
Abstract
The agronomic performances of giant miscanthus (Miscanthus x giganteus) and switchgrass (Panicum virgatum L.) grown as bioenergy grasses are still unclear in North Carolina, due to a relatively short period of introduction. The objectives of the study were to compare the biomass yield and annual N removal of perennial bioenergy grasses and the commonly grown coastal bermudagrass [Cynodon dactylon (L.) Pers.], and to determine the optimum N rates and harvest practices for switchgrass and miscanthus. A 4-year field trial of the grasses under five annual harvest frequencies (May/Oct, June/Oct, July/Oct, Aug/Oct, and October only) and five annual N rates (0, 67,134, 202, and 268 kg N ha−1) was established at a research farm in Eastern North Carolina in 2011. Across harvest treatments and N rates, greatest biomass was achieved in the second growth year for both miscanthus (19.0 Mg ha−1) and switchgrass (15.9 Mg ha−1). Grasses demonstrated no N response until the second or the third year after crop establishment. Miscanthus reached a yield plateau with a N rate of 134 kg ha−1 since achieving plant maturity in 2013, whereas switchgrass demonstrated an increasing fertilizer N response from 134 kg N ha−1 in the third growth year (2014) to 268 kg N ha−1 in the fourth growth year (2015). The two-cut harvest system is not recommended for bioenergy biomass production in this region because it does not improve biomass yield and increased N removal leads to additional costs.
Similar content being viewed by others

Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
Production of cellulosic biofuels from plant biomass could reduce dependence on fossil fuels. Giant Miscanthus (Miscanthus × giganteus) and switchgrass (Panicum virgatum L.) are emerging as dedicated energy crops due to high biomass yield and low fertilizer requirements [3, 11, 18,19,20]. Water availability is considered a key factor limiting yield of these grasses [18]. The North Carolina Coastal Plain region lies within a humid, temperate area, which has the potential to produce high biomass yield for bioenergy grasses with proper harvesting and nutrient management. Currently, hay production from coastal bermudagrass [Cynodon dactylon (L.) Pers.] is an important part of the local agriculture. Since grasses used for hay and biofuel production share similarities in methods of planting and harvesting, farmers could diversify or alternate their production systems with bioenergy grasses if they bring higher economic returns.
Both miscanthus and switchgrass have high nutrient use efficiencies due to recycling by rhizomes within the plant after senescence [4, 45]. Yield response to N by the two grasses have been well documented and results varied with soil and climate conditions [10, 36, 37, 48]. Cadoux and co-workers [6] reviewed a total of 11 miscanthus studies in Europe that involved N yield response. Five of them had no yield response to N due to high initial soil mineral N content. For example, no N response was found in a 14-year miscanthus yield experiment in England [9]. Schwarz et al. [43] reported a lack of response to N by miscanthus over 3 years in Austria. The six studies with N responses in England, Germany, Italy, and Turkey had optimum N rates ranging from 40 to 120 kg ha−1 over 3–5 years [1, 5, 10, 12, 29]. In general, the biomass increase from these N inputs was moderate. Nitrogen studies with N15 recovered 14, 20, and 26% of the applied N in the aboveground miscanthus biomass during the first three growing seasons in England, which was relatively low compared with other crops [7, 8]. Miguez et al. [34] used statistical models on miscanthus research data from Europe to predict a negligible impact of applied N in the first growing year followed by a relatively small long-term impact in subsequent years.
Miscanthus yield response to N application showed varying results in the USA. Arundale et al. [3] reported the biomass yield of miscanthus after 5 years of establishment increased by 25%, with a linear response up to 202 kg N ha−1 year−1 across seven locations in Illinois. In a 2-year trial in Oklahoma, no N response was found up to 168 kg N ha−1 year−1 for miscanthus that was one year old at the start of the experiment [24]. In experiments starting with 1-year-old miscanthus at two North Carolina sites, Haines and co-workers [16] reported no N response up to 135 kg ha−1 year−1 in 2 of 3 years at one site and in either of 2 years at the second site. Despite multiple miscanthus trials in the Southeastern U.S., there is limited information regarding yield response to N or N use efficiency [13, 26, 37].
Studies on switchgrass N response in the USA have been extensively reported and reviewed. After compiling data from 39 switchgrass field trials, Wullschleger et al. [48] indicated switchgrass responded in general to 100 kg N ha−1. The largest N response of switchgrass was reported in Oklahoma with 448 kg N ha−1 year−1over 3 years [47]. Stout and Jung [46] reported N recovery rates of switchgrass ranging from 22 to 57% with 84 kg N ha−1 year−1 applied in Pennsylvania at four locations over 3 years. In the Northern Appalachian region switchgrass recovered 25–33% of N at applied rates up to180 kg ha−1 year−1 over 3 years [44]. Liu et al. [30] reported switchgrass recovered on average 31% N from biosolids amendments which contained plant-available N (PAN) rates of 180 kg ha−1 on three sites in Virginia. However, some studies reported lower N recovery rates for switchgrass. For example, a multi-site study including South Dakota, New York, Iowa, Oklahoma, and Virginia reported an N recovery rate of switchgrass one year after planting below 10% with up to 112 kg ha−1 N year−1applied at all locations over 2 years [36]. Nitrogen response among 19 location and year combinations varied from none to 56 or 112 kg N ha−1year−1 and was attributed to variances in soil N content [22].
Timing and frequency of harvests are important factors for bioenergy grass production. Thomason and co-workers [47] reported that harvest frequency played a more important role in switchgrass yield for a 3-year study than applied N rates. Miscanthus and switchgrass are typically harvested in the winter after senescence, allowing aboveground nutrient translocation to rhizomes and reducing nutrient removal [28, 45]. A late harvest, however, might decrease biomass yield due to leaf loss [21, 49]. Some studies have indicated increased biomass yield with two-cut or early harvesting systems [4, 14, 40]. For example, a study in Denmark showed miscanthus harvested in early fall produced approximately 40% more biomass than a late harvest in winter [27]. Switchgrass harvested in Oklahoma by a two-cut system produced 30% more biomass than a one-cut system over 4 years [25]. Although yields might increase, greater removal of nutrients during early or two-cut systems could lead to soil nutrient depletion and require more fertilizer for subsequent growing seasons [15, 25, 30, 35, 40]. Another possible side effect of early harvests is declined biomass yield over time. Perrish [39] reported a 30% miscanthus yield decrease after 3 years of early harvesting in September compared with winter harvesting in Illinois, indicating early harvest might decrease the stand longevity. A study in Italy reported that two-cut systems increased biomass production of switchgrass during the first 2 years but yields declined significantly during the last 2 years of the study due to loss of plant stand [35].
Studies comparing the combined effects of both N fertilization and harvest timing and frequency on miscanthus and switchgrass have not been conducted in the Southeastern U.S. The objectives of the study were to: (i) compare the biomass yield and annual nutrient removal of perennial bioenergy grasses and the commonly grown coastal bermudagrass and (ii) determine the optimum N rates and harvest practices for switchgrass and miscanthus in order to provide guidelines for perennial bioenergy grass productions across the Coastal Plain area of North Carolina and other regions with similar climate and soil properties.
Materials and Methods
Site Description
The experiment was conducted at the NCSU Williamsdale Biofuels Field Laboratory (34°46′ N, 78°6′ W) in Wallace, North Carolina. The site was previously used for fertilized corn production. The soil was a Noboco loamy fine sand (fine-loamy, siliceous, subactive, thermic Oxyaquic Paleudults). On-site weather data during the research period were acquired from the North Carolina State climate office (Raleigh, NC, USA). Throughout the experimental period, average growing season (April to September) temperature was relatively consistent with the 30-year means (Table 1). Annual rainfall for 2012, 2013, and 2015 exceeded the 30-year average, but was 428 mm below the 30-year average in 2014.
Experimental Design, Sampling, and Analysis
Treatments were arranged in a split-split plot randomized complete block design with three replications. Main plots were three grass species: switchgrass, giant miscanthus, and coastal bermudagrass. Sub plots were five treatments with either single or dual annual harvests (May/Oct, June/Oct, July/Oct, Aug/Oct, and Oct only). Four of the treatments (May/Oct, June/Oct, July/Oct, and Aug/Oct) were harvested twice per year with the first harvest in May, June, July, and Aug, respectively. All of the second harvests for the two-cut treatments and the October-only treatment were cut on the same day, usually after the first killing frost. Each of the harvest treatment sub plots were divided into five sub-sub plots with different annual N rates (0, 67,134, 202, and 268 kg N ha−1) chosen to encompass the entire range of dry matter yield responses for the three grasses. The size of each sub-sub plot was 5 × 5 m. Plugs of miscanthus cv. “Nagara” and switchgrass cv. “Performer” were planted with a transplanter on May 23, 2011. Miscanthus was planted with an in-row spacing of 92 cm and between-row spacing of 100 cm. Switchgrass had equal in-row and between-row spacings of 50 cm. Coastal bermudagrasss cv. “Ranchero Frio” seed was drilled into plots on July 5, 2011 at the rate of 37 kg ha−1. Urea coated with the urease inhibitor NBPT (Agrotain Ultra, Koch Agronomic Services, LLC, Wichita, KS, USA) was broadcast by hand on plots in late March before the shoot emergence each year since 2012. Pre-emergence herbicide S-metolachlor plus atrazine (Bicep II Magnum, Syngenta, Greensboro, NC, USA) and post-emergence herbicide Paraquat (Gramoxone, Syngenta, Greensboro, NC, USA) were used to control weeds wherever necessary each year. A forage plot harvester (Wintersteiger Inc., Salt Lake City, UT, USA) was used to harvest the grasses and record the wet biomass yield. Due to the young plant stand in 2012, miscanthus was not harvested in May or June for the May/Oct or June/Oct harvest treatments. In 2015, the harvest scheme for all the coastal bermudagrass plots was changed to five harvests per year from the same plot (May, June, July, Aug, and Oct) to represent a common harvest practice for bermudagrass hay production in the region.
Plant subsamples from each sub-sub plot were collected at each harvest and dried at 65 °C for three days to determine moisture content and dry biomass yield. Afterward, subsamples were ground to pass a 1-mm sieve (Christy-Turner, Ipswich, Suffolk, UK) small scale hammer mill and analyzed by the NC Department of Agriculture and Consumer Services (NCDA&CS) plant tissue testing laboratory to determine N concentration [31]. In May 2011, before establishment of the research plots, soil samples were collected at each of the three designated blocks to a 1.2 m depth in four segments (0–30, 30–60, 60–90, and 90–120 cm) with a 5-cm diameter hydraulic probe. Three cores were collected from each block and mixed together by depth interval. Soil samples composed of 10 soil cores collected with a 1.7-cm diameter hand probe were taken at the 0–20 cm depth from the zero N sub-sub plots before fertilization in March from 2012 to 2014 and at the end of the study in December 2015.
Soil samples were tested by routine procedures at the North Carolina Department of Agriculture and Consumer Services (NCDA&CS) soil testing laboratory [17]. Nutrient analysis on Mehlich-3 soil extracts were performed using inductively coupled plasma (ICP) [33]; cation exchange capacity (CEC) was determined by summation of basic cations (excluding Na) and buffer acidity [32]; soil pH was determined on a 1:1 soil/water volume ratio [33].
Crop species, harvest treatments, N rates, and years were considered as fixed effects and replications were considered as random effects. Year was treated as a repeated measure. Biomass yield and N removal data were analyzed using the MIXED procedure in SAS version 9.3 [42] to determine main effects and interactions among the fixed factors. The statistical model applied autoregressive structure to account for temporal data collection across years. The SLICE option in the MIXED procedure was used to perform a partitioned analysis for the N rates × crop × year interaction, where year and crop effects were fixed to identify the simple effects of N rates on crop species in certain years. In cases where simple effects of N rate were significant (P < 0.05) in a year-crop species combination, biomass yield response to N rates was fit to a linear plateau model using the GLM procedure in SAS as described by Anderson and Nelson [2]. Mean separation of yield and N removal were performed using the DIFF option of the LSMEANS statement in the MIXED procedure at the respective interaction levels. The means were compared by Fisher’s protected least significant difference (LSD) at the 5% probability level.
Result and Discussion
Soil Chemical Properties
Soil analysis before crop establishment in 2011 demonstrated most nutrients were concentrated in the 0–30 cm depth, except for soil cations (K, Ca, and Mg) which were more evenly distributed to a depth of 60 cm (Table 2). Throughout the research period, soil P and K levels for samples from the 0–20 cm depth were defined as “high” or “very high” for North Carolina soil tests [17], which means these were not limiting nutrients. The only exception was that soil K concentrations in the switchgrass treatment declined to a “medium” category by the end of 2015. There was no significant decrease in soil P level until the end of the study in 2015, when the P level dropped approximately 50 mg kg−1 compared with the values in March 2012.
Biomass Yield
The four-way interaction of year × crop species × harvest treatment × N rate was not significant. Also, none of the two-way or three-way interactions on biomass yield which included both N rate and harvest treatment were significant (Table 3). Therefore, data were pooled over either harvest treatment or N rate.
The impact of the harvest treatments were evidenced by a significant (P < 0.0001) year × crop species × harvest treatment interaction (Table 3). Since the year × crop species × N rate interaction was not significant (P = 0.7728), the impact of N rate was investigated by the crop-year species combinations with a positive N response identified in Table 4.
Biomass yield differences among crops suggested by the significant crop species × year interaction (Table 3) were evaluated by comparisons between years of species yields averaged across harvest treatments and N rates (Table 5). Miscanthus had the greatest yield among crop species, with values of 19.0 Mg ha−1 in 2013, 17.1 Mg ha−1 in 2014, and 16.7 Mg ha−1 in 2015. Maximum miscanthus yield in 2013 could be an indicator of crop stand maturity that is usually reported at 2 or 3 years after establishment by other studies [10, 19]. The miscanthus yields achieved in this study were higher than or similar to those reported in the Southeastern U.S. [13, 16, 37]. Switchgrass had similar yield to miscanthus in 2012 (13.9 Mg ha−1) and 2013 (15.9 Mg ha−1), but yielded significantly less than miscanthus in 2014 (12.3 Mg ha−1) and 2015 (12.5 Mg ha−1). These combined yields, however, were close to a national average of 14 Mg ha−1 reported by Wullschleger et al. [48]. Comparing at the regional scale, the yields were lower than those reported by Palmer et al. (20.1 Mg ha−1) [37] within the same region of this study but higher than the yield of 9.4 Mg ha−1 in the Georgia Coastal Plain area [26]. Although not produced for bioenergy, the traditional bermudagrass hay yield was the lowest among grasses in every year, with an average annual yield of 5.4 Mg ha−1 (Table 5). A multiple harvest system for bermudagrass adopted in 2015 did not improve yield compared with the one- and two-cut systems used in 2012–2014.
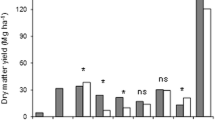
The significant interaction of year × crop species × harvest treatment was evaluated by comparing annual crop species and harvest treatment yields averaged across N rates (Fig. 1). In general, two-cut systems did not produce more biomass yield for all the three grasses than the one-cut treatment throughout the research period. The two-cut system only provided the greatest biomass yield in 2012 for switchgrass and bermudagrass. The two-cut system did not produce significantly greater yield than the one-cut treatment for miscanthus in any year. In 2012, the highest yield of switchgrass was achieved in treatments with dual cuts in July/October and August/October. In 2013, switchgrass under the July/October and August/October harvest treatments still produced the greatest biomass among the two-cut systems but there was no significant difference in yield when compared with the October-only harvest treatment. In 2014 and 2015, the October-only single harvest treatment produced greater switchgrass biomass yield than any of the two-cut treatments. Declines in switchgrass yield over time from the two-cut harvest systems had also been reported by other studies: Thomason et al. [47] and Monti et al. [35] observed switchgrass productivity declined after 2 to 4 years under two- or three-cut harvest management. The yield decline was usually accompanied by a reduction in tiller density, and was attributed to a physiological stress with extra nutrient uptake from the regrowth after the first cut that disrupted the nutrient remobilization to belowground biomass after grass senescence each fall [35, 41]. Furthermore, reduced rainfall in July and August of 2014 and in July 2015 may have limited the production of switchgrass biomass after the first cut (Table 1).

Biomass yields by year, crop species and harvest treatment averaged across N rates. In 2012, the May/October and June/October treatments for miscanthus were treated as missing values because no harvests were made in either May or June to the young crop stand. In 2014, the June/October harvest treatment was missing for bermudagrass. In 2015, the bermudagrass plots were uniformly harvested for five times so there was no harvest treatment effect. Mean separation of biomass yield was performed within years by crop species. Different letters within a given year and crop species indicate that yields of harvest treatments are statistically different (P < 0.05). Error bars are shown as standard errors
The miscanthus stand was too young to harvest until July 2012 (Fig. 1). From 2013 to 2015, yield of miscanthus under the two-cut harvest systems did not decline relative to the October-only harvest treatment. This indicated miscanthus might be more adapted than switchgrass to the two-cut systems, possibly due to greater nutrient reserves in the belowground biomass or higher nutrient translocation efficiency than in switchgrass. Bermudagrass yields increased under the two-cut treatments, but total annual biomass was less than for miscanthus and switchgrass, especially in 2013 and 2014. When averaged across the 4 years, 88% of the miscanthus and 86% of the switchgrass yields in August/October harvest treatments came from the first harvest. The reduced yields from the second harvest in the August/October treatment were associated with a short growing period after the first harvest. From the economic perspective, the efficiency of a second harvest after an initial August harvest might be low because the harvesting and bailing comprises a significant part of production cost [35, 38].
Yield Response to Fertilizer N
Yield response curves to applied N rates (Fig. 2) were fit to the year/crop species combinations with significant N effects (Table 4) at the P = 0.05 level. Nitrogen applications had significant effects on yield in six of the 12 year-crop species combinations, and most occurred during the later years of the experiment. Consistently, since 2013, miscanthus reached a yield plateau with 134 kg N ha−1 (Fig. 2). The slope of the miscanthus yield response (i.e., biomass produced per unit N fertilizer) increased over time due to a gradual decrease in biomass yield from the unfertilized treatments. Switchgrass did not respond to N applications until 2014. The slopes of switchgrass yield responses were almost parallel in 2014 and 2015. The greater maximum yield in 2015 for switchgrass was obtained with a greater N requirement relative to 2014. The only N response for bermudagrass occurred in 2015 when the harvest scheme was changed to five consecutive cuts per year. All the three grasses showed increasing yield responses to N over time. In contrast to the unfertilized treatment, miscanthus yield increased from 2013 to 2015 by 16, 35, and 41% to a yield plateau of 19.0 Mg ha−1; switchgrass yield increased by 36% to 14.1 Mg ha−1 with 134 kg N ha−1 in 2014 and by 49% in a linear response up to 268 kg N ha−1 in 2015; bermudagrass biomass increased by 49% in 2015 to a yield plateau of 6.7 Mg ha−1 when fertilized with 201 kg N ha−1.

Linear or linear plateau relationships between biomass yield of grass species and applied N. Only year/crop species combinations with significant responses to N are presented
The lack of an N response by miscanthus and switchgrass after 1 year of establishment (2012) is in accordance with multiple studies over a wide range of geographical locations [16, 22, 25, 34]. The transition from no response to applied N to increasing dependence on the external N supply coincides with the depletion of native soil N by crop removals over time. This process might be faster in the Southeastern Coastal Plain region where soils have a coarse texture and are low in organic matter content. With relatively large amounts of precipitation in the fall and winter, carry-over effects of applied N are unlikely due to nitrate leaching. A combination of limited organic matter and nitrate leaching might lead to an increased response to fertilizer N over time.
Crop N Recovery
Apparent N recovery is defined as the N removal in harvests for a given fertilizer N treatment minus N removal of the treatment without applied N [23]. Apparent N recovery (ANR) of the year-crop species combinations with significant responses to applied N are presented in Fig. 3. The slope of the relation between apparent N recovery and applied N indicates the proportion of fertilizer N removed by the crop and was estimated by linear regression. Miscanthus recovered a higher percentage of applied N (53%) in 2015 than in 2013 (45%) or 2014 (42%), probably due to a shrinking native soil N pool and an expanding root system to intercept more nutrients. Switchgrass recovered similar portions of applied N (41%) to miscanthus in 2014 up to the yield plateau at 134 kg N ha−1. In 2015, however, the percentage of recovered N for switchgrass (34%) was identical to bermudagrass and was considerably lower than miscanthus. The lower percentage of applied N recovery for switchgrass and bermudagrass in 2015 might explain their needs for higher N rates to achieve maximum yields (Fig. 2). The switchgrass N recovery values were notably higher than those reported by Owens et al. [36] at multiple locations across the USA, probably due to differences in harvest managements, climate, and soil types. The combination of the sandy textured soil and humid climate in the Coastal Plain area results in a relatively lower N content in soil than Midwest U.S.

Apparent N recovery (ANR) (N removal at each N rate – N removal at 0 N rate) of harvested biomass as a function of applied N. The slopes represent the proportions of applied N recovered in aboveground biomass. Fertilizer N treatments beyond the yield plateau with excess N accumulation were excluded
Crop N Removal
Since there were significant interactions for both year × crop species × harvest treatment and year × crop species × N rates interaction (Table 3), crop N removal data were organized by year, crop species and N rates averaged across harvest treatments (Table 6) and by year, crop species and harvest treatment averaged across N rates (Table 7). When analyzed by year × crop species × N rates, the effect of applied N was more pronounced on N removal than on biomass yield (Table 3). This was likely due to plant “luxury N uptake.” A pattern of N removal response to N rates can be observed in most year-crop species combinations (Table 6). Although biomass yields for all the three grasses lacked a response to applied N in 2012 (Table 4), there were notable increased N removals up to 201 kg ha−1 applied N for bermudagrass and 134 kg N ha−1 for switchgrass (Table 6). Lack of increased miscanthus N removal with increasing N rates in 2012 might be due to less developed roots of the young plant stand. In 2013 and 2014, bermudagrass removed significantly less N than miscanthus and switchgrass as a result of substantially lower biomass yields (Fig. 1 and Table 5). With adoption of the five-cut system in 2015, bermudagrass N removal increased 2.7-fold between 0 and 268 kg applied N ha−1 (Table 6). The difference in N removal between treatments of 0 and 268 kg N ha−1 for miscanthus and switchgrass increased with time from 2013 to 2015 (73, 84, and 131 kg N ha−1 for miscanthus and 44, 70, and 91 kg ha−1 for switchgrass). Diminishing native soil N supply over time promoted the need for external N supply.
When analyzed by year × crop species × harvest treatments averaged across N rates, at least one of the two-cut treatments removed significantly more N than the Oct only harvest in 9 out 12 year-crop combinations (Table 7). This result was consistent with other studies that included different harvest times or frequencies as a factor [25, 30, 35, 38]. From 2013 to 2015, the two-cut harvest treatments with the most N removal for miscanthus removed 36% (2013), 20% (2014), and 50% (2015) more N than the October-only harvest treatment. The same pattern could also be observed among switchgrass harvest treatments except in 2015 due to a significantly greater yield under the October-only harvest treatment. In general, the greater N removal under two-cut systems was attributed to a greater N concentration in harvested biomass during the summer. Since there was no benefit of increased biomass yield when harvested twice per year, the extra N removal under a two-cut systems made it undesirable compared with a single harvest system after senescence; the latter would enable aboveground nutrient translocation to belowground biomass and reduce the N removal at harvest. Since there was a yield decline over time with switchgrass plots under the two-cut systems, a single harvest after senescence would avoid the physiological stress associated with the two-cut harvest systems and improve long-term sustainability.
Conclusions
In the Coastal Plain region of North Carolina, both miscanthus and switchgrass produced significantly greater biomass yields than the conventionally grown coastal bermudagrass during a four-year field trial. The two-cut harvest system, with no advantage in biomass yield and with greater N removal, provided no advantage for bioenergy grass production. There was no response to N fertilizer by miscanthus or switchgrass until the second year after crop establishment. Miscanthus demonstrated a stable yield plateau at N applications of 134 kg ha−1 year−1 since achieving plant maturity in 2013. A fixed rate of recommended N was not feasible for switchgrass, because N responses increased from 0 to 268 kg ha−1 across the 2012–2015 period. An incremental N rate recommendation might be more appropriate for switchgrass production as residual soil N levels decline.
References
Acaroglu M, Aksoy AS (2005) The cultivation and energy balance of Miscanthus x giganteus production in Turkey. Biomass Bioenergy 29:42–48
Anderson RL, Nelson LA (1987) Linear-plateau and plateau-linear-plateau models: usefulness in evaluating nutrient responses. Tech bull 283. North Carolina State Univ, Raleigh
Arundale RA, Dohleman FG, Thomas BV, Long SP (2013) Nitrogen fertilization does significantly increase yields of stands of Miscanthus × giganteus and Panicum virgatum in multiyear trials in Illinois. Bioenergy Res 7:408–416
Beale CV, Long SP (1997) Seasonal dynamics of nutrient accumulation and partitioning in the perennial C-4-grasses Miscanthus ×giganteus and Spartina cynosuroides. Biomass Bioenergy 12:419–428
Boehmel C, Lewandowski I, Claupein W (2008) Comparing annual and perennial energy cropping systems with different management intensities. Agric Syst 96:224–236
Cadoux S, Riche AB, Yates NE, Machet JM (2012) Nutrient requirements of Miscanthus x giganteus: conclusions from a review of published studies. Biomass Bioenergy 38:14–22
Christian DG, Poulton PR, Riche AB, Yates NE (1997) The recovery of 15N-labelled fertilizer applied to Miscanthus x giganteus. Biomass Bioenergy 12:21–24
Christian DG, Poulton PR, Riche AB, Yates NE, Todd AD (2006) The recovery over several seasons of 15N-labelled fertilizer applied to Miscanthus x giganteus ranging from 1 to 3 years old. Biomass Bioenergy 30:125–133
Christian DG, Riche AB, Yates NE (2008) Growth, yield and mineral content of Miscanthus x giganteus grown as a biofuel for 14 successive harvests. Ind Crop Prod 28:320–327
Clifton-Brown JC, Breuer J, Jones MB (2007) Carbon mitigation by the energy crop, miscanthus. Glob Change Biol 13:2296–2307
Clifton-Brown JC, Lewandowski I, Andersson B (2001) Performance of 15 miscanthus genotypes at five sites in Europe. Agron J 93:1013–1019
Cosentino SL, Patane C, Sanzone E, Copani V, Foti S (2007) Effects of soil water content and nitrogen supply on the productivity of Miscanthus x giganteus Greef et Deu in a Mediterranean environment. Ind Crop Prod 25:75–88
Fedenko JR, Erickson JE, Woodard KR et al (2013) Biomass production and composition of perennial grasses grown for bioenergy in a subtropical climate across Florida, USA. Bioenergy Res 6:1082–1093
Fike JH, Parrish DJ, Wolf DD et al (2006) Long-term yield potential of switchgrass-for-biofuel systems. Biomass Bioenergy 30:198–206
Guretzky JA, Biermacher JT, Cook BJ, Kering MK, Mosali J (2011) Switchgrass for forage and bioenergy: harvest and nitrogen rate effects on biomass yields and nutrient composition. Plant Soil 339:69–81
Haines SA, Gehl RJ, Havlin JL, Ranney TG (2014) Nitrogen and phosphorus fertilizer effects on establishment of giant miscanthus. BioEnergy Res 8:17–27
Hardy DH, Tucker MR, Stokes CE (2014) Crop fertilization based on North Carolina soil tests. North Carolina Department of Agriculture and Consumer Services, Agronomic Division. Circular no. 1, Raleigh. http://www.ncagr.gov/agronomi/pdffiles/obook.pdf . Accessed 5 July 2017
Heaton EA, Voigt TB, Long SP (2004) A quantitative review comparing the yields of two candidate C4 perennial biomass crops in relation to nitrogen, temperature and water. Biomass Bioenergy 27:21–30
Heaton E, Dohleman FG, Long SP (2008) Meeting US biofuel goals with less land: the potential of miscanthus. Glob Change Biol 14:2000–2014
Heaton EA, Dohleman FG, Long SP (2009) Seasonal nitrogen dynamics of Miscanthus × giganteus and Panicum virgatum. GCB Bioenergy 1:297–307
Himken M, Lammel J, Neukirchen D et al (1997) Cultivation of miscanthus under west European conditions: seasonal changes in dry matter production, nutrient uptake and remobilization. Plant Soil 189:117–126
Hong CO, Owens VN, Bransby D et al (2014) Switchgrass response to nitrogen fertilizer across diverse environments in the USA: a regional feedstock partnership report. Bioenergy Res 7:777–788
Israel DW, Smyth TJ (2015) Crop utilization of nitrogen in swine lagoon sludge. Commun Soil Sci Plant Anal 46:1525–1539
Kering MK, Butler TJ, Biermacher JT, Guretzky JA (2012) Biomass yield and nutrient removal rates of perennial grasses under nitrogen fertilization. Bioenergy Res 5:61–70
Kering MK, Butler TJ, Biermacher JT et al (2013) Effect of potassium and nitrogen fertilizer on switchgrass productivity and nutrient removal rates under two harvest systems on a low potassium soil. Bioenergy Res 6:329–335
Knoll JE, Anderson WF, Strickland TC et al (2012) Low-input production of biomass from perennial grasses in the Coastal Plain of Georgia, USA. Bioenergy Res 5:206–214
Larsen SU, Jorgensen U, Kjeldsen JB, Lærke PE (2014) Long-term miscanthus yield influenced by location, genotype, row distance, fertilization and harvest season. Bioenergy Res 7:620–635
Lewandowsk I, Heinz A (2003) Delayed harvest of Miscanthus—influences on biomass quantity and quality and environmental impacts of energy production. Eur J Agron 19:45–63
Lewandowski I, Schmidt U (2006) Nitrogen, energy and land use efficiencies of miscanthus, reed canary grass and triticale as determined by the boundary line approach. Agric Ecosyst Environ 112:335–346
Liu XJA, Fike JH, Galbraith JM, Fike WB (2015) Biosolids amendment and harvest frequency affect nitrogen use dynamics of switchgrass grown for biofuel production. Bioenergy Res 8:560–569
McGinnis M, Stokes C, Cleveland B (2014) NCDA&CS plant tissue analysis guide. Plant/Waste/Solution/Media Analysis Section. North Carolina Department of Agriculture and Consumer Services, Agronomic Division, Raleigh
Mehlich A (1976) New buffer pH method for rapid estimation of exchangeable acidity and lime requirement of soils. Commun Soil Sci Plant Anal 7:637–652
Mehlich A (1984) Mehlich 3 soil test extractant: a modification of Mehlich 2 extractant. Commun Soil Sci Plant Anal 15:1409–1416
Miguez FE, Villamil MB, Long SP, Bollero GA (2008) Meta-analysis of the effects of management factors on Miscanthus x giganteus growth and biomass production. Agric For Meteorol 148:1280–1292
Monti A, Bezzi G, Pritoni G, Venturi G (2008) Long-term productivity of lowland and upland switchgrass cytotypes as affected by cutting frequency. Bioresour Technol 99:7425–7432
Owens VN, Viands DR, Mayton HS et al (2013) Nitrogen use in switchgrass grown for bioenergy across the USA. Biomass Bioenergy 58:286–293
Palmer IE, Gehl RJ, Ranney TG, Touchell D, George N (2014) Biomass yield, nitrogen response, and nutrient uptake of perennial bioenergy grasses in North Carolina. Biomass Bioenergy 63:218–228
Pedroso GM, Hutmacher RB, Putnama D et al (2013) Yield and nitrogen management of irrigated switchgrass systems in diverse ecoregions. Agron J 105:311–320
Perrish AS (2013) Yield response to nitrogen fertilization and harvest timing on a mature Miscanthus x giganteus stand. Dissertation, University of Illinois at Urbana-Champaign, Urbana, Illinois
Reynolds J, Walker C, Kirchner M (2000) Nitrogen removal in switchgrass biomass under two harvest systems. Biomass Bioenergy 19:281–286
Sanderson MA, Read JC, Reed RL (1999) Harvest management of switchgrass for biomass feedstock and forage production. Agron J 91:5–10
SAS Institute (2011) SAS/STAT User's Guide. Version 9.3. SAS Inst, Cary
Schwarz H, Liebhard P, Ehrendorfer K, Ruckenbauer P (1994) The effect of fertilization on yield and quality of Miscanthus sinensis ‘Giganteus’. Ind Crop Prod 2:153–159
Staley TE, Stout WL, Jung GA (1991) Nitrogen use by tall fescue and switchgrass on acidic soils of varying water holding capacity. Agron J 83:732–738
Strullu L, Cadoux S, Preudhomme M et al (2011) Biomass production and nitrogen accumulation and remobilisation by Miscanthus×giganteus as influenced by nitrogen stocks in belowground organs. Field Crop Res 121:381–391
Stout WL, Jung GA (1995) Biomass and nitrogen accumulation in switchgrass: effects of soil and environment. Agron J 876:63–669
Thomason WE, Raun WR, Johnson GV, Taliaferro CM, Freeman KW, Wynn KJ, Mullen RW (2004) Switchgrass response to harvest frequency and time and rate of applied nitrogen. J Plant Nutr 27:1199–1226
Wullschleger SD, Davis EB, Borsuk ME, Gunderson CA, Lynd LR (2010) Biomass production in switchgrass across the United States: database description and determinants of yield. Agron J 102:1158–1168
Zub HW, Arnoult S, Brancourt-Hulmel M (2011) Key traits for biomass production identified in different miscanthus species at two harvest dates. Biomass Bioenergy 35:637–651
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article

Cite this article
Wang, Z., Jot Smyth, T., Crozier, C.R. et al. Yield and Nitrogen Removal of Bioenergy Grasses as Influenced by Nitrogen Rate and Harvest Management in the Coastal Plain Region of North Carolina. Bioenerg. Res. 11, 44–53 (2018). https://doi.org/10.1007/s12155-017-9876-x
Published:
Issue Date:
DOI: https://doi.org/10.1007/s12155-017-9876-x