
Abstract
Hydrogen is a renewable fuel that can be biologically produced by green algae in closed anaerobic photobioreactors with light and organic carbon as energy sources. This research aimed to investigate the influence of sulfur concentration and light intensity on hydrogen and ethanol production, as well as on nutrient removal by Chlamydomonas reinhardtii (CC425) in batch cultures. The strain was cultivated in two phases: in the first step, the cultures were maintained under aerobic conditions to obtain biomass; in the second step, the biomass was transferred to closed anaerobic photobioreactors for gas generation under continuous illumination. A factorial design was accomplished to improve the biomass production in the first step, with light variation, pH, and glucose addition. Afterward, light intensity and sulfur concentration were varied to test hydrogen production in the second step. The best hydrogen production occurred in photobioreactors without sulfur addition (average increase of 7 times in the production) and under higher light intensity the productivity was 37% higher than lower light intensity (39.64 ± 2.44 µmol H2 L−1 h−1). There was an effect of sulfur concentration in the ethanol production and under higher light intensity the production was higher (203.20 ± 31.49 mg L−1). Furthermore, under conditions with the presence of sulfur, there was greater removal of ammoniacal nitrogen (5.3%), phosphate (85.0%), COD (9.1%) and acetic acid (97.2%). This research demonstrates the efficient production of hydrogen by C. reinhardtii and it shows that the process can be associated with ethanol production and nutrient removal.
Similar content being viewed by others

Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
Microalgae are a highly diversified group of photoautotrophic microorganisms, which grow quickly when adapted to the environment and can be cultivated in different conditions from open lagoons and even in closed photobioreactor systems [1]. They have the ability to accumulate lipids and sugars in their biomass, depending on the species and cultivation conditions, which is a good option for biofuel production, such as biodiesel, obtained from lipids in biomass, methane, from the biodigestion of algal biomass, ethanol by the fermentation of sugars and hydrogen from direct or indirect biophotolysis [1,2,3,4]. The advantage of using hydrogen as an energy source is because it is renewable and clean, generating only water in its combustion, and it is harmless to living beings and the environment [3].
Regarding the hydrogen production by microalgae, this biogas can be produced through the enzyme hydrogenase by direct or indirect biophotolysis pathways. Direct biophotolysis occurs from the photolysis of water by photosystem II (PSII) in the chloroplast during photosynthesis, releasing oxygen, electrons and protons which are directed through the plastoquinone to photosystem I (PSI). The electrons in PSI pass through a transport chain to the final electron acceptor, ferredoxin, which reduces NADP to NADPH which is then oxidized in the CO2 fixation reactions in the Calvin cycle [5]. Ferredoxin may also transport electrons to the hydrogenase, where protons are converted into molecular hydrogen [6]. However, hydrogenase is an oxygen-sensitive enzyme and the presence of oxygen can inhibit hydrogen production [7].
Therefore, oxygen evolution needs to be minimized and the indirect biophotolysis is an alternative to hydrogen production. In indirect biophotolysis, hydrogen production occurs from oxidation of organic carbon under anaerobic conditions and directs the flow of electrons through ferredoxin to the PSI (photofermentation) or to the fermentative pathway in dark (dark fermentation). Thus, the indirect pathway in the PSI occurs regardless of PSII [5, 6, 8, 9]. Furthermore, different types of ferredoxins and genes associated with the expression of the hydrogenase structure have been identified in C. reinhardtii [10, 11], and their expression interferes with hydrogen production in different algal strains.
Thus, for the continuous production of hydrogen the formation of an anaerobic environment needs to be induced. Several researchers have reported that nutritional deprivation, mainly of sulfur, supports the achievement of anaerobiosis [12, 13], as this element is a structural part of PSII proteins in the chloroplast and its deprivation causes a decrease in photosynthetic activity [6, 7, 13]. Consequently, there is a decrease in oxygen production during photosynthesis which is then consumed by respiration, a function that is maintained under these conditions. Therefore, there is compensation between oxygen consumption and production, and the algal metabolism is responsible for providing an anaerobic environment and consequent hydrogen production [7].
However, for hydrogen production by indirect biophotolysis to occur and in an anaerobic environment, it is necessary to grow the microalgae heterotrophically. Some green algae, such as Chlamydomonas, have this ability and can grow using acetate as the only organic carbon source, in addition to having a versatile fermentative metabolism [9, 14] and the potential for production of biofuels under autotrophic, mixotrophic or heterotrophic conditions [6, 15].
Therefore, considering the presence of specific cellular metabolism of Chlamydomonas microalgae and the compensatory point between respiration and photosynthesis provided by sulfur deprivation, there are several factors that can interfere and are important in hydrogen production, such as pH [16], nutritional stress by phosphorus or nitrogen [17], as well as light intensity and the effect of low sulfur concentrations, which are being investigated in order to the improve the process [8, 13, 18]. Furthermore, some researchers reported that a greater amount of biomass of microalgae allows higher hydrogen production efficiency [6,7,8, 19].
Moreover, the versatile fermentative metabolism of Chlamydomonas and its ability to use organic molecules such as acetate, allows this microalgae to generate other products, such as organic acids and ethanol, through fermentation pathways [7, 8, 20]. Thus, since part of the anaerobic metabolism occurs in the chloroplast, the electron flow can be directed to the PSI to reduce hydrogen or to form fermentation by-products, in the process of photofermentation [6, 9]. In addition to being potential hydrogen and by-product producers, green algae can also be used in wastewater treatment, mainly when they are associated with bacteria [21, 22], for example. In anaerobic digestion, acidogenic bacteria can break down the organic matter into organic acids, such as acetate, by acetogenesis process [23], and microalgae can use this acetate to grow heterotrophically and produce hydrogen by indirect biophotolysis [9, 14] and assist in removing nutrients.
Therefore, the purpose of this research was to improve biomass production of C. reinhardtii (CC425) and to evaluate the influence of sulfur concentration and light intensity in hydrogen production in batch anaerobic photobioreactor closed systems under continuous illumination, as well as to evaluate the by-product production, such as ethanol and organic acids, and the nutrient removal efficiency by this organism to contribute to future research and applications of environmental and economic interest.
Material and Methods
Strain and Inoculum Maintenance
The strain of Chlamydomonas reinhardtii CC425 (cw15 arg2 sr-u-2–60 mt +) was provided by the National Renewable Energy Laboratory. The strain was maintained in axenic culture Tris–acetate-phosphate, TAP medium [24], with arginine supplementation (100 mg L−1), at 24 ± 0.5 °C, pH 7.2, 12 h light/dark cycle, and light intensity of 60 μmol m−2 s−1 obtained with cool white fluorescent tubular lamps in an air-conditioned room. The TAP medium has acetic acid as a source of organic carbon at a concentration of 17.5 mmol L−1 and 506.5 µmol L−1 of sulfur.
Biomass Improvement Experiment: Factorial Design
In order to improve the biomass concentration, a factorial design with a central point (24 + 1) was used to generate a matrix with 17 experiments. Experiments were carried out in TAP medium [24] under continuous illumination at two levels of temperature (24 and 32 °C), pH (6.2 and 8.2), light intensities (200 and 500 μmol m−2 s−1), and with or without glucose addition (0 and 1 g L−1), as well as a central condition in the intermediate level of these 4 parameters (28 °C, pH 7.2, 350 μmol m−2 s−1 and 0.5 g L−1 of glucose). The strain maintenance conditions were used as the control condition in the factorial design experiment. The experiment was performed with batch cultures in triplicate using glass tubes closed with screw lids (20 mL) until the stabilization of the stationary phase. The biomass concentration of factorial design was measured by weighing the total suspended solids [25] and was calculated based on the difference between the final and initial biomass values.
Experimental Procedure
The experiments were carried out in triplicate simultaneously in an incubator with temperature control (411 FPD Ethiktechnology). The strain was cultivated under two experimental steps with arginine supplementation (100 mg L−1). The first step was utilized to obtain the biomass needed for the second step, which was to hydrogen production.
The First Step: Biomass Production
To obtain biomass in the first step, the culture was maintained under aerobic growth conditions in the TAP medium [24], according to the best result of the improvement biomass concentration in the factorial design (continuous illumination of 200 μmol m−2 s−1 with a cool white lamp; pH 6.2 and 32 ± 0.1 °C) in 500 mL Erlenmeyer flasks with 300 mL of culture, until the middle of the exponential phase. In the first step, an Erlenmeyer flask was used to obtain biomass for each photobioreactor of the second step.
The Second Step
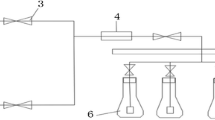
The second step was performed to induce hydrogen production through the anaerobic phototrophic process in batch cultures under either sulfur deprivation in TAP-S medium (0 µmol L−1 of sulfur) or sulfur limitation in TAP50S medium (50 µmol L−1 of sulfur) (Table 1). The biomass of Erlenmeyer flasks with 300 mL of culture from the first step was centrifuged in 2000 rpm for 10 min, washed twice with culture medium selected for each condition and suspended in 500 mL Duran glass bottles with 300 mL of medium and sealed with a butyl cover (photobioreactor). The temperature and pH from the first step were maintained, and there was a variation of the concentration of the sulfur (0 or 50 µmol L−1) and light intensity (60 or 200 μmol m−2 s−1), as shown in Table 1 (12 photobioreactors: 4 conditions in triplicate). The atmospheric conditions of the photobioreactors’ headspace were altered by adding nitrogen gas for 10 min. The nitrogen gas was previously sterilized in a filter with 0.2 μm porosity.
Biomass of the photobioreactors was estimated at the point when the algae were transferred from the 1st to the 2nd experimental step. Samples of 3 mL were taken from each photobioreactor for the analyses, and 0.5 mL of the samples were fixed with Lugol’s acetic solution and stored in the dark until cell count. The biomass, in terms of cell density was measured by cell counting using a Fuchs Rosenthal chamber in an Olympus BX5 microscope [25], and by the chlorophyll a amount, using the extraction method with 80% ethanol [26].
Hydrogen Production
Aliquots of 500 µL of gas of headspace of photobioreactors were collected every 12 h using a syringe and valve (push button valve 22,285) to measure the gas composition by gas chromatography (Shimadzu GC-2010) using a thermal conductivity detector (TCD) and argon as a carrier gas. Experiments were carried out until the hydrogen production was stabilized. The hydrogen results obtained by gas chromatography were calculated based on the regression method with calibration curves (r2 = 0.996). The hydrogen production curves are shown in µmol L−1 of hydrogen/mg L−1 of chlorophyll a.
Furthermore, other parameters were calculated from the data, such as the maximum production of hydrogen per chlorophyll a per liter of culture (in µmol of H2/mg chlorophyll a, represented by Pmax) and per mol (in µmol of H2/liter of culture, represented by H), hydrogen volume (milliliters of H2/liter of culture, represented by VolH2), which were calculated using the ideal gas equation, and hydrogen production rate (µmax) by Gompertz mathematical model [27] (Eq. (1)). Equation (1) represents the accumulated production of hydrogen (H) (µmol H2 L−1), t is the hydrogen production time (hours), and e is Euler’s number (2.71828). The equation results in parameters A (hydrogen production potential in µmol L−1), µmax (hydrogen production rate in µmol H2 L−1 h−1) and k (lag phase time). The specific hydrogen production rate, µspecific (µmol H2 mg Chl a−1 h−1), was calculated from the µmax divided by the initial biomass in chlorophyll a (mg L−1) of the photobioreactor.
By-product Production: Ethanol and Organic Acids
To analyze ethanol and organic acids concentrations, 2 mL of the sample at the end of the second step were collected from photobioreactors. For acetic acid, the same amount of sample was taken at the beginning and at the end of the photobioreactors. All samples were filtered through a 0.2 µm membrane before analysis. Afterwards, these compounds were detected using gas chromatography in a Shimadzu GC-2010/FID with an HP-INNOWAX capillary column [28].
Biochemical Analysis
The biochemical characterization of dry biomass was performed with the samples from the initial photobioreactor (sacrifice reactor) and at the end of the hydrogen production phase. The dry biomass was obtained by centrifuging the cultures followed by heat treatment in an oven at 60 °C for 24 h of the pelletized biomass until a constant weight was achieved. Carbohydrates were determined using the phenol colorimetric method [29] and the total protein was estimated by the total nitrogen analysis TKN [25] with a protein conversion factor of 4.71 [30].
Nutrient Removal
The ammoniacal nitrogen and phosphate concentrations, as well as chemical oxygen demand (COD) were analyzed from 10 mL of samples, filtered through a 0.2 µm membrane, taken from the initial and at the end of photobioreactors of the second step, during the hydrogen production process. Nutrient removal was calculated based on the difference between the initial and final concentrations of nitrogen, phosphate, and COD, shown in percentage. The nutrient analyses were carried out according to APHA [25]. Furthermore, the acetic acid removal (analyzed as described above) was also calculated by the difference between the initial and final concentrations.
Analysis of Results
The Protimiza Experimental Design® software was used to analyze the factorial design of improvement of C. reinhardtii biomass and to find the parameters that had positive effects on the biomass concentration through the Pareto Chart with a significance level of 5%. Furthermore, the Statistica 7.0® software was used for the Analysis of Variance (ANOVA) and Tukey post-hoc tests to differences among the biomass concentration. The best result of biomass production was selected from the highest averages of biomass concentration combined with the Pareto Chart.
The results of the hydrogen production parameters were processed using the Gompertz mathematical model according to Zwietering et al. [27] in the statistical software Origin Pro 8.0®. Analysis of Variance (ANOVA) and Tukey post-hoc tests were used to assess the differences among the hydrogen production parameters, such as the hydrogen production rate, maximum production of hydrogen, as well as biomass concentration by chlorophyll a and cell density. Furthermore, the same statistical comparisons were made of the amount of nutrient removal (ammoniacal nitrogen, phosphate, COD, and acetic acid) and ethanol produced. The T-Student test was used to compare the initial and final carbohydrate and protein amounts of the biomass in the photobioreactors. The probability value (p) of ≤ 0.05 was adopted to indicate the significant differences.
Results
Improvement of C. reinhardtii Biomass
The results of factorial design of improvement of C. reinhardtii biomass (Table 2) showed that a higher temperature and more acidic pH improve the biomass concentration of C. reinhardtii. The ANOVA analysis confirmed that there was a difference in the biomass concentration between the conditions tested (17 experiments of design and control) (F(17,37) = 51.27, p < 0.0001). In general, the best biomass concentration conditions, without any statistical differences between them, occurred at 32 °C and pH 6.2, regardless of the light intensity and glucose addition (conditions 10, 11 and 13), except condition 12. The central point (condition 9) also obtained a higher biomass concentration and there were no differences among the conditions mentioned above.
The analyze of Pareto Chart (Fig. 1) indicated effect positive of temperature (t = 11.62, p < 0.00001), effect negative of pH and light intensity ( t = − 10.04 and − 3.75, p ≤ 0.0006), and a positive interaction of variables temperature and pH (t = 3.28, p = 0.0022). In addition, there was no effect of glucose addition. Although there is no difference in the biomass concentration among conditions 9, 10, 11 and 13 (Table 2), the results from the Pareto Chart showed that higher pH values and light intensity can impair the biomass concentration. Therefore, condition 10 was chosen as the best one to obtain biomass in the first step (32 °C, pH 6.2 and 200 μmol m−2 s−1) instead of another condition with greater light intensity and glucose addition.

Pareto Chart showing the effect of each factor and their interactions in improving the biomass concentration of C. reinhardtii. The variables are indicated for X1 (temperature), X2 (pH), X3 (light intensity) and X4 (glucose addition). The bold vertical bar indicates the significance level of 5%
Hydrogen and Ethanol Production
There was no statistical difference in the initial biomass of the first step transferred to the photobioreactors between all conditions. The cell density was on average 6.9 × 10−6 ± 0.49 cell mL−1 and chlorophyll a concentration 13.7 ± 1.09 mg L−1. The experiments were carried out until the H2 production was stabilized (180 h).
The best result was obtained from the condition without sulfur and under higher light intensity (condition 2: 285.74 ± 27.01 µmol H2 mg Chl a−1 and 3.16 ± 0.05 µmol H2 mg Chl a−1 h−1). The second-best result was obtained under the same conditions but under lower light intensity (condition 1: 209.79 ± 2.55 µmol mg Chl a−1 and 2.01 ± 0.10 µmol H2 mg Chl a−1 h−1). The hydrogen production curves (Fig. 2) and the VolH2, H and µmax (Table 3) also confirmed these results.

Pmax (µmol H2 mg Chl a−1). Conditions 1 to 4 are represented by cond. and were described in Table 1. The bars indicate the standard deviation (n = 3)
Furthermore, the ANOVA analysis confirmed that there was an effect of light and sulfur concentration (Table 3). In all these parameters of hydrogen production, the second condition was statistically better than all other conditions (p ≤ 0.0008) and the first condition was better compared to the third and fourth conditions (p ≤ 0.0002) (Table 3).
The highest ethanol production occurred in the condition with sulfur and light of 200 μmol m−2 s−1 (203.20 ± 31.49 mg L−1) (Fig. 3), although this number was statistically higher when compared only to conditions 1 (139.30 ± 10.31 mg L−1). According to the ANOVA analysis, only the sulfur concentration affected the ethanol production (F (1,8) = 8.08, p = 0.02). Under all conditions, there was simultaneous production of organic acids, such as isobutyric, butyric and isovaleric acids, but these were below the quantification limit of the method.

Concentration (mg L−1) of ethanol under conditions 1 to 4. The bars indicate the standard deviation (n = 3). Symbols show statistical differences, according to ANOVA. * Difference with condition 1 (p = 0.034)
Biochemical Composition of Biomass
The initial, accumulated in the first step, and final carbohydrate and protein biomass composition in the second step were different under most of the conditions (Fig. 4). In general, there was a consumption of carbohydrate in cultures, however only there were statistical differences in conditions under higher light intensity, regardless of the presence of sulfur (average consumption of 38%). The ANOVA analysis confirmed that there was an effect of light in consumption of carbohydrate (F(1,8) = 30.20, p = 0.0006), while in protein consumption this effect occurred only of sulfur concentration (F(1,8) = 133.13, p = 0.000003). However, in all conditions, there was significant protein consumption during the hydrogen production, and this consumption was on average 27% higher in conditions with higher hydrogen production (1 and 2 conditions).

Carbohydrate (A) and protein (B) (mg) per mg of algal biomass under conditions 1 to 4. The biomass compositions accumulated in the first step is represented by initial bars (black), and biomass compositions at the end of the second step (gray bars). The bars indicate the standard deviation (n = 3). Symbols show statistical differences, according to ANOVA. * Difference between the initial and the final biomass of the bioreactors in each condition (p ≤ 0.013)
Nutrient Removal
The percentage of ammoniacal nitrogen removal in the photobioreactors (Fig. 5), occurred only under conditions 3 and 4 (6.11 and 4.57%, respectively), and there was an effect of the sulfur concentration in the photobioreactors (F(1,8) = 73.45, p = 0.00003). However, the decrease in protein levels in biomass may have obscured the results of ammoniacal nitrogen removal. Phosphate removal (Fig. 5), in general, was higher under conditions that yielded lower hydrogen production, and according to ANOVA analysis, there was an effect of sulfur concentration (F(1,8) = 113.42, p = 0.000005), and of light intensity (F(1,8) = 69.83, p = 0.000032).

Percentage of nutrient removal, ammoniacal nitrogen, phosphate, chemical oxygen demand (COD), and acetic acid under conditions 1 to 4. The bars indicate the standard deviation (n = 3). Symbols show statistical differences, according to ANOVA. * Difference with conditions 1 and 2 in the ammoniacal nitrogen uptake (p ≤ 0.004). # Difference with condition 2 in the orthophosphate uptake (p ≤ 0.0002). λ Difference with conditions 1, 2 and 4 in the COD uptake (p ≤ 0.006). γ Difference with conditions 1and 2 in the COD uptake (p ≤ 0.007). ε Difference with condition 2 in the COD uptake (p = 0.038). α Difference with conditions 1 and 2 in the acetic acid uptake (p ≤ 0.0003)
The sulfur concentration and light intensity also affected organic carbon removal (Fig. 5), as measured by COD, according to ANOVA (F(1,8) = 155.55 and 33.71, respectively, p ≤ 0.0004). The higher organic carbon removal also occurred under lower hydrogen production conditions. Acetic acid, with an average initial concentration of 910 mg L−1 in the photobioreactors, was consumed in all conditions (Fig. 5). Under conditions with sulfur addition there was statistically the highest consumption of acetic acid (average consumption of 97.2%) compared to conditions without sulfur addition (mean consumption of 57.2%). The ANOVA analysis confirmed there was an effect of sulfur concentration in the acid acetic removal (F(1,8) = 186.55, p = 0.000001) and there was no light intensity effect.
Discussion
The biomass improvement experiment showed that mainly the pH and the light intensity interfered in obtaining biomass, corroborating with previous research that claims that physical and chemical factors affect the growth of microalgae [31, 32]. Thus, a higher amount of biomass was obtained in the first step with a lower pH (6.2) and light intensity of 200 μmol m−2 s−1. It is believed that the result of improved biomass of C. reinhardtii applied in the first step of the experimental procedure provided the inoculation of a greater amount of biomass in the photobioreactors and favored the hydrogen production in the second step as studies have reported that an increased amount of biomass can improve the hydrogen production yield [6,7,8, 19].
The results described above demonstrate a significant influence of sulfur concentration and light intensity in hydrogen production by C. reinhardtii (CC425). The best results were obtained with sulfur-deprived (0 µmol L−1 of sulfur), and higher light intensity supported better hydrogen production, 3573.07 µmol L−1, and 2623.66 µmol L−1 at the lowest light intensity (conditions 2 and 1, respectively) for 180 h, agree with previous literature that used similar methodology, and were even higher compared to some of the reported results [33, 34].
In order to improve photobiological hydrogen production, there must be a balance between respiration and photosynthesis, and this can be achieved by varying the concentration of nutrients in the culture medium, such as sulfur, nitrogen and phosphorus [17]. Sulfur deprivation decreases photosynthetic activity and consequently decreases oxygen production, which is consumed by respiration. There is a compensatory point between photosynthesis and respiration that results in an anaerobic environment [5, 7, 8, 35]. Reports show that low sulfur concentrations (around 12.5 to 100 µmol L−1) can improve the compensatory point between photosynthesis and respiration and obtain better results in hydrogen production [12, 13].
However, other researchers obtained negative results when low sulfur concentration was added when compared to total sulfur deprivation [35]. The addition above 50 µmol L−1 of sulfur can delay the onset of hydrogen production and decrease the final yield of hydrogen produced [12]. The same occurred in this research, and when comparing the conditions without or with sulfur addition (comparison of conditions 1 with 3 and 2 with 4), it was observed that the hydrogen productivity was on average 6 times higher when there was total sulfur deprivation.
The lowest hydrogen production in sulfur limitation conditions (50 µmol L−1 of sulfur) could be due to an increased photosynthetic rate caused by sulfur presence. Consequently, an increase in oxygen production may have occurred, thus resulting in the inhibition of hydrogen production. Another factor associated with sulfur concentration is light intensity, which must be varied in parallel with the sulfur concentration, so that the best compensatory point between photosynthesis and respiration to make the anaerobic environment can be achieved [6, 8, 35]. Wild and mutant strains may also respond differently to sulfur concentrations and light intensities, and variations in this compensatory point may also occur. It has been stated that approximately 27 µmol m−2 s−1 is the best light intensity to achieve this compensation point with the CC425 strain [32], which is lower than the intensities used in this study and which possibly prevented from obtaining positive results upon sulfur addition.
In this research, it was evident that the light intensity was another factor that interfered in hydrogen production, mainly in sulfur-deprived conditions in which the production was, respectively, 73.4% higher under 200 µmol m−2 s−1 compared to 60 µmol m−2 s−1. Although higher light intensities can inhibit the hydrogenase due to higher photosynthetic rate [36], in sulfur-deprived cultures there is low evolution of oxygen through photosynthesis, then this inhibition by light is not expected to occur [18]. Several researchers analyzed the influence of light intensity in hydrogen production in sulfur deprived cultures, with different strains of Chlamydomonas, and some obtained better results at lower intensities (around 12 to 40 µmol m−2 s−1) [18, 37] and others at higher intensities (around 100 to 300 µmol m−2 s−1) [7, 38], although researchers affirm that the optimal light intensity is 50 to 200 µmol m−2 s−1 for hydrogen production [39].
Different light intensities, light:dark cycles, light absence, and sulfur deprivation are also factors that aid in the investigation of metabolic pathways involved in hydrogen and other fermentation by-products. In this study, acetic acid was used as a source of organic carbon and it was consumed in all conditions, although the consumption was higher in the photobioreactors containing sulfur (average consumption of 57.2% under sulfur-free conditions and 97.2% with sulfur addition). As discussed above, under these conditions hydrogen production was lower, probably due to the increase in the photosynthetic rate, as previously discussed. This consumption is one of the factors responsible for maintaining the anaerobic environment and providing reductant for hydrogen production via indirect photoproduction (photofermentation) [5]. Furthermore, acetic acid can also be used by microalgae as a substrate for respiration and starch accumulation, and after 24 h of anoxia acetate, acetate production by the fermentative acidogenic pathways can be observed, and there is a superposition between its consumption and production [8]. Therefore, the results suggest that in the presence of additional sulfur, the higher acetic acid consumption rate is probably due to its lower production by the fermentation pathway, justifying and corroborating the lower hydrogen production that occurs under these conditions.
As the system becomes anaerobic, the ability of microalgae to modify their metabolism to anaerobic fermentation can lead to the generation of organic acids, ethanol, carbon dioxide and hydrogen through multiple fermentation pathways through anaerobic decomposition of pyruvate [7, 8, 20, 40]. However, considering that an important part of the pyruvate anaerobic metabolism occurs in the chloroplast, where the hydrogenase is located, there is competition for a reductant between the hydrogen production and other fermentation pathways [6, 9].
The production of ethanol measured under all experimental conditions in this work confirms the occurrence of the alternative fermentative metabolic pathways. Ethanol production was higher under higher light intensity and with sulfur (average of 203.2 mg L−1). The results of this research were better, compared to those of other researchers that utilized similar methodology and the same strain, who reported 37 mg L−1 [8] and 57 mg L−1 of ethanol [40], but under lower light intensity (100 µmol m−2 s−1) and a more basic pH (around 7.2 to 7.7). It is known that low pH favor ethanol production, which explains the better ethanol production results in this study. However, it has also been shown that high ethanol production decreases hydrogen photoproduction due to the competition for the reductant that occurs between these two pathways [16]. An alternative to increase ethanol production is to produce this compound from microalgal biomass as they accumulate carbohydrate in the biomass [4]. Biorefineries can use residual microalgal biomass in bioethanol production processes and obtain more satisfactory results, such as 3.83 g L−1 of ethanol from Chlorococum sp. biomass by fermentation [41] and 4.17 g L−1 of ethanol from Chlorella vulgaris biomass by enzymatic hydrolysis and fermentation [4, 42].
Besides hydrogen and ethanol production by the fermentative pathways, the more acidic pH and nutritional sulfur limitation modify the protein metabolic pathway causing an increase in the degradation of proteins and carbohydrates [16]. It has been reported that the decrease in carbohydrates and protein content occurs concomitantly with hydrogen photoproduction in sulfur-deprived cultures [15, 16, 43], although the carbohydrate accumulation can occur during the hydrogen production due to the presence of acetic acid and mixotrophic metabolism of Chlamydomonas [38]. This information corroborates the results obtained in this research, in which there was a decrease in protein concentrations in all photobioreactors, especially under deprived sulfur, which show higher hydrogen production (consumption of protein 111 times higher in deprived sulfur conditions than conditions with sulfur). Carbohydrate consumption, also observed in all conditions, probably occurred for ATP generation needed to conduct metabolic processes that require energy during anaerobiosis [9, 15, 43].
In addition to the hydrogen and ethanol production by the CC425 strain in this research, the results also demonstrated that it is possible to associate the cultivation of microalgae with wastewater treatment. Recent studies have shown the application of microalgae in wastewater treatment, mainly using Chlorophyceae [21, 22, 44]. However, there is a lack of research on Chlamydomonas genus in the literature.
The results demonstrated mainly the removal of acetic acid, as previously discussed, and the occurrence of phosphate uptake, with an average removal of 72.8%, which may have occurred by its adsorption to the cell surface and assimilation into biomass [44]. Phosphate enters the cell by active transport through the cell membrane and can be used in the formation of ATP molecules [22]. Nitrogen uptake was not observed at significant levels, possibly due to the increased accumulation of nitrogen compounds in the liquid medium caused by the decrease in biomass protein under all tested conditions. Furthermore, it was observed that the nitrogen removal occurred under the conditions, in which there was less decrease in biomass protein, corroborating the results. Indeed, organic matter uptake was greater under conditions of lower hydrogen production. Under higher hydrogen production conditions, an increase in acetic acid production, as previously discussed, may have occurred due to the acidogenic pathway and may have influenced this result, as previously discussed.
In general, the greatest removal of nutrients occurred in the photobioreactors under sulfur limitation, in which there was the lowest hydrogen production. These data can be an indication that under these conditions there was a higher photosynthetic rate, and consequently it did not provide a totally anaerobic environment [6, 12]. Thus, it may have contributed to the greater removal of nutrients, since photosynthetic microorganisms can be efficient for this purpose in aerobic conditions, although a high concentration of dissolved oxygen may limit the removal [22, 44, 45]. Therefore, a microaerobic environment may have reduced the hydrogen production, due to the inhibition of hydrogenase, consequently causing metabolic deviations to fermentative pathways and higher production of ethanol in these conditions, in addition to having provided greater nutrient removal.
It has been shown that microalgae biomass bioflocculation, or immobilized-cell cultivation systems, can improve the reduction in nutritive agent in media [21]. In this research, the assays were performed with suspended-cell cultivation systems, which may have decreased the nutrient removal in media. Therefore, research with flocculated biomass of CC425 strain in the wastewater treatment is the aim of future research. In addition, the microalgae may be associated with other microorganisms. Algal–bacterial consortia can be combined for biohydrogen generation and wastewater treatment. In this case, the aerobic bacteria can consume the oxygen produced by the algae in photosynthesis, providing an anaerobic environment and contributing to the hydrogen production [45]. Furthermore, if there is an anaerobic digestion pathway, fermentative bacteria break down organic matter in wastewater into soluble acids, alcohols, hydrogen gases and carbon dioxide. Acetate can be generated from these products by acidogenic bacteria through the process of acetogenesis during the dark fermentation step [23, 46]. In the second step, microalgae having mixotrophic or heterotrophic metabolism, such as Chlamydomonas, are able to grow in closed systems and use this acetate produced as an organic carbon source to produce hydrogen by photoheterotrophic pathway (indirect biophotolysis) [9, 14, 46].
Therefore, the results of this research confirm that hydrogen production occurs through direct water biophotolysis and from carbohydrates breakdown, from the starch reserve or from acetic acid. The occurrence of each pathway may vary according to the different conditions studied. The relative contribution of each of the electron sources may depend on factors such as the strain, extent of damage of PSII, culture conditions and metabolic restrictions [46], such as light intensity and sulfur concentration, all of which affected the results obtained in this research. In addition, the results of this research demonstrate applicability of C. reinhardtii cultivation (CC425) in anaerobic photobioreactors and provide an important discussion of factors that influence hydrogen production, as well as the contribution of using this strain for other purposes of environmental and economic interest.
Conclusion
The improvement of obtaining biomass in the first step may be an important measure to obtain better hydrogen production in the second step. Factors such as sulfur concentration and light intensity influenced the production of hydrogen by C. reinhardtii (CC425), and the best results occurred in the absence of sulfur and under higher light intensity. The results also demonstrate that it is possible to produce ethanol and remove nutrients with this green algae strain in closed anaerobic systems. The most efficient ethanol production occurred in photobioreactors under higher light intensity and in the presence of sulfur, and acid acetic and phosphate removal efficiency were mainly observed, regardless of the condition tested. Therefore, this research demonstrates the efficient hydrogen production by strain C. reinhardtii (CC425) and shows that hydrogen production by this microorganism can be associated with ethanol production and wastewater treatment in closed anaerobic photobioreactors.
Data Availability
The datasets generated during and/or analyzed during the current study are available in the University of São Paulo repository. Persistent web link to datasets: https://teses.usp.br/teses/disponiveis/18/18138/tde-21032017-100636/pt-br.php
References
Razeghifard R (2013) Algal biofuels. Photosynth Res 117(1–3):207–219. https://doi.org/10.1007/s11120-013-9828-z
Demirbas A (2010) Use of algae as biofuel sources. Energy Convers Manag 51(12):2738–2749. https://doi.org/10.1016/j.enconman.2010.06.010
Dubini A, Ghirardi ML (2015) Engineering photosynthetic organisms for the production of biohydrogen. Photosynth Res 123(3):241–253. https://doi.org/10.1007/s11120-014-9991-x
de Farias Silva CE, Bertucco A (2019) Bioethanol from microalgal biomass: a promising approach in biorefinery. Braz Arch Biol Technol 62:1–14. https://doi.org/10.1590/1678-4324-2019160816
Ghirardi ML (2015) Implementation of photobiological H2 production: the O2 sensitivity of hydrogenases. Photosynth Res 125(3):383–393. https://doi.org/10.1007/s11120-015-0158-1
Antal TK, Krendeleva TE, Tyystjärvi E (2015) Multiple regulatory mechanisms in the chloroplast of green algae: relation to hydrogen production. Photosynth Res 125(3):357–381. https://doi.org/10.1007/s11120-015-0157-2
Tamburic B, Zemichael FW, Maitland GC, Hellgardt K (2011) Parameters affecting the growth and hydrogen production of the green alga Chlamydomonas reinhardtii. Int J Hydrogen Energy 36(13):7872–7876. https://doi.org/10.1016/j.ijhydene.2010.11.074
Antal T, Petrova E, Slepnyova V et al (2020) Photosynthetic hydrogen production as acclimation mechanism in nutrient-deprived Chlamydomonas. Algal Res 49:101951. https://doi.org/10.1016/j.algal.2020.101951
Catalanotti C, Yang W, Posewitz MC, Grossman AR (2013) Fermentation metabolism and its evolution in algae. Front Plant Sci 4:150. https://doi.org/10.3389/fpls.2013.00150
Posewitz MC, King PW, Smolinski SL, Smith RD, Ginley AR, Ghirardi ML, Seibert M (2005) Identification of genes required for hydrogenase activity in Chlamydomonas reinhardtii. Biochem Soc Trans 33(Pt1):102–104. https://doi.org/10.1042/BST0330102
Peden EA, Boehm M, Mulder DW, Davis R, Old WM, King PW, Ghirardi ML, Dubini A (2013) Identification of global ferredoxin interaction networks in Chlamydomonas reinhardtii. J Biol Chem 288(49):35192–35209. https://doi.org/10.1074/jbc.M113.483727
Kosourov S, Tsygankov A, Seibert M, Ghirardi ML (2002) Sustained hydrogen photoproduction by Chlamydomonas reinhardtii: effects of culture parameters. Biotechnol Bioeng 78(7):731–740. https://doi.org/10.1002/bit.10254
Zhang L, Happe T, Melis A (2002) Biochemical and morphological characterization of sulfur-deprived and H2-producing Chlamydomonas reinhardtii (green alga). Planta 214(4):552–561. https://doi.org/10.1007/s004250100660
Mus F, Dubini A, Seibert M, Posewitz MC, Grossman AR (2007) Anaerobic acclimation in Chlamydomonas reinhardtii: anoxic gene expression, hydrogenase induction, and metabolic pathways. J Biol Chem 282(35):25475–25486. https://doi.org/10.1074/jbc.M701415200
Gonzalez-Ballester D, Jurado-Oller JL, Fernandez E (2015) Relevance of nutrient media composition for hydrogen production in Chlamydomonas. Photosynth Res 125(3):395–406. https://doi.org/10.1007/s11120-015-0152-7
Kosourov S, Seibert M, Ghirardi ML (2003) Effects of extracellular pH on the metabolic pathways in sulfur-deprived, H2-producing Chlamydomonas reinhardtii cultures. Plant Cell Physiol 44(2):146–155. https://doi.org/10.1093/pcp/pcg020
Pongpadung P, Liu J, Yokthongwattana K, Techapinyawat S, Juntawong N (2015) Screening for hydrogen-producing strains of green microalgae in phosphorus or sulphur deprived medium under nitrogen limitation. Sci Asia 41:97–107. https://doi.org/10.2306/scienceasia1513-1874.2015.41.097
Laurinavichene T, Tolstygina I, Tsygankov A (2004) The effect of light intensity on hydrogen production by sulfur-deprived Chlamydomonas reinhardtii. J Biotechnol 114(1–2):143–151. https://doi.org/10.1016/j.jbiotec.2004.05.012
Vargas SR, Santos PVD, Zaiat M, Calijuri MDC (2018) Optimization of biomass and hydrogen production by Anabaena sp. (UTEX 1448) in nitrogen-deprived cultures. Biomass Bioenergy 111:70–76. https://doi.org/10.1016/j.biombioe.2018.01.022
Hallenbeck PC, Benemann JR (2002) Biological hydrogen production; Fundamentals and limiting processes. Int J Hydrogen Energy 27(11–12):1185–1193. https://doi.org/10.1016/S0360-3199(02)00131-3
Pham TL, Bui MH (2020) Removal of nutrients from fertilizer plant wastewater using Scenedesmus sp.: formation of bioflocculation and enhancement of removal efficiency. J Chem 8094272. https://doi.org/10.1155/2020/8094272
Gonçalves AL, Pires JCM, Simões M (2017) A review on the use of microalgal consortia for wastewater treatment. Algal Res 24(B):403–415. https://doi.org/10.1016/j.algal.2016.11.008
Harun R, Davidson M, Doyle M et al (2011) Technoeconomic analysis of an integrated microalgae photobioreactor, biodiesel and biogas production facility. Biomass Bioenergy. https://doi.org/10.1016/j.biombioe.2010.10.007
Gorman DS, Levine RP (1965) Cytochrome f and plastocyanin: their sequence in the photosynthetic electron transport chain of Chlamydomonas reinhardi. Proc Natl Acad Sci U S A 54(6):1665–1669. https://doi.org/10.1073/pnas.54.6.1665
APHA (2011) Standard methods for the examination of water and wastewater, 21st edn. American Public Health Association, American Water Works Association (AWWA); Water Environment Federation (WEF), Washington
Nusch EA (1980) Comparison of different methods for chlorophyll and pheopigment determination. Arch Hydrobiol Bech 14:14–36
Zwietering MH, Jongenburger I, Rombouts FM, Van’t Riet K (1990) Modeling of the bacterial growth curve. Appl Environ Microbiol 56(6):1875–1881. https://doi.org/10.1128/aem.56.6.1875-1881.1990
Adorno M, Hirasawa J, Varesche M (2014) Development and validation of two methods to quantify volatile acids. Am J Anal Chem 5(7):406–414. https://doi.org/10.4236/ajac.2014.57049
Dubois M, Gilles KA, Hamilton JK, Rebers PA, Smith F (1956) Colorimetric method for determination of sugars and related substances. Anal Chem 28(3):350–356. https://doi.org/10.1021/ac60111a017
Lourenço SO, Barbarino E, Lavín PL, Marquez UML, Aidar E (2004) Distribution of intracellular nitrogen in marine microalgae: calculation of new nitrogen-to-protein conversion factors. Eur J Phycol 39(1):17–32. https://doi.org/10.1080/0967026032000157156
Sun H, Zhao W, Mao X, Li Y, Wu T, Chen F (2018) High-value biomass from microalgae production platforms: strategies and progress based on carbon metabolism and energy conversion. Biotechnol Biofuels 11:227. https://doi.org/10.1186/s13068-018-1225-6
Rühle T, Hemschemeier A, Melis A, Happe T (2008) A novel screening protocol for the isolation of hydrogen producing Chlamydomonas reinhardtii strains. BMC Plant Biol 8:107. https://doi.org/10.1186/1471-2229-8-107
Kosourov SN, Ghirardi ML, Seibert M (2011) A truncated antenna mutant of Chlamydomonas reinhardtii can produce more hydrogen than the parental strain. Int J Hydrogen Energy 36(3):2044–2048. https://doi.org/10.1016/j.ijhydene.2010.10.041
Xu L, Wang Q, Wu S, Li D (2014) Improvement of hydrogen yield of lba-transgenic Chlamydomonas reinhardtii caused by increasing respiration and impairing photosynthesis. Int J Hydrogen Energy 39(25):13347–13352. https://doi.org/10.1016/j.ijhydene.2014.04.178
Laurinavichene TV, Fedorov AS, Ghirardi ML, Seibert M, Tsygankov AA (2006) Demonstration of sustained hydrogen photoproduction by immobilized, sulfur-deprived Chlamydomonas reinhardtii cells. Int J Hydrogen Energy 31(5):659–667. https://doi.org/10.1016/j.ijhydene.2005.05.002
Markov SA, Eivazova ER, Greenwood J (2006) Photostimulation of H2 production in the green alga Chlamydomonas reinhardtii upon photoinhibition of its O2-evolving system. Int J Hydrogen Energy 31(10):1314–1317. https://doi.org/10.1016/j.ijhydene.2005.11.017
Pyo Kim J, Duk Kang C, Hyun Park T, Kim MS, Sim SJ (2006) Enhanced hydrogen production by controlling light intensity in sulfur-deprived Chlamydomonas reinhardtii culture. Int J Hydrogen Energy 31(11):1585–1590. https://doi.org/10.1016/j.ijhydene.2006.06.026
Grossman AR, Catalanotti C, Yang W, Dubini A, Magneschi L, Subramanian V, Posewitz MC, Seibert M (2011) Multiple facets of anoxic metabolism and hydrogen production in the unicellular green alga Chlamydomonas reinhardtii. New Phytol 190(2):279–288. https://doi.org/10.1111/j.1469-8137.2010.03534.x
Dasgupta CN, Jose Gilbert J, Lindblad P, Heidorn T, Borgvang SA, Skjanes K, Das D (2010) Recent trends on the development of photobiological processes and photobioreactors for the improvement of hydrogen production. Int J Hydrogen Energy 35(19):10218–10238. https://doi.org/10.1016/j.ijhydene.2010.06.029
Ghirardi ML, Subramanian V, Wecker MSA, Smolinski S, Antonio RV, Xiong W, Gonzalez-Ballester D, Dubini A (2018) Survey of the anaerobic metabolism of various laboratory wild-type Chlamydomonas reinhardtii strains. Algal Res 35:355–361. https://doi.org/10.1016/j.algal.2018.05.002
Harun R, Danquah MK, Forde GM (2010) Microalgal biomass as a fermentation feedstock for bioethanol production. J Chem Technol Biotechnol 85(2):199–203. https://doi.org/10.1002/jctb.2287
Vargas SR, dos Santos PV, Giraldi LA, Zaiat M, Calijuri MC (2018) Anaerobic phototrophic processes of hydrogen production by different strains of microalgae Chlamydomonas sp. FEMS Microbiol Lett 365(9). https://doi.org/10.1093/femsle/fny073
Wollmann F, Dietze S, Ackermann JU, Bley T, Walther T, Steingroewer J, Krujatz F (2019) Microalgae wastewater treatment: biological and technological approaches. Eng Life Sci 19(12):860–871. https://doi.org/10.1002/elsc.201900071
Tang CC, Tian Y, Liang H, Zuo W, Wang ZW, Zhang J, He ZW (2018) Enhanced nitrogen and phosphorus removal from domestic wastewater via algae-assisted sequencing batch biofilm reactor. Bioresour Technol 250:185–190. https://doi.org/10.1016/j.biortech.2017.11.028
Yao S, Lyu S, An Y, Lu J, Gjermansen C, Schramm A (2019) Microalgae–bacteria symbiosis in microalgal growth and biofuel production: a review. J Appl Microbiol 126(2):359–368. https://doi.org/10.1111/jam.14095
Marín-Navarro J, Esquivel MG, Moreno J (2010) Hydrogen production by Chlamydomonas reinhardtii revisited: Rubisco as a biotechnological target. World J Microbiol Biotechnol 26:1785–1793. https://doi.org/10.1007/s11274-010-0359-x
Acknowledgements
The authors would like to acknowledge the technical support of the Biological Processes Laboratory, Biotoxicological Laboratory of Continental Water and Effluents, the National Renewable Energy Laboratory for the donation of the Chlamydomonas reinhardtii, and Dr. Maria L. Ghirardi for her helpful advice in preparing the manuscript.
Funding
This work was supported by the Coordenação de Aperfeiçoamento de Pessoal de Nível Superior – Brasil (CAPES) (Code 001) and by the Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq).
Author information
Authors and Affiliations
Contributions
All authors contributed to the study conception and design. Conceptualization (Sarah Regina Vargas, Marcelo Zaiat, Maria do Carmo Calijuri); Methodology (Sarah Regina Vargas), Investigation (Sarah Regina Vargas), Formal analysis (Sarah Regina Vargas), Writing- Original draft preparation (Sarah Regina Vargas); Resources (Marcelo Zaiat, Maria do Carmo Calijuri), Supervision (Marcelo Zaiat, Maria do Carmo Calijuri), Project administration (Maria do Carmo Calijuri); Writing- Reviewing and Editing (Marcelo Zaiat, Maria do Carmo Calijuri); Funding acquisition (Maria do Carmo Calijuri). All authors read and approved the final manuscript.
Corresponding author
Ethics declarations
Ethics Approval and Consent to Participate
Not applicable.
Consent for Publication
All the authors mutually consent for publication this manuscript and state that has not been previously published in another journal.
Competing Interests
The authors have no conflicts of interest to declare that are relevant to the content of this article.
Additional information
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
About this article

Cite this article
Vargas, S.R., Zaiat, M. & Calijuri, M.d. Influence of Sulfur and Light Intensity in Nutrient Removal, and Hydrogen and Ethanol Production by Improved Biomass of Chlamydomonas reinhardtii in Batch Anaerobic Photobioreactors. Bioenerg. Res. 15, 218–229 (2022). https://doi.org/10.1007/s12155-021-10296-y
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s12155-021-10296-y