC-MSCs and R + S (as 100%), or R + PHA-L + UC-MSCs and R + PHA-L (as 100%) as described previously [15, 16].

Quality test results of frozen-thawed IMSUT-CORD for the clinical trial. A Morphological features of IMSUT-CORD before harvesting the cells. B Surface markers of IMSUT-CORD. A representative flow cytometry histogram image of 10 products, processed individually; the table shows mean ± SD % of the products (n = 10). C Inhibitory effect of allogeneic mixed lymphocyte reaction (MLR) and phytohemagglutinin-L (PHA-L) stimulation by co-culture of UC-MSCs. Mean ± SD % of inhibition of MLR and PHA-L stimulation by IMSUT-CORD is shown (n = 10). D Factors in the supernatant of allogeneic mixed lymphocyte reaction (MLR) with or without IMSUT-CORD. Supernatant concentration of IFN-γ, TNF-α, and prostaglandin E2, with or without IMSUT-CORD co-culture; mean ± SD (n = 10). Statistical analysis is shown. E Influence of IFN-γ stimulation on IMSUT-CORD. Expression of HLA-DR, CD80, CD86, PD-L1, and PD-L2 of IMSUT-CORD examined using flow cytometry in the absence or presence of IFN-γ and shown as mean ± SD of % positivity. Fold expression of indoleamine 2,3-dioxygenase 1 (IDO-1) of IMSUT-CORD normalized by glyceraldehyde-3-phosphate dehydrogenase (GAPDH) with or without IFN-γ stimulation, determined using quantitative real-time PCR (n = 10)
Analysis of IFN-γ, TNF-α, and prostaglandin E2 concentrations in the supernatant of allogeneic MLR co-cultured with IMSUT-CORD
Supernatant concentrations of IFN-γ, and TNF-α with or without IMSUT-CORD co-culture, were analyzed using the cytokine beads assay (BD CBA Human Th1/Th2/Th17 Cytokine Kit, BD) according to the manufacturer’s instructions. The beads were acquired using FACS Canto II flow cytometer and analyzed using the FCAP Array v3.0 software (BD). Prostaglandin E2 expression was analyzed using ELISA (Prostaglandin E2 Parameter Assay Kit, R&D systems, MN) according to the manufacturer’s instructions.
Quantitative transcription-polymerase chain reaction (RT-qPCR) analysis
Fold change in indoleamine 2,3-dioxygenase 1 (IDO-1) expression in IMSUT-CORD normalized to glyceraldehyde-3-phosphate dehydrogenase (GAPDH) expression with or without IFN-γ stimulation at the dose of 100 ng/mL was determined using RT-qPCR (n = 10). Primers used were listed in supplementary Table2. Relative quantification was performed using the comparative cycle threshold (Ct) (2−ΔΔCt) method, where ΔΔCt = mean ΔCt (target gene) – mean ΔCt (housekeeping gene). Details of RT-qPCR conditions were previously reported [12].
Trial design and patients
In this dose escalation, open-label, phase I clinical trial, patients were enrolled from July 2018 to June 2020 at three sites. This trial, registered as the Umin Clinical Trial Registry (UMIN-CTR: UMIN000032819), was conducted in accordance with the principles of the Declaration of Helsinki and Good Clinical Practice guidelines after the approval of each Institutional Review Board (IRB #29-32, 30-21, 2020-1) and regulatory authority. Written informed consent was obtained from all patients.
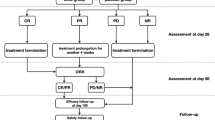
aGVHD stage/grade was defined according to the 1994 Consensus Conference on Acute GVHD grading criteria [18]. In response to aGVHD, a complete response (CR) was defined as the complete resolution of aGVHD; partial response (PR) as a decrease in organ stage or grade; no response (NR) as no change in grade; progression (PG) as progressive worsening in grade; and mixed response (MR) as a mixture of a decrease and increase in organ stages of aGVHD. Steroid-refractory aGVHD was defined as the PG of aGVHD for 3 days with standard-dose steroid administration or NR after 5 days of therapy. Twenty- to seventy-year-old standard steroid therapy-resistant patients with Grade II to IV aGVHD after HSCT for hematological malignancies resistant to standard steroid-containing therapy were eligible. Patients not treated with standard steroid therapy for aGVHD, who were not in remission at HSCT except for myelodysplastic syndrome with or without leukemic transition and myeloproliferative diseases, with severe complications of vital organs tested positive for viral infections or were allergic to gentamicin and amphotericin B, utilized in the processing and culture, or DMSO, were excluded. The concentrations of gentamicin and amphotericin B are lower than the molecular weight in the final product because they were used during early processing and diluted; therefore, as instructed by the Japanese regulatory authorities, we used this as an exclusion criterion.
Cryopreserved IMSUT-CORD was rapidly thawed in 37 °C water bath and drawn into the syringe and slowly injected intravenously without washing and dilution over 10 min. Before IMSUT-CORD injection, hydrocortisone and chlorpheniramine, were infused to prevent infusion-related reactions. IMSUT-CORD was administered twice per week (designated as one cycle) in two cycles, and patients who showed PR or MR with no severe adverse events were allowed to undergo two additional cycles. Dose escalation was conducted following the 3 + 3 dose-escalation scheme. Doses were as follows: cohort 1, 1 × 106 cells/kg body weight (BW); cohort 2, 2 × 106 cells/kg BW; cohort-1, 0.5 × 106 cells/kg BW. The initial dose was 1 × 106 cells/kg BW (cohort 1), while the subsequent dose was 2 × 106 cells/kg BW (cohort 2), when there was no Dose-limiting toxicity (DLT) in cohort1.
To monitoring the adverse events associated with IMSUT-CORD injection, laboratory studies, viral tests, electrocardiogram (ECG), echocardiography were done according to the schedule. Chest X-ray, computed tomography, and bone marrow aspiration test were performed, if necessary. Vital signs, including ECG monitoring, percutaneous oxygen saturation concentration (SpO2 levels), and blood pressure, were monitored from the IMSUT-CORD injection to two hours after injection.
The primary endpoint was safety, and the secondary endpoint was efficacy, including the overall response and change of stage/grade of organs damaged by aGVHD. Adverse events and laboratory test results were evaluated until 11 weeks after the final IMSUT-CORD administration. DLT was assessed within 7 days of the final injection of IMSUT-CORD according to the Common Terminology Criteria for Adverse Events (CTCAE) ver. 4.0-JCOG http://www.jcog.jp/doctor/tool/ctcaev5.html: (1) CTCAE grade 4 neutropenia with < 5% blastic cells in the BM; (2) non-hematologic toxicity ≥ grade 3 (controllable infusion reaction excluded); (3) and requirement of > 7 days for the next administration process due to adverse events. Chronic GVHD (cGVHD) was evaluated during a long-term follow-up according to NIH consensus criteria [19].
Patient blood test and immune cell analysis
Immunological studies were conducted, separate from the clinical trial, after obtaining written informed consent and approval from each Institutional Review Board.
The absolute immune cell counts, comprising CD3+T, CD3−CD56+natural killer (NK), CD3+CD4+T, CD3+CD8+T, CD19+B, and CD4+CD25+regulatory T (Treg) cells, in the peripheral blood were measured by counting white blood cells stained directly with antibodies in a BD Trucount™ tube containing beads for quantification (BD). Briefly, 100 μL of blood, antibodies, and 7-AAD was added into a Trucount tube and red blood cells were lysed using 1 mL of Pharm Lyse (BD). After staining cells according to reagent instructions, the cells were acquired and analyzed using FACSCanto II flow cytometer to quantify immune cells.
Cytokine and chemokine assay in patient blood
We evaluated human cytokine and chemokine concentrations in frozen-thawed plasma collected on the day of IMSUT-CORD administration (before administration), and 1, 2, 3, 4, 6, 8, and 11 weeks after final administration. Cytokines and chemokines were analyzed using a Cytometric bead array; Human Inflammation Panel (13-plex; BioLegend, USA) and Human Proinflammatory Chemokine Panel (13-plex; BioLegend), respectively. All samples were analyzed in triplicate, according to the manufacturer’s instructions. Bead fluorescence readings were obtained using FACS Canto II. Data were analyzed using LEGENDplex version 8.0 software (BioLegend).
We also measured C–C motif chemokines, such as chemokine (C–C motif) ligand 2 (CCL)2 and CCL11 levels in the supernatant of MLR co-cultured with UC-MSCs (n = 3) using a Human Proinflammatory Chemokine Panel (13-plex; BioLegend). To exclude the FBS influence and attachment of them, we set the Transwell insert (Corning, MA, USA) between UC-MSCs and MLR and cultured without FBS supplement. The method for MLR setting was the same as described above.
Statistical analysis
The sample size was determined via the trial study design (early phase I safety study with 3 + 3 dose-escalation protocol). Data from patients who dropped out from the study due to additional treatments for aGVHD were included in any analysis when possible. Overall survival was analyzed using a Kaplan–Meier curve. A linear mixed model was used to assess time-dependent changes for some measurements while considering correlation with patients. Spearman’s correlation coefficients were calculated to assess the association between immune cells, cytokines, and chemokines with the treatment response, including the aGVHD grade. Differences between groups were analyzed with JMP 10.0.2 software (SAS Institutes, Cary, NC, USA). Statistical analyses were performed with the Turkey–Kramer test or T test, and P < 0.05 was regarded as statistically significant.
Results
IMSUT-CORD characteristics
IMSUT-CORD were adherent with spindle-shaped morphology (Fig. 1A), positive for CD105, CD73, and CD90 and negative for CD45, CD34, CD11b, CD19, and HLA-DR (Fig. 1B). In an MLR, the triggering of responder T cell proliferation by allogeneic dendritic cells was efficiently inhibited to 8.56 ± 6.57% in CD4-positive cells and 9.87 ± 7.26% in CD8-positive cells via its co-culture with IMSUT-CORD (Fig. 1C). The proliferation of responder cells treated with phytohemagglutinin-L (PHA-L; Sigma-Aldrich) was also inhibited to 16.33 ± 6.31% in CD4-positive cells and 17.14 ± 4.46% in CD8-positive cells (Fig. 1C). Co-culture with IMSUT-CORD in the MLR system was accompanied by a significant reduction in IFN-γ and TNF-α levels as shown using cytokine beads assay and the induction of prostaglandin E2 using ELISA (Fig. 1D). Flow cytometry analysis showed that IMSUT-CORD cells were negative for HLA-DR, CD80, and CD86 even at high concentrations of IFN-γ (Fig. 1E). PD-L2 was constitutively expressed in UC-MSCs, whereas PD-L1 expression was induced upon IFN-γ addition (Fig. 1E, F). The abundant induction of indoleamine 2,3-dioxygenase 1 (IDO-1) in IMSUT-CORD, as one of the key immunoregulators in the presence of IFN-γ was confirmed using quantitative real-time PCR (Fig. 1E). No chromosomal abnormalities were observed in product cells after G-banding analysis.
Patient characteristics and GVHD response
Seven patients were enrolled in this study, and six agreed to participate in immunological studies. Four were treated in cohort 1 and three were treated in cohort 2 (Table 1). The median age was 43 years (range: 25–62 years); three patients were male and four were females. The prophylaxis of aGVHD is also shown in Table 1. All patients were administered prednisolone or methylprednisolone as first-line therapy against aGVHD.
Two patients had grade III aGVHD, whereas five had grade II aGVHD (Table 2). Among the patients with grade III aGVHD, patient 2 demonstrated the following stages of organ involvement: skin, 3; liver, 1; gastrointestinal tract (GI tract), 2. Patient 3 demonstrated the following stages of organ involvement: skin, 1; liver, 3; GI tract, 0. Among patients with grade II aGVHD, patient 1 demonstrated skin, 1; liver, 0; GI tract 1, and the skin alone was affected in three patients, and the GI tract alone affected the remaining patient. In cohort 2, single organ involvement was observed. The median number of days from HSCT to the first administration of IMSUT-CORD was 57 days (range, 41–125 days). Three patients (patients 2, 3, and 6) received the first IMSUT-CORD injection after 100 days of HSCT, due to the late-onset aGVHD during the tapering of GVHD prophylaxis reagents. One patient underwent two cycles of IMSUT-CORD; two three cycles; and four four cycles. In cohort 1, one patient achieved CR, two PR, and one MR after 4 weeks from the first administration. In cohort 2, two patients achieved PR and one NR after 4 weeks from the first administration and the patients with PR reached CR after 12 weeks from the first administration (Table 2). Finally, overall, three patients achieved CR and two achieved PR possessing continuous duration for more than 28 days. All patients except patient 2 were alive during the long-term follow-up period of 327 days (median; range: 104–712 days). The overall survival rate was 85.7% (90% CI 62.2–100) at day 100 (Supplementary Fig. 1).
Long-term follow-up revealed four of six evaluable patients developed cGVHD; one developed cGVHD within the monitoring period of adverse events and the remaining three developed after then. Patient 1 received cholecystectomy for cholecystitis due to gall stone and discontinued PSL after 37 weeks from the first administration of IMSUT-CORD, and liver dysfunction with elevated alkaline phosphatase developed after 49 weeks from the first administration, which improved without any immunosuppressant. Patient 3 had bronchiolitis obliterans (BO) after pneumonia and septic shock caused by Streptococcus Pneumoniae after 36 weeks from the first administration and developed into severe cGVHD with BO and dry eye and mouth controlled with tacrolimus and PSL. Patient 4 persisted skin aGVHD and transited into mild skin cGVHD treated with low-dose steroid therapy after 7 weeks from the first administration. Patient 7 showed cGVDH with dry eye treated after 27 weeks from the first administration.
Safety evaluations
No DLT was found in any patient. Adverse events until 11 weeks after the final IMSUT-CORD administration were observed in all patients and listed in Table 3. Fourteen out of total seventy-six adverse events were either associated or possibly associated with IMSUT-CORD administration. In 3/4 patients in cohort 1, one patient experienced hot flashes (grade 1) and upper abdominal pain (grade 1), one had a hypoxia (grade 1), and one had a headache (grade 1). Patients recovered from these symptoms without medication. In cohort 2, 2/3 patients showed either of the following adverse events: one event of increased γ-glutamine transferase levels (grade 1), hot flashes (grade 1), somnolence (grade 1), peripheral vascular pain at the injection site (grade 1), thrombocytopenia (grade 1), controllable hypertension (grade 3), fibrinogen decreased (grade 2), or headache (grade 2) and two events of peripheral neuropathies (grade 2). Patients recovered from all symptoms except peripheral neuropathy, although it could not be overlooked that this symptom may have been affected by other drugs. No serious adverse events related, or possibly related, to IMSUT-CORD administration were observed.
Immunological response to IMSUT-CORD
To assess the immunological influence of IMSUT-CORD, immunological properties during and after IMSUT-CORD therapy were monitored in six patients except patient 6. Blood samples were obtained on the day of IMSUT-CORD administration (before administration), and 1, 2, 3, 4, 6, 8, and 11 weeks after the final administration. Therefore, the data on the administration of IMSUT-CORD reflect the status before administration or the influence of the prior injections. CD14+ monocytes, CD3−CD56+ NK cells, CD19+ B cells, CD3+CD4+ T cells, and CD3+CD8+ T cells in the peripheral blood were analyzed by flow cytometry (Fig. 2A–E). In patients 1, 2, and 3, the absolute number of monocytes in peripheral blood showed an increasing tendency, whereas that in patients 4, 5, and 7 with skin GVHD showed no change or decreasing tendency during IMSUT-CORD administrations. Overall, there was no significant difference in monocytes change (Fig. 2A, Table 4). Absolute numbers of CD3−CD56+NK cells were significantly increased during IMSUT-CORD therapy in all patients except Patient 2, who developed liver GVHD (Fig. 2B, Table 4). NK cell numbers correlated with treatment response (R2 = 0.498), whereas they were not correlated with aGVHD grade.

Immune cells during and after UC-MSC administration in peripheral blood. A CD14+ monocytes, B CD3−CD56+ NK cells, C CD19+ B cells, D CD3+CD4+T cells, E CD3+CD8+T cells. Data are shown in terms of the absolute number of peripheral blood cells in respective cases. PB, peripheral blood; (n = 6). NS, not significant
IL-12p70, IL-17A, and IL-33 levels were significantly suppressed during IMSUT-CORD therapy compared with those before injection, although they were not correlated with aGVHD grade or treatment response (Table 4). In these few patients, no significant correlation was observed between other inflammatory cytokines and aGVHD grades or therapeutic effects.
Levels of chemokines, such as CCL11 and CCL2, increased during IMSUT-CORD therapy, whereas MIP1α levels decreased. Notably, CCL11 levels were inversely correlated with aGVHD and positively correlated with the treatment response (Table 4).
A clinical course of Patient 3 with grade III aGVHD (skin, 1; liver, 3; GI tract, 0), one of the representative responders to IMSUT-CORD was shown in Fig. 3. He was diagnosed with hepatic GVHD after pathological analysis, accompanied by an increase in liver transferase (ALT, AST, and γ-GTP) and a total bilirubin (Fig. 3A). The liver transferase was decreased after administration of IMSUT-CORD, and correspondingly levels of serum inflammatory cytokines, such as IL-18, IL-17A, IL-23, and IL-33 decreased rapidly after the first administration of IMSUT-CORD (Fig. 3B). Although his total bilirubin was delayed to be decreased after IMSUT-CORD and diagnosed as PR at the end of the observation period, he finally achieved CR with a normal range of total bilirubin on day 174.

Clinical course of patient 03. A Hepatic function test results, including results for AST, ALT, ALP, γ-GTP, and total bilirubin (T bil), and B Inflammatory cytokines, including IFN-γ, IL-8, IL-12p70, IL-17A, IL-18, IL-23, and IL-33, before and after the administration of UC-MSCs
CCL2 and CCL11 secretion in MLR with IMSUT-CORD
To understand the mechanism for the elevation in CCL2 and CCL11 levels in the peripheral blood (PB) of patients during IMSUT-CORD administration (Table 4), we measured the CCL2 and CCL11 levels in the supernatant of MLR co-cultured with UC-MSCs (n = 3) using chemokine beads assay. UC-MSCs constitutively secreted CCL2, which was significantly induced in large quantities upon co-culture with MLR (Fig. 4A). The mean CCL2 levels in the supernatant of MNCs, UC-MSCs, MLR, and MLR co-cultured with UC-MSCs were 74.73, 519.7, 176.33, and 12,167.2 pg/mL, respectively. CCL11 was secreted from the activated MNCs in allogeneic MLR and increased significantly in the supernatant of MLR co-cultured with UC-MSCs (Fig. 4B). The mean CCL11 levels in the supernatant of MNCs, UC-MSCs, MLR, and MLR co-cultured with UC-MSCs were 0, 0, 68.94, and 108.66 pg/mL, respectively.

CCL2 and CCL11 levels in the supernatant of MLR co-cultured with UC-MSCs. A CCL2 levels. B CCL11 levels. CCL2 and CCL11 levels in the supernatant of peripheral blood mononuclear cells (MNCs), umbilical cord-derived mesenchymal stromal cells (UC-MSCs), mixed lymphocyte reaction (MLR), and MLR co-cultured with UC-MSCs. Data are shown as the mean ± SD (pg/mL) for three different donors with triplicate measurements. *P < 0.05
Discussion
Here, we conducted a phase I clinical trial for the treatment-resistant aGVHD with clinical-grade serum-free manufactured IMSUT-CORD. The immunosuppressive properties based on our previous report on UC-MSCs [12] were adopted to the quality tests for shipping the IMSUT-CORD. The demonstrated results showed that each lot of IMSUT-CORD were equivalent and functional. The IMSUT-CORD possessed the characteristics of UC-MSCs. They are characterized by the absence of HLA-DR, CD80, and CD86 markers, even in the presence of IFN-γ, while HLA-DR is induced in BM-MSCs by IFN-γ stimulation as previously reported [12]. The IMSUT-CORD inhibited the proliferation of CD4- and CD8-positive T cells stimulated by allogeneic dendritic cells in a MLR and showed the significant induction of IDO-1 and PD-L1 [15, 16].
Certain BM-MSC-based drugs, such as Remestemcel-L® [20, 21], Prochymal® [20], and Temsel HS Inj® [22,23,24] have been approved in Canada, USA, and Japan, respectively, based on their safety and effectiveness in treatment-resistant aGVHD. Wu et al. [25] first reported that steroid-resistant gastrointestinal and hepatic aGVHD improved dramatically after each of the four infusions of UC-MSCs administered to two patients, without adverse effects. There are not many of clinical trials as the pure treatment with UC-MSCs against aGVHD among many clinical trials, including that for the prevention of graft failure and severe aGVHD in HSCT for patients with aplastic anemia [10, 26, 27]. However, most of MSCs were cultured with xenogeneic serum (FBS)-containing medium. No severe adverse events related, or possibly related, to IMSUT-CORD administration were observed and tolerable for the continuation of the IMSUT CORD administration. To the best of our knowledge, this is the first study to report that serum-free manufactured UC-MSCs; IMSUT-CORD are safe and effective in some patients with steroid-resistant aGVHD.
Long-term follow-up showed that four out of 6 evaluable patients had cGVHD after reaching complete or partial response. One patient with skin aGVHD, which could not obtain CR developed cGVHD, while the remaining patients developed cGVHD after 6 months from IMSUT-CORD therapy when the effect of IMSUT-CORD might have already disappeared. Two of three patients (Patient 01 and 03) had inflammation (Cholecystitis) or infection (Pneumonia and sepsis) events prior development of cGVHD. The development of cGVHD in those patients seemed related to the prior inflammatory events rather than steroid or other immunosuppressant tapering. We cannot speculate on the influence of IMSUT-CORD treatment on the development of cGVHD, but we need to take care of the trigger episode of inflammation to prevent severe cGVHD.
In our study, Patient 2 presented stage 1 liver GVHD initially but progressed to liver dysfunction with elevated bilirubin during the administration of IMSUT-CORD. In contrast, Patient 3 with stage 3 liver aGVHD showed continuous elevation in bilirubin levels during the administration of IMSUT-CORD, then slow recovery to the normal range, while rapidly improving liver transferase levels together with a reduction of inflammatory cytokines from the first administration of IMSUT-CORD administration. The control of the liver GVHD remains unresolved without the suitable prognostic factors [24, 28]. Interestingly, the number of CD3−CD56+NK cells during IMSUT-CORD administration, significantly increased in all patients except Patient 2, who developed liver GVHD. NK cell number also correlated with IMSUT-CORD treatment rather than aGVHD grade. The influence of MSC on NK cells appears to be dependent on the surrounding environment. Boissel et al. showed that irradiated UC-MSCs feeder cells can increase the expansion of CB NK cells [29]. Vacca et al. reported that decidua MSCs (dMSCs) sustain the differentiation of CD34+ cell to CD56bright CD16− NK cells [30]. On the other hand, the same group demonstrate that dMSCs suppress IL-15-induced NK cell proliferation, inhibit the perforin and granzyme levels and NK cell cytotoxicity. Furthermore, TGF-β released by MSCs convert peripheral blood CD16+NK cells into CD16− NK cells [31]. Like M1 and M2 macrophage, NK cells which are classified into type 1 and type2 show different immunological functions [32], although the patient NK cells in this study remained unclassified. In the context of GVHD, Olson et al. [33] reported that NK cells suppress GVHD without causing GVHD. The authors showed that mice receiving donor NK and T cells exhibited improved survival and decreased GVHD scores compared with controls receiving donor T cells alone. Further, activated NK cells mediate the direct lysis of re-isolated GVHD-inducing T cells in vitro. Yamazaki et al. also demonstrated that the CD56+ cell dose in peripheral blood stem cells allografts is significantly inversely correlated with the incidence of GVHD, suggesting that NK cells have a suppressive effect on aGVHD [34]. Therefore, NK cell transition might be an indicator of aGVHD response to IMSUT-CORD.
Th17 cells producing IL-17 contribute to the development of GVHD. Levels of inflammatory cytokines, including IL-12, IL-17, and IL-33 (ST2), were decreased during IMSUT-CORD therapy, although they showed no correlation with aGVHD grade or treatment in such a small sample set. IL-33, together with regenerating islet-derived protein 3 alpha are reportedly correlated with acute GVHD [35,36,37]. These cytokines might be vise-versa to activate MSCs and be suppressed by MSCs. The IMSUT-CORD can be activated upon the stimulation with IFN-γ resulting in the induction of IDO-1, while IFN-γ and TNF-α were suppressed after the co-culture of IMSUT-CORD in allogeneic MLR in vitro. High levels of inflammatory cytokines in patient 3 might activate IMSUT-CORD resulting in the rapid suppression inflammatory cytokines and improvement of liver function.
The importance of chemokines in aGVHD treatment has not been elucidated. Damaged tissues or activated lymphocytes secrete chemokines that recruit MSCs, some of which are secreted by MSCs to recruit other cells. We found that CCL2 known as monocyte chemoattractant protein-1(MCP-1) and CCL11 (Eotaxin) levels were elevated during IMSUT-CORD administration and CCL11 was significantly correlated to the IMSUT-CORD therapy. However, the functions of chemokines are not fully understood in relation to MSC and MSC-associated immunotherapy. It is demonstrated that microvesicles from cytochalasin B-induced murine adipose-derived MSCs show a greater immunosuppressive potency after an increase in CCL11 levels than those from parental MSCs [38]. Additionally, CCL11 causes MSCs to traverse the BM endothelium, in a basal-to-apical and apical-to-basal direction [39]. Recently, Cruz-Barrera et al., through secretomic and transcriptomic analyses, demonstrated that the expression of CCL2 and CCL11 was upregulated by 2.29 folds in PHA-treated peripheral blood MNC/UC-MSC co-cultures compared to that in untreated UC-MSCs. They suggested that monocyte reprograming from M1 to M2 triggered by UC-MSCs may be related to the induction of CCL2 expression [40]. Furthermore, Zhang et al. reported that CCL2 levels in supernatants of the co-culture comprising IL-2–activated T cells and allogeneic BM-MSCs were significantly higher than those in the MSC monoculture due to the upregulation of CCR2 expression in T cells. They concluded that MSCs can enhance the chemotaxis of activated T cells through CCL2-CCR2 signaling in vitro [41], although the role of CCL11 remained unclear. To confirm which part secreted CCL2 and CCL11, not only CCL2 but also CCL11 levels (but less) were significantly increased in the supernatant of the allogeneic MLR co-cultured with UC-MSCs compared with those in the MLR alone, suggesting that IMSUT-CORD might have some influence on the elevation of these chemokines. In contrast, we could not mRNA of their representative receptors CCR2, CCR3, and CCR5 in IMSUT-CORD, as analyzed by qRT-PCR (Supplementary table 3). We do not know how much CCL2 and CCL11 secreted from UC-MSCs product (i.e., IMSUT-CORD) influence on the increase of these chemokines in the plasma of the patients treated with IMSUT-CORD, but our data suggested these chemokines may have some function in aGVHD. Further study is necessary to elucidate the CCL2/CCL11-CCRs axis in the context of MSC and immunosuppression.
Although further studies are needed due to the limitations of a phase 1 trial with such a small patient population, monitoring of the immunological cell population and cytokine and chemokine profile is valuable to identify the prognostic biomarkers to determine the success of aGVHD treatment with UC-MSCs. In conclusion, IMSUT-CORD cultured and cryopreserved in a serum-free solution was safe for patients with treatment-resistant-aGVHD and can be used to conduct efficacy-confirmation clinical trials.
Data availability statement
Deidentified individual participant data that underlie the reported results will be made available 3 months after publication for a period of 5 years after the publication date. Requests should be sent to tokikoni@ims.u-tokyo.ac.jp.
References
Reshef R, Saber W, Bolaños-Meade J, Chen G, Chen Y-B, Ho V, et al. Acute GVHD diagnosis and adjudication in a multicenter trial: a report from the BMT CTN 1202 biorepository study. J Clin Oncol. 2021;39:1878–87.
Harris A, Young R, Devine S, Hogan W, Ayuk F, Bunworasate U, et al. International, multicenter standardization of acute graft-versus-host disease clinical data collection: a report from the Mount Sinai acute GVHD international consortium. Biol Blood Marrow Transplant. 2016;22:4–10.
Penack O, Marchetti M, Ruutu T, Aljurf M, Bacigalupo A, Bonifazi F, et al. Prophylaxis and management of graft versus host disease after stem-cell transplantation for haematological malignancies: updated consensus recommendations of the European Society for Blood and Marrow Transplantation. Lancet Haematol. 2020;7:e157–67.
Le Blanc K, Rasmusson I, Götherstrom C, Seidel C, Sundberg B, Sundin M, et al. Mesenchymal stem cells inhibit the expression of CD25 (interleukin-2 receptor) and CD38 on phytohaemagglutinin-activated lymphocytes. Scand J Immunol. 2004;60:307–15.
Le Blanc K, Frassoni F, Ball L, Locatelli F, Roelofs H, Lewis I, et al. Mesenchymal stem cells for treatment of steroid-resistant, severe, acute graft-versus-host disease: a phase II study. Lancet. 2008;371:1579–86.
Hashmi S, Ahmed M, Murad M, Litzow M, Adams R, Ball L, et al. Survival after mesenchymal stromal cell therapy in steroid-refractory acute graft-versus-host disease: systematic review and meta-analysis. Lancet Haematol. 2016;3:e45-52.
Le Blanc K, Rasmusson I, Sundberg B, Gotherstrom C, Hassan M, Uzunel M, et al. Treatment of severe acute graft-versus-host disease with third party haploidentical mesenchymal stem cells. Lancet. 2004;363:1439–41.
Galleu A, Milojkovic D, Deplano S, Szydlo R, Loaiza S, Wynn R, et al. Mesenchymal stromal cells for acute graft-versus-host disease: response at 1 week predicts probability of survival. Br J Haematol. 2019;185:89–92.
Agata H, Asahina I, Watanabe N, Ishii Y, Kubo N, Ohshima S, et al. Characteristic change and loss of in vivo osteogenic abilities of human bone marrow stromal cells during passage. Tissue Eng Part A. 2010;16:663–73.
Nagamura-Inoue T, Nagamura F. Umbilical cord blood and cord tissue bank as a source for allogeneic use. IntechOpen. 2021. pp. 31–55 (Chapter 3).
Ishige I, Nagamura-Inoue T, Honda MJ, Harnprasopwat R, Kido M, Sugimoto M, et al. Comparison of mesenchymal stem cells derived from arterial, venous, and Wharton’s jelly explants of human umbilical cord. Int J Hematol. 2009;90:261–9.
He H, Nagamura-Inoue T, Takahashi A, Mori Y, Yamamoto Y, Shimazu T, et al. Immunosuppressive properties of Wharton’s jelly-derived mesenchymal stromal cells in vitro. Int J Hematol. 2015;102:368–78.
Shimazu T, Mori Y, Takahashi A, Tsunoda H, Tojo A, Nagamura-Inoue T. Serum- and xeno-free cryopreservation of human umbilical cord tissue as mesenchymal stromal cell source. Cytotherapy. 2015;17:593–600.
Mori Y, Ohshimo J, Shimazu T, He H, Takahashi A, Yamamoto Y, et al. Improved explant method to isolate umbilical cord-derived mesenchymal stem cells and their immunosuppressive properties. Tissue Eng Part C Methods. 2015;21:367–72.
Kurogi H, Takahashi A, Isogai M, Sakumoto M, Takijiri T, Hori A, et al. Umbilical cord derived mesenchymal stromal cells in microcarrier based industrial scale culture sustain the immune regulatory functions. Biotechnol J. 2021;16: e2000558.
He H, Takahashi A, Mukai T, Hori A, Narita M, Tojo A, et al. The immunomodulatory effect of triptolide on mesenchymal stromal cells. Front Immunol. 2021;12: 686356.
Narita M, Watanabe N, Yamahira A, Hashimoto S, Tochiki N, Saitoh A, et al. A leukemic plasmacytoid dendritic cell line, PMDC05, with the ability to secrete IFN-alpha by stimulation via Toll-like receptors and present antigens to naive T cells. Leuk Res. 2009;33:1224–32.
Przepiorka D, Weisdorf D, Martin P, Klingemann HG, Beatty P, Hows J, et al. 1994 Consensus conference on acute GVHD grading. Bone Marrow Transplant. 1995;15:825–8.
Filipovich AH, Weisdorf D, Pavletic S, Socie G, Wingard JR, Lee SJ, et al. National Institutes of Health consensus development project on criteria for clinical trials in chronic graft-versus-host disease: I. Diagnosis and staging working group report. Biol Blood Marrow Transplant. 2005;11:945–56.
Kurtzberg J, Prockop S, Teira P, Bittencourt H, Lewis V, Chan KW, et al. Allogeneic human mesenchymal stem cell therapy (remestemcel-L, Prochymal) as a rescue agent for severe refractory acute graft-versus-host disease in pediatric patients. Biol Blood Marrow Transplant. 2014;20:229–35.
Locatelli F, Algeri M, Trevisan V, Bertaina A. Remestemcel-L for the treatment of graft versus host disease. Expert Rev Clin Immunol. 2017;13:43–56.
Muroi K, Miyamura K, Okada M, Yamashita T, Murata M, Ishikawa T, et al. Bone marrow-derived mesenchymal stem cells (JR-031) for steroid-refractory grade III or IV acute graft-versus-host disease: a phase II/III study. Int J Hematol. 2016;103:243–50.
Muroi K, Miyamura K, Ohashi K, Murata M, Eto T, Kobayashi N, et al. Unrelated allogeneic bone marrow-derived mesenchymal stem cells for steroid-refractory acute graft-versus-host disease: a phase I/II study. Int J Hematol. 2013;98:206–13.
Murata M, Terakura S, Wake A, Miyao K, Ikegame K, Uchida N, et al. Off-the-shelf bone marrow-derived mesenchymal stem cell treatment for acute graft-versus-host disease: real-world evidence. Bone Marrow Transplant. 2021;56:2355–66.
Wu KH, Chan CK, Tsai C, Chang YH, Sieber M, Chiu TH, et al. Effective treatment of severe steroid-resistant acute graft-versus-host disease with umbilical cord-derived mesenchymal stem cells. Transplantation. 2011;91:1412–6.
Ding L, Han DM, Zheng XL, Yan HM, Xue M, Liu J, et al. Infusion of haploidentical hematopoietic stem cells combined with mesenchymal stem cells for treatment of severe aplastic anemia in adult patients yields curative effects. Cytotherapy. 2022;24:205–12.
Zu Y, Zhou J, Fu Y, Fang B, Liu X, Zhang Y, et al. Feasibility of reduced-dose posttransplant cyclophosphamide and cotransplantation of peripheral blood stem cells and umbilical cord-derived mesenchymal stem cells for SAA. Sci Rep. 2021;11:253.
Resnick IB, Barkats C, Shapira MY, Stepensky P, Bloom AI, Shimoni A, et al. Treatment of severe steroid resistant acute GVHD with mesenchymal stromal cells (MSC). Am J Blood Res. 2013;3:225–38.
Boissel L, Tuncer HH, Betancur M, Wolfberg A, Klingemann H. Umbilical cord mesenchymal stem cells increase expansion of cord blood natural killer cells. Biol Blood Marrow Transplant. 2008;14:1031–8.
Vacca P, Vitale C, Montaldo E, Conte R, Cantoni C, Fulcheri E, et al. CD34+ hematopoietic precursors are present in human decidua and differentiate into natural killer cells upon interaction with stromal cells. Proc Natl Acad Sci USA. 2011;108:2402–7.
Keskin DB, Allan DS, Rybalov B, Andzelm MM, Stern JN, Kopcow HD, et al. TGFbeta promotes conversion of CD16+ peripheral blood NK cells into CD16− NK cells with similarities to decidual NK cells. Proc Natl Acad Sci USA. 2007;104:3378–83.
Bogunia-Kubik K, Lacina P. Non-KIR NK cell receptors: role in transplantation of allogeneic haematopoietic stem cells. Int J Immunogenet. 2021;48:157–71.
Olson JA, Leveson-Gower DB, Gill S, Baker J, Beilhack A, Negrin RS. NK cells mediate reduction of GVHD by inhibiting activated, alloreactive T cells while retaining GVT effects. Blood. 2010;115:4293–301.
Yamasaki S, Henzan H, Ohno Y, Yamanaka T, Iino T, Itou Y, et al. Influence of transplanted dose of CD56+ cells on development of graft-versus-host disease in patients receiving G-CSF-mobilized peripheral blood progenitor cells from HLA-identical sibling donors. Bone Marrow Transplant. 2003;32:505–10.
Reichenbach DK, Schwarze V, Matta BM, Tkachev V, Lieberknecht E, Liu Q, et al. The IL-33/ST2 axis augments effector T-cell responses during acute GVHD. Blood. 2015;125:3183–92.
Choi SW, Braun T, Henig I, Gatza E, Magenau J, Parkin B, et al. Vorinostat plus tacrolimus/methotrexate to prevent GVHD after myeloablative conditioning, unrelated donor HCT. Blood. 2017;130:1760–7.
Solan L, Kwon M, Carbonell D, Dorado N, Balsalobre P, Serrano D, et al. ST2 and REG3alpha as predictive biomarkers after haploidentical stem cell transplantation using post-transplantation high-dose cyclophosphamide. Front Immunol. 2019;10:2338.
Gomzikova MO, Aimaletdinov AM, Bondar OV, Starostina IG, Gorshkova NV, Neustroeva OA, et al. Immunosuppressive properties of cytochalasin B-induced membrane vesicles of mesenchymal stem cells: comparing with extracellular vesicles derived from mesenchymal stem cells. Sci Rep. 2020;10:10740.
Smith H, Whittall C, Weksler B, Middleton J. Chemokines stimulate bidirectional migration of human mesenchymal stem cells across bone marrow endothelial cells. Stem Cells Dev. 2012;21:476–86.
Cruz-Barrera M, Florez-Zapata N, Lemus-Diaz N, Medina C, Galindo CC, Gonzalez-Acero LX, et al. Integrated analysis of transcriptome and secretome from umbilical cord mesenchymal stromal cells reveal new mechanisms for the modulation of inflammation and immune activation. Front Immunol. 2020;11: 575488.
Zhang YL, Qiao SK, Xing LN, Guo XN, Ren JH. Mesenchymal stem cells enhance chemotaxis of activated T cells through the CCL2-CCR2 axis in vitro. Bull Exp Biol Med. 2021;172:263–9.
Acknowledgements
We thank Yamamoto Y. and Miharu Y. at the Institute of Medical Science, University of Tokyo (IMSUT) for manufacturing products, performing quality management, and providing technical support. We thank Saito Y. at IMSUT Hospital for clinical trial preparations. We thank Tsunoda H., Sugita M., Satoh N., and the staff at NTT Medical Center Hospital (Tokyo), and Yamaguchi S. and the staff at Yamaguchi Hospital, for performing UC tissue and CB collection after obtaining informed consent. We thank the clinical coordinators and nurses at IMSUT Hospital, Komagome Hospital, and Toranomon Hospital for their continuous support during the clinical trial and Dr. Hanaoka and all staff at the Clinical Research Center of Chiba University Hospital. We thank Dr. Narita M at the School of Health Sciences, Faculty of Medicine, Niigata University, for providing PMDC05 cell line.
Funding
This work was supported by the Japan Agency for Medical Research and Development (AMED) (Project number: 17bk0104070h0001/18bk0104070h0002/19bk0104070h0003/20bk01041h0001/21bk0104109h0002). AMED No. 21be0804004h0001 supports the stable delivery of the source of somatic stem cells to researchers and companies as cord blood and UC-MSC medical product.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
The study was partially supported by the joint research study of the University of Tokyo with Rohto Pharmaceutical Co. Ltd. in the development of culture medium and processing (December 2014, December to March 2020) and the joint study of the University of Tokyo with Human Life Cord Japan Inc. in the development of banking and clinical application (September 2017–). Nagamura-Inoue is the senior medical advisor of Human Life Cord Japan Inc.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Below is the link to the electronic supplementary material.
About this article

Cite this article
Nagamura-Inoue, T., Kato, S., Najima, Y. et al. Immunological influence of serum-free manufactured umbilical cord-derived mesenchymal stromal cells for steroid-resistant acute graft-versus-host disease. Int J Hematol 116, 754–769 (2022). https://doi.org/10.1007/s12185-022-03408-7
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s12185-022-03408-7