
Abstract
The t(8;21) translocation is the most common cytogenetic abnormality in acute myeloid leukemia (AML). Although t(8;21) AML patients have a relatively favorable prognosis, relapse is a frequent occurrence, underscoring the need to develop novel therapeutic approaches. Here, we showed that t(8;21) AML is characterized by frequent mutation and overexpression of CCND2. Analysis of 19 AML cell lines showed that t(8;21) AML cells had lower IC50 values for the selective CDK4/6 inhibitors palbociclib and abemaciclib than non-t(8;21) AML cells. CDK4/6 inhibitors caused cell cycle arrest at G1 phase and impaired cell proliferation in t(8;21) AML cells. CDK4/6 inhibition decreased MAP-ERK and PI3K-AKT-mTOR signaling pathway activity, induced LC3B-I to LC3B-II conversion, and enhanced autophagosome formation, suggesting autophagy induction. Treatment of t(8;21) AML cells with the autophagy inhibitors chloroquine (CQ) or LY294002 in combination with the CDK4/6 inhibitor abemaciclib significantly increased the percentage of apoptotic (Annexin V positive) cells, whereas CQ or LY294002 single treatment had no significant effects. The effectiveness of co-inhibiting CDK4/6 and autophagy was confirmed in primary t(8;21) AML cells. The results suggest that the combination of CDK4/6 and autophagy inhibitors had a synergistic effect on inducing apoptosis, suggesting a novel therapeutic approach for the treatment of t(8;21) AML.
Similar content being viewed by others
Avoid common mistakes on your manuscript.
Introduction
Acute myeloid leukemia (AML) is a genetically and clinically heterogeneous disease [1]. t(8;21))(q22;q22)/RUNX1-RUNX1T1 [also known as AML1-ETO, hereafter referred to as t(8;21)] is one of the most frequent cytogenetic abnormalities in AML. The incidence of t(8;21) is 12–14% in pediatric AML and approximately 7% in adult AML [2,3,4,5,6]. Although t(8;21) AML patients have a more favorable prognosis than other cytogenetic subgroups, nearly 40% of t(8;21) AML patients experience relapse [7,8,9,10]. Therefore, novel therapeutic approaches based on a better understanding of the biology of t(8;21) AML need to be developed.
The RUNX1-RUNX1T1 fusion gene blocks myeloid differentiation, although the abnormality alone is not sufficient to induce leukemia [11, 12]. Additional abnormalities including driver mutations are required for leukemogenesis, and comprehensive analyses of genetic variations have been performed to identify these mutations. Several groups including ours recently identified novel CCND2 mutations in t(8;21) AML [13,14,15]. CCND2 encodes cyclin D2, which is one of three D-type cyclins (cyclin D1–3). D-type cyclins form complexes with CDK4/6 and induce cell cycle progression from G1 to S phase by phosphorylating the retinoblastoma protein [16, 17]. Most CCND2 mutations are located on the PEST domain, which is present in short-lived proteins and plays a role in their degradation [18]. CCND2 PEST domain mutations stabilize cyclin D2 and promote cell proliferation [13]. Deregulation of the cell cycle is a hallmark of cancer, and CDK4/6 inhibitors are currently in clinical trials for the treatment of solid cancers including breast cancer [19]. The efficacy of these inhibitors for improving survival rates has been reported [19, 20]. Because abemaciclib, a potent CDK4/6 inhibitor, is effective in cancer cells with deregulation of D-type cyclins [21], we hypothesized that CDK4/6 inhibitors may be effective for t(8;21) AML cells.
Here, we show that CCND2 mutations/overexpression is common in t(8;21) AML, and we demonstrate the effect of CDK4/6 inhibitors on blocking cell cycle progression and inhibiting the proliferation of t(8;21) AML cells. CDK4/6 inhibition induced autophagy in t(8;21) AML cells. Autophagy is an intracellular degradation system that recycles cellular constituents by engulfing them into double-membrane vesicles called autophagosomes [22]. The conversion of light chain (LC)3B-I to LC3B-II is a marker for autophagy induction [23]. Autophagy is regulated by several kinases: activation of class I phosphoinositide 3-kinase (PI3K) suppresses autophagy via the PI3K-AKT-mTOR pathway. By contrast, activation of class II and III PI3Ks contributes to initiation and progression of autophagy [24, 25]. MAP-ERK signaling pathway also negatively regulates autophagy [26]. Autophagy is associated with resistance to chemotherapy in cancer, and autophagy inhibition is a promising strategy for overcoming resistance [27, 28]. Several studies of solid tumors report CDK4/6 inhibition-induced autophagy and show that targeting both CDK4/6 and autophagy is a promising therapeutic strategy [29,30,31]. Therefore, we examined the effect of combination treatment with a CDK4/6 inhibitor and an autophagy inhibitor, and found that the two inhibitors acted synergistically to induce apoptosis in t(8;21) cells. These data suggest a novel therapeutic option based on the mechanism underlying the biology of t(8;21) AML.
Methods
CCND2 mutations and expression
Published data from mutational analyses of t(8;21) AML were searched. For pediatric t(8;21) AML, mutational data from 149 patients were collected from St. Jude Children’s Research Hospital tissue resource core facility and the Japanese Pediatric Leukemia/Lymphoma Study Group (JPLSG) AML-05 study [13, 15]. For adult t(8;21) AML, mutational data from 134 patients were collected from Cancer and Leukemia Group B (CALGB)/Alliance for Clinical Trials in Oncology (Alliance) trials and the University Hospital of Ulm [13, 14]. Lollipop plots for visualizing CCND2 mutations were generated using ProteinPaint (https://pecan.stjude.org/proteinpaint/).
CCND2 expression data from 1707 AML patients in the GSE13159, GSE15434, GSE61804, GSE14468, and Cancer Genome Atlas (TCGA) deposited to BloodSpot (https://servers.binf.ku.dk/bloodspot/) were compared between t(8;21) AML and non-t(8;21) AML patients [32]. Data on CCND2 mRNA expression in AML cell lines were also collected from the Cancer Cell Line Encyclopedia (CCLE) database (https://portals.broadinstitute.org/ccle) and compared between t(8;21) AML cell lines (Kasumi-1 and SKNO-1) and other AML cell lines [33].
Primary AML cells
Primary AML cells were obtained from the bone marrow of a pediatric t(8;21) AML patient at Kyoto University Hospital after provision of informed consent and institutional review board approval. The study was conducted in accordance with the Declaration of Helsinki.
AML cell lines
Kasumi-1, SKNO-1, ML-2, HL-60, HEL, MV4-11, NB-4, KG-1a, Kasumi-6, KG-1, KO52, MOLM-16, U937, Kasumi-3, UF-1, CMK-86, and MOLM-13 cell lines were purchased from the German Collection of Microorganisms and Cell Cultures (DSMZ, Braunschweig, Germany). THP-1 and NOMO-1 cells were purchased from the Japanese Collection of Research Bioresources Cell Bank (Ibaraki, Japan). These cell lines were cultured in Roswell Park Memorial Institute 1640 medium supplemented with 10% or 20% fetal bovine serum and 1% penicillin/streptomycin at 5% CO2 and 37 °C.
Compounds
Palbociclib (PD0332991) and abemaciclib (LY2835219) were obtained from AdooQ BioScience (Irvine, CA, USA). Both compounds were dissolved in dimethyl sulfoxide (DMSO) and used at a concentration of 500 nM according to previously published data [15]. Chloroquine (CQ) (#C6628, Sigma-Aldrich, St. Louis, MO, USA) was dissolved in PBS and used at a concentration of 10 µM. LY294002 (#129-04,861, FUJIFILM Wako Pure Chemical Corporation, Osaka, Japan) was dissolved in DMSO and used at a concentration of 1 µM.
Small hairpin RNA (shRNA) interference
shRNA targeting human cyclin D2 (shCCND2) (CTGCAATATGGGAACAAAT) and the control shRNA against luciferase (shLuc) were designed and cloned into pENTR4-H1tetOx1 and CS-RfA-ETBsd purchased from RIKEN BRC. Lentivirus was produced in HEK293T cells with psPAX2 and pMD2.G, and cells were transfected using polyethyleneimine (Sigma-Aldrich) for 48 h. The virus was collected, cells were infected, and infected cells were selected using Blasticidin S (Sigma-Aldrich).
Small interfering RNA (siRNA)
Experiments used siRNA targeting the human ATG7 transcript (siATG7) (sc-41447, Santa Cruz Biotechnology, Dallas, TX, USA) and control siRNA (siControl) (sc-37007, Santa Cruz Biotechnology, Dallas, TX, USA). Kasumi-1 cells (4 × 105 cells/mL) were cultured at 37 °C in RPMI 1640 medium containing 10% fetal bovine serum without 1% penicillin/streptomycin, and then transfected with siRNA/Lipofectamine RNAiMAX (Invitrogen) complexes diluted in Opti-MEM Reduced Serum Medium (Gibco) (final siRNA concentration, 50 nM), according to the manufacturer’s protocol. Two days after transfection, whole-cell lysates were prepared to determine knockdown efficiency by immunoblot analysis. Three days after transfection, cells were collected for apoptosis analysis.
Cell fertility assay
Cells (0.8–2 × 105/mL) were seeded according to the respective growing speed. SF reagent (Nacalai Tesque, Kyoto, Japan) was added after 48 h, and the absorbance was measured using the Tecan i-control infinite 200 (Tecan, Männedorf, Switzerland). The relative value was calculated based on the absorbance of the control.
Cell proliferation assay
Cells (2 × 105/mL) were cultured in DMSO in the presence of palbociclib or abemaciclib and counted daily for 4–5 days. Data are presented as the mean ± standard error (SE) of three independent experiments.
Cell cycle analysis
Cells (2 × 105/mL) were cultured in DMSO in the presence of palbociclib or abemaciclib for 72 h. Then, cells were stained with propidium iodide solution (PI) and analyzed using a FACS Canto II flow cytometer (BD Biosciences, San Jose, CA, USA).
Microarray and pathway analyses
Kasumi-1 and SKNO-1 cells were treated with DMSO or abemaciclib for 72 h. Cells were harvested and RNA was extracted using an RNase mini kit (QIAGEN). Microarrays were performed on GeneChip Arrays (Clariom™ S Assay, Human) (Filgen, Nagoya, Japan). The data were analyzed using the Microarray Data Analysis Tool Ver 3.2 (Filgen).
A total of 468 genes exhibiting a strong transcriptomic change (> 1.5-fold change in expression in DMSO-treated cells vs. abemaciclib-treated cells in both Kasumi-1 and SKNO-1 cell lines) were called. Pathway analysis was performed using the Kyoto Encyclopedia of Genes and Genomes (KEGG) database.
Immunoblot analysis
Cells were washed in PBS and lysed in RIPA buffer including a protease inhibitor cocktail (Nacalai Tesque). Whole-cell lysates were separated using 15% SDS polyacrylamide gels (SDS-PAGE) and transferred to Immobilon-P transfer membranes (Merck, Darmstadt, Germany). Membranes were blocked with Blocking One reagent (Nacalai Tesque) for 1 h at room temperature and incubated with the following primary antibodies overnight at 4 °C: anti-cyclin D2 (1/1000; #10,934-1-AP, Proteintech, Rosemont, IL, USA), anti-LC3B (1/1000; #L7543, Sigma-Aldrich), anti-cleaved caspase3 (1/500; #9661, CST, Danvers, MA, USA), or anti-GAPDH (1/3000; sc-47724, Santa Cruz Biotechnology, Dallas, TX, USA). Membranes were then incubated with HRP-conjugated anti-mouse IgG or anti-rabbit IgG (1/4000; #NA931; NA934, GE Healthcare Bio-Sciences, Little Chalfont, UK) for 1 h at room temperature. Protein bands were detected using an HRP Novex ECL Chemiluminescent Substrate Reagent Kit (Invitrogen, Carlsbad, CA, USA) and ChemiDoc™ XRS + Imager (Bio-Rad Laboratories, Hercules, CA, USA).
Transmission electron microscopy
Cells (2.5 × 106) cultured in DMSO containing palbociclib or abemaciclib were pelleted by centrifugation, fixed in 4% paraformaldehyde/2% glutaraldehyde/0.1 M phosphate buffer at 4℃, washed in isotonic phosphate-buffered sucrose, re-fixed in phosphate-buffered 1% OsO4, dehydrated through a graded series of ethanol solutions, and embedded in Luveak 812 (Nacalai Tesque). Thin sections (70–90 nm thick) were cut with a diamond knife on an EM UC6 ultramicrotome (Leica, Wetzlar, Germany), stained with uranyl acetate and lead citrate, and observed using a Hitachi H-7650 electron microscope (Hitachi, Tokyo, Japan).
Apoptosis assay
Cells were seeded at a density of 2 × 105 cells/mL and treated as indicated for 24 h. The collected cells were washed once with PBS- and suspended in 100 μL Annexin V binding buffer and 5 μL Annexin V using an apoptosis detection kit (BioLegend, San Diego, CA, USA). After addition of 10 µL PI and incubation at room temperature for 20 min, samples were prepared in 300 µL PBS and analyzed using the FACS Canto II flow cytometer (BD Biosciences).
Statistical analysis
Continuous variables were compared using the Student’s t-test or Mann–Whitney U-test. A P-value of < 0.05 was considered statistically significant. Statistical significance was defined as follows: P < 0.05 (*), P < 0.01 (**), or P < 0.001 (***).
Results
Frequent CCND2 mutations and high CCND2 expression in t(8;21) AML
To examine the frequency of CCND2 mutations in t(8;21) AML patients, we analyzed 149 pediatric patients from St. Jude Children’s Research Hospital tissue resource core facility and the JPLSG AML-05 study, and 134 adult patients from CALGB/Alliance trials and the University Hospital of Ulm. In 149 pediatric t(8;21) AML patients, 13 mutations were detected in 11 patients (11/149, 7.4%) (Fig. 1a). In 134 adult t(8;21) AML patients, 14 mutations were detected in 12 patients (12/134, 9.0%). In both cohorts, CCND2 mutations were located on the PEST domain, suggesting that the mutations stabilize the cyclin D2 protein.

Frequent CCND2 mutations and high CCND2 expression in t(8;21) AML. a Domain structure and location of CCND2 mutations in pediatric and adult t(8;21) AML patients. CCND2 mutations were detected in 11 of 149 pediatric t(8;21) AML patients and 12 of 134 adult t(8;21) AML patients. b CCND2 mRNA expression data from 1707 patients [non-t(8;21) AML patients (n = 1609); t(8;21) AML patients (n = 98)] in the GSE13159, GSE15434, GSE61804, GSE14468, and Cancer Genome Atlas (TCGA) deposited to BloodSpot (https://servers.binf.ku.dk/bloodspot/) were compared. c CCND2 mRNA expression in 34 AML cell lines [non-t(8;21) AML cell lines (n = 32) and t(8;21) AML cell lines (n = 2, Kasumi-1 and SKNO-1)] from the CCLE database was compared. d Kasumi-1 cells were transfected with shLuc or shCCND2. Immunoblot analysis showed significantly lower expression of cyclin D2 in shCCND2-transfected cells than in shLuc-transfected cells. e Kasumi-1 cells transfected with shCCND2 showed impaired cell proliferation. f. Cell cycle analysis showed a significantly lower percentage of cells in the S/G2/M phase in shCCND2-transfected Kasumi-1 cells compared with in shLuc-transfected cells. Data are presented as the mean ± SE of three independent experiments. *P < 0.05, **P < 0.01
Next, we compared CCND2 expression in t(8;21) AML patients with that in other AML patients. RNA expression data from 1707 patients in the GSE13159, GSE15434, GSE61804, GSE14468, and TCGA deposited to BloodSpot (https://servers.binf.ku.dk/bloodspot/) were analyzed. The results showed that CCND2 expression t(8;21) in AML patients (n = 98) was significantly higher than that in other AML patients (n = 1609) (P = 0.003, Fig. 1b). In AML cell lines, those with t(8;21) (n = 2: Kasumi-1 and SKNO-1; both CCND2 wild-type) showed higher CCND2 expression than those without (n = 32); however, the difference was not statistically significant (P = 0.18, Fig. 1c), probably due to the very small sample size.
To determine the role of high CCND2 expression in t(8;21) AML, we performed shRNA knockdown analysis. Kasumi-1 cells transfected with shCCND2 showed a lower rate of cell proliferation than those transfected with shLuc (control) (Fig. 1d, e). Cell cycle analysis showed that the percentage of cells in S/G2/M phase was significantly lower in shCCND2-transfected Kasumi-1 cells than in the controls, suggesting that cells were arrested in G1 phase (Fig. 1f; Supplementary Fig. 1). These results suggest that the frequency of CCND2 mutations and CCND2 expression are increased in t(8;21) AML, and high CCND2 expression plays an important role in t(8;21) AML cell proliferation.
Efficacy of CDK4/6 inhibitors in t(8;21) AML cell lines
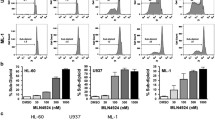
Because CCND2 is not a druggable target, we examined the effect of CDK4/6 inhibitors (palbociclib and abemaciclib) on t(8;21) AML cells. The IC50 (50% inhibitory concentration) of CDK4/6 inhibitors was screened using 19 AML cell lines including two t(8;21) AML cell lines (Fig. 2a). There was a moderate correlation in IC50 values between palbociclib and abemaciclib (R2 = 0.56). The IC50 values for both palbociclib and abemaciclib were significantly lower in the two t(8;21) AML cell lines than in the non-t(8;21) AML cell lines (Fig. 2a, b).

CDK4/6 inhibitors induce cell cycle arrest in t(8;21) AML cell lines. a IC50 of abemaciclib and palbociclib in 19 AML cell lines including two t(8;21) and 17 non-t(8;21) AML cell lines. b Raw data from IC50 screening according to FAB subtypes of cell lines. c t(8;21) AML cells treated with palbociclib or abemaciclib showed impaired cell proliferation. d, e Treatment with CDK4/6 inhibitors decreased the percentage of S/G2/M phase cells in t(8;21) AML cells. Data are presented as the mean ± SE of three independent experiments. *P < 0.05, ***P < 0.001
Investigation of the mechanism of action of CDK4/6 inhibitors in t(8;21) AML cell lines showed that the inhibitors impaired proliferation in Kasumi-1 and SKNO-1 cells (Fig. 2c). In addition, treatment with CDK4/6 inhibitors decreased the percentage of S/G2/M phase cells detected by flow cytometry, suggesting that CDK4/6 inhibition arrested cells in G1 phase (Fig. 2d, e). We also examined the effect of CDK4/6 inhibition on proliferation in shLuc-transfected and shCCND2-transfected Kasumi-1 cells. We found that shLuc + abemaciclib and shCCND2 + abemaciclib cells gave similar results in that both showed a significant decrease in proliferation compared with shLuc and shCCND2 cells (Supplementary Fig. 2). These results indicate that CDK4/6 inhibitors may be effective for the treatment of t(8;21) AML.
CDK4/6 inhibitors induce autophagy in t(8;21) AML cells
To identify potential therapeutic approaches in combination with CDK4/6 inhibitors in t(8;21) AML, we performed microarray analysis to examine the effects of CDK4/6 inhibition in addition to cell cycle arrest. Kasumi-1 and SKNO-1 cell lines were incubated with DMSO or abemaciclib, and RNA expression levels were compared. The number of genes showing a significant change (> 1.5-fold up or down in DMSO vs. abemaciclib) was 2562 in Kasumi-1 cells, 1560 in SKNO-1 cells, and 468 in both cell lines (Fig. 3a, b). Pathway analysis of the 468 genes changed in both cell lines was performed using the KEGG database, and pathways with P < 0.05 were extracted. In addition to the pathways associated with the cell cycle (regulation of sister chromatid separation, retinoblastoma gene, and cell cycle), the MAP-ERK and (Class I) PI3K-AKT-mTOR signaling pathways were downregulated by CDK4/6 inhibition (Fig. 3c). Because these pathways are involved in autophagy regulation via mTOR (Fig. 3d), we focused on examining autophagy in subsequent experiments.

Microarray and pathway analysis of t(8;21) AML cell lines treated with abemaciclib. a Comparison of gene expression between DMSO-treated control cells and abemaciclib-treated cells in the Kasumi-1 and SKNO-1 cell lines. b The number of genes showing a significant change (> 1.5-fold up or down) was 2562 in Kasumi-1 cells, 1560 in SKNO-1 cells, and 468 in both cells. c Pathway analysis of the 468 genes changed in both cell lines was performed using the KEGG database, and pathways with P < 0.05 were extracted. The upregulated pathways are indicated in red characters and the downregulated pathways in blue. MAP-ERK and PI3K-AKT-mTOR signaling pathways are highlighted in bold. d Schematic figure showing the contribution of the MAP-ERK and PI3K-AKT-mTOR signaling pathways to autophagy regulation via mTOR
Assessment of the effect of CDK4/6 inhibition on autophagy in t(8;21) AML cells showed that inhibitor treatment induced LC3B-I to LC3B-II conversion in both Kasumi-1 and SKNO-1 cells (Fig. 4a, b). Transmission electron microscopic examination of autophagosome formation detected a large number of autophagosomes in the cytoplasm of Kasumi-1 and SKNO-1 cells treated with CDK4/6 inhibitors, whereas few autophagosomes were detected in control samples (Fig. 4c, d). These results suggest that autophagy is induced by CDK4/6 inhibition in t(8;21) AML cells.

Induction of autophagy in t(8;21) AML cells treated with CDK4/6 inhibitors. a, b Immunoblot analysis showing the effect of CDK4/6 inhibitors on inducing LC3B-I to LC3B-II conversion in t(8;21) AML cells. Data from immunoblots were quantified by densitometric analysis (n = 4). c, d The formation of double-membrane structures (autophagosomes) in t(8;21) AML cells treated with CDK4/6 inhibitors was observed by TEM. The black arrows indicate autophagosomes. *P < 0.05, **P < 0.01
CDK4/6 and autophagy inhibitors synergistically induce apoptosis in t(8;21) AML cells
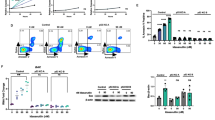
Autophagy is involved in the resistance to chemotherapy in cancer cells [27]. Therefore, we hypothesized that autophagy inhibition may be a promising therapeutic approach. Two autophagy inhibitors with different mechanisms of action were selected: CQ neutralizes the acidic pH of intracellular vesicles and blocks lysosomal degradation, and LY294002 inhibits class III PI3Ks [34]. The percentage of Annexin V positive (apoptotic) cells was determined by flow cytometry in Kasumi-1 cells. The percentage of apoptotic cells did not differ significantly between cells treated with CQ or LY294002 and control cells (control vs. CQ: P = 0.24, control vs. LY294002: P = 0.22) (Fig. 5a, b). However, the percentage of apoptotic cells was significantly higher in cells treated with autophagy inhibitors in combination with abemaciclib than in cells treated with abemaciclib alone (abemaciclib vs. abemaciclib plus CQ: P = 0.00001, abemaciclib vs. abemaciclib plus LY294002: P = 0.03). Consistently, treatment with both abemaciclib and autophagy inhibitors upregulated the expression of cleaved caspase3, a marker of apoptosis (Fig. 5c). The combinatorial effect was confirmed by silencing the autophagy-related protein ATG7 using small interfering RNA in abemaciclib-treated t(8;21) AML cells (Supplementary Fig. 3).

CDK4/6 and autophagy inhibitors in combination synergistically induce apoptosis in t(8;21) AML cells. a, b The percentage of apoptotic (Annexin V positive) cells was examined by flow cytometry. The percentage of apoptotic cells did not differ significantly between chloroquine (CQ) or LY294002 treated Kasumi-1 cells and control cells. The percentage of apoptotic cells was significantly higher in Kasumi-1 cells treated with autophagy inhibitors in combination with abemaciclib than in cells treated with abemaciclib alone. c The expression of cleaved caspase 3 was examined by immunoblotting. Combination treatment with abemaciclib and autophagy inhibitors upregulated cleaved caspase 3 expression. d, e The percentage of apoptotic cells was examined using a primary t(8;21) AML sample. The number of apoptotic cells did not differ significantly between CQ-treated cells and control cells. The percentage of apoptotic cells was significantly higher in primary t(8;21) AML cells treated with CQ plus abemaciclib than in cells treated with abemaciclib alone. f Schematic figure showing that co-inhibition of CDK4/6 and autophagy enhances apoptosis in t(8;21) AML cells, which are characterized by frequent CCND2 mutations or high CCND2 expression. Data are presented as the mean ± SE of three independent experiments. *P < 0.05, ***P < 0.001; ns not significant
To confirm the result using a t(8;21) AML cell line, we examined the efficacy of co-inhibiting CDK4/6 and autophagy in a primary bone marrow sample from a pediatric t(8;21) AML patient. Treatment of primary t(8;21) AML cells with CQ resulted in LC3B-I to LC3B-II conversion, suggesting induction of autophagy (Supplementary Fig. 4). The percentage of apoptotic cells did not differ significantly between control cells and cells treated with CQ (P = 0.40) (Fig. 5d, e). However, the percentage of apoptotic cells was significantly higher after treatment with autophagy inhibitors plus abemaciclib than after treatment with abemaciclib alone (P = 0.049). These results suggest that CDK4/6 and autophagy inhibitors synergistically induce apoptosis in primary t(8;21) AML cells (Fig. 5f).
Discussion
In this study, we showed that CCND2 mutations located on the PEST domain occur at a high rate in t(8;21) AML. CCND2 mutations in t(8;21) AML were reported by three independent groups although the number of patients analyzed in each study was small. We showed that the frequency of CCND2 mutations was comparable between pediatric and adult t(8;21) AML patients (7.4% and 9.0%, respectively), suggesting that the genetic abnormality is a common feature in t(8;21) AML regardless of patient age. The frequency of CCND2 mutations in patients with non-core binding factor (CBF)-AML is 0.4% (6/1426), supporting the high frequency of CCND2 mutations in t(8;21) AML [14]. We also showed that CCND2 expression in t(8;21) AML patients is higher than that in non-t(8;21) AML patients. This result may be explained by a recent study showing that CCND2 is a crucial transcriptional target of RUNX1-ETO protein and that this fusion protein upregulates CCND2 expression [35]. CCND2 mutations located in the PEST domain stabilize cyclin D2 protein [13]. Therefore, the effect of CCND2 mutations on CCND2 expression would be reflected at the protein level. Comparison of cyclin D2 protein expression between patients with CCND2 wild-type and those with the CCND2 mutant would be an interesting topic for future study.
Among the 19 AML cell lines analyzed, t(8;21) AML cell lines (Kasumi-1 and SKNO-1) showed high sensitivity to CDK4/6 inhibition. In the IC50 screening, three non-t(8;21) AML cell lines with FAB-M2 (HL-60, Kasumi-6, and KO52) were included. Despite the fact that these cell lines share the same FAB-subtype with Kasumi-1 and SKNO-1 cells, the IC50 values were > 10-fold higher than those of t(8;21) AML cell lines. This difference may be explained by the fact that deregulation of D-type cyclins increases the sensitivity of cancer cells to CDK4/6 inhibitors [21]. Several non-t(8;21) AML cell lines were also sensitive to CDK4/6 inhibitors, suggesting that these cell lines may harbor other D-cyclin activating features including CCND3 amplification. Although CDK4/6 inhibitors are currently in clinical trials for the treatment of various solid tumors, there are few clinical trials assessing their efficacy in leukemia [19]. Clinical trials for AML, especially for t(8;21) AML, should be considered. We also showed that the proliferation curves of shLuc-transfected and shCCND2-transfected Kasumi-1 cells are similar when CDK4/6 is inhibited. This result is reasonable because CCND2 and CDK4/6 share the same pathway: cyclin D2 forms complexes with CDK4/6 and induces cell cycle progression [16, 17].
CDK4/6 inhibition induced autophagy in t(8;21) AML cells. Despite the promising results of clinical trials of CDK4/6 inhibitors in solid tumors, various resistance mechanisms against CDK4/6 inhibition have been reported [36, 37]. In this study, combination treatment with CDK4/6 and autophagy inhibitors induced apoptosis, suggesting that autophagy is involved in the mechanism of resistance to CDK4/6 inhibition in t(8;21) AML. Targeting both CDK4/6 and autophagy showed promising results with respect to treatment of solid tumor cells [30, 31], supporting the notion that inhibiting autophagy may be a key strategy for overcoming the resistance to CDK4/6 inhibition. CQ, one of the autophagy inhibitors used in this study, is a well-tolerated antimalarial drug, and several clinical trials combining CQ with anticancer therapies are ongoing [27, 38]. Thus, CQ may be a promising agent in combination with CDK4/6 inhibitors.
In conclusion, the present results indicate that inhibition of CDK4/6 and autophagy may be a novel and promising biomarker-driven therapeutic strategy for the treatment of t(8;21) AML. The results of this study were obtained using in vitro analyses, and in vivo experiments are necessary to confirm the results in the future.
References
Papaemmanuil E, Gerstung M, Bullinger L, Gaidzik VI, Paschka P, Roberts ND, et al. Genomic classification and prognosis in acute myeloid leukemia. N Engl J Med. 2016;374:2209–21.
Grimwade D, Hills RK, Moorman AV, Walker H, Chatters S, Goldstone AH, et al. Refinement of cytogenetic classification in acute myeloid leukemia: determination of prognostic significance of rare recurring chromosomal abnormalities among 5876 younger adult patients treated in the United Kingdom Medical Research Council trials. Blood. 2010;116:354–65.
Harrison CJ, Hills RK, Moorman AV, Grimwade DJ, Hann I, Webb DK, et al. Cytogenetics of childhood acute myeloid leukemia: United Kingdom Medical Research Council Treatment trials AML 10 and 12. J Clin Oncol. 2010;28:2674–81.
von Neuhoff C, Reinhardt D, Sander A, Zimmermann M, Bradtke J, Betts DR, et al. Prognostic impact of specific chromosomal aberrations in a large group of pediatric patients with acute myeloid leukemia treated uniformly according to trial AML-BFM 98. J Clin Oncol. 2010;28:2682–9.
Raimondi SC, Chang MN, Ravindranath Y, Behm FG, Gresik MV, Steuber CP, et al. Chromosomal abnormalities in 478 children with acute myeloid leukemia: clinical characteristics and treatment outcome in a cooperative pediatric oncology group study-POG 8821. Blood. 1999;94:3707–16.
Byrd JC, Mrozek K, Dodge RK, Carroll AJ, Edwards CG, Arthur DC, et al. Pretreatment cytogenetic abnormalities are predictive of induction success, cumulative incidence of relapse, and overall survival in adult patients with de novo acute myeloid leukemia: results from Cancer and Leukemia Group B (CALGB 8461). Blood. 2002;100:4325–36.
Marcucci G, Mr´ozek K, Ruppert AS, Maharry K, Kolitz JE, Moore JO, et al. Prognostic factors and outcome of core binding factor acute myeloid leukemia patients with t(8;21) differ from those of patients with inv(16): a Cancer and Leukemia Group B study. J Clin Oncol. 2005;23:5705–17.
Schlenk RF, Benner A, Krauter J, Büchner T, Sauerland C, Ehninger G, et al. Individual patient data-based meta-analysis of patients aged 16 to 60 years with core binding factor acute myeloid leukemia: a survey of the German Acute Myeloid Leukemia Intergroup. J Clin Oncol. 2004;22:3741–50.
Jourdan E, Boissel N, Chevret S, Delabesse E, Renneville A, et al. Prospective evaluation of gene mutations and minimal residual disease in patients with core binding factor acute myeloid leukemia. Blood. 2013;121:2213–23.
Krauth MT, Eder C, Alpermann T, Bacher U, Nadarajah N, Kern W, et al. High number of additional genetic lesions in acute myeloid leukemia with t(8;21)/RUNX1-RUNX1T1: frequency and impact on clinical outcome. Leukemia. 2014;28:1449–588.
Higuchi M, O'Brien D, Kumaravelu P, Lenny N, Yeoh EJ, Downing JR. Expression of a conditional AML1-ETO oncogene bypasses embryonic lethality and establishes a murine model of human t(8;21) acute myeloid leukemia. Cancer Cell. 2002;1:63–74.
Yuan Y, Zhou L, Miyamoto T, Iwasaki H, Harakawa N, Hetherington CJ, et al. AML1-ETO expression is directly involved in the development of acute myeloid leukemia in the presence of additional mutations. Proc Natl Acad Sci U S A. 2001;98:10398–403.
Faber ZJ, Chen X, Gedman AL, Boggs K, Cheng J, Ma J, et al. The genomic landscape of core-binding factor acute myeloid leukemias. Nat Genet. 2016;48:1551–6.
Eisfeld AK, Kohlschmidt J, Schwind S, Nicolet D, Blachly JS, Orwick S, et al. Mutations in the CCND1 and CCND2 genes are frequent events in adult patients with t(8;21)(q22;q22) acute myeloid leukemia. Leukemia. 2017;31:1278–85.
Matsuo H, Yoshida K, Fukumura K, Nakatani K, Noguchi Y, Takasaki S, et al. Recurrent CCND3 mutations in MLL-rearranged acute myeloid leukemia. Blood Adv. 2018;2:2879–89.
Xiong Y, Zhang H, Beach D. D type cyclins associate with multiple protein kinases and the DNA replication and repair factor PCNA. Cell. 1992;71:505–14.
Bates S, Parry D, Bonetta L, Vousden K, Dickson C, Peters G. Absence of cyclin D/cdk complexes in cells lacking functional retinoblastoma protein. Oncogene. 1994;9:1633–40.
Rogers S, Wells R, Rechsteiner M. Amino acid sequences common to rapidly degraded proteins: the PEST hypothesis. Science. 1986;234:364–8.
Ingham M, Schwartz GK. Cell-cycle therapeutics come of age. J Clin Oncol. 2017;35:2949–59.
Turner NC, Ro J, André F, Loi S, Verma S, Iwata H, et al. Palbociclib in hormone-receptor-positive advanced breast cancer. N Engl J Med. 2015;373:209–19.
Gong X, Litchfield LM, Webster Y, Chio LC, Wong SS, Stewart TR, et al. Genomic aberrations that activate D-type cyclins are associated with enhanced sensitivity to the CDK4 and CDK6 inhibitor abemaciclib. Cancer Cell. 2017;32:761–76.
Mizushima N. Autophagy: process and function. Genes Dev. 2007;21:2861–73.
Mizushima N, Yoshimori T. How to interpret LC3 immunoblotting. Autophagy. 2007;3:542–5.
Yu X, Long YC, Shen HM. Differential regulatory functions of three classes of phosphatidylinositol and phosphoinositide 3-kinases in autophagy. Autophagy. 2015;11:1711–28.
Bilanges B, Posor Y, Vanhaesebroeck B. PI3K isoforms in cell signalling and vesicle trafficking. Nat Rev Mol Cell Biol. 2019;20:515–34.
Sridharan S, Jain K, Basu A. Regulation of autophagy by kinases. Cancers (Basel). 2011;3:2630–54.
Levy JMM, Towers CG, Thorburn A. Targeting autophagy in cancer. Nat Rev Cancer. 2017;17:528–42.
Kamitsuji Y, Kuroda J, Kimura S, Toyokuni S, Watanabe K, Ashihara E, et al. The Bcr-Abl kinase inhibitor INNO-406 induces autophagy and different modes of cell death execution in Bcr-Abl-positive leukemias. Cell Death Differ. 2008;15:1712–22.
Valenzuela CA, Vargas L, Martinez V, Bravo S, Brown NE. Palbociclib-induced autophagy and senescence in gastric cancer cells. Exp Cell Res. 2017;360:390–6.
Vijayaraghavan S, Karakas C, Doostan I, Chen X, Bui T, Yi M, et al. CDK4/6 and autophagy inhibitors synergistically induce senescence in Rb positive cytoplasmic cyclin E negative cancers. Nat Commun. 2017;8:15916.
Ji Y, Liu X, Li J, Xie X, Huang M, Jiang J, et al. Use of ratiometrically designed nanocarrier targeting CDK4/6 and autophagy pathways for effective pancreatic cancer treatment. Nat Commun. 2020;11:4249.
Bagger FO, Kinalis S, Rapin N. BloodSpot: a database of healthy and malignant haematopoiesis updated with purified and single cell mRNA sequencing profiles. Nucleic Acids Res. 2019;47:D881–D88585.
Ghandi M, Huang FW, Jané-Valbuena J, Kryukov GV, Lo CC, McDonald ER 3rd, et al. Next-generation characterization of the Cancer Cell Line Encyclopedia. Nature. 2019;569:503–8.
Vakifahmetoglu-Norberg H, Xia HG, Yuan J. Pharmacologic agents targeting autophagy. J Clin Invest. 2015;125:5–13.
Martinez-Soria N, McKenzie L, Draper J, Ptasinska A, Issa H, Potluri S, et al. The oncogenic transcription factor RUNX1/ETO corrupts cell cycle regulation to drive leukemic transformation. Cancer Cell. 2018;34:626–42.
Herrera-Abreu MT, Palafox M, Asghar U, Rivas MA, Cutts RJ, Garcia-Murillas I, et al. Early adaptation and acquired resistance to CDK4/6 Inhibition in estrogen receptor-positive breast cancer. Cancer Res. 2016;76:2301–13.
Formisano L, Lu Y, Servetto A, Hanker AB, Jansen VM, Bauer JA, et al. Aberrant FGFR signaling mediates resistance to CDK4/6 inhibitors in ER+ breast cancer. Nat Commun. 2019;10:1373.
Onorati AV, Dyczynski M, Ojha R, Amaravadi RK. Targeting autophagy in cancer. Cancer. 2018;124:3307–18.
Acknowledgements
This work was supported by JSPS KAKENHI Grant Number JP19K16832 and a research Grant from the Japanese Society of Hematology. This work is supported in part by the Practical Research for Innovative Cancer Control (JP19ck0106329) from the Japan Agency for Medical Research and Development (AMED).
The authors thank Prof. Junko Takita, Dr. Itaru Kato, and Dr. Takashi Mikami (Department of Pediatrics, Graduate School of Medicine, Kyoto University) for providing primary AML cells. The authors also thank Keiko Furuta and Haruyasu Kohda (Division of Electron Microscopic Study, Center for Anatomical Studies, Graduate School of Medicine, Kyoto University) for technical assistance.
Author information
Authors and Affiliations
Contributions
KN and HM analyzed sequencing data and performed the functional analysis. YH and MH helped with data acquisition. KN, HM, YH, MH, AK, MN, YN-A, YK, and SA contributed to the interpretation of data. KN and HM prepared the figures and wrote the manuscript. HM, YK, and SA supervised the project.
Corresponding author
Ethics declarations
Conflict of interest
The authors declare no competing financial interests.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Electronic supplementary material
Below is the link to the electronic supplementary material.
About this article

Cite this article
Nakatani, K., Matsuo, H., Harata, Y. et al. Inhibition of CDK4/6 and autophagy synergistically induces apoptosis in t(8;21) acute myeloid leukemia cells. Int J Hematol 113, 243–253 (2021). https://doi.org/10.1007/s12185-020-03015-4
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s12185-020-03015-4