
Abstract
Humans are continuously exposed to ionizing radiation throughout life from natural sources that include cosmic, solar, and terrestrial. Much harsher natural radiation and chemical environments existed during our planet’s early years. Mammals survived the harsher environments via evolutionarily-conserved gifts ̶ a continuously evolving system of stress-induced natural protective measures (i.e., activated natural protection [ANP]). The current protective system is differentially activated by stochastic (i.e., variable) low-radiation-dose thresholds and when optimally activated in mammals includes antioxidants, DNA damage repair, p53-related apoptosis of severely-damaged cells, reactive-oxygen-species (ROS)/reactive-nitrogen-species (RNS)- and cytokine-regulated auxiliary apoptosis that selectively removes aberrant cells (e.g., precancerous cells), suppression of disease promoting inflammation, and immunity against cancer cells. The intercellular-signaling-based protective system is regulated at least in part via epigenetic reprogramming of adaptive-response genes. When the system is optimally activated, it protects against cancer and some other diseases, thereby leading to hormetic phenotypes (e.g., reduced disease incidence to below the baseline level; reduced pain from inflammation-related problems). Here, some expressed radiation hormesis phenotypes and related mechanisms are discussed along with their implications for disease prevention and therapy.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
“At the early stages of evolution, increasingly complex organisms developed powerful defense mechanisms against such adverse radiation effects as mutation and malignant change. These effects originate in the cell nucleus, where the DNA is their primary target. That evolution has apparently proceeded for so long is proof, in part, of the effectiveness of living things’ defenses against radiation” Zbigniew Jaworowski 1999
“The notion of radiation hormesis, that exposure to low levels of ionizing radiation could produce beneficial effects, developed seriously in the late 1950’s, and was, to most radiation scientists, incredible…More recent understanding of the mechanisms of radiation damage and repair, and discoveries of induction of gene expression by radiation and other genotoxic agents make it seem inevitable that under suitable conditions, irradiation will produce beneficial effects.” Howard Ducoff 2002
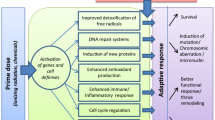
Life on Earth is estimated to have originated about 3.9 billion years ago in a more hostile natural radiation environment (Karam and Leslie 1999; Bhatia 2008; Jaworowski 2008). The radiation exposures involved both low linear-energy-transfer (LET) (e.g., beta and gamma radiations) and high-LET (e.g., alpha radiation) sources. The level of natural background radiation exposure of life forms during that era is estimated to have been five-fold larger than for recent times (Karam and Leslie 1999). Mammals later emerged, and in doing so, survived via adapting to the harsh radiation and also oxygen environments. The adaptation over a long period of time led to the present-day hierarchical system of mild-stress activated natural protection (ANP). The known components of the ANP system include antioxidants, DNA damage repair, p53-related apoptosis of severely damaged cells, p53-independent apoptosis of precancerous and other aberrant cells, suppression of disease promoting inflammation, and immunity against cancer (Kondo 1998; Wolff 1998; Sakai et al. 2003, 2006; Bauer 2007; Feinendegen et al. 2007a,b; Koana et al. 2007; Liu 2007; Cohen 2008; Nakatsukasa et al. 2008, 2010; Tubiana 2008; Averbeck 2009; Scott et al. 2009; Tubiana et al. 2009; Takatori et al. 2010; Calabrese and Calabrese 2013; Kataoka 2013; Calabrese and Dhawan 2014). The indicated components are differentially activated by low radiation doses and dose rates.
The radiation ANP system is responsible for the hormesis phenotype (mild-stress-stimulated reduced impairment status for and individual or reduced frequency of impairment among a population) for stochastic effects of irradiation such as cancer. Hormetic dose responses for stochastic radiation effects generally have a U or J shape, reflective of low-dose stimulation and high-dose inhibition (Luckey 1980; Ducoff 2002; Calabrese and Baldwin 1999, 2003) and the related stimulatory and inhibitory stochastic thresholds (Scott 2004, 2005; Scott et al. 2009).
Some key studies of components of the radiation ANP system are discussed in the next section in relation to different hormesis phenotypes expressed at the molecular, cellular, tissue, organ, or organism level. Also discussed is the current knowledge about biological mechanisms responsible for different radiation-stimulated hormesis phenotypes and the related recognized and potential health benefits, which are largely unknown by the majority of the publics worldwide. Papers evaluated were published as early as 1927 and as recent as 2014. Oddly, much of the cutting-edge research has been carried out in Japan, the only nation directly impacted by nuclear-weapons detonations during war. Only peer-reviewed publications were considered.
While most of the results presented are based on research by others, some of the results were derived by our research group which participated in the U. S. Department of Energy, Office of Science (BER) Low Dose Radiation Research Program that was halted by the recent government sequester.
Benefits of low and moderate radiation doses
Some studies demonstrating beneficial effects of low and moderate radiation doses are described in this section. Where mechanisms of radiation action have been proposed, they are also discussed.
Low-dose radiation stimulates antioxidant production, protecting from oxidative damage
Low-dose radiation induces antioxidant defenses (Yamaoka et al. 1998; Kojima et al. 1998a,b; Kataoka 2013). As an example, a whole-body X-ray dose of 200 mGy increased superoxide dismutase (SOD), glutathione peroxidase (GPx), and GPx mRNA in spleens of C57BL/6NJcl and BALB/c mice, which was not the case for a large dose of 4,000 mGy (Kataoka 2013). Another study suggested that the levels of reduced glutathione (GSH), glutathione reductase (GR), γ-glutamylcysteine synthetase (γ-GCS), and thioredoxin (TRX) increased in liver shortly after whole body irradiation with 500 mGy of gamma rays (Kojima et al. 1998a). In addition, the levels of GSH, GR, γ-GCS, and TRX increased in the brain shortly after 500 mGy of gamma rays (Kojima et al. 1998b). The activation of antioxidant functions is mediated via transcriptional regulation of the γ-GCS gene, predominantly through the activator protein-1 binding site in its promoter region (Kawakita et al. 2003). These findings support the view that exposure to low radiation doses (mild stresses) increase protective antioxidants which may contribute to hormetic phenotypes. Stochastic low-dose-radiation thresholds are likely involved as well as intercellular signaling, which may be epigenetically regulated (i.e., epiregulated; Scott et al. 2009).
Low-dose radiation activates DNA damage repair and protective apoptosis
DNA double strand breaks are considered the most serious type of genomic damage. Sophisticated homeostatic mechanisms have evolved to correct or compensate for such damage, which occurs with relative high frequency as a result of normal metabolic activities within cells and normal microenvironmental changes. Currently, there are three known mechanisms of repair of double-stand breaks: non-homologous end joining, microhomology-mediated end joining, and homologous recombination. Low-dose-radiation stochastic thresholds are likely involved in DNA double-strand-break repair activation and may involve intercellular communications arising as an epiregulated cell-community-wide (epicellcom) process (Scott 2011a).
Some information related to DNA damage repair activation by low radiation doses has been derived from studies of radiation-induced mutations. A sex-linked recessive lethal mutation assay was performed by Koana et al. (2007) in Drosophila melanogaster using immature spermatocytes and spermatogonia irradiation with 150-kVp X rays at a high (500 mGy/min) or low (50 mGy/min) rate. The mutation frequency in the sperm irradiated with a low dose at a low rate was significantly lower than that for controls, whereas irradiation with a high dose and rate resulted in a significant increase in the mutation frequency (i.e., hormetic response; low-dose protection and high-dose harm). When cells deficient in DNA excision repair were used instead of wild-type cells, low-dose irradiation at a low rate did not reduce the mutation frequency (i.e., no evidence for radiation ANP). These findings are consistent with the possibility that error-free DNA repair functions were activated as an epicellcom process by low-dose/low-dose-rate irradiation and that this repaired spontaneous DNA damage throughout the target cell population as well as radiation-related damage, thus producing a practical threshold for induced mutation-related harm (e.g., mutation-facilitated cancer). The findings are not in line with the widely-used, linear-no-threshold (LNT) hypothesis as it relates to mutation and cancer induction.
The LNT hypothesis was initially justified on the basis of the dose–response function for mutation induction in germ cells of Drosophila melanogaster interpreted to be of the LNT type based on the very high X-ray doses employed (Muller 1927, 1954); however, a more recent and more reliable study (Ogura et al. 2009) using gamma rays that included orders of magnitude lower radiation doses delivered at 22.4 mGy/h revealed that a strong adaptive response (hormetic phenotype) occurs at doses less than about 100 mGy with a significant reduction (p <0.01) in the mutation frequency to below the spontaneous level at a dose of 0.5 mGy. Because there is on average less than 1 electron track (from ionizations) per cell at the indicated absorbed dose, this is likely a protective bystander effect that relates to epigenetic activation (epiactivation) of adaptive-response genes (Scott et al. 2009). Thus, the initial basis for use of the LNT risk model has been invalidated. Interestingly, the 0.5 mGy dose up-regulated heat-shock-protein- and apoptosis-related genes as well as other mild-stress-response genes; however, DNA-repair-related genes were not up-regulated (Ogura et al. 2009). Somewhat higher doses appear to be needed for up-regulation of DNA repair genes (Rothkamm and Löbrich 2003). Rather than reliance only on DNA repair for tissue protection, damaged cells may be removed via selective apoptosis as a mild-stress response when signaled to divide (Rothkamm and Löbrich 2003). These adaptive responses are probably regulated epigenetically. If so, each likely represents an epicellcom process (Scott 2011a).
Low-dose radiation prevents harm from high doses
Many studies have demonstrated that a low radiation dose can prevent harm from subsequent high radiation dose. This type of radiation adaptive response has been called radiation conditioning hormesis (Calabrese et al. 2007). Miura (2004) reviewed the extensive literature related to studies demonstrating that a small radiation dose protected from harm from a subsequent large radiation dose. The author put forth the evolutionary perspective that organisms living in an aerobic environment in order to survive were forced to evolve effective cellular strategies to detoxify reactive oxygen species. In addition to diverse antioxidant enzymes and compounds, DNA repair enzymes, and disassembly systems, which eliminate damaged proteins, regulatory systems that control transcription, translation, and activation have also developed. According to Miura (2004), the adaptive responses, especially those to radiation, are defensive regulation mechanisms by which oxidative stress (e.g., from a conditioning low radiation dose) elicits a response against damage from a subsequent severe stress (e.g., challenging high radiation dose). The activation of DNA damage repair pathways is known to be an important component of the radiation adaptive response (Wolf 1992).
A novel adaptive response study design was used by Day et al. (2007). Using a sensitive in vivo chromosomal inversion assay (pKZ1 mouse prostate model), they demonstrated for the first time an adaptive response when a low X-ray dose (0.01–1 mGy) was given several hours after a high dose (1,000 mGy). This type of adaptive response has been called radiation post exposure conditioning hormesis (Calabrese et al. 2007). The adaptive responses in this study were of similar magnitude to the adaptive responses previously observed in the indicated test system when the low dose was given first. A chromosomal inversion adaptive response was also observed when a 1-mGy X-ray dose was preceded or followed by a dose of 0.01 mGy. This was the first example of an adaptive response when both doses are low. The authors stated that their data agree with previous reports of an on-off mechanism of adaptive response. They concluded that the induction of an adaptive response by a low dose after a high damaging dose provides evidence that the mechanisms underlying radiation adaptive responses are not due to prevention of damage induced by the high dose but to modulation of the cellular response to this damage.
Low-dose radiation stimulates selective removal of precancerous and other aberrant cells via intercellular signaling
The ability of a precancerous cell to escape natural anticancer signals imposed on them by neighboring cells and the microenvironment is an important stage in tumorigenesis (Portess et al. 2007). Portess et al. characterized a system of intercellular induction of apoptosis whereby nontransformed cells selectively remove neoplastically transformed cells from co-culture via cytokine, ROS and RNS signaling. This has been called a protective apoptosis mediated (PAM) process (Scott 2008a,b). Portess et al. (2007) demonstrated that irradiation of nontransformed cells with low doses of either high-LET alpha particles or low-LET gamma rays led to stimulation of intercellular induction of apoptosis (i.e., the PAM process). Using scavengers and inhibitors they confirmed the involvement of ROS/RNS signaling and the importance of transformed cell secreted NADPH oxidase in the selectivity of the system against transformed cells. Absorbed radiation doses as low as 2 mGy of gamma rays and 0.29 mGy of alpha radiation were sufficient to produce an observable increase in transformed cell apoptosis. However, this adaptive response process saturated at very low doses (50 mGy for gamma rays and 25 mGy for alpha radiation) under the exposure scenarios employed. By applying a neutralizing antibody assay, the researchers also confirmed an important role for transforming growth factor β (TGF-β) in the radiation-induced intercellular signaling. They also indicated that the protective system may represent natural anticancer mechanisms stimulated by extremely low doses of ionizing radiation (i.e., very mild stress).
Tremme and Bauer (2013) more recently studied signaling between irradiated transformed (or tumor) and unirradiated nontransformed cells using a co-culture system involving both cells types, with a focus on the PAM process. They demonstrated that low-dose gamma rays substantially increased superoxide anion production in oncogenically transformed cells and tumor cells but not nontransformed cells. The enhancement was radiation-dose-independent for doses in the range 20 − 200 mGy, consistent with characteristics of an epicellcom process. The transfer of a few irradiated transformed cells to nonirradiated control cultures (bystander study) was sufficient for transmission of a signal to induced superoxide anion production in the nonirradiated cells. SiRNA-related knockdown and reconstitution experiments demonstrated that TGF-β1 was involved in the protective bystander effect triggered by low-dose gamma rays in this experimental system. A kinetic analysis by Tremme and Bauer (2013) revealed that the enhanced superoxide anion production was substantially reduced before the release of bystander signaling via activated TGF-β.
Other researchers demonstrated that low doses of low-LET photon radiation can lead to a reduction in the neoplastic transformation frequency to below the spontaneous level (Azzam et al. 1996; Redpath and Antoniono 1998, 2001, 2003) while high doses lead to elevated transformation frequencies that increase as the dose increases further (i.e., hormetic responses). The reduction in the spontaneous frequency may relate to intercellular signaling between transformed and non-transformed cells leading to selective removal of the transformed cells, but this has not been investigated by the indicated researchers.
Redpath’s group also studied the importance of dose rate in connection with radiation ANP against neoplastic transformation. A dose rate threshold (approximately 1 mGy/day) was revealed for ANP (Elmore et al. 2008).
Low-dose radiation suppresses inflammation and thereby protects from inflammatory diseases
A multitude of experimental evidence has accumulated showing that low radiation doses functionally modulates a variety of inflammatory processes (Rödel et al. 2012). The indicated modulations include hampered leukocyte adhesion to endothelial cells, a reduced activity of the inducible nitric oxide synthase, and lowered oxidative burst in macrophages (Rödel et al. 2012).
Cigarette smoke contains the chemical benzo[a]pyrene (BaP) that when metabolized in the body produces the inflammation-promoting carcinogen BaP diol epoxide (BPDE) which has been demonstrated to induced lung tumors in animal models. Further, cigarette smoking is known to cause inflammation and related lung cancer in humans, which has been implicated to be suppressed by low-level exposure to radon in the home (Thompson et al. 2008; Scott 2011b). Because BPDE modifies the microenvironment (e.g., stromal cells) of potential-cancer-causing lung epithelial cells (if undergoing neoplastic transformation), Chen et al. (2012) investigated whether low-dose-gamma rays could modulate the in vitro response of stromal cells to BPDE. The strategy employed was based on neoplastic transformation of human bronchial epithelial cells (HBEC) being an important step in the lung carcinogenesis process. The researchers employed a cell-culture-media-transfer approach and revealed that BPDE induces secretion of the pro-inflammatory cytokines interleukin-6 (IL-6), CLCX1 and CLCX5 from human lung fibroblast, and more importantly, a single low dose (90 mGy) of gamma rays inhibited the secretion (a novel hormetic phenotype).
Chen et al. (2012) also investigated the mechanism by which IL-6 secretion by fibroblasts promotes human bronchial epithelial cell (HBEC) transformation. Condition media from cigarette-smoke-carcinogen-treated fibroblast (cell line HFL1) strongly induced phosphorylation of STAT3 in an IL-6-dependent manner in HBEC. Direct application of IL-6 markedly potentiated BPDE-induced HBEC transformation, supporting the finding that IL-6 secretion from fibroblasts facilitates HBEC transformation. The finding that low-dose gamma rays suppresses fibroblast-derived, IL-6-mediated transformation is supportive of complementary observations of Vicent et al. (2012) discussed below.
Vicent et al. (2012) conducted gene expression analysis comparing normal mouse lung fibroblast and cancer-associated fibroblasts (CAF) from mice. The researchers identified a set of genes (called secreted genes [relates to their associated proteins]) that correlate with the CAF phenotype. The indicated set of secreted genes is called a “gene signature”. The gene signature for CAFs was found to be an independent marker of poor survival in patients with non-small-cell lung cancer. Genes comprising the gene signature were up-regulated in normal lung fibroblast after they were exposed to tumor cells for a prolonged period, implicating that lung fibroblast can be influenced by bystander tumor cells leading to taking on a CAF-like phenotype. Functional studies revealed important roles for IL-6 → IL-6R and CLF1 → CNTFR signaling in promoting non small cell lung cancer. Based on the work of Chen et al. (2012), low-dose gamma rays would be expected to suppress IL-6 → IL-6R signaling and may also suppress CLF1 → CNTFR signaling.
Nakatsukasa et al. (2008) studied the effect of low-dose gamma rays on collagen-induced arthritis (CIA) in DBA/IJ mice. The mice were immunized with inflammation-promoting type II collagen and exposed to low-dose gamma rays (500 mGy per week for 5 weeks). Paw swelling, redness, and bone degradation were suppressed by irradiation, which also delayed the onset of pathological changes and reduced severity of arthritis. The production of tumor necrosis factor-alpha (TNF-α), interferon-gamma (IFN-γ), and IL-6, which have important roles in the onset of CIA, was suppressed by low-dose gamma irradiation. The level of anti-type II collagen antibody, which is essential for the onset of CIA, was also lower in irradiated CIA mice. The population of plasma cells was increased in the CIA mice; however, low-dose irradiation blocked the increase. Because regulatory T cells (Tregs) are known to be involved in suppression of autoimmune disease, the population of CD4+CD25+Foxp3+ Tregs was evaluated by the researchers. Interestingly, a significant increase in Tregs was observed in the irradiated CIA mice. Overall, the data points to low-dose gamma rays attenuating CIA through suppression of pro-inflammatory cytokines and autoantibody production, and induction of Tregs. Based on a follow-up study with a similar design, the researchers concluded that the results obtain suggest that a major mechanism of attenuation of the pathology of collagen-induced arthritis by repeated 500 mGy gamma irradiation is up-regulation of Treg cells concomitantly with suppression of IL-6 and IL-17 production (Nakatsukasa et al. 2010 ).
Low-dose radiation stimulates anticancer and other immunity
Sakai et al. (2003) using a mouse model for skin-tumor induction by injected chemical carcinogen (0.5 mg of 20-methylcholanthrene [MC] in olive oil), examined the reduction in MC-induced skin tumors as a result of chronic exposure to Cs-137 gamma rays at different dose rates (0.3, 0.95, or 2.6 mGy/h). Thirty-five days after the start of irradiation, mice were injected via the groin with MC and radiation exposure was then continued at the same dose rate as before the injection. Cumulative tumor incidences after 216 days following MC injection were 94 % in mice irradiated at 0.3 mGy/h, 76 % at 0.95 mGy/h, 89 % at 2.6 mGy/h, and 94 % in non-irradiated control mice. Only the result (76 %) for the 0.95 mGy/h group was significantly below (p <0.05) the controls. The protection afforded by the chronic low-dose-rate exposure to low-LET radiation was attributed to a hierarchy of adaptive response mechanisms that include antioxidant capacity, repair of DNA damage, removal of neoplastically transformed cells via apoptosis, and removal of proliferating cancer cells by the immune system. Indeed, Sakai and his colleagues were one of the first groups to recognize the hierarchical nature of radiation adaptation.
Internal exposure to high-level cigarette smoke carcinogen benzo[a]pyrene (BaP) causes inflammation and in mouse models has been demonstrated to lead to multiple lung tumors per animal. Using chemopreventative agents, researcher have successfully protected from BaP-exposure related lung cancers by using select agents that reduce the dose of ultimate carcinogen (e.g., BPDE) that arises in the body via metabolism of BaP. However, such studies do not relate to boosting the body’s natural defenses against cancer. Given that low-dose radiation suppresses cancer-facilitating inflammation, one might expect that low-dose radiation may reduce the number of lung tumors in mice exposed to high-level BaP, provided anti-inflammatory genes are not epigenetically silenced via the high level BaP exposure. Bruce et al. (2012) examined the effects of injected BaP alone or in combination with fractionated low-dose gamma radiation (60 – 600 mGy) on the induction of lung adenomas in A/J mice. The results obtained showed that 600 mGy to the total body delivered in six biweekly fractions of 100 mGy starting 1 month after BaP injection significantly reduced the number of lung tumors (adenomas) induced per animal. The 60 mGy group (10 mGy fractions) did not reveal any radiation protection against BaP-induced lung adenomas, which suggests that DNA double-strand-break repair (which should be induced by both 10 and 100 mGy fractions [Rothkamm and Löbrich 2003]) may not explain the protection observed for the 600 mGy group. Suppression of inflammation and stimulation of anticancer immunity appear more plausible explanations of the protection afforded by the six 100 mGy fractions. The data of Bruce et al. (2012) also indicated that the six biweekly doses of 100 mGy suppressed the occurrence of spontaneous hyperplastic foci in the lung, although this suppression failed to reach statistical significance when analyzed based on average foci per lung, possibly related to the small sample sizes used for the control and test groups.
Kojima et al. (2004) examined whether the increase of glutathione level induced by low-dose gamma rays is involved in the appearance of enhanced natural killer (NK) activity and antibody-dependent cellular cytotoxicity (ADCC), leading to a suppression of tumor growth in Ehrlich solid tumor-bearing mice. NK cell activity in ICR mouse splenocytes significantly increased from 4 to 6 h after whole-body exposure to 500 mGy and thereafter decreased almost to the background level by 24 h post exposure. ADCC also had a similar pattern over time. Reduced glutathione exogenously added to splenocytes in culture obtained from normal mice enhanced both NK activity and ADCC in a dose-dependent manner. Tumor growth was also examined in tumor-bearing mice. Tumor growth after inoculation was significantly delayed by low-dose irradiation. The results suggested that low-dose gamma rays activate immune functions via an induction of glutathione, leading to a delay of tumor growth.
Studies by Hayase et al. (2013) investigate the influence of repeated 500-mGy gamma-ray dose on the Th1/Th2 immunity balance in Ehrlich-Solid-Tumor-bearing mice. The repeated doses significantly delayed the growth of tumors. The cytotoxic activities of natural killer cells and cytotoxic T lymphocytes were enhanced after repeated irradiations. The irradiation increased the production of IFN-γ by splenocytes of tumor-bearing mice with there Interleukin (IL-4) unchanged, resulting in an increased IFN-γ/IL-4 ratio, a hallmark of a Th1 shift. Irradiation also increased IL-12 production and levels of reduced-glutathione in macrophages.
Cheda et al. (2004a,b) demonstrated that single exposure of mice to 100 or 200 mGy of X rays led to significant inhibition of the development of artificial tumor metastases (from injected tumor cells) in the lungs and that the effect was related in part to the enhanced activity of natural killer cells. In another study by the same group, they demonstrated that inhibition of the tumor metastases by single exposure of mice to 100 or 200 mGy of X rays results, to a large extent, from stimulation of the cytocidal activity of macrophages, which secrete enhanced amount of nitric oxide (Nowosielska et al. 2006).
Takahashi and Kojima (2006) examined the effect of ingested radon (222Rn, t1/2 = 3.82 days, alpha particle energy =5.49 MeV) in disease suppression using two experimental disease mouse models (radon concentrations in drinking water at the time of use varied over a wide range). Model 1: Ingestion exposure of five-week-old SPF NC/Nga mice to radon significantly delayed the progression of atopic dermatitis-like skin lesions induced by picrylchloride (2,4,6-trinitrochlorobenzene). Model 2: The number of pulmonary metastatic foci in six-week-old male C57BL/6 mice inoculated with B16 melanoma cells 2 weeks after start of radon ingestion was reduced significantly by the ingested radon. In addition, the IFNγ/IL-4 ratio in splenocytes from BALB/c mice immunized with DNP-Ascaris was significantly increased by ingested water containing radon. These results were interpreted by Takahashi and Kojima (2006) to indicate beneficial modulation of the immune system by the ingested radon.
Thompson et al. (2008) in an epidemiological study of lung cancer in association with residential radon exposure revealed a reduction in lung cancer cases for persons living in homes with radon levels near and at the U.S. Environmental Protection Agency’s action level of 4 pCi/L. Further, the dose–response relationship was found to be hormetic, supporting the view that low-level radiation stimulates the body’s natural defenses and protects from cancer induction by other agents (e.g., cigarette smoke carcinogens). In an analysis of the data of Thompson et al. (2008) by Scott (2011b) using a hormetic relative risk (HRR) model that allows assessing the percentage of the population that benefits from radiation ANP, it was found that everyone may benefit from radiation ANP for living in homes with radon levels near to (below and above) and at the EPA’s action level. In addition, the analysis of the data revealed that eliminating radon from the home in such cases could actually lead to an increase in the risk of lung cancer due to a loss of radiation ANP against cancer.
With the HRR model, the threshold dose for radiation hormesis varies for different individuals (Scott 2011a, b). Similarly, the higher threshold dose that suppresses adaptive-response mechanisms also varies between different individuals. The distributions of the beneficial low-dose threshold and harmful high-dose threshold were the basis of the J-shape, dose–response curve for lung cancer relative risk. Variable thresholds are also expected for other cancer types, with the threshold distributions depending on the individual and cancer type. Variable thresholds are also expected to apply to other diseases. Whether doses below the minimum hormetic dose are harmful or not is unknown.
Low-dose-radiation exposure during gestation causes lasting protective epigenetic changes in male offspring
To determine if deleterious or protective epigenetic changes occur when exposed to low-dose radiation doses during gestation, Bernal et al. (2013) utilized the viable yellow agouti (A vy ) mouse model (Duhl et al. 1994 ). This mouse strain is very sensitive to environmental agents that alter the fetal epigenome. Variable expression of the A vy metastable epiallele is regulated by epigenetic modifications, such as cytosine-phosphate-guanine (CpG) site methylation and histone marks that are established early during development in and around the cryptic promoter in a transgene upstream of the Agouti gene (Bernal et al. 2013). A transgene is an exogenous gene that is introduced into an organism so that it will have a new characteristic that can be transmitted to offspring. Metastable epialleles are variably expressed in genetically identical individuals because of epigenetic modifications of genes that occur during early development. Hypomethylation of the alternative promoter leads to inappropriate Agouti gene expression in all tissues in Avy mice (Bernal et al. 2013). This leads to a yellow coat color (morbidity-promoting phenotype) and also antagonizes the melanocortin 4 receptor (MC4R) in the hypothalamus, which causes the animals to become obese and to develop cancer and diabetes at high frequencies. Exposure to low-dose radiation (14 to 30 mGy) led to protective epigenetic changes (coat color shifted from yellow towards brown [p <0.01]) in a sex-specific manner, with male offspring benefiting (reduced risks for obesity, cancer, and diabetes) more than female offspring. The protective changes were inhibited by antioxidants, indicating that the adaptive protection induced by mild radiation stress during gestation is mediated at least in part via ROS.
More data are accumulating from experimental animal studies showing that mild stresses at a young age can protect via epigenetic reprogramming from damage from severe stresses later in life as well as slow the rate of ageing and increase lifespan. Single or multiple exposures to low doses of otherwise harmful agents (e.g., radiation, heat stress, ROS) can cause a variety of anti-ageing and lifespan-extending effects that appear to relate to epigenetic reprogramming of genes as an adaptive response (Vaiserman 2011). The early-in-life-induced epigenetic reprogramming is not limited to specific genes or genomic regions but rather is consistent with a genome-wide epigenomic response (Vaiserman 2011).
Low-dose radiation slows ageing and prolongs life
Nomura et al. (2013) evaluated the effect of continuous exposure to low-dose-rate gamma rays on lifespan of mice with accelerated ageing caused by mutation of the klotho gene. While control mice did not survive more than 80 days, more than 10 % of mice exposed continuously to 0.35 or 0.7 mGy/h of gamma rays from 40 days after birth survived more than 80 days. Four percent of the irradiated mice survived for more than 100 days. Low rate gamma-ray exposure significantly increased plasma calcium concentration in the mutant mice, and concomitantly increased hepatic catalase activity. The researchers concluded that their data suggest that low-dose-rate ionizing radiation can prolong the lifespan of mice in certain settings.
Low-dose radiation stimulates beneficial Wnt/β → catenin signaling, neural stem cell proliferation, and nerve cell production in the hippocampus
Nerve cell production (neurogenesis) in the hippocampus is actively involved in neural circuit plasticity and learning of mammals; however, learning capacity can decrease significantly with ageing and with the aging-related neurodegenerative disorder Alzheimer’s disease. Increasing evidence points to Wnt/β → catenin signaling as being critical in control of proliferation and differentiation of neural stem cells or progenitors in the hippocampus. Wei et al. (2012) studied the influence of low-dose (300 mGy) X rays on neurogenesis in the hippocampus of mice. Low-dose X rays induced significant increases in Wnt1, Wnt3a, Wnt5a, and β-catenin expression in both neural stem cells and in situ hippocampus. Low-dose radiation also enhanced neurogenesis in hippocampus. In addition, it promoted cell survival and reduced apoptotic death of neuronal stem cells. In contrast, a high dose of 3,000 mGy caused detrimental effects on signaling and neurogenesis (i.e., hormetic dose–response relationship). The research findings were interpreted by Wei et al (2012) to reveal beneficial effects of low-dose radiation related to stimulating neural stem cell prolifereation, nerogenesis of hippocampus, and animal learning, possibly via stimulating Wnt/β → catenin signaling cascades, suggesting its translational application role in devising new therapy for aging-related neurodegenerative disorders, especially Alzheimer’s disease.
Moderate radiation doses reduce asthma severity
Mitsunobu et al. (2003) examined in a pilot study the bronchial asthma mitigating effect of combined radon and thermal therapy of human subjects. The therapy is based on the knowledge that increased systemic production of ROS by activated inflammatory cells have a pathophysiological role in bronchial asthma that might be mitigated via increased antioxidants stimulated by radiation exposure (from inhaled radon). The aim of the study was to evaluate the clinical efficacy of combined radon and thermal therapy on asthma in relation to antioxidant enzymes and lipid peroxide. The combined therapy was performed once a week. All subjects went to a hot bathroom with a high concentration of radon and nasal inhalation of radon from a hot spring was performed for 40 min under conditions of high humidity. The room temperature was 48° Celsius and the room average radon concentration was 2,080 Bq/cubic meter. On day 28 the catalase and superoxide dismutase activities were significantly increased compared to controls while the lipid peroxide level was significantly decreased. In addition, the forced expiratory volume in one second was significantly increased at the indicated follow-up time. The researchers concluded that their pilot study demonstrated that combined radon + thermal therapy improved the pulmonary function of asthmatics by increasing antioxidant enzymes in the respiratory tract.
Park et al. (2013) investigated whether low-dose radiation exacerbates allergic asthma responses in C57BL/6 mice sensitized and challenged with ovalbumin (OVA) to induce asthma. The mice were irradiated (whole-body) once daily for 3 consecutive days with a 667 mGy dose of gamma rays given 24 h before each OVA challenge. Repeated gamma irradiation reduced OVA-specific IgE levels, the number of inflammatory cells (including mast cells), goblet cell hyperplasia, collagen deposition, airway hyper-responsiveness, expression of inflammatory cytokines, the chmokine system CC ligand 2 (CCL2) with its receptor CC receptor 2 (CCR2), as well as nuclear factor kappa B (NF-κB) and activator protein-1 activities. These factors were increased in broncheoalveolar lavage cells and lung tissue of OVA-challenged mice. The data obtained was interpreted to suggest that low radiation doses enhance Foxp3+ and IL-10-producing Tregs, which reduce OVA-induced allergic airway inflammation and tissue remodeling through the down-regulation of migration by the CCL2/CCR2 chemokine axis and activation of mast cells via OX40/OX40L in lung tissue of OVA-challenged mice.
Discussion
It is now recognized based on research as early as 1975 that low radiation doses are not harmful and may invoke hormetic phenotypes via stimulating the body’s natural defenses (Ducoff 1975; Kondo 1993; Hashimoto et al. 1999; Mitchel et al. 2003; Sakai et al. 2003; Nowosielska et al. 2006, 2010; Pollycove 2006; Sakai 2006; Lacoste-Collin et al. 2007; Jaworowski 2008; Pollycove and Feinendegen 2008; Rithidech and Scott 2008; Sanders and Scott 2008; Scott et al. 2009; Cuttler 2010; Jaworowski 2010a,b; Sanders 2010; Vaiserman 2011; Nomura et al. 2013). Components of the indicated low-dose-radiation stimulated protective effects have been observed in a wide variety of organisms, ranging from simple prokaryotes to higher eukaryotes suggesting that the phenomena are evolutionary conserved (Makinodan and James 1990). Depending on the radiation dose rate and radiation dose level, components of the hierarchy of protective mechanisms (increased antioxidants, selective apoptosis of aberrant cells, DNA damage repair in cells with repairable injury, apoptosis of severely damage cells, inflammation suppression, and enhanced anticancer immunity) are differentially activated by low dose radiation and can suppress cancer induction by high-level-cigarette-smoke and other chemical carcinogens, may lead to life lasting beneficial epigenetic changes when exposed during gestation, can reduce the severity of asthma, can relieve pain from arthritis, may reduce the severity of diabetes, may aid in treating Alzheimer’s disease, and could slow the rate of ageing. Variable thresholds for radiation hormesis are expected to apply for different individuals and for different endpoints. For the same individual and for cancer prevention, variable hormesis thresholds (radiation doses) are also expected for different cancer types.
Unfortunately the potential enormous benefits of low-dose radiation that were discussed here are largely unknown to the majority of the general public who has been mislead by influential organizations and individuals to believe that any amount of radiation is harmful, no matter how small.
It is clear that low-dose-radiation ANP is responsible for the radiation hormesis phenotype for stochastic effects such as mutations, neoplastic transformation, and cancer. The hierarchical ANP system is at least in part epiregulated, e.g., via microRNAs (miRNAs) and other means (Vaiserman 2011; Bernal et al. 2013). Emerging data suggest that mild stress (e.g., low-level radiation) can rapidly alter the biogenesis of miRNAs, the expression of mRNA, and the activities of miRNA-protein complexes (Leung and Sharp 2010). As a sensor of environmental and other stresses, the tumor suppressor gene p53 is activated by DNA damage. It also regulates the expression of specific miRNAs at the transcription and processing levels, which can lead to activation of adaptive-response genes that differentially control the hierarchal system of radiation ANP.
Components of the presumed epiregulated system of natural protection can be activated by gamma-ray doses as low as 0.5 mGy above natural background as was demonstrated by Ogura et al. (2009) for suppression of spontaneous mutations and the activation likely involves rapid epireprogramming of the genome (Scott et al. 2009). This allows for rapidly mounting DNA damage repair, antioxidant production, protective cytokine (e.g., low-level TGF-β1) releases, etc. At a gamma-ray dose of 0.5 mGy, the small number of spontaneous mutant cells in the target population is unlikely to be directly destroyed by radiation tracks (e.g., ionization election tracks after gamma- or X-ray exposure). Thus, the adaptive response (hormetic phenotype) observed by Ogura et al. (2009) is likely an epiregulated protective bystander effect implemented as an epicellcom process.
Radiation doses much higher than current natural background radiation levels promote epigenetically silencing (episilencing [Scott et al. 2009]) of adaptive-response genes and can significantly suppress the immune system (Liu 2007), facilitating development of cancer and other diseases. As might be expected, protective epigenetic changes appear to be orders of magnitude more likely after low radiation doses than an induced mutation of a given type in a given gene (Scott 2013 ). Low-dose radiation epiactivation of adaptive response genes and their episilencing by high radiation doses provides a basis for hormetic (U- or J-shaped) dose–response relationships for mutation induction, neoplastic transformation and cancer.
The natural protection system appears to be more efficiently (i.e., over a wider dose range) activated by low-LET radiation than high-LET radiation and this may relate to a link between the microdose (dose to very small targets like the cell nuclei) spatial and temporal distributions and the intensity of the protective intercellular signaling associated with ANP (Feinendegen et al. 2007b). More cells are hit when low-LET radiation is used than for high-LET radiation, for a given macroscopic radiation dose (i.e., absorbed dose representing energy deposition per unit macroscopic mass), because the low-LET radiation dose is more uniformly distributed.
The benefit of radiation ANP for children is less clear than for adults (Scott et al. 2008). There is however evidence for chronic low-rate exposure to gamma rays protecting children from sporadic cancer (Chen et al. 2007).
Life lasting beneficial epiregulated hormetic phenotypes (less obesity, fewer cancers, fewer diabetics) brought on by exposure to low radiation doses during gestation were demonstrated by Bernal et al. (2013) for offspring using the very sensitive viable yellow agouti (A vy) mouse model. Male offspring benefited more than did female. Whether the same benefits occur in humans is unknown, but could be investigated by comparing populations residing in high natural background radiation areas with those residing in low natural background radiation areas.
Previous ecological studies that compared disease prevalence in regions of high natural background radiation regions with that for low natural background radiation regions have demonstrated associations between disease reduction and increasing background radiation levels (Luckey 1991; Mifune et al. 1992; Sawat 2000; Kojima et al. 2002; Hart 2010, 2011a, b; Fornalski and Dobrzyński 2012).
Regarding the future, there is great potential for low-dose radiation therapy for certain diseases and for use of low-dose radiation in disease prevention (e.g., for heavy smokers) and for improved treatment of intense pain associated with inflammatory diseases. This includes debilitating diseases such as cancer, arthritis, diabetes, and Alzheimer’s. Hopefully, these benefits will be soon achieved and become commonplace. A worldwide research initiative focusing on radiation hormesis-phenotype-related mechanisms and implications for disease therapy and prevention could lead to huge dividends for all. A major current obstacle is the invalid LNT hypothesis whose use by regulatory agencies should be abandoned because of the enormous harm it has already caused (Orient 2014). Another obstacle is the lack of research support for low-dose radiation studies in the United States.
The discussion is ended with the following LNT hypothesis-related quote from 20 years ago:
“Existing data obtained with beams of electrons, protons, X ray photons, incorporated tritium and 125I, demonstrate that hundreds of electrons may traverse a cell for inactivation and millions may be required for cancer induction. If the linear extrapolation were valid these numbers would be reduced to one. The contradictions suggest serious reconsideration of accepted radiation protection standards.” Katz and Waligorski (1994)
Abbreviations
- ADCC :
-
antibody-dependent cellular cytotoxicity
- ANP :
-
activated natural protection
- B[a]P :
-
benzo[a]pyrene
- BPDE :
-
benzo[a]pyrene diolepoxide
- CAF :
-
cancer-associated fibroblasts
- CCL2 :
-
chmokine system CC ligand 2
- CCR2 :
-
receptor CC 2
- CIA :
-
collagen-induced arthritis
- CLF1 :
-
cytokine-like factor 1
- CNTFR :
-
ciliary neurotropic factor receptor
- DNA :
-
deoxyribonucleic acid
- epiactivation :
-
epigenetic activation
- epiregulated :
-
epigenetically regulated
- epicellcom :
-
epigenetically regulated cell community wide
- epireprogramming :
-
epigenetic reprogramming
- episilencing :
-
epigenetically silencing
- γ-GCS :
-
γ-glutamylcysteine synthetase
- GPx :
-
glutathione peroxidase
- GR :
-
glutathione reductase
- GSH :
-
reduced glutathione
- HBEC :
-
human bronchial epithelial cells
- HRR :
-
hormetic relative risk
- IFN-γ :
-
interferon-gamma
- IL-6 :
-
interleukin-6
- IL-6R :
-
interleukin-6 receptor
- p53 :
-
tumor protein
- LET :
-
linear energy transfer
- MC :
-
methylcholanthrene
- MC4R :
-
melanocortin 4 receptor
- miRNA :
-
microRNA
- NK :
-
natural killer
- PAM :
-
protective-apoptosis-mediated
- OVA :
-
ovalbumin
- RNS :
-
reactive nitrogen species
- ROS :
-
reactive oxygen species
- SiRNA :
-
small interfering RNA
- SOD :
-
superoxide dismutase
- SPF :
-
specific pathogen free
- STAT 3 :
-
signal transducer and activator of transcription 3
- TGF-β :
-
transforming growth factor β
- TNF-α :
-
tumor necrosis factor alpha
- Tregs :
-
regulatory T cells
- TRX :
-
thioredoxin
References
Averbeck D (2009) Does scientific evidence support a change from the LNT model for low-dose radiation risk extrapolation? Health Phys 95:493–504
Azzam EI, de Toledo SM, Raaphorst GP, Mitchel RE (1996) Low-dose ionizing radiation decreases the frequency of neoplastic transformation to a level below the spontaneous rate in C3H 10T1/2 cells. Radiat Res 146:369–373
Bauer G (2007) Low dose radiation and intercellular induction of apoptosis: potential implications for control of oncogenesis. Int J Radiat Biol 83:873–888
Bernal AJ, Dolinoy DC, Huang D, Skaar DA, Weinhouse C, Jirtle RL (2013) Adaptive radiation-induced epigenetic alterations mitigated by antioxidants. FASEB J 27(2):665–671
Bhatia AL (2008) Radiation exposure and evolutionary perspectives with special reference to neutral theory. Int J Low Radiat 5(2):113–122
Bruce V, Belinsky SA, Gott K, Liu Y, March T, Scott B, Wilder J (2012) Low-dose gamma-radiation inhibits benzo[a]pyrene-induced lung adenoma development in A/J mice. Dose Response 10:516–526
Calabrese EJ, Baldwin LA (1999) Reevaluation of the fundamental dose–response relationship. A new database suggests that the u-shaped, rather than the sigmoidal, curve predominates. BioSci 49(9):725–732
Calabrese EJ, Baldwin LA (2003) The hormetic dose–response model is more common than the threshold model in toxicology. Toxicol Sci 71:246–250
Calabrese EJ, Calabrese V (2013) Reduction of arthritic symptoms by low-dose radiation therapy (LD-RT) is associated with an anti-inflammatory phenotype. Int J Radiat Biol 89:278–l286
Calabrese EJ, Dhawan G (2014) Historical use of x-rays: treatment of inner ear infections and prevention of deafness. Hum Exp Toxicol 33(5):542–553
Calabrese EJ, Bachmann KA, Bailer AJ et al (2007) Biological stress response terminology: Integrating the concepts of adaptive response ad preconditioning stress within a hormetic dose–response framework. Toxicol Appl Pharmacol 222:122–128
Cheda A, Wrembel-Wargocka J, Lisiak E, Nowosielska EM, Marciniak M, Janiak MK (2004a) Single low does of X-rays inhibit development of experimental tumor metastases and trigger the activities of NK cells in mice. Radiat Res 161:335–340
Cheda A, Wrembel-Wargocka J, Lisiak E, Marciniak M, Nowosielska EM, Janiak MK (2004b) Inhibition of the development of pulmonary tumor nodules and stimulation of the activity of NK cells and macrophages in mice by single low doses of low-LET radiation. Int J Low Radiat 1:171–179
Chen WL, Luan YC, Shieh MC, Chen ST, Kung HT, Soong KL, Yeh YC, Chou TS, Mong SH, Wu JT, Sun CP, Deng WP, Wu MF, Shen ML (2007) Effects of cobalt-60 exposure on health of Taiwan residents suggest new approach needed in radiation protection. Dose Response 5:63–75
Chen W, Xu X, Bai L, Padilla MT, Gott KM, Leng S, Tellez CS, Wilder JA, Belinsky SA, Scott BR, Lin Y (2012) Low dose gamma-irradiation inhibits IL-6 secretion from human lung fibroblasts that promotes bronchial epithelial cell transformation by cigarette-smoke carcinogens. Carcinogenesis 33(7):1368–1374
Cohen BL (2008) The linear no-threshold theory of radiation carcinogenesis should be rejected. J Am Phys Surg 13(3):70–76
Cuttler JM (2010) Commentary on using LNT for radiation protection and risk assessment. Dose Response 8(3):378–383
Day TK, Zeng G, Hooker AM, Bhat M, Scott BR, Turner DR, Sykes PJ (2007) Adaptive response for chromosomal inversions in pKZ1 mouse prostate induced by low doses of X radaition delivered after a high dose. Radiat Res 167(6):682–692
Ducoff HS (1975) Forms of increased longevity of Tribolium after X-irradiation. Exp Gerontol 10:189–193
Ducoff HS (2002) Radiation hormesis: incredible or inevitable? Korean J Biol Sci 6:187–193
Duhl DM, Vrieling H, Miller KA, Wolff GL, Barsh GS (1994) Neomorphic agouti mutations in obese yellow mice. Nat Genet 8:6–11
Elmore E, Lao XY, Kapadia R, Giedzinski E, Limoli C, Redpath JL (2008) Low doses of very low-dose-rate low-LET radiation suppress radiation-induced neoplastic transformation in vitro and induce an adaptive response. Radiat Res 169(3):311–318
Feinendegen LE, Paretzke HG, Neumann RD (2007a) Damage propagation in complex biological systems following exposure to low doses of ionizing radiation. Atoms Peace 1:336–354
Feinendegen LE, Pollycove M, Neumann RD (2007b) Whole-body responses to low-level radiation exposure: new concepts in mammalian radiobiology. Exp Hematol 35(Suppl 1):37–46
Fornalski KW, Dobrzyński L (2012) The cancer mortality rate in high natural radiation areas in Poland. Dose Response 10(4):541–562
Hart J (2010) Mean cancer mortality rates in low versus high elevation counties in Texas. Dose Response 8(4):448–455
Hart J (2011a) Cancer mortality for a single race in low versus high elevation counties in the U.S. Dose response 9(3):348–355
Hart J (2011b) Lung cancer in Oregon. Dose Response 9(3):410–415
Hashimoto S, Shirato H, Hosokawa M, Nishioka T, Kuramitsu Y, Matsushita K, Kobayashi M, Miyasaka K (1999) The suppression of metastases and the change in host immune response after low-dose total-body irradiation in tumor-bearing rats. Radiat Res 151:717–724
Hayase H, Ohshima Y, Takahashi M, Kojima S (2013) The enhancement of Th1 immunity and the suppression tumor growth by low-dose γ-radiation. Int J of Low Radiat 5:275–298
Jaworowski Z. (1999) Radiation risk and ethics. Phys Today, September:24 − 29
Jaworowski A (2008) The paradigm that failed. Int J Low Radiat 5(2):151–155
Jaworowski Z (2010a) Observations of the Chernobyl disaster and LNT. Dose Response 8(2):148–171
Jaworowski Z (2010b) Radiation hormesis − a remedy for fear. Hum Exp Toxicol 29:263–270
Karam PA, Leslie SA (1999) Calculation of background beta-gamma radiation dose through geologic time. Health Phys 77(6):662–667
Kataoka T (2013) Study of the antioxidative effects and anti-inflammatory effects in mice due to low-dose X-irradiation or radon inhalation. J Radiat Res 54(4):587–596
Katz R, Waligorski MPR (1994) On the linear extrapolation to low doses. Radiat Prot Dosimetry 52(1–4):197–199
Kawakita Y, Ikekita M, Kurozumi R et al (2003) Increase of intracellular glutathione by low-dose gamma-ray irradiation is mediated by transcription factor AP-1 in RAW 264.7 cells. Biol Pharm Bull 36:19–23
Koana T, Okada MO, Ogura K, Tsujimura H, Sakai K (2007) Reduction of background mutations by low-dose X irradiation of Drosophila spermatocytes at a low dose rate. Radiat Res 167:217–221
Kojima S, Matsuki O, Nomura T et al (1998a) Induction of mRNAs for glutathione synthesis-related proteins in mouse liver by low doses of γ-rays. Biochem Biophys Acta 1381:312–318
Kojima S, Matsuki O, Nomura T et al (1998b) Localization of glutathione and induction of glutathione synthesis-related proteins in mouse brain by low doses of γ rays. Brain Res 808:262–269
Kojima S, Ishida H, Takahasji M, Yamaoka K (2002) Elevation of glutathione induced by low-dose gamma rays and its involvement in increased natural killer activity. Radiat Res 157:275–280
Kojima S, Nakayama K, Ishida H (2004) Low dose γ-rays activate immune function via induction of glutathione and delay tumor growth. J Radiat Res 45:33–39
Kondo S (1993) Health effects of low-level radiation. Kinki University Press, Osaka, Japan, Medical Physics Publishing, Madison, WI, USA
Kondo S (1998) Apoptotic repair of genotoxic tissue damage and the role of p53 gene. Mutat Res 402:311–319
Lacoste-Collin L, Jozan S, Cances-Lauwers V, Pipy B, Gasset G, Caratero C, Courtade-Saïdo M (2007) Effect of continuous irradiation with very low doses of gamma rays on life span and the immune system in SJL mice prone to B-cell lymphoma. Radiat Res 168:725–732
Leung AKL, Sharp PA (2010) MicroRNA functions in stress responses. Mol Cell 40:205–215
Liu S-Z (2007) Cancer control related to stimulation of immunity by low-dose radiation. Dose Response 5:39–47
Luckey TD (1980) Hormesis with ionizing radiation. CRC Press, Florida
Luckey TD (1991) Radiation Hormesis. CRC Press, Boca Raton
Makinodan T, James SJ (1990) T cell potentiation by low dose ionizing radiation: possible mechanisms. Health Phys 59(1):29–34
Mifune M, Sobue T, Arimoto H, Komoto Y, Kondo S, Tanooka H (1992) Cancer mortality survey in a spa area (Misasa, Japan) with a high radon background. Jpn J Cancer Res 83:1–5
Mitchel REJ, Jackson JS, Morrison DP, Carlisle SM (2003) Low doses of radiation increase the latency of spontaneous lymphomas and spinal osteosarcomas in cancer prone, radiation sensitive Trp53 heterozygous mice. Radiat Res 159:320–327
Mitsunobu F, Yamaoka K, Hanamoto K, Kojima S, Hosaki Y, Ashida K, Sugita K, Tanizaki Y (2003) Elevation of antioxidant enzymes in the clinical effects of radon and thermal therapy for bronchial asthma. J Radiat Res 44:95–99
Miura Y (2004) Oxidative stress, radiation adaptive responses, and aging. J Radiat Res 45(3):357–372
Muller HJ (1927) Artificial transmutation of the gene. Science 46:84–87
Muller HJ (1954) The manner of production of mutations by radiation. Radiat Biol, vol 1. Chap. 8, McGraw-Hill Book Co. Inc.: 475 − 626
Nakatsukasa H, Tsukimoto M, Ohshima Y, Tago F, Masada A, Kojima S (2008) Suppressing effect of low-dose gamma-ray irradiation on collagen-induced arthritis. J Radiat Res 49:381–389
Nakatsukasa H, Tsukimoto M, Tokuaga A, Kojima S (2010) Repeated gamma irradiation attenuates collagen-induced arthritis via up-regulation of regulator T cells but not by damaging lymphocytes directly. Radiat Res 174:313–324
Nomura T, Sakai K, Ogata H, Magae J (2013) Prolongation of life span in the accelerated aging klotho mouse model, by low-dose-rate continuous γ irradiation. Radiat Res 179:717–724
Nowosielska EM, Wrembel-Wargocka J, Cheda A, Lisiak E, Janiak MK (2006) Enhanced cytotoxic activity of macrophages and suppressed tumor metastases in mice irradiated with low dose x-rays. J Radiat Res 47:229–236
Nowosielska EM, Cheda A, Wrembel-Wargochka J, Janiak MK (2010) Immunological mechanism of the low-dose radiation-induced suppression of cancer metastases in a mouse model. Dose Response 8(2):209–226
Ogura K, Magae J, Kawakami Y, Koana T (2009) Reduction in mutation frequency by very low-dose gamma irradiation of Drosophila melanogaster germ cells. Radiat Res 171:1–8
Orient J (2014) Fukushima and reflections on radiation as a terror weapon. J Am Phys Surg 19(2):48–55
Park BS, Hong GU, Ro JY (2013) Foxp3 + -Treg cells enhanced by repeated low-dose gamma-irradiation attenuate ovalbumin-induced allergic asthma in mice. Radiat Res 179(5):570–583
Pollycove M (2006) Radiobiological basis of low-dose irradiation in prevention and therapy of cancer. Dose Response 5(1):26–38
Pollycove M, Feinendegen L (2008) Low dose radioimmunotherapy for cancer. Hum Exp Toxicol 27(2):169–175
Portess DI, Bauer G, Hill M et al (2007) Low-dose irradiation of nontransformed cells stimulate the selective removal of precancerous cells via intercellular induction of apoptosis. Cancer Res 67:1246–1253
Redpath JL, Antoniono RJ (1998) Induction of an adaptive response against spontaneous neoplastic transformation in vitro by low-dose gamma radiation. Radiat Res 149(5):517–520
Redpath JL, Liang D, Taylor TH, James C, Christie E, Elmore E (2001) The shape of the dose–response curve for radiation-induced neoplastic transformation in vitro: Evidence for an adaptive response against neoplastic transformation at low doses of low-LET radiation. Radiat Res 156:700–707
Redpath JL, Short SC, Woodcock M, Johnston PJ (2003) Low-dose reduction in transformation frequency compared to unirradiated controls: the role of hyper-radiosensitivity to cell death. Radiat Res 159(3):433–436
Rithidech K, Scott BR (2008) Evidence for radiation hormesis after in vitro exposure of human lymphocytes to low doses of ionizing radiation. Dose Response 6:252–271
Rödel F, Frey B, Gaipl U, Keilholz L, Fournier C, Manda K, Scöllnberger H, Hildebrandt G, Rödel C (2012) Modulation of inflammatory immune reactions by low-dose radiation: molecular mechanisms and clinical application. Curr Med Chem 19:1741–1750
Rothkamm K, Löbrich M (2003) Evidence for a lack of DNA double-strand break repair in human cells exposed to very low x-ray doses. Proc Natl Acad Sci U S A 100(9):5057–5062
Sakai K (2006) Enhancement of bio-protective functions by low dose/dose-rate radiation. Dose Response 4:327–332
Sakai K, Hoshi Y, Nomura T, Oda T, Iwasaki T, Fujita K, Yamada T, Tanooka H (2003) Suppression of carcinogenic process in mice by chronic low dose rate gamma-irradiation. Int J Low Radiat 1:142–146
Sanders CL (2010) Radiation hormesis and the linear-no-threshold assumption, 1st edn. Springer, Heidelberg
Sanders CL, Scott BR (2008) Smoking and hormesis as confounding factors in radiation pulmonary carcinogenesis. Dose Response 6:53–79
Sawat A (2000) The immunobiology of low-dose total-body irradiation: more questions than answers. Radiat Res 153:599–604
Scott BR (2004) A biological-based model that links genomic instability, bystander effects, and adaptive response. Mutat Res 568(1):129–143
Scott BR (2005) Stochastic thresholds: a novel explanation of nonlinear dose–response relationships for stochastic radiobiological effects. Dose Response 3(4):547–567
Scott BR (2008a) It’s time for a new low-dose-radiation risk assessment paradigm − one that acknowledges hormesis. Dose Response 6:333–351
Scott BR (2008b) Low-dose risk extrapolation fallacy associated with the linear-no-threshold model. Hum Exp Toxicol 27:163–168
Scott BR (2011a) Modeling DNA double-strand break repair kinetics as an epiregulated cell-community-wide (epicellcom) response to radiation stress. Dose Response 9:579–601
Scott BR (2011b) Residential radon appears to prevent lung cancer. Dose Response 9:444–464
Scott BR (2013) First generation stochastic gene episilencing (STEP1) model and applications to in vitro cancinogen exposure. Dose Response 11:9–28
Scott BR, Sanders CL, Mitchel REJ, Boreham DR (2008) CT scans may reduce rather than increase the risk of cancer. J Am Phys Surg 13:8–11
Scott BR, Belinsky SA, Leng S, Lin L, Wilder JA, Damiani LA (2009) Radiation-stimulated epigenetic reprogramming of adaptive-response genes in the lung: an evolutionary gift for mounting adaptive protection against lung cancer. Dose Response 7:104–131
Takahashi M, Kojima S (2006) Suppression of atopic dermatitis and tumor metastasis in mice by small amounts of radon. Radiat Res 165:337–342
Takatori M, Hattori S, Yagi M (2010) Clinical significance of low-dose radiation therapy: radiation hormesis. Int J Low Rad 7(6):51–519
Thompson RE, Nelson DF, Popkin JH, Popkin Z (2008) Case–control study of lung cancer risk from residential radon exposure in Worchester County, Massachusetts. Health Phys 94(3):228–241
Tremme J, Bauer G (2013) Low-dose gamma irradiation enhances superoxide anion production by nonirradiated cells through TGF-β1-dependent bystander signaling. Radiat Res 179:422–432
Tubiana M (2008) The 2007 Marie Curie prize: the linear no threshold relationship and advances in our understanding of carcinogenesis. Int J Low Radiat 5(3):173–204
Tubiana MF, Feinendegen LE, Yang C, Kaminski JM (2009) The linear no-threshold relationship is inconsistent with radiation biologic and experimental data. Radiology 251:13–22
Vaiserman AM (2011) Hormesis and epigenetics: is there a link? Ageing Res Rev 10:413–421
Vicent S, Sayles LC, Vaka D, Khatri P, Gavaert O, Chen R, Zheng Y, Gillespie AK, Clarke N, Xu Y, Shrager J, Hoang CD, Plevritis S, Butte AJ, Seweet-Cordero EA (2012) Cross-species functional analysis of cancer-associated fibroblasts identifies a critical role for CLCF1 and IL-6 in non-small cell lung cancer in vivo. Cancer Res 72(22):5744–5756
Wei L-C, Ding Y-X, Liu Y-H, Duan L, Bai Y, Shi M, Chen L-W (2012) Low-dose radiation stimulates Wtn/β-catenin signaling, neural stem cell proliferation and neurogenesis of the mouse hippocampus in vitro and in vivo. Curr Alzheimer Res 9:278–289
Wolf S (1992) “Is radiation all bad? The search for adaptation”, the Failla memorial lecture. Radiat Res 131(2):117–123
Wolff S (1998) The adaptive response in radiobiology: evolving insights and implications. Environ Health Perspect 106:277–283
Yamaoka K, Kojima S, Takahashi M et al (1998) Change in glutathione peroxidase synthesis along with that of superoxide dismutase synthesis in mice spleens after low-dose X-ray irradiation. Biochem Biophys Acta 1381:265–270
Acknowledgments
The preparation of this manuscript was supported in part by the Office of Science (BER), U.S. Department of Energy, Grant No. DE-FG02-09ER64783 and in part by Lovelace Respiratory Research Institute. The paper is dedicated to the memory of Professor Howard Ducoff who made numerous important contributions to radiation hormesis research through his work with insects as well as through the work of those he mentored during his life, including the author of this paper. The author (BRS) is a founder member of Scientists for Accurate Radiation Information (SARI; www.radiationeffects.org), a group that includes several authors of radiation hormesis publications. The author confirms independence from the sponsors; the content of the article has not been influenced by the sponsors.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article

Cite this article
Scott, B.R. Radiation-hormesis phenotypes, the related mechanisms and implications for disease prevention and therapy. J. Cell Commun. Signal. 8, 341–352 (2014). https://doi.org/10.1007/s12079-014-0250-x
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s12079-014-0250-x