
Abstract
Both childhood abuse and Catechol-O-methyltransferase (COMT) Val158Met polymorphism are implicated in aggression; however, their interactive effects on aggression and the related psychological mechanisms have not been clearly established. Furthermore, the Dark Triad traits (i.e., Machiavellianism, narcissism, and psychopathy), which are closely related to aggression, may mediate the abuse-aggression link. The current study aims to examine how Dark Triad traits mediate the childhood abuse × COMT polymorphism genotype-aggression link. A total of 680 incarcerated Chinese males completed the Reactive-Proactive Aggression Questionnaire, Adverse Childhood Experiences Questionnaire, and the Dirty Dozen Scale. DNA was extracted from their venous blood. When controlling for the inter-correlation between the two types of aggression, the results show indirect effects of childhood abuse on proactive aggression through all three Dark Triad traits, but only through narcissism on reactive aggression. Additionally, the COMT Val158Met polymorphism moderated the relation between childhood abuse and psychopathy; specifically, carriers of the Val/Val homozygote who perceived higher levels of childhood abuse demonstrated higher levels of psychopathy, which was related to higher proactive aggression. These findings have implications for understanding mechanisms of the abuse-aggression link.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Aggression is a risk factor for violent crime and violent behavior in jail (Sarchiapone et al., 2009) and has serious ramifications for inmates’ well-being and safety. It is of paramount importance to have a parsimonious theoretical and conceptual understanding of the antecedents of aggression among inmates to help improve forensic risk assessment and prison security (Gardner et al., 2015). Biological and environmental factors are posited to influence proximal personality traits and situational perceptions that ultimately increase aggression (Allen et al., 2018). Thus, biological factors (e.g., genetic polymorphisms) may interact with environmental exposures (e.g., gene-environment interaction) to influence aggressive traits. A particularly relevant environmental contributor to aggression is childhood abuse, as the prevalence of childhood abuse is higher among incarcerated samples compared to the general population (Bodkin et al., 2019; Wolff & Shi, 2012). However, not everyone who experiences abuse goes on to engage in aggressive behavior (Choi et al., 2019). This could be explained by the moderating effects of genetic vulnerabilities on violent behavior or aggressive traits (Caspi et al., 2002; Moffitt, 2005). To this end, the current study further investigates the moderating role of genotype and the mediating role of personality on the relations between childhood abuse and aggression among a sample of incarcerated males.
Childhood abuse is an environmental modifier empirically associated with aggressive behavior (Debowska et al., 2017). A recent meta-analysis indicates that individuals exposed to maltreatment experiences during childhood are more likely to display violent behaviors in adulthood (Fitton et al., 2020). However, this observed maltreatment-aggression link may depend on the type of maltreatment. A meta-analysis of 33 longitudinal studies found that physical, sexual, and emotional childhood abuse are more strongly associated with aggressive behaviors than general antisocial behaviors, while neglect increased the risk of general antisocial involvement (Braga et al., 2017). Thus, it appears that physical, sexual, and emotional forms of childhood abuse may act as environmental modifiers contributing to aggression.
Genetic dispositions may be one of the most salient biological factors associated with aggression as findings consistently indicate that aggression is partly heritable, with genetic factors explaining nearly 50% of the observed variance in aggression (Odintsova et al., 2019). The catechol-O-methyltransferase (COMT) gene, in particular, may be the most relevant candidate genes related to aggression (Qayyum et al., 2015). The COMT gene is responsible for metabolizing catecholamine neurotransmitters like dopamine, which is essential to executive functioning in the prefrontal cortex (Diamond, 2007). Specifically, the valine (Val) allele (vs. Methionine [Met] allele) at codon 158 (i.e., rs4680) is associated with higher activation of COMT enzyme (Lachman et al., 1996), which increases prefrontal dopamine catabolism and reduces executive functioning (Chen et al., 2004; Goldberg et al., 2003). Impaired executive functioning (e.g., low self-control and planning) is related to antisocial behavior including criminality, physical aggression, and clinical syndromes such as psychopathy (Ogilvie et al., 2011).
Research examining the link between the COMT Val158Met polymorphisms and aggression yields mixed results. For example, Caspi et al. (2008) and Monuteaux et al. (2009) found that the Val homozygotes were more aggressive (e.g., more likely to engage in aggressive conduct behaviors and be convicted of criminal offenses) compared to the Met carriers in samples of individuals with attention-deficit/hyperactivity disorder. Conversely, Albaugh et al. (2010) found that youth carrying the Met allele scored higher on both direct (e.g., physically attacks people) and relational aggression (e.g., easily jealous) compared to the Val homozygotes. Disparities in the COMT-aggression link may suggest the interactive effect of genetic and environmental factors.
Interaction between the COMT Val158Met polymorphism and childhood abuse on aggression and aggression-related traits varies across studies. For example, the Val homozygotes who experienced abuse or other stressors during childhood are more likely to display high levels of aggression compared to carriers of the Met allele (Hygen et al., 2015). Additional results in a sample of adults show that Val allele carriers who experienced sexual abuse have a greater disposition toward anger than the Met homozygotes (Perroud et al., 2010). Contrarily, studies in adolescent samples found that Met allele carriers, compared to the Val-homozygotes, display more externalizing problems (Nederhof et al., 2012) and higher reactive aggression (Zhang et al., 2016) after experiencing childhood stressors, such as parental divorce (Nederhof et al., 2012) and low positive parenting (Zhang et al., 2016). These inconsistent findings may be attributed to the use of different samples and varying measures of aggression (Qayyum et al., 2015). Taken together, it seems opportune for future research to narrow the focus of the COMT-aggression link to specific aggression types within specific populations (Qayyum et al., 2015), such as incarcerated males with relatively higher aggression.
Aggression is often classified as being either reactive or proactive (Dodge & Coie, 1987; Fite et al., 2018). Reactive aggression is considered a “hot” and retaliatory form of aggression, and is typically conceptualized within the frustration-aggression model (Gelles & Berkowitz, 1994). Alternatively, proactive aggression is framed within social learning paradigms (Berkowitz, 1978; Card & Little, 2007; Fite et al., 2018) and involves goal-oriented and socially dominant aggression. Previous studies indicate that different types of childhood abuse may be differentially related to reactive and proactive aggression (Richey et al., 2016; Zhu et al., 2020). However, more research needs to test whether the interaction between childhood abuse and COMT influences the observed variation in the manifestation of these aggressive subtypes (Zhang et al., 2016).
Personality traits are potential mediators of the Gene × Environment effects on aggression (Allen et al., 2018). Three personality traits comprising the Dark Triad (i.e., Machiavellianism, narcissism, and psychopathy; Paulhus & Williams, 2002) are associated with childhood adversity (Csatho & Birkas, 2018) and aggression (Jones & Neria, 2015). The Dark Triad traits also seem to mediate the link between childhood abuse and cyberbullying (Kircaburun et al., 2019), as well as the link between early adversity and interpersonal aggression (Figueredo et al., 2020). From the perspective of the Life History Theory (Roff, 2002), the Dark Triad traits emerge when adapting to adverse, harsh, and unpredictable environments (Brumbach et al., 2009; Plouffe et al., 2020). Several studies report significant associations between the Dark Triad traits and different types of aggressive behavior, such as bullying (Baughman et al., 2012), cyberbullying (Kircaburun et al., 2019), and intimate partner violence (Kiire, 2017). Additionally, variance in narcissism and psychopathy is also explained by genetic and non-shared environmental factors, while Machiavellianism may only be attributable to environmental effects (Veselka et al., 2011). Taken together, these findings suggest that the Dark Triad traits may mediate the link between childhood abuse and trait aggression, however, it is not yet clear how the COMT Val158Met polymorphism modifies the abuse-aggression link as mediated by personality. Accordingly, this study tests whether the direct and indirect effect of childhood abuse on trait aggression is moderated by the COMT Val158Met polymorphism which is then be mediated by Dark Triad traits. Due to mixed results of the moderation effect by the COMT genotypes, no specific direction regarding the moderation effect was hypothesized.
Method
Participants
Participants were recruited from a centralized male prison in a southern province of China who were convicted of crimes in several cities and towns surrounding this province. Participants included male inmates without a recorded diagnosis of a severe somatic pathology or psychopathological disorder upon intake. Male inmates were chosen given that they display higher levels of aggression (Falk et al., 2017) and are more likely to commit violent crimes than female inmates (Anderson & Huesmann, 2007). To achieve a statistical power level of 0.80, with effect sizes (partial R square) between 0.01 to 0.03 and α = 0.05 for G × E interaction under the assumption of a linear multiple regression fixed model (Wang et al., 2019), power analysis using G*Power (Faul et al., 2009) indicated the recommended minimum sample size to detect gene-environment interaction effects was 256 to 779. Participants were recruited only if they were within one month of their imprisonment and had completed the required prison adaptation and health services administrative courses.
Invitation letters were distributed to participants following completion of the prison adaption courses discussed above. An initial sample of 774 incarcerated Chinese males were contacted, with 752 of these inmates accepting the invitation for participation. These participants were provided with invitation letters and informed consent for the current study. Participants were compensated with a notepad as an incentive following data collection. Data collection was carried out by a group of trained researchers acquainted with the aims and measures of the study. Participants’ demographics (e.g., age, educational level, and criminal offenses) were provided by prison administrative staff. Participants’ biological material (i.e., three milliliters of venous blood) was collected by certified phlebotomists from the prison infirmary on a weekday morning during a routine physical examination. Biological material was then sent to the Northwest University for genotyping. All aspects of this study were approved by the Ethics Committee of the College of Psychology, Shenzhen University.
The final analytic sample included 680 participants (Mage = 32.49 years, SD = 9.20, 18–74 years; 5.3% of non-Han ethnicity, n = 36). A total of 72 participants were removed from the initial recruitment sample because they either voluntarily dropped out of the study, provided incomplete responses to questionnaires, or did not provide biological material. The education levels of the analytic sample were as follows: 29.9% below middle school, 57.8% middle school, and 12.3% above middle school (including vocational school, high school, and college). Participants were incarcerated for a variety of offenses including violent crimes (n = 196, 28.8%), property crimes (n = 327, 48.1%), drug crimes (n = 73, 10.7%), sex crimes (n = 38, 5.6%), and other crimes (n = 46, 6.8%).
Measures
Childhood Abuse
Childhood abuse experiences were assessed with the abuse subscale of the Chinese version (Fan et al., 2011) of the Adverse Childhood Experiences Questionnaire (Dong et al., 2004). The abuse subscale includes two items, respectively, for emotional and physical abuse with five response categories (never = 1, once or twice = 2, sometimes = 3, often = 4, or very often = 5), and four items for sexual abuse with two responses (yes = 1, no = 0). Experiencing each type of abuse was coded as 1, else as 0, according to the criteria by Dong et al. (2004), which generates three binary indicators. Confirmatory factor analysis indicated that the three-factor abuse subscale provided a good fit to our data (χ2 = 21.231, df = 17, RMSEA = 0.02, CFI = 0.99, TLI = 0.99, SRMR = 0.06). Referring to the previous study (Cheong et al., 2017), the three abuse types were averaged (Cronbach’s α = 0.72) to obtain a score of childhood abuse (M = 0.19, SD = 0.26).
Reactive and Proactive Aggression
Reactive and proactive aggression was assessed with the Chinese version (Zhang et al., 2014) of the Reactive–Proactive Aggression Questionnaire (Raine et al., 2006). This scale consists of 23 items, scored on a 3-point Likert scale ranging from 0 (never) to 2 (often). Confirmatory factor analysis indicated that the two-factor construct provided a good fit to our data (χ2 = 735.69, df = 229, RMSEA = 0.06, CFI = 0.89, TLI = 0.88, SRMR = 0.05). The Cronbach’s α was 0.84 for reactive aggression and 0.87 for proactive aggression. Scales were averaged into reactive (M = 0.82, SD = 0.37) and proactive (M = 0.21, SD = 0.29) aggression scores, with a higher value indicating higher levels of trait aggression.
Dark Triad Traits
The Dark Triad traits (i.e., Machiavellianism, narcissism, and psychopathy) were assessed by the Chinese version (Geng et al., 2015) of the Dirty Dozen scale (Jonason & Webster, 2010). The Dirty Dozen scale includes 12 items with four items for each trait, scored on a Likert scale ranging from 1 (strongly disagree) to 7 (strongly agree). Confirmatory factor analysis indicated that the three-factor model provided a good fit to our data (χ2 = 240.73, df = 51, RMSEA = 0.07, CFI = 0.92, TLI = 0.89, SRMR = 0.05). The Cronbach’s α for Machiavellianism, narcissism, and psychopathy were 0.80, 0.84, and 0.76, respectively. An average score was derived for each trait (Machiavellianism, M = 2.30, SD = 1.29; narcissism, M = 4.34, SD = 1.65; psychopathy, M = 2.53, SD = 1.39), with higher scores indicating higher levels for that specific trait.
Genotyping
Genomic DNA was extracted from venous blood using the TIANamp Genomic DNA Kit (DP304; Tiangen Biotech Co., Ltd., Beijing, China). The COMT Val158Met polymorphism was amplified by polymerase chain reaction (PCR) with an upstream primer, 5’-CCAGCGGATGGTGGATTTCGCACGC-3’ and a downstream primer 5’-TGGGGGGGTCTTTCCTCAGCC-3’. The genotyping procedures and details were described in a previous study (Gong et al., 2013). Upon genotyping, six samples were randomly selected from each genotype group for sequencing to determine the specific genotypes. The genotypes were scanned with the Multi-Spectral imaging System. Participants genotypes were classified as the Val/Val (n = 396, 58.2%), Met/Val (n = 220, 32.4%), and Met/Met (n = 64, 9.4%), which is consistent with the distribution of this genotype across other samples taken from Asian ancestries (Sindermann et al., 2018). The distribution of the genotype deviated from the Hardy–Weinberg equilibrium (χ2 = 15.387, p < 0.001). Owing to a small number of participants with the Met/Met genotype, participants’ genotypes were coded as 0 = Met allele carriers (Met/Met and Met/Val) and 1 = Val homozygotes (Val/Val) according to the previous studies (Hygen et al., 2015; Zhang et al., 2016).
Statistical Analyses
Continuous variables (age, childhood abuse, proactive and reactive aggression, and Dark Triad traits) and the ordinal variable (educational level) were standardized prior to analyses, to create similar scales and to avoid multicollinearity. Considering the correlation between proactive and reactive aggression, we took standardized residuals while controlling for the other aggression scale as an additional indicator of the outcomes in the following analysis (Raine et al., 2006). This method allows us to assess the “refined” proactive or reactive aggression measure independent of the other aggression scale effect. The analyses then proceeded in a series of linked steps. First, we ruled out potential gene-environment correlations (rGE) by assessing bivariate correlations and ANOVAs (Kim-Cohen et al., 2006) between the COMT genotype and all the variables in the analysis. Following this, and in line with suggestions from previous candidate gene-environment interaction (cGxE) studies (Dick et al., 2015), we then entered gene-by-covariate (i.e., age, educational level) and environment-by-covariate interaction terms into a series of conditional process models. The first model was a mediation model (Model 4 in PROCESS 3.5 macro; Hayes, 2013) examining the mediating roles of Dark Triad traits (measured with the Dirty Dozen scale) on the relation between abuse and reactive/proactive aggression. Then, a moderation model (Model 1 in PROCESS) was built to test the interactive effects of childhood abuse and the COMT genotype on reactive/proactive aggression and Dark Triad traits. Finally, a conditional process moderated-mediation model (Model 8 in PROCESS) was applied to assess whether the Dark Triad traits mediated the abuse-aggression relation as moderated by the COMT genotype.
We used the Bias-Corrected Bootstrap approach with 10,000 bootstrap resamples to compute the 95% confidence intervals (CI) for estimating effects. If the 95% CI does not contain zero, then the direct or indirect effects are considered significant. The index of moderated-mediation (Hayes, 2015) was calculated to examine differences in indirect effects between the two genetic groups. An index that is significantly different from zero indicates that the indirect effects are significantly different between the two genotypes. As our participants included non-Han ethnicities (n = 36), the robustness of results were examined through a sensitivity analysis (Zhang et al., 2018) after excluding these participants.
Results
Preliminary Analysis
Table S1 presents means, SDs, and bivariate correlations among study variables. The COMT Val158Met polymorphism was not significantly correlated with childhood abuse (r = − 0.001, p = 0.990), and there was no significant difference in childhood abuse between the two COMT genotypes (MMet carriers = 0.185 vs. MVal/Val = 0.185, t = 0.013, p = 0.990). These results indicate that any rGE effects would be small in the present study, though correlations among unmeasured genes and environments (e.g., parental phenotypes and positive environment) cannot be ruled out.
Simple Mediation Analyses
As shown in Figs. 1A and 1B, Machiavellianism, narcissism, and psychopathy mediated the abuse-aggression links (b = 0.030–0.044 for reactive aggression and b = 0.012–0.077 for proactive aggression). Childhood abuse had a significant direct effect on reactive aggression (b = 0.146, SE = 0.034, p < 0.001) and proactive aggression (b = 0.069, SE = 0.034, p = 0.042). When controlling the inter-correlation between the two aggression types, different paths emerged (see Fig. 1C and 1D). Specifically, only narcissism mediated the abuse-reactive aggression link (b = 0.046, CI [0.022, 0.076]), while all the Dark Triad traits mediated the abuse-proactive aggression link (b = 0.073, CI [0.039, 0.115] for Machiavellianism; b = − 0.018, CI [− 0.035, − 0.006] for narcissism; b = 0.037, CI [0.017, 0.061] for psychopathy). Childhood abuse had a significant direct effect on “refined” reactive aggression (b = 0.131, SE = 0.038, p < 0.001), but not on “refined” proactive aggression (b = − 0.023, SE = 0.037, p = 0.536), which implies narcissism partially mediated the abuse-reactive aggression link, while all the Dark Triad traits fully mediated the abuse-proactive aggression link.

The simple mediation model: Effects of childhood abuse through Machiavellianism, narcissism, and psychopathy on reactive (panel A), proactive (panel B), “refined” reactive (panel C), and “refined” proactive (panel D) aggression. Note. 95% Boot CI of indirect effects are shown in brackets with significant coefficients highlighted in bold. *p < .05. **p < .01. ***p < .001
Simple Moderation Analyses
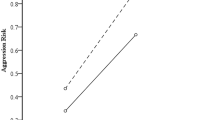
Simple moderation analyses (see Table S2 and S3) showed that the COMT genotype did not moderate the relationship between abuse and aggression (b = 0.101, SE = 0.076, p = 0.185 for reactive aggression; b = 0.149, SE = 0.076, p = 0.050 for proactive aggression). Moderation effects remained the same for the “refined” types of aggression. The COMT genotype moderated the association between childhood abuse and psychopathy (b = 0.162, SE = 0.075, p = 0.031), but not Machiavellianism (b = 0.137, SE = 0.076, p = 0.072) or narcissism (b = − 0.009, SE = 0.078, p = 0.910). Simple slopes tests showed that the effect of childhood abuse on psychopathy was larger among the Val/Val carriers (b = 0.274, SE = 0.048, p < 0.001) than the Met carriers (b = 0.111, SE = 0.058, p = 0.055).
Moderated Mediation Analyses
As shown in Figs. 2A and 2B, the indices of moderated mediation were only significantly different from zero for psychopathy (reactive aggression: index = 0.033, CI [0.003, 0.070]; proactive aggression: index = 0.043, CI [0.004, 0.086]), but not for Machiavellianism (reactive aggression: index = 0.015, CI [− 0.002, 0.042]; proactive aggression: index = 0.041, CI [− 0.004, 0.099]) or narcissism (reactive aggression: index = − 0.003, CI [− 0.049, 0.045]; proactive aggression: index = − 0.001, CI [− 0.014, 0.014]). Moderated mediation analyses (see Table S4) showed that the conditional indirect effects of childhood abuse on aggression via psychopathy were higher in the Val homozygotes (b = 0.056, CI [0.028, 0.091] for reactive aggression; b = 0.072, CI [0.040, 0.110] for proactive aggression) than in the Met carriers (b = 0.023, CI [0.001, 0.051] for reactive aggression; b = 0.029, CI [0.001, 0.062] for proactive aggression). The COMT genotype did not moderate the direct effect of childhood abuse on reactive (b = 0.055, SE = 0.067, p = 0.413) and proactive aggression (b = 0.067, SE = 0.066, p = 0.315).

The moderated mediation model: Effects of childhood abuse through Machiavellianism, narcissism, and psychopathy on reactive (panel A), proactive (panel B), “refined” reactive (panel C), and “refined” proactive (panel D) aggression conditionally. Note. Index indicates the moderated mediation effects, and 95% Boot CI of the index is shown in brackets with significant coefficients highlighted in bold. *p < .05. **p < .01. ***p < .001
As shown in Figs. 2C, 2D, and Table S5, when controlling the inter-correlation between the two types of aggression, the direct and indirect effects for both genotypes remained similar to those found in the simple mediation model (see Figs. 1C, 1D). The only exception was the moderated indirect effect via psychopathy (index = 0.028, CI [0.003, 0.062]): The conditional indirect effect of childhood abuse on “refined” proactive aggression was higher in the Val homozygotes (b = 0.048, CI [0.022, 0.080]) than in the Met carriers (b = 0.020, CI [0.001, 0.044]).
Supplementary Analyses
The sensitivity analyses indicated that the results of the moderated mediation models for the “refined” reactive and proactive aggression remained almost the same (see supplementary material Table S6 and Figure S1) after excluding participants of non-Han ethnicity (n = 36).
Discussion
This study examined relations between childhood abuse and aggression in a sample of incarcerated males. We found that the Dark Triad traits mediated the abuse-aggression link and that the COMT Val158Met polymorphism moderated the effect of childhood abuse on psychopathy. Specifically, among those who reported higher levels of childhood abuse, carriers of the Val homozygote showed higher levels of psychopathy and higher levels of aggression compared to the Met carriers. We found additional distinct paths after controlling for the inter-correlation between reactive and proactive aggression. Specifically, the three Dark Triad traits mediated the abuse and “refined” proactive aggression link, with a relatively greater effect through psychopathy for the Val carriers. Only narcissism mediated the abuse and “refined” reactive aggression link.
Similar to previous studies (Figueredo et al., 2020; Kircaburun et al., 2019), simple mediation model in the current study found that the Dark Triad traits mediated the abuse-aggression relation. In other words, individuals who experienced more childhood abuse had higher Dark Triad traits which, in turn, were related to higher levels of aggression. These findings is consistent with Life History Theory (Roff, 2002), and suggest a possibility that experiences of abuse may increase the adoption of fast life history strategies. Fast life history strategies are characterized by present-oriented behaviors and proclivities, such as those captured by the Dark Triad, aimed at maximizing immediate rewards and mating efforts (Csatho & Birkas, 2018; Jonason et al., 2010). Thus aggression, a fast strategy to survive in an early environment that is perceived as threatening, may be adopted by people who have higher Dark Triad traits (McDonald et al., 2012). However, further research is needed to test the potential causality from childhood abuse to Dark Triad, because a reverse or bidirectional path from the Dark Triad to childhood abuse cannot be completely ruled out given the correlational nature of cross-sectional design in the current study.
For the “refined” types of aggression, all the three Dark Triad traits fully mediated the abuse-proactive aggression link, while only narcissism partially mediated the abuse-reactive aggression in male inmates. This finding adds an important contribution to the literature on linkages among childhood abuse, Dark Triad traits, and aggression in samples of inmates, who have relatively high aggression, and even more robust links between childhood maltreatment and psychopathic traits (Plouffe et al., 2020). Specifically, it suggests that incarcerated Chinese males may engage in more reactive and proactive aggression via the different links between early exposure to childhood abuse and Dark Triad traits. This is consistent with prior research which reports that, among males, psychopathy was strongly associated with proactive aggression, while agreeableness was the most important predictor for reactive aggression (Dinic & Wertag, 2018). Broadly, the current findings further suggest that childhood abuse may influence development of an impaired sense of morality and even less empathy, which is related to higher proactive aggression, in male inmates. Future research should test these findings in general populations of males and females.
Notably, childhood abuse was positively associated with higher narcissism, which was related to higher “refined” reactive aggression but lower “refined” proactive aggression. Within the theoretical framework of Dark Triad traits, narcissism involves a grandiose sense of self (e.g., arrogance, Du et al., 2021). This type of grandiose narcissism is associated with both types of aggression, but in opposite directions across different studies (e.g., positive in Du et al., 2021 and negative in Knight et al., 2018). The current study supports the findings that grandiose narcissism, which is relatively higher in men (Grijalva et al., 2015), may develop as a coping mechanism to childhood abuse (Keene & Epps, 2016; Kernberg, 2014). Narcissism and reactive aggression may act as a kind of defensive response to extreme environmental adversity (Csatho & Birkas, 2018; Kjærvik & Bushman, 2021). Individuals may also report using fewer proactively aggressive strategies when faced with a need to achieve social (e.g., narcissistic) goals which may also coincide with a belief in not harming others seen as inferior to themselves (Knight et al., 2018). Alternatively, the partial mediating effect of reactive aggression may indicate that other paths explain relations between childhood abuse and reactive aggression. Different traits and associated paths like this should be investigated in the future studies. It is also noteworthy that proactive and reactive aggression often share mixed motives (Kjærvik & Bushman, 2021). Hence, future works should explore the motivations for using different types of aggression when examining the abuse-aggression link.
The findings did not replicate previous research on the moderating role of the Val158Met polymorphism on the direct link between abuse and aggression (e.g., Hygen et al., 2015; Perroud et al., 2010). However, results did support the finding that the Val158Met polymorphism moderated the effect of abuse on psychopathy, which was associated with increased proactive aggression. Consistent with Zhang et al. (2018), it seems possible that there could be a decrease in executive function (e.g., behavioral shift) in carriers of the Val homozygotes who experienced childhood abuse compared to carriers of the Met homozygote. This also further supports findings that the Val homozygotes with higher COMT enzyme activity have worse performance in executive functions (Chen et al., 2004; Goldberg et al., 2003), which is associated with psychopathy (Ogilvie et al., 2011). Thus, Val/Val carriers may be more likely to develop personality traits consistent with psychopathy in response to early adversity.
Collectively, the current study found abuse-aggression links among incarcerated Chinese males, with different mediation effects of the Dark Triad traits for reactive and proactive aggression. The COMT genotype also moderated the mediating effect of psychopathy on proactive aggression. These findings support Life History Theory, indicating that early environmental adversity can influence Dark Triad traits, which are then related to aggression in adulthood especially among samples of incarcerated males. These findings contribute to a more comprehensive view of the antecedents of trait aggression for male inmates who display relatively high trait aggressiveness. Specifically, childhood abuse is associated with higher reactive aggression via narcissism and higher proactive aggression via all the Dark Triad traits. Increased psychopathy was especially pronounced among the Val homozygotes with childhood abuse.
Limitations
There were several limitations to consider when interpreting the findings of this study. First, the sample used for genotyping might not be representative of all incarcerated populations or general populations, as it only consisted of male inmates. Additionally, genotype distributions showed a significant deviation from Hardy–Weinberg Equilibrium. This is most likely due to the unique homogeneity of the sample. Future studies need to examine whether the gene-moderated mediation path from childhood abuse to aggression through Dark Triad traits generalizes to female inmates and broader populations. Second, the current study only examined one candidate gene (i.e., COMT) and the findings have not been replicated in an independent sample due to limited sample sizes of inmates. As suggested by Dick et al. (2015), using genome-wide analyses (GWAS) and replication samples are warranted to corroborate and extend the present findings. Third, the effect of rGE and other confounders cannot be entirely ruled out because other relevant covariates were unavailable (e.g., familial phenotypes, substance use, prior criminal history). Fourth, causal relationships cannot be established based on the current cross-sectional study with self-reported measures only. Future longitudinal studies using other methods for measuring childhood abuse are recommended.
Research Implications
Despite the limitations, this study adds to the literature examining mechanisms associated with the relation between abuse and aggression among incarcerated males. Using a cross-sectional design, we tested whether the COMT genotype would moderate the effect of childhood abuse on proactive and reactive aggression through the Dark Triad traits in a sample of incarcerated males. In addition to observed differences in the paths for reactive and proactive aggression, we found that the effects of childhood abuse on proactive aggression via psychopathy was larger for the Val homozygotes. These findings provide evidence for indirect and direct relations among environmental, genetic, and personality factors on aggressive traits. These findings may be helpful for developing risk assessment tools and tailored behavioral interventions for male inmates.
Availability of data and material
Due to the sensitive nature of participants, the data cannot be available to access in public.
Code availability
Not applicable.
References
Albaugh, M. D., Harder, V. S., Althoff, R. R., Rettew, D. C., Ehli, E. A., Lengyel-Nelson, T., Davies, G. E., Ayer, L., Sulman, J., Stanger, C., & Hudziak, J. J. (2010). COMT Val158Met genotype as a risk factor for problem behaviors in youth. Journal of the American Academy of Child and Adolescent Psychiatry, 49(8), 841–849. https://doi.org/10.1016/j.jaac.2010.05.015
Allen, J. J., Anderson, C. A., & Bushman, B. J. (2018). The General Aggression Model. Current Opinion in Psychology, 19, 75–80. https://doi.org/10.1016/j.copsyc.2017.03.034
Anderson, C. A., & Huesmann, L. R. (2007). Human aggression: A social-cognitive view. In The SAGE Handbook of Social Psychology: Concise Student Edition (pp. 259–288). SAGE Publications Ltd. https://doi.org/10.4135/9781848608221.n12
Baughman, H. M., Dearing, S., Giammarco, E., & Vernon, P. A. (2012). Relationships between bullying behaviours and the Dark Triad: A study with adults. Personality and Individual Differences, 52(5), 571–575. https://doi.org/10.1016/j.paid.2011.11.020
Berkowitz, L. (1978). Whatever happened to the frustration-aggression hypothesis? American Behavioral Scientist, 21(5), 691–708. https://doi.org/10.1177/000276427802100505
Bodkin, C., Pivnick, L., Bondy, S. J., Ziegler, C., Martin, R. E., Jernigan, C., & Kouyoumdjian, F. (2019). History of childhood abuse in populations incarcerated in Canada: A systematic review and meta-analysis. American Journal of Public Health, 109(3), e1–e11. https://doi.org/10.2105/AJPH.2018.304855
Braga, T., Gonçalves, L. C., Basto-Pereira, M., & Maia, Â. (2017). Unraveling the link between maltreatment and juvenile antisocial behavior: A meta-analysis of prospective longitudinal studies. Aggression and Violent Behavior, 33, 37–50. https://doi.org/10.1016/j.avb.2017.01.006
Brumbach, B. H., Figueredo, A. J., & Ellis, B. J. (2009). Effects of harsh and unpredictable environments in adolescence on development of life history strategies. Human Nature, 20(1), 25–51. https://doi.org/10.1007/s12110-009-9059-3
Card, N., & Little, T. (2007). Differential relation of instrumental and reactive aggression with maladjustment: Does adaptivity depend of function? In Aggression and Adaptation: The Bright Side to Bad Behavior (pp. 107–134). https://doi.org/10.4324/9780203936900
Caspi, Avshalom, McClay, Joseph, Moffitt, Terrie E.., Mill, Jonathan, Martin, Judy, Craig, Ian W.., et al. (2002). Role of genotype in the cycle of violence in maltreated children. Science, 297(5582), 851–854. https://doi.org/10.1126/science.1072290
Caspi, A., Langley, K., Milne, B., Moffitt, T. E., O’Donovan, M., Owen, M. J., Tomas, M. P., Poulton, R., Rutter, M., Taylor, A., Williams, B., & Thapar, A. (2008). A replicated molecular genetic basis for subtyping antisocial behavior in children with attention-deficit/hyperactivity disorder. Archives of General Psychiatry, 65(2), 203–210. https://doi.org/10.1001/archgenpsychiatry.2007.24
Chen, J., Lipska, B. K., Halim, N., Ma, Q. D., Matsumoto, M., Melhem, S., Kolachana, B. S., Hyde, T. M., Herman, M. M., Apud, J., Egan, M. F., Kleinman, J. E., & Weinberger, D. R. (2004). Functional analysis of genetic variation in Catechol-O-Methyltransferase (COMT): Effects on mRNA, protein, and enzyme activity in postmortem human brain. The American Journal of Human Genetics, 75(5), 807–821. https://doi.org/10.1086/425589
Cheong, E. V., Sinnott, C., Dahly, D., & Kearney, P. M. (2017). Adverse childhood experiences (ACEs) and later-life depression: Perceived social support as a potential protective factor. British Medical Journal Open, 7(9), e013228. https://doi.org/10.1136/bmjopen-2016-013228
Choi, K. W., Stein, M. B., Dunn, E. C., Koenen, K. C., & Smoller, J. W. (2019). Genomics and psychological resilience: A research agenda. Molecular Psychiatry, 24(12), 1770–1778. https://doi.org/10.1038/s41380-019-0457-6
Csatho, A., & Birkas, B. (2018). Early-life stressors, personality development, and fast life Strategies: An evolutionary perspective on malevolent personality features. Frontiers in Psychology, 9, 305. https://doi.org/10.3389/fpsyg.2018.00305
Debowska, A., Willmott, D., Boduszek, D., & Jones, A. D. (2017). What do we know about child abuse and neglect patterns of co-occurrence? A systematic review of profiling studies and recommendations for future research. Child Abuse & Neglect, 70, 100–111. https://doi.org/10.1016/j.chiabu.2017.06.014
Diamond, A. (2007). Consequences of variations in genes that affect dopamine in prefrontal cortex. Cerebral Cortex, 17(suppl_1), i161–i170. https://doi.org/10.1093/cercor/bhm082
Dick, D. M., Agrawal, A., Keller, M. C., Adkins, A., Aliev, F., Monroe, S., Hewitt, J. K., Kendler, K. S., & Sher, K. J. (2015). Candidate gene–environment interaction research: Reflections and recommendations. Perspectives on Psychological Science, 10(1), 37–59. https://doi.org/10.1177/1745691614556682
Dinic, B. M., & Wertag, A. (2018). Effects of Dark Triad and HEXACO traits on reactive/proactive aggression: Exploring the gender differences. Personality and Individual Differences, 123, 44–49. https://doi.org/10.1016/j.paid.2017.11.003
Dodge, K., & Coie, J. (1987). Social-information-processing factors in reactive and proactive aggression in children’s peer groups. Journal of Personality and Social Psychology, 53(6), 1146–1158. https://doi.org/10.1037/0022-3514.53.6.1146
Dong, M., Anda, R. F., Felitti, V. J., Dube, S. R., Williamson, D. F., Thompson, T. J., Loo, C. M., & Giles, W. H. (2004). The interrelatedness of multiple forms of childhood abuse, neglect, and household dysfunction. Child Abuse & Neglect, 28(7), 771–784. https://doi.org/10.1016/j.chiabu.2004.01.008
Du, T. V., Miller, J. D., & Lynam, D. R. (2021). The relation between narcissism and aggression: A meta‐analysis. Journal of Personality, jopy.12684. https://doi.org/10.1111/jopy.12684
Falk, Ö., Sfendla, A., Brändström, S., Anckarsäter, H., Nilsson, T., & Kerekes, N. (2017). Personality and trait aggression profiles of male and female prison inmates. Psychiatry Research, 250, 302–309. https://doi.org/10.1016/j.psychres.2016.12.018
Fan, Y., Pan, H., Zhang, Z., Zhang, N., Wang, C., & Ye, D. (2011). Correlative analysis between ACEs and coping styles among medicos. Chin J Sch Health, 32(07), 774–776.
Faul, F., Erdfelder, E., Buchner, A., & Lang, A.-G. (2009). Statistical power analyses using G*Power 3.1: Tests for correlation and regression analyses. Behavior Research Methods, 41(4), 1149–1160. https://doi.org/10.3758/BRM.41.4.1149
Figueredo, A. J., Black, C. J., Patch, E. A., Heym, N., Ferreira, J. H. B. P., Varella, M. A. C., Defelipe, R. P., Cosentino, L. A. M., Castro, F. N., Natividade, J. C., Hattori, W. T., Pérez-Ramos, M., Madison, G., & Fernandes, H. B. F. (2020). The cascade of chaos: From early adversity to interpersonal aggression. Evolutionary Behavioral Sciences. https://doi.org/10.1037/ebs0000241
Fite, P. J., Craig, J., Colder, C. R., Lochman, J. E., & Wells, K. C. (2018). Proactive and reactive aggression. In R. J. R. Levesque (Ed.), Encyclopedia of Adolescence (pp. 2870–2879). Springer International Publishing. https://doi.org/10.1007/978-3-319-33228-4_211
Fitton, L., Yu, R., & Fazel, S. (2020). Childhood maltreatment and violent outcomes: A systematic review and meta-analysis of prospective studies. Trauma Violence & Abuse, 21(4), 754–768. https://doi.org/10.1177/1524838018795269
Gardner, B. O., Boccaccini, M. T., Bitting, B. S., & Edens, J. F. (2015). Personality Assessment Inventory scores as predictors of misconduct, recidivism, and violence: A meta-analytic review. Psychological Assessment, 27(2), 534–544. https://doi.org/10.1037/pas0000065
Gelles, R. J., & Berkowitz, L. (1994). Aggression: Its causes, consequences, and control. Contemporary Sociology, 23(4), 575. https://doi.org/10.2307/2076412
Geng, Y., Sun, Q., Huang, J., Zhu, Y., & Han, X. (2015). Dirty Dozen and Short Dark Triad: A Chinese validation of two brief measures of the Dark Triad. Chinese Journal of Clinical Psychology, 23(02), 246–250. https://doi.org/10.16128/j.cnki.1005-3611.2015.02.013
Goldberg, T. E., Egan, M. F., Gscheidle, T., Coppola, R., Weickert, T., Kolachana, B. S., Goldman, D., & Weinberger, D. R. (2003). Executive subprocesses in working memory: Relationship to Catechol- O-methyltransferase Val158Met genotype and schizophrenia. Archives of General Psychiatry, 60(9), 889. https://doi.org/10.1001/archpsyc.60.9.889
Gong, P., Shen, G., Li, S., Zhang, G., Fang, H., Lei, L., Zhang, P., & Zhang, F. (2013). Genetic Variations in COMT and DRD2 Modulate Attentional Bias for Affective Facial Expressions. PLoS ONE, 8(12), e81446. https://doi.org/10.1371/journal.pone.0081446
Grijalva, E., Newman, D. A., Tay, L., Donnellan, M. B., Harms, P. D., Robins, R. W., & Yan, T. (2015). Gender differences in narcissism: A meta-analytic review. Psychological Bulletin, 141(2), 261–310. https://doi.org/10.1037/a0038231
Hayes, A. F. (2013). Introduction to mediation, moderation, and conditional process analysis: A regression-based approach (pp. xvii, 507). Guilford Press.
Hayes, A. F. (2015). An index and test of linear moderated mediation. Multivariate Behavioral Research, 50(1), 1–22. https://doi.org/10.1080/00273171.2014.962683
Hygen, B. W., Belsky, J., Stenseng, F., Lydersen, S., Guzey, I. C., & Wichstrom, L. (2015). Child exposure to serious life events, COMT, and aggression: Testing differential susceptibility theory. Developmental Psychology, 51(8), 1098–1104. https://doi.org/10.1037/dev0000020
Jonason, P. K., Koenig, B. L., & Tost, J. (2010). Living a fast life. Human Nature, 21(4), 428–442. https://doi.org/10.1007/s12110-010-9102-4
Jonason, P. K., & Webster, G. D. (2010). The dirty dozen: A concise measure of the dark triad. Psychological Assessment, 22(2), 420–432. https://doi.org/10.1037/a0019265
Jones, D. N., & Neria, A. L. (2015). The Dark Triad and dispositional aggression. Personality and Individual Differences, 86, 360–364. https://doi.org/10.1016/j.paid.2015.06.021
Keene, A. C., & Epps, J. (2016). Childhood physical abuse and aggression: Shame and narcissistic vulnerability. Child Abuse & Neglect, 51, 276–283. https://doi.org/10.1016/j.chiabu.2015.09.012
Kernberg, O. F. (2014). An overview of the treatment of severe narcissistic pathology. The International Journal of Psychoanalysis, 95(5), 865–888. https://doi.org/10.1111/1745-8315.12204
Kiire, S. (2017). Psychopathy rather than Machiavellianism or narcissism facilitates intimate partner violence via fast life strategy. Personality and Individual Differences, 104, 401–406. https://doi.org/10.1016/j.paid.2016.08.043
Kim-Cohen, J., Caspi, A., Taylor, A., Williams, B., Newcombe, R., Craig, I. W., & Moffitt, T. E. (2006). MAOA, maltreatment, and gene-environment interaction predicting children’s mental health: New evidence and a meta-analysis. Molecular Psychiatry, 11(10), 903–913. https://doi.org/10.1038/sj.mp.4001851
Kircaburun, K., Jonason, P., Griffiths, M. D., Aslanargun, E., Emirtekin, E., Tosuntaş, Ş. B., & Billieux, J. (2019). Childhood emotional abuse and cyberbullying perpetration: The role of dark personality traits. Journal of Interpersonal Violence, 088626051988993. https://doi.org/10.1177/0886260519889930
Kjærvik, S. L., & Bushman, B. J. (2021). The link between narcissism and aggression: A meta-analytic review. Psychological Bulletin, 147(5), 477–503. https://doi.org/10.1037/bul0000323
Knight, N. M., Dahlen, E. R., Bullock-Yowell, E., & Madson, M. B. (2018). The HEXACO model of personality and Dark Triad in relational aggression. Personality and Individual Differences, 122, 109–114. https://doi.org/10.1016/j.paid.2017.10.016
Lachman, H., Papolos, D., Saito, T., Yu, Y.-M., Szumlanski, C., & Weinshilboum, R. (1996). Human catechol-O-methyltransferase pharmacogenetics: Description of a functional polymorphism and its potential application to neuropsychiatric disorders. Pharmacogenetics, 6(3), 243–250.
McDonald, M. M., Donnellan, M. B., & Navarrete, C. D. (2012). A life history approach to understanding the Dark Triad. Personality and Individual Differences, 52(5), 601–605. https://doi.org/10.1016/j.paid.2011.12.003
Moffitt, T. E. (2005). The New Look of Behavioral Genetics in Developmental Psychopathology: Gene-Environment Interplay in Antisocial Behaviors. Psychological Bulletin, 131(4), 533–554. https://doi.org/10.1037/0033-2909.131.4.533
Monuteaux, M. C., Biederman, J., Doyle, A. E., Mick, E., & Faraone, S. V. (2009). Genetic risk for conduct disorder symptom subtypes in an ADHD sample: Specificity to aggressive symptoms. Journal of the American Academy of Child & Adolescent Psychiatry, 48(7), 757–764. https://doi.org/10.1097/CHI.0b013e3181a5661b
Nederhof, E., Belsky, J., Ormel, J., & Oldehinkel, A. J. (2012). Effects of divorce on Dutch boys’ and girls’ externalizing behavior in Gene × Environment perspective: Diathesis stress or differential susceptibility in the Dutch Tracking Adolescents’ Individual Lives Survey study? Development and Psychopathology, 24(3), 929–939. https://doi.org/10.1017/S0954579412000454
Odintsova, V. V., Roetman, P. J., Ip, H. F., Pool, R., Van der Laan, C. M., Tona, K.-D., Vermeiren, R. R. J. M., & Boomsma, D. I. (2019). Genomics of human aggression: Current state of genome-wide studies and an automated systematic review tool. Psychiatric Genetics, 29(5), 170–190. https://doi.org/10.1097/YPG.0000000000000239
Ogilvie, J. M., Stewart, A. L., Chan, R. C. K., & Shum, D. H. K. (2011). Neuropsychological measures of executive function and antisocial behavior: A meta-analysis*. Criminology, 49(4), 1063–1107. https://doi.org/10.1111/j.1745-9125.2011.00252.x
Paulhus, D. L., & Williams, K. M. (2002). The Dark Triad of personality: Narcissism, Machiavellianism, and psychopathy. Journal of Research in Personality, 36(6), 556–563. https://doi.org/10.1016/S0092-6566(02)00505-6
Perroud, N., Jaussent, I., Guillaume, S., Bellivier, F., Baud, P., Jollant, F., Leboyer, M., Lewis, C. M., Malafosse, A., & Courtet, P. (2010). COMT but not serotonin-related genes modulates the influence of childhood abuse on anger traits. Genes, Brain and Behavior, 9(2), 193–202. https://doi.org/10.1111/j.1601-183X.2009.00547.x
Plouffe, R. A., Wilson, C. A., & Saklofske, D. H. (2020). Examining the relationships between childhood exposure to intimate partner violence, the Dark Tetrad of personality, and violence perpetration in adulthood. Journal of Interpersonal Violence, 0886260520948517. https://doi.org/10.1177/0886260520948517
Qayyum, A., Zai, C. C., Hirata, Y., Tiwari, A. K., Cheema, S., Nowrouzi, B., Beitchman, J. H., & Kennedy, J. L. (2015). The role of the Catechol-o-methyltransferase (COMT) gene Val158Met in aggressive behavior, a review of genetic studies. Current Neuropharmacology, 13(6), 802–814. https://doi.org/10.2174/1570159X13666150612225836
Raine, A., Dodge, K., Loeber, R., Gatzke-Kopp, L., Lynam, D., Reynolds, C., Stouthamer-Loeber, M., & Liu, J. H. (2006). The reactive-proactive aggression questionnaire: Differential correlates of reactive and proactive aggression in adolescent boys. Aggressive Behavior, 32(2), 159–171. https://doi.org/10.1002/ab.20115
Richey, A., Brown, S., Fite, P. J., & Bortolato, M. (2016). The role of hostile attributions in the associations between child maltreatment and reactive and proactive aggression. Journal of Aggression, Maltreatment & Trauma, 25(10), 1043–1057. https://doi.org/10.1080/10926771.2016.1231148
Roff, D. (2002). Life history evolution. Sinauer Associates. https://doi.org/10.1016/B978-0-12-384719-5.00087-3
Sarchiapone, M., Carli, V., Cuomo, C., Marchetti, M., & Roy, A. (2009). Association between childhood trauma and aggression in male prisoners. Psychiatry Research, 165(1), 187–192. https://doi.org/10.1016/j.psychres.2008.04.026
Sindermann, C., Luo, R., Zhang, Y., Kendrick, K. M., Becker, B., & Montag, C. (2018). The COMT Val158Met polymorphism and reaction to a transgression: Findings of genetic associations in both Chinese and German samples. Frontiers in Behavioral Neuroscience, 12, 148. https://doi.org/10.3389/fnbeh.2018.00148
Veselka, L., Schermer, J. A., & Vernon, P. A. (2011). Beyond the Big Five: The Dark Triad and the supernumerary personality inventory. Twin Research and Human Genetics, 14(2), 158–168. https://doi.org/10.1375/twin.14.2.158
Wang, M., Zheng, X., Xia, G., Liu, D., Chen, P., & Zhang, W. (2019). Association between negative life events and early adolescents’ depression: The moderating effects of Catechol-O-methyltransferase (COMT) Gene Val158Met polymorphism and parenting behavior. Acta Psychologica Sinica, 51(8), 903. https://doi.org/10.3724/SP.J.1041.2019.00903
Wolff, N., & Shi, J. (2012). Childhood and adult trauma experiences of incarcerated persons and their relationship to adult behavioral health problems and treatment. International Journal of Environmental Research and Public Health, 9(5), 1908–1926. https://doi.org/10.3390/ijerph9051908
Zhang, H., Li, J., Yang, B., Ji, T., Long, Z., Xing, Q., Shao, D., Bai, H., Sun, J., & Cao, F. (2018). The divergent impact of catechol-O-methyltransferase (COMT) Val158Met genetic polymorphisms on executive function in adolescents with discrete patterns of childhood adversity. Comprehensive Psychiatry, 81, 33–41. https://doi.org/10.1016/j.comppsych.2017.11.004
Zhang, W., Cao, C., Wang, M., Ji, L., & Cao, Y. (2016). Monoamine Oxidase A (MAOA) and Catechol-O-Methyltransferase (COMT) gene polymorphisms interact with maternal parenting in association with adolescent reactive aggression but not proactive aggression: Evidence of differential susceptibility. Journal of Youth and Adolescence, 45(4), 812–829. https://doi.org/10.1007/s10964-016-0442-1
Zhang, W., Jia, S., Chen, G., & Zhang, W. (2014). Reliability and validity of reactive-proactive aggression questionnaire in college students. Chinese Journal of Clinical Psychology, 22(02), 260–263. https://doi.org/10.16128/j.cnki.1005-3611.2014.02.022
Zhu, W., Chen, Y., & Xia, L.-X. (2020). Childhood maltreatment and aggression: The mediating roles of hostile attribution bias and anger rumination. Personality and Individual Differences, 162, 110007. https://doi.org/10.1016/j.paid.2020.110007
Acknowledgements
We thank the Key Laboratory of Resource Biology and Biotechnology in Western China (Ministry of Education) for their Genotyping analyses.
Funding
This study was supported by the National Natural Science Foundation of China [grant number 31700982, 31600928]; the Shenzhen Humanities & Social Sciences Key Research Bases of the Center for Mental Health, Shenzhen university; Natural Science Foundation of Guangdong Province (2022A1515011097) to Jinting Liu and the Ministry of Education of Humanities and Social Science Foundation (19YJAZH021) to Pingyuan Gong.
Author information
Authors and Affiliations
Contributions
Jieting Zhang: Conceptualization, Methodology, Writing—Original Draft, Supervision.
Junhui Leng:Data Curation, data analysis, Writing—Original Draft.
Cuimei Yang: Data curation, Validation, Writing—Review and Editing.
Eric M. Cooke: Writing – Review, Revision and Editing.
Pingyuan Gong: Methodology, Conceptualization, Genotyping.
Jinting Liu: Methodology, Conceptualization, Resources.
Corresponding authors
Ethics declarations
Ethical Approval
All aspects of this study were approved by the Ethics Committee of the College of Psychology, Shenzhen University.
Consent to participate
All participants were provided with invitation letters and informed consent which explicitly stated the purpose of the study, participation incentives, and the voluntary nature of their participation as well as their rights during the study and the confidentiality of their personal information used in the study.
Consent to publish
Informed consents were obtained from participants prior to the study.
Conflicts of interest
The authors declare that they have no conflict of interest.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Below is the link to the electronic supplementary material.
Rights and permissions
About this article

Cite this article
Zhang, J., Leng, J., Yang, C. et al. Dark Triad traits mediate the interaction between childhood abuse and COMT Val158Met polymorphism on aggression among incarcerated Chinese males. Curr Psychol 42, 21164–21174 (2023). https://doi.org/10.1007/s12144-022-03092-0
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s12144-022-03092-0