
Abstract
The Canadian reference method for the enumeration of Listeria monocytogenes (Health Canada method MFLP-74) was modified by the addition of a layer of non-selective tryptone soy agar (TSA) to the Oxford, Palcam, and (or) Rapid’L.mono selective agars, to improve the recovery of stressed L. monocytogenes from frozen foods. The performance of the standard selective agars versus the selective agars with an additional TSA agar layer (TAL agars) showed that for each of the frozen food items tested (vegetables, shrimp and meatballs), as well as for all data combined, L. monocytogenes counts were significantly higher (up to 20% higher) in TAL agars in relation to the standard agars (p < 0.0001, α = 0.05). A selectivity assessment showed that the 50 inclusivity L. monocytogenes strains tested produced true positive reactions on the TAL agars, and the 30 exclusivity bacterial strains tested produced true negative results on the TAL agars, indicating that the specificity of the selective agars was not altered by the additional non-selective TSA layer. In light of the 2016 Listeria outbreak linked to frozen fruits and vegetables in the USA, the results of this method investigation seem to be timely and particularly important for the regulatory testing of frozen foods.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Listeria monocytogenes is a Gram-positive bacterial pathogen widely distributed in the environment, and frequently found in food products (reviewed by Farber and Peterkin 1991). Ingestion of food contaminated with L. monocytogenes can result in listeriosis, which manifests either as a severe systemic infection with a high fatality rate (Schlech 2000; Schlech et al. 1983), or as febrile gastroenteritis (Aureli et al. 2000; Dalton et al. 1997; Schlech 1997). Listeriosis outbreaks are most often associated with ready-to-eat (RTE) food products, which are consumed without prior cooking and thus regarded as high-risk foods (Farber and Peterkin 1991). In Canada, RTE foods are classified into two categories (Health Canada 2011). Category 1 includes those RTE products in which L. monocytogenes is capable of growth to high numbers, even at refrigeration temperatures. Category 2 RTE foods include those products that do not support the growth of L. monocytogenes, or where this bacterium will not grow above the regulatory action limit of 100 colony-forming units per gram (CFU/g).
The Canadian government has a zero tolerance policy for L. monocytogenes present in Category 1 RTE food types. However, a maximum level of 100 CFU/g of L. monocytogenes is placed on Category 2 RTE products (e.g., frozen foods). In these products, the levels of L. monocytogenes should not exceed 100 CFU/g throughout their stated shelf-life (Health Canada 2011). Therefore, the Canadian policy considers both the presence/absence of L. monocytogenes (tested by Health Canada method MFHPB-30) (Pagotto et al. 2011a), and the actual levels of L. monocytogenes in a food product (tested by Health Canada method MFLP-74) (Pagotto et al. 2011b), according to the potential for the organism to grow in that particular food product, i.e., its RTE category. However, only certain RTE foods are subject to the provisions of the Listeria policy, i.e., foods that have been subjected to some form of processing (most often cooking), and (or) foods that have been treated to extend their shelf-life (e.g., frozen foods) (Health Canada 2011).
In food, L. monocytogenes is exposed to environmental stresses caused by processing and preserving treatments, which often include exposure to high and low temperatures (Besse 2002). Heat and freezing treatments may cause microorganisms to be killed or sub-lethally injured, although some may be able to survive without injury (Berry et al. 2009; Wu and Fung 2001; Wu et al. 2001b). Using the definitions reviewed by Wu (2008), physiologically and (or) structurally injured bacterial cells are unable to grow and form colonies on selective media that support the growth of non-injured bacteria (Golden et al. 1988). Food products contaminated with stressed cells of a bacterial pathogen thus pose a potentially undetectable health risk, because sub-lethally injured cells (which will not grow on selective media during enrichment and [or] isolation) have the capability to later repair, return to a normal physiological state, acquire cross-resistance to other stresses, and regain their ability to cause disease (Wesche et al. 2009; Wu 2008). Therefore, from a risk-based regulatory perspective, microbiology methods should be able to detect both injured and non-injured cells from foods. That is, bacterial enumeration methods should allocate provisions to stimulate the repair and growth of injured bacterial cells; a process known as resuscitation. Resuscitation may be achieved by providing optimal growth conditions for a brief period, for instance, incubation for 1–4 h at a suitable temperature in a nutrient-rich, non-selective medium (ISO 1998; Wu 2008). A simple and practical resuscitation method is that originally reported by Kang and Fung (1999), which consists in overlaying one or two thin layers of non-selective TSA onto a pre-solidified plate of selective agar. This modification, known as TAL (for thin agar layer), is applicable to virtually any bacterial enumeration method that uses selective agars for the isolation of a target stressed bacterial pathogen (Wu and Fung 2001, Wu et al. 2001a, b). The TAL modification, however, not always results in enhanced recovery of stressed cells (Lavieri et al. 2014), nor always produces significantly higher counts in relation to standard selective agars (Wu et al. 2001b; Yuste et al. 2003), suggesting that the effectiveness of this method modification may depend on multiple factors, having to be specifically evaluated for different pathogens in different food matrices and for different stress-mediated injuries.
In MFLP-74, a portion of sample is blended in a prescribed diluent, surface-plated directly onto at least two selective agars (one of which has to be the Oxford selective agar), and incubated under specified conditions of time and temperature to allow the formation of identifiable colonies of L. monocytogenes (Pagotto et al. 2011b). Although it is logical to expect that L. monocytogenes cells present in the tested food items may be injured by the physiological stress imposed by food processing and storage, MFLP-74 does not include a resuscitation step, as does method ISO 11290-2, which is the European reference method for L. monocytogenes enumeration (ISO 1998). Thus, the hypothesis for this study is that direct plating of food suspensions on selective medium without resuscitation would underestimate L. monocytogenes actual numbers (Besse and Colin 2004). The specific objective was to develop and validate the TAL agar modification for the enumeration of stressed L. monocytogenes (artificially injured in vitro by heat and freezing), in the context of the Canadian regulatory method MFLP-74, as applied for the testing of frozen foods at the Canadian Food Inspection Agency (CFIA).
Materials and Methods
Bacterial Strains and General Culture Conditions
The L. monocytogenes strains and control isolates used for the pilot and validation studies have been previously characterized by the CFIA Dartmouth Laboratory (Table 1). The bacterial isolates used for the selectivity assessment are listed in Online Resource 1 (ESM_1.pdf), all being available from the CFIA Dartmouth Laboratory culture collection. Fifty L. monocytogenes strains representing a range of serotypes from clinical, environmental and food sources were used in the inclusivity study, and 30 cultures of microorganisms known to cause interference of (or be closely related to) L. monocytogenes were selected for the exclusivity study. Stock bacterial cultures were maintained below −70 °C on cryobeads in Microbank™ vials (Pro-Lab Diagnostics). To activate stock cultures, cryobeads were routinely streaked onto Tryptone Soy Agar with 0.6% yeast extract (TSA-YE; Difco), incubated at 35 °C for 18 h, and checked for purity by assessing typical colony morphology. An isolated colony from the primary TSA-YE plate was streaked onto a Tryptone Soy Agar (TSA; Difco) slant, incubated at 35 °C for 18 h, and then held at 4 °C for up to 1 month, as a working culture. For experimentation, a loopful from the working culture of each isolate was streaked onto TSA-YE and incubated for 18 h at 35 °C before use.
Food Samples
The pilot study was completed with frozen cooked shrimp, and for the validation study frozen mixed vegetables, frozen cooked shrimp and frozen cooked meatballs were tested. The food products were either purchased from a retail establishment in Dartmouth, Nova Scotia, or received as laboratory retail samples by the CFIA Dartmouth Microbiology Laboratory, as part of routine inspection programs. The frozen foods were stored below −15 °C until used.
Heat-Stressing (Health Canada 2011) of L. monocytogenes Strain MI-0497
Isolated colonies of L. monocytogenes from a freshly grown TSA-YE plate were transferred to 10 ml of ambient temperature sterile 0.45% saline (Remel) to produce a suspension of 3.0 McFarland units. This suspension was tenfold serially diluted in 0.1% peptone water (Difco), plated onto ACC Petrifilm™ (3 M™) and incubated at 35 °C for 48 h to retrospectively confirm the bacterial concentration in CFU/ml (Health Canada 2015a). Next, 10 ml of this 3.0 McFarland suspension was transferred into a 250-ml sterile Erlenmeyer screw-cap flask that had been pre-warmed to 55 °C. The flask was initially vortexed for 10 s and then incubated at 55 °C for 39 min in a water bath, mixing every 5 min. The flask was then cooled at ambient temperature (approx. 25 °C) for 20 min and vortexed for 10 s. The level of stress (%) was then evaluated as described below, and the heat-stressed inoculum was held at 4 °C for 2 days, until assessment of the level of stress was completed.
Freeze-Stressing (Wu et al. 2001a) of L. monocytogenes Strain MI-0493
A broth culture was prepared by inoculating one isolated L. monocytogenes colony from a freshly grown TSA-YE plate into 10 ml of Tryptone Soy Broth with 0.6% yeast extract (TSB-YE, Difco) in a 24-mm glass screw-cap tube, and incubating at 35 °C for 24 h. This overnight broth culture was diluted 1:100 in fresh TSB to achieve a cell density of approx. 107 CFU/ml, and then diluted again 1:100 by transferring 50 μl to a 24 mm glass screw-cap tube containing 5 ml of 0.1% peptone at ambient temperature. The tube (with approx. 105 L. monocytogenes CFU/ml) was closed tightly, vortexed for 10 s, placed in a −20 °C freezer for 2 h, and then thawed at ambient temperature (approx. 25 °C) for 2 h. The level of stress (%) was then evaluated as described below, and the freeze-stressed inoculum was kept at 4 °C for 2 days, until assessment of the % stress was completed.
Evaluation of Stress
Heat- or freeze-stressed L. monocytogenes were tenfold serially diluted in 0.1% peptone and spread plated onto non-selective TSA-YE agar and selective Oxford agar (OXF, Oxoid CM0856B agar base with supplement SR0140). Plates were incubated at 35 °C for 48 h and colonies were counted. The percent of injured cells was calculated as % stress = 100 − [(mean count on OXF / mean count on TSA-YE) × 100] (Health Canada 2011). The target was to achieve ≥50% stressed cells, which would grow on TSA-YE plates but not on selective OXF plates.
Method Development Phase
In method MFLP-74, OXF is listed as the mandatory plating agar for enumeration of L. monocytogenes (Pagotto et al. 2011b), and therefore, this was the agar selected for the development phase of the project. TSA was used as the nonselective medium for the additional agar layer (TAL). Two arrangements of OXF-TSA were prepared: single-step TAL and two-step TAL. For single-step TAL, a 14 ml aliquot of sterilized OXF agar was first poured into standard Petri dishes (100 × 15 mm) and allowed to solidify at room temperature. Then, a single sterilized TSA agar aliquot of 14 ml (tempered to 48 °C) was overlaid on the solidified OXF plate (Kang and Fung 1999). For two-step TAL, the initial 14-ml layer of OXF agar was overlaid with two 7-ml layers of TSA (Wu and Fung 2001), with each layer allowed to independently solidify and dry at room temperature. To determine the effect of holding time of the TAL agar on the ability to recover heat-stressed and non-stressed L. monocytogenes strain MI-0497 cells, three batches of each of the two presentations of OXF-TSA agar were prepared 1, 2 and 7 days prior to plating with an inoculum. Heat-stressed and non-stressed cell suspensions of L. monocytogenes were prepared (having an expected number of 106 CFU/ml) and then serially diluted in 0.1% peptone (having an expected number of 125–250 CFU/ml). A 1-ml aliquot of the cell suspensions were directly spread plated in triplicate on each of the six OXF-TSA agar types (emerging from the combination of two TAL presentations and three holding times) independently from any food product. One experiment was run using non-stressed cells and two experiments were run (each using three replicates for each condition) according to the method for L. monocytogenes isolation and enumeration described below in the context of the pilot study and Fig. 1. Heat-stressed and non-stressed cell suspensions were also plated in triplicate onto OXF and TSA-YE to serve as controls for each experiment.

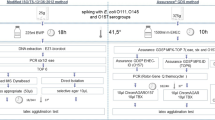
Diagram showing the sequential testing performed in parallel on frozen food items spiked with stressed L. monocytogenes. For the pilot study, OXF, PAL, OXF-TSA, and PAL-TSA were used, and for method validation OXF, RLM, OXF-TSA, and RLM-TSA were used
Pilot Study to Assess the Effectiveness of the TAL Modification
Health Canada Guidelines for the Evaluation of Quantitative Microbiological Methods (Health Canada 2015b) were used to design a pilot study for the effectiveness of a TAL modification of selective agars for the testing of one frozen food product. OXF and Palcam (PAL, Oxoid CM877 agar base with supplement SR150) were the selective agars chosen to enumerate L. monocytogenes. Results from the method development phase (described above) indicated that the performance of the single-step TAL was equivalent to the two-step TAL. Thus, the single-step TAL modification (14 ml TSA layered on 14 ml of selective agar) was adopted both for the pilot study and for method validation.
Only one type of frozen food product (pre-cooked shrimp) was used in the pilot study, which was spiked with artificially heat-stressed cells, to emulate a combination of high-temperature and freezing stresses potentially present in pre-cooked frozen foods. Three independent experiments, each consisting of 30 paired samples, were run. The 30 samples emerged from six replicates of five spiking levels (0, 0.5, 5, 50, and 500 L. monocytogenes CFU/g, designated as target inoculation levels A to E, respectively). To achieve these target inoculation levels, suspensions of stressed cells of L. monocytogenes strain MI-0497 were diluted according to assumed values of CFU/ml, and adjusted to 10× the target concentration, so that the inoculation levels (in CFU/g) could be delivered in 0.1 ml aliquots. These aliquots were then randomly dispensed onto the surface of 10 g of frozen product as small droplets. Aliquots (0.1 ml) of the inoculum were also directly spread in triplicate onto both TSA-YE and OXF plates, and incubated at 35 °C for 48 h. The actual CFU/ml values of the prepared inocula were retrospectively determined from the triplicate counts on TSA-YE, and used to calculate the inoculation levels (e.g., those reported in the third column of Tables 3 and 4). Ratios of the actual OXF/TSA-YE plate counts were used to calculate the percent stress of the inoculum for further reference. The spiked samples were then held below −15 °C for 2 weeks prior to sample analysis to emulate a possible real-life scenario for pre-cooked frozen foods.
Each of the 30 samples was processed as a 10 g portion in filtered stomacher bags (Whirl-pak™) and analyzed by both the reference method MFLP-74, and alternative MFLP-74 method with TAL modification, so that paired results were obtained for each sample (paired sample). Therefore, a total of 90 paired samples (30 paired samples/run × 3 independent runs) were processed. Figure 1 shows the general experimental workflow followed to isolate and enumerate L. monocytogenes. Briefly, 40 ml of 0.1% peptone was added to the 10 g samples (to achieve a 1:5 dilution) and stomached for 2 min (AES, Smasher). Immediately, 1 ml of the diluted sample was plated over triplicate plates (i.e., 3 plates per dilution, at ~0.333 ml/plate) for each of the four agar types tested (OXF, OXF-TSA, PAL, PAL-TSA). That is, 4 × 1-ml aliquots per dilution were plated on a total of 12 plates (3 plates of each of the four agar types tested). Plates were incubated for 48 ± 4 h at 35 °C, and then typical L. monocytogenes colonies (black colonies on OXF and OXF-TSA, and dark gray-green or black colonies on PAL and PAL-TSA) were enumerated. Positive colonies were confirmed as described in MFHPB-30 (Pagotto et al. 2011a) by scoring hemolysis, carbohydrate utilization (xylose, rhamnose, mannitol) and motility. Results were recorded separately for each of the four agars, but only the results from the agar type with the largest number of colonies were used to calculate the final CFU/g (PAL vs OXF, or PAL-TSA vs OXF-TSA), as prescribed in method MFLP-74 (Pagotto et al. 2011b).
Blank cultures, as well as positive and negative control strains (Table 1) were employed as specified in method MFLP-74. When no L. monocytogenes colonies were found in the lowest dilution plated, samples were reported as having <5 CFU/g. Health Canada method MFHPB-33 (Health Canada 2015a) was used to analyze a sample (10 g) of the frozen food tested, to determine the level of background microbial aerobic counts (AC).
Method Validation and Selectivity Assessment of TAL-Modified MFLP-74
Health Canada Guidelines for the Evaluation of Quantitative Microbiological Methods (Health Canada 2015b) were used to conduct the validation of the TAL modification for method MFLP-74, to be used at the CFIA for the testing of frozen foods. The study design, sample preparation, spiking, calculated inoculation levels, percent stress, and enumeration were as described above for the pilot study with the following exceptions and changes: First, three types of frozen food products were used, vegetables, pre-cooked shrimp, and pre-cooked meatballs. Second, the frozen vegetable samples were spiked with artificially freeze-stressed cells of L. monocytogenes strain MI-0493, whereas the pre-cooked meatball samples were spiked with artificially heat-stressed cells of strain MI-0497. Third, method validation samples were tested as five replicates of six spiking levels (0, 5, 50, 100, 500, and 1500 L. monocytogenes CFU/g of food, designated as target inoculation levels A to F, respectively). Fourth, the inocula of stressed L. monocytogenes were prepared in a 1-ml total volume (instead of the 0.1 ml used in the pilot study). Fifth, Rapid’L.mono (RLM, Bio-Rad #356–3694, on which L. monocytogenes produces blue colonies) and RLM-TSA were selected as the second agars (besides OXF and OXF-TSA) to enumerate L. monocytogenes in the MFLP-74 and TAL modification methods, respectively. Therefore, the result from the agar type with the largest number of colonies (either OXF vs RLM, or OXF-TSA vs RLM-TSA) was used to calculate the final CFU/g, as prescribed in method MFLP-74 (Pagotto et al. 2011b). Sixth, for enumeration, dilution-plated RLM and RLM-TSA were incubated for 48 ± 4 h at 37 °C (instead of 35 °C used in the pilot study for OXF and PAL). Seventh, in contrast to the pilot study that had three runs per food sample, the validation study had only one run for each of three frozen food products (the total number of paired samples being kept at 90).
Method validation also included a selectivity assessment consisting of an inclusivity study (ability of the alternative method to detect the target organism) and an exclusivity study (lack of interference on the alternative method by a relevant range of non-target organisms), which were performed according to Health Canada guidelines (Health Canada 2015b). The selectivity assessment was conducted on OXF, OXF-TSA, RLM and RLM-TSA agars directly inoculated with the non-stressed test organisms, independently from any food product. Pure cultures of the selectivity strains (listed in Online Resource, 1 ESM_1.pdf) were prepared from frozen stock cultures and working seed cultures, as described above in the Bacterial Strains and General Culture Conditions section. A single colony from a fresh TSA-YE plate of each strain tested was inoculated into a tube containing 4 ml of Brain Heart Infusion (BHI) broth. The tubes were then incubated at 35 °C overnight (18–24 h), followed by dilution to adjust the cell concentration to 100 CFU/ml for target strains, and 1000 CFU/ml for non-target strains. These adjusted cultures were then plated, 1 ml on triplicate plates (i.e., 3 plates per dilution, at ~0.333 ml/plate) on the four selective agars tested, incubated for 48 ± 4 h at 37 °C, and analyzed for the presence or absence of colonies showing the typical morphology for L. monocytogenes. Colonies were also characterized as prescribed in method MFHPB-30 (Pagotto et al. 2011b) for positive or negative reactions on carbohydrate utilization, hemolysis, and motility.
Statistical Analysis
All enumeration data were transformed to log10 CFU/g. Un-inoculated samples or samples with a result of <5 L. monocytogenes CFU/g were arbitrarily considered to have 1 CFU/g and were thus represented as log10 1 = 0, for statistical evaluation. Before performing any statistical significance tests, linearity of correlative data was visually verified through a scatter diagram, and the overall relationship between methods (and that the points were randomly distributed within the groups) was assessed. The Grubb’s outlier test was used to ensure that there were no outlying points. The reference method (MFLP-74) was compared to the TAL modification, for the ability to recover and enumerate stressed L. monocytogenes cells by means of a linearity test, and a linear regression analysis (95% confidence interval) was performed for each food item.
Two statistical tests were used to evaluate the significance of the differences between methods. A paired Student’s t test was used to directly analyze differences in sets of paired samples (development phase, pilot study and method validation data). For the latter two, significance was determined by comparing the means for each method by experimental run (n = 30) and for all three runs combined (n = 90) for each frozen food product. A one-way ANOVA test, performed with Tukey’s Contrasts, was used for multiple comparisons between data not necessarily paired, e.g., comparisons between OXF, RLM, OXF-TSA and RLM-TSA in the validation study, or between single-step and two-step TAL agars. The minimum level of significance was set for p < 0.05, (α = 0.05). Statistical analyses were completed with Microsoft Office Excel (Data Analysis ToolPack 2010) and R Commander (Fox 2005, version 3.2.3).
Results and Discussion
Stress Levels of the Inocula
Health Canada stipulates that for the evaluation of processed foods by quantitative microbiological methods, microorganisms must be stressed before inoculation, equilibration and analysis (Health Canada 2015b). The recommendation is for samples to be spiked with test microorganisms that are injured by exposure to sub lethal treatments that simulate, as close as possible, the food processing conditions (Health Canada, 2011). Frozen fresh vegetables (where freezing is the primary source of stress) were spiked only with artificially freeze-stressed L. monocytogenes, whereas pre-cooked frozen products were spiked with artificially heat-stressed L. monocytogenes, and subsequently frozen. The aim was to achieve ≥50% stress in the inocula, which was on target for all the experimental runs. The stress levels of the L. monocytogenes inocula used in the two experiments run for the development phase were 67 and 56%. The L. monocytogenes inocula utilized to spike the frozen cooked shrimp in the three independent experiments of the pilot study were 92, 87, and 67% stressed. The inocula used to spike the food samples during method validation were 54, 72, and 79% stressed, for frozen vegetables, frozen cooked shrimp, and frozen cooked meatballs, respectively.
The Aerobic Microflora of Frozen Foods Did Not Interfere with TAL Agar Performance
Enumeration of L. monocytogenes in selective media can be complicated by the growth of background microflora (Golden et al. 1988). The non-selective TSA layer of TAL agars could further complicate enumeration and interfere with the identification of L. monocytogenes by potentially favoring the overgrowth of background organisms. Frozen vegetables and frozen shrimp samples had a mean AC of 150 and 440 CFU/g, respectively, whereas frozen meatballs had a mean AC of <3.3 CFU/g. Overall, only a low number (≤50 CFU/ml) of non-target colonies were observed in the 1 ml aliquots plated on the TAL agars from the frozen vegetable and frozen shrimp samples, suggesting that the antimicrobial compounds from the selective agar plate effectively diffused into the TSA layer and inhibited the growth of background microflora, on time to prevent overgrowth. Often, due to the low concentration of L. monocytogenes cells in relation to unknown levels of various background organisms, enumeration by direct plating methods may be ineffective, dependent on the type of food (Golden et al. 1988; Besse and Colin 2004).
The Single-Step OXF-TSA Constitutes a Simple and Practical TAL Modification
In the development phase of the work, two arrangements of OXF-TSA agar and three holding times were assessed for their ability to induce repair of stressed L. monocytogenes cells. Overall, the mean OXF-TSA counts for all paired conditions were significantly different from the mean OXF counts (p < 0.0001, α = 0.05, Table 2), suggesting that OXF-TAL permitted the recovery of stressed L. monocytogenes cells. No significant differences (α = 0.05) were found between the mean counts on TSA-YE, OXF-TSA and OXF agars for recovery of non-stressed cells (Table 2), indicating that OXF-TSA does not favor the growth of non-stressed cells in a non-specific manner. It is, therefore, reasonable to surmise that the significantly higher counts of stressed cells observed on OXF-TSA in relation to OXF are due to the specific resuscitation of stressed cells, rather than to a non-specific nutritional or growth effect that favors the growth of both stressed and non-stressed cells.
When the mean counts of heat-stressed cells from the single-step OXF-TSA agar (6.39 ± 0.22 Log10CFU/g, n = 6 means) were compared with the mean counts from the two-step OXF-TSA agar (6.55 ± 0.06 Log10CFU/g, n = 6 means), no significant differences were found by applying a paired Student’s t test (p = 0.18, α = 0.05). Similarly, a one-way ANOVA test showed that there were no significant differences between the mean enumeration results using heat-stressed cells obtained from the OXF-TSA agars prepared 1 day (6.32 ± 0.23 Log10CFU/g, n = 6 means), 2 days (mean = 6.55 ± 0.06 Log10CFU/g, n = 6 means) and 7 days (6.52 ± 0.04 Log10CFU/g, n = 6 means) before testing (p = 0.15, α = 0.05), indicating that the selective overlay agar can be prepared in advance and held for up to 7 days before plating, with no deleterious effect on the recovery of injured L. monocytogenes. Although Wu and Fung (2001) reported that two 7-ml layers of TSA worked better than a single 14-ml layer for slowing down the selective agents’ migration into the TSA upper layers, Hajmeer et al. (2001) found that the double-step TAL did not have a practical advantage over the single-step TAL in the recovery of injured E. coli O157:H7. Those authors also found that TAL agars held for up to 7 days would perform similarly to 1-day-old agars (Hajmeer et al. 2001). The data showing the growth of L. monocytogenes strain MI-0497 on OXF-TSA plates, therefore, aligns well with the findings of Hajmeer et al. (2001).
Since the goal was to develop a TAL modification that was practical and efficient, the single-step TAL modification was selected for all subsequent experiments. It was decided not to develop methods that use <5 ml thin agar layers (e.g., Kang and Fung 1999) because it is difficult to obtain a uniform and leveled coverage of the solidified selective agar. In addition, overlaying a pre-solidified agar layer of TSA on a solidified plate of selective agar (Yan et al. 2006) was regarded as highly impractical in regulatory testing laboratories, particularly when large batches of media are required and the handling of the pre-solidified non-selective agar layer would increase the likelihood of contamination.
The TAL Modification Enhances the Recovery of Stressed L. monocytogenes Cells
Results from the pilot study (Table 3) and method validation (Table 4) clearly indicated that the TAL modification of the three selective agars tested provided an advantageous resuscitation step to enhance the recovery of stressed cells.
The mean counts from the three independent experimental runs of the pilot study are presented separately in Table 3. The mean counts listed by agar type can be found in supplemental Online Resource 2 (ESM_2.pdf). The linear relationship shown in Fig. 2 indicates a good correlation between standard agars vs TAL-modified agars. The linear correlation coefficients (R 2) for the three runs of frozen cooked shrimp were 0.977, 0.996, and 0.994, respectively (and 0.980 for all the data combined), suggesting that both methods responded well to the spiked levels of the target organism. No outlying values were identified for the data of either the TAL modification or the standard method. However, the paired Student’s t test demonstrated that L. monocytogenes counts differed significantly between the two methods (t = 7.49, p < 0.0001, α = 0.05), with higher counts being consistently registered on the TAL agars. When the combined data from the three runs was compared, the number of L. monocytogenes cells recovered on TAL agars (mean = 1.21 Log10CFU/g, n = 84) was overall 23.4% greater than the cells recovered on the standard agars (mean = 0.92 Log10CFU/g, n = 84).

Scatter plot showing the linearity of the pilot study data, comparing mean enumeration results of stressed L. monocytogenes by the TAL method (vertical axis) relative to the reference method MFLP-74 (horizontal axis) for three trials with frozen cooked shrimp. Upper error bar = One standard deviation above the mean values (n = 6) for the TAL method, lower error bar = One standard deviation under the mean values (n = 6) for the reference method. The linear regression equation and the correlation coefficient for the line shown are given at the top of the plot
The pilot study indicated that the TAL modification warranted formal validation. For this, two additional frozen foods were included and PAL was exchanged for RLM as the second selective agar, besides OXF. This was prompted by the fact that both OXF and PAL do not distinguish between Listeria species, whereas RLM allows a rapid visual distinction between L. monocytogenes and Listeria spp. In addition, RLM is the routine second agar used at the CFIA for methods MFLP-74 and MFHPB-30. Table 4 shows the comparative mean counts from MFLP-74 and TAL-modified MFLP-74, separated by food type. The mean counts listed by agar type can be found in supplemental Online Resource 2 (ESM_2.pdf). The linear relationship between the two methods (Fig. 3) indicated a good correlation, with R2 = 0.984, 0.989, and 0.997 for frozen vegetables, frozen cooked shrimp, and frozen cooked meatballs, respectively, and R 2 = 0.985 for all data combined. No outlying values were identified for the data of either MFLP-74 or the TAL-modified MFLP-74. For all data combined, the paired Student’s t test showed a significant difference (t = 7.80, p < 0.0001, α = 0.5) between the two methods, with MFLP-74 consistently giving lower bacterial counts. Statistically significant differences were also observed when each food item was evaluated individually: frozen vegetables (t = 5.02, p < 0.0001, α = 0.5), frozen cooked shrimp (t = 3.55, p < 0.001, α = 0.5), and frozen cooked meatballs (t = 5.45, p < 0.0001, α = 0.5).

Scatter plot showing the linearity of validation study data, comparing mean enumeration results of stressed L. monocytogenes by the TAL method (vertical axis) relative to the reference method MFLP-74 (horizontal axis) for the three food items indicated in the graph legend. Upper error bar = One standard deviation above the mean values (n = 5) for the TAL method, lower error bar = One standard deviation under the mean values (n = 5) for the reference method. The linear regression equation and the correlation coefficient for the line shown are given at the top of the plot
TAL Agars Are Effective Regardless of the Selective Media Used
Golden et al. (1988) observed that colonies formed by non-injured and freeze-injured L. monocytogenes cells exhibited no detectable morphological differences, regardless of the selective agar tested, albeit colonies arising from heat-injured cells were smaller than those from non-injured cells. Furthermore, Kang and Fung (1999) noted that the additional TSA layer did not hinder black color production by heat-injured L. monocytogenes cultured on modified Oxford medium (MOX), and Wu et al. (2001a) reported that cold-injured L. monocytogenes cells formed typical colonies on MOX-TSA (Wu et al. 2001a, b). In the work presented, it was observed that stressed L. monocytogenes produced positive reactions on TAL-modified OXF-TSA, PAL-TSA and RLM-TSA that were indistinguishable from those produced by non-injured control cells, as well as very similar to those produced by non-injured and stress-injured cells on standard OXF, PAL and RLM agars. The only changes noted when the TAL agars were compared to the standard selective agars, were slightly muted color presentations on OXF-TSA (presumptive black colonies with a black halo and sunken center) (Fig. 4a, b), presumptive blue colonies without a yellow halo on RLM-TSA (Fig. 4c, d), and presumptive colonies with a gray-green coloration, a black sunken center, and a black halo on a cherry-red background on PAL-TSA (not shown).

Photographic composite showing the morphology of colonies formed by heat-stressed cells of L. monocytogenes recovered on standard and TAL-modified agar media used in this study. A OXF, B OXF-TSA, C PALCAM, D PALCAM-TSA, E RLM, and F RLM-TSA. Replicate aliquots of the same sample dilution were plated in A and B, C and D, and in E and F. Insets showing colony close-ups (a to f) are enlargements of the areas marked by the dotted lined squares shown in A to F
To determine whether the performance of the TAL modification depended on the selective agar utilized, or if a particular agar would outperform others, a one-way ANOVA test was conducted where results from the method validation were grouped using Tukey’s contrasts for pairwise comparisons of counts on OXF, RLM, OXF-TSA, and RLM-TSA, and for the three frozen food items tested. Assessing all data combined, but excluding results for the non-spiked samples (inoculation level A), a significant difference was observed between OXF and RLM-TSA (p = 0.02, α = 0.05, CI = 0.038–0.892), but not between the other agar types (Fig. 5). The analysis of enumeration counts paired by inoculation level (Table 5) showed a trend of a significant difference between more than one combination of agars as the inoculation level increased. At the highest inoculation level (1500 CFU/g), the only non-significant difference was between RLM and OXF-TAL (p = 0.996, α = 0.05, CI, −0.149-0.177). The combined data suggested that, as expected, enumeration counts varied in an agar- and inoculum size-dependent manner, and that the use of RLM-TSA has the highest likelihood of providing the best estimation of L. monocytogenes levels.

Bar graph showing mean recovery of stressed L. monocytogenes, as Log10 CFU/g, on OXF, RLM, OXF-TSA and RLM-TSA agars (Agar Type) for the combined three food items used in the validation study. Different letters (above the bars) indicate significant differences between media, p < 0.05, α = 0.05. Error bars represent one standard deviation above the mean (n = 75). Note: Data for the non-spiked samples (0 CFU/g = inoculation level A) were excluded from the analysis
Finally, the results from the inclusivity study showed that all 50 L. monocytogenes strains (listed in Online Resource ESM_1.pdf) produced colonies with typical morphologies on OXF-TSA and RLM-TSA. In addition, all 50 strains grown on the TAL agars had positive carbohydrate utilization, hemolysis and motility reaction tests. The exclusivity study showed that OXF-TSA and RLM-TSA agars suppressed the growth of non-L. monocytogenes strains (listed in Online Resource 1 ESM_1.pdf) as effectively as OXF and RLM. It seems that despite the presence of the non-selective TSA layer, the selective agents were effective inhibitors of non-target microorganisms, with the exception of Rhodococcus equi which was able to grow on OXF-TSA and RLM-TSA but not on OXF and RLM. However, R. equi colonies were atypical, suggesting that the differential components of the selective agar were still effective in the TAL. It should be noted that although Staphylococcus aureus and Staphylococcus epidermidis were able to grow and give a non-typical reaction on OXF-TSA, these two organisms also grew on OXF agar to similar extents. As expected, non-target Listeria spp. gave typical reactions on OXF-TSA (presumptive black colonies with a black halo and sunken center with a slightly muted color reaction), but were later confirmed as negative through the carbohydrate utilization, hemolysis and motility tests. Non-target Listeria spp. produced negative presumptive colonies on RLM-TSA, confirming that the differential agents from the RLM layer diffused through the non-selective layer, to differentiate between target and non-target colonies. These results indicate that OXF-TSA and RLM-TSA are fit for purpose and, therefore, meet the performance criteria required for quantitative methods, as specified in the Health Canada quantitative guidelines (Health Canada 2015a).
Food preservation by freezing slows down chemical changes and prevents microbial growth, thereby extending the shelf life of products (reviewed by Berry et al. 2009). Frozen vegetables are not regarded as high-risk foods and are rarely associated with food-borne outbreaks since pathogens such as L. monocytogenes are unable to proliferate at freezing temperatures, and cooking is recommended prior to consumption (Berry et al. 2009; Health Canada 2011). Nevertheless, a recent large-scale voluntary recall of frozen fruits and vegetables (including 358 consumer products sold under 42 separate brands, and at least 98 other products recalled by other firms) was undertaken in the USA, as routine testing detected L. monocytogenes (FDA 2016a). In fact, there were listeriosis cases later confirmed to be associated with these recalled products (FDA 2016b). As a result, food recall warnings were issued in Canada, and the CFIA conducted a food safety investigation, which eventually led to the recall of additional frozen products (14 recall warnings for a variety of frozen products and brands) (CFIA 2016). Moreover, in early 2015, an outbreak of invasive listeriosis linked to ice cream products was identified in the USA (CDC 2015). Although the vast majority of the population did not become ill after ingesting a low level of L. monocytogenes in the contaminated ice cream (Pouillot et al. 2015), the aforementioned cases stress the importance of having a method that accurately detects low-levels of L. monocytogenes contamination (including stressed and non-stressed bacterial cells) in frozen products, typically regarded as low-risk due to the fact that they do not support the growth of this pathogen.
Conclusion
The experimental data presented supports the working hypothesis formulated for this study, i.e., that method MFLP-74 (which lacks a provision for resuscitation of stressed cells) would underestimate the number of L. monocytogenes cells actually present in frozen food products. The TAL modification emerged as a simple, effective and very practical method modification that allowed the repair of stress-injured L. monocytogenes cells and enhanced their recovery from frozen food products. Enhanced recovery of L. monocytogenes is particularly relevant in foods contaminated near the Canadian action level, as this could mean the regulatory acceptance or rejection of an unsafe food product, depending on whether the standard MFLP-74 method or the TAL-modified method is utilized, respectively. A practical scenario of such importance was recently provided by the 2016 Listeria outbreak linked to frozen fruits and vegetables in the USA, which also triggered a national recall in Canada. Although this particular outbreak recall was not based on Listeria enumeration results, an accurate enumeration method is important for the regulatory testing of frozen foods which may be contaminated with L. monocytogenes near the Canadian regulatory action level of 100 CFU/g. Therefore, it is recommend that the TAL modification of method MFLP-74 be adopted in the regulatory testing of frozen food products in Canada, on the understanding that additional validation work could expand the scope of assessment to other RTE foods for which CFIA conducts routine L. monocytogenes enumeration.
References
Aureli P, Fiorucci GC, Caroli D, Marchiaro G, Novara O, Leone L, Salmaso S (2000) An outbreak of febrile gastroenteritis associated with corn contaminated by Listeria monocytogenes. N Engl J Med 342:1236–1241
Berry M, Fletcher J, McClure P, Wilkinson J (2009) Effects of freezing on nutritional and microbiological properties of foods. In: Frozen Food Science and Technology. pp 26–50
Besse NG (2002) Influence of various environmental parameters and of detection procedures on the recovery of stressed L. monocytogenes: a review. Food Microbiol 19:221–234
Besse NG, Colin P (2004) Enumerating listeria monocytogenes in food: a problem of low numbers. J Rapid Methods Autom Microbiol 12:83–105
CDC (2015) Multistate outbreak of listeriosis linked to Blue Bell Creameries products http://www.cdc.gov/listeria/outbreaks/ice-cream-03-15/. (retrieved 2016–05-11)
CFIA (2016) Food Recall Warning – Harmonie brand, ICA brand, Co-op brand, Western Family brand and unbranded frozen vegetable products recalled due to Listeria monocytogenes http://www.inspection.gc.ca/about-the-cfia/newsroom/food-recall-warnings/complete-listing/2016-05-07/eng/1462667105675/1462667109404 (retrieved 2016-05-11)
Dalton CB, Austin CC, Sobel J, Hayes PS, Bibb WF, Graves LM, Swaminathan B, Proctor ME, Griffin PM (1997) An outbreak of gastroenteritis and fever due to Listeria monocytogenes in milk. N Engl J Med 336:100–105
Farber JM, Peterkin PI (1991) Listeria monocytogenes, a food-borne pathogen. Microbiol Rev 55:476–511
Food and Drug Administration (FDA) (2016a) CRF Frozen Foods Expands Voluntary Recall to Include All Frozen Vegetable and Fruit Products Due to Possible Health Risk http://www.fda.gov/Safety/Recalls/ucm497297.htm (retrieved 2016-09-24)
Food and Drug Administration (FDA) (2016b) FDA Investigated Listeria Outbreak Linked to Frozen Vegetables https://www.fda.gov/Food/RecallsOutbreaksEmergencies/Outbreaks/ucm499157.htm (retrieved 2016-09-24)
Fox J (2005) The R commander: a basic statistics graphical user interface to R. J Stat Softw 14(9):1–42
Golden DA, Beuchat LR, Brackett RE (1988) Evaluation of selective direct plating media for their suitability to recover uninjured, heat-injured, and freeze-injured Listeria monocytogenes from foods. Appl Environ Microbiol 54:1451–1456
Hajmeer MN, Fung DYC, Marsden JL, Milliken GA (2001) Effects of preparation method, age, and plating technique of thin agar layer media on recovery of Escherichia coli O157:H7 injured by sodium chloride. J Microbiol Methods 47:249–253
Health Canada (2011) Procedure for the Development and Management of Food Microbiological Methods. Annex 4.2: Procedure for stressing microorganisms in artificially contaminated foods. Compendium of Analytical Methods. Health Products and Food Branch, Ottawa, Canada
Health Canada (2015a) MFHPB-33: Enumeration of total aerobic bacteria in food products and food ingredients using 3M™ Petrifilm™ Aerobic Count Plates. Compendium of Analytical Methods, Vol. 2. Health Products and Food Branch, Ottawa, Canada
Health Canada (2015b) Procedure for the Development and Management of Food Microbiological Methods. Part 3: Guidelines for the evaluation of quantitative food microbiological methods. Compendium of Analytical Methods, Vol. 1. Health Products and Food Branch, Ottawa, Canada
ISO (1998) ISO 11290-2:1998, Microbiology of food and animal feeding stuffs - horizontal method for the detection and enumeration of Listeria monocytogenes - part 2: enumeration method. International Organization for Standardization, Geneva
Kang D-H, Fung DYC (1999) Thin agar layer method for recovery of heat-injured Listeria monocytogenes. J Food Prot 62:1346–1349
Lavieri NA, Sebranek JG, Cordray JC, Dickson JS, Jung S, Manu DK, Mendoça AF, Brehm-Stecher BF, Stock J, Stalder KJ (2014) Evaluation of the thin agar layer method for the recovery of pressure-injured and heat-injured Listeria monocytogenes. J Food Prot 77:828–831
Pagotto F, Hébert K, Farber J (2011a) MFHPB-30: isolation of Listeria monocytogenes and other Listeria spp. from foods and environmental samples. Compendium of analytical methods. Vol. 3. Health products and food branch, Ottawa, Canada
Pagotto F, Trottier Y, Upham J, Iugovaz I (2011b) MFLP-74: enumeration of listeria monocytogenes in foods. Compendium of analytical methods, Vol. 3. Health products and food branch, Ottawa, Canada
Pouillot R, Klonz K, Chen Y, Burall L, Macarisin D, Doyle M, Bally KM, Strain E, Datta AR, Hammack TS, Van Doren JM (2016) Infectious dose of Listeria monocytogenes in outbreak linked to ice cream, United States, 2015 Emerg Infect Dis 22:2113–2119
Schlech WF III (1997) Listeria gastroenteritis—old syndrome, new pathogen. N Engl J Med 336:130–132
Schlech WF III (2000) Epidemiology and clinical manifestations of Listeria monocytogenes infection. In: Fischetti VA, Novick RP, Ferretti JJ, Portnoy DA, Rood JI (eds) Gram-positive pathogens. American Society of Microbiology, Washington, D.C., pp 473–479
Schlech WF III, Lavigne PM, Bortolussi RA, Allen AC, Haldane EV, Wort AJ, Hightower AW, Johnson SE, King SH, Nicholls ES, Broome CV (1983) Epidemic listeriosis—evidence for transmission by food. N Engl J Med 308:203–206
Wesche AM, Gurtler JB, Marks BP, Ryser ET (2009) Stress, sublethal injury, resuscitation, and virulence of bacterial foodborne pathogens. J Food Prot 72:1121–1138
Wu VCH (2008) A review of microbial injury and recovery methods in food. Food Microbiol 25:735–744
Wu VCH, Fung DYC (2001) Evaluation of thin agar layer method for recovery of heat-injured foodborne pathogens. J Food Sci 66:580–583
Wu VCH, Fung DYC, Kang DH (2001a) Evaluation of thin agar layer method for recovery of cold-injured foodborne pathogens. J Rapid Methods Autom Microbiol 9:11–25
Wu VCH, Fung DYC, Kang DH, Thompson LK (2001b) Evaluation of thin agar layer method for recovery of acid-injured foodborne pathogens. J Food Prot 64:1067–1071
Yan Z, Gurtler JB, Kornacki JL (2006) A solid agar overlay method for recovery of heat-injured Listeria monocytogenes. J Food Prot 69:428–431
Yuste J, Capellas M, Pla R, LLorens S, Fung DYC, Mor-Mur M (2003) Use of conventional media and thin agar layer method for recovery of foodborne pathogens from pressure-treated poultry products. J Food Sci 68:2321–2324
Acknowledgments
The authors gratefully acknowledge Jacqueline Upham and Jodi Elliot for contributions made during planning of the preliminary experiments and Elizabeth Boutilier for providing technical assistance during the selectivity study.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Funding
This work was funded by internal research grants provided by the Canadian Food Inspection Agency.
Conflict of Interest
Julie McLennon declares that she has no conflict of interest. Antonela Borza declares that she has no conflict of interest. Mikaela Eisebraun declares that she has no conflict of interest. Rafael Garduño declares that he has no conflict of interest.
Ethical Approval
This article does not contain any studies with human participants or animals performed by any of the authors.
Informed Consent
Informed consent is not applicable in this study.
Rights and permissions
About this article

Cite this article
McLennon, J., Borza, A., Eisebraun, M. et al. Improved Recovery of Stressed Listeria monocytogenes from Frozen Foods. Food Anal. Methods 11, 403–414 (2018). https://doi.org/10.1007/s12161-017-1011-2
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s12161-017-1011-2