
Abstract
The Early Permian marine strata exposed at the Siang and Subansiri districts, Arunachal Pradesh are known for brachiopod and molluscan fossils. A stratigraphic review and palaeoenvironmental, palaeoecological, and palaeobiogeographic interpretation of these Early Permian fossils have been dealt here on the basis of additional fossil and rock materials. Our study confirms the following: (1) the brachiopod-dominated assemblage represents Early Permian, most likely the Sakmarian and younger, age; (2) all marine fossils were suspension feeding groups, representing normal marine condition. Based on the associated sediments and ecologies of taxa, the autecology of the assemblage represents an oxic, soft substrate condition, typical of shallow shelf environment; the plant remains of the upper Khelong/Bhareli Formation, however, indicate a coastal environment; (3) the depositional paleoenvironment indicates the waning phase of a marine glacial episode; and (4) this Early Permian bivalve-brachiopod assemblage show Gondwanan affinity, having strong faunal similarity with the East and West Australia, Nepal and North India (Kashmir) which were part of the Indoralian Province.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
1 Introduction
The Permian was an interval of some major tectonic changes including the breakup of the peri-Gondwana and spreading of the Neotethys ocean. During this interval, the northern part of India, Tibet, Pamir, and Karakorum (Pakistan) were drifting apart from the Gondwana land (Angiolini et al. 2007; Shen et al. 2013). The remnants of the marine habitats of the faunal assemblage of these areas have been the focus of much research efforts (Nakazawa et al. 1975; Singh 1988; Singh and Archbold 1993; Shen et al. 2003; Angiolini et al. 2005). In the north-eastern part of India, the Early Permian fossil-bearing marine sediments of Arunachal Pradesh occur in an east–west strip and are associated with the coal-bearing Gondwana (Acharyya et al. 1975; Kesari 2010), although detailed works on these rocks are mostly lacking.
These aforementioned sediments of Early Permian age, exposed along Siang, Kameng, Subansiri, and Papum Pare districts in Arunachal Pradesh are lithologically quite diverse (Roy Chowdhury and Sinha 1984). From the Darjeeling foothills to the West Siang district, the Gondwana sequence of Eastern Himalaya has been mapped (Jain and Thakur 1975), but the continuity of the sediments in the direction of the strike is truncated east of Siang river by the Siang Fracture Zone (Nandi 1975). Laskar (1959) proposed that the northern boundary of these rocks exposed at Siang and Subansiri districts show thrusted nature of contact. Later, Laskar and Roy Chowdhury (1977) added that these units are basically bounded by two regional thrusts – the northern part upthrusted against the Siwaliks by the Main Boundary Thrust (MBT), and the southern part associated with older metamorphosed and migmatized rocks (Roy Chowdhury and Sinha 1984).
Extensive tectonic disturbances make it difficult to identify the continuity of the lithofacies laterally. The Early Permian sediments are conformably lying above the Miri Formation, a sequence of cross bedded-ripple marked, sandstone intercalated with fine shale but the stratigraphic status of this unit is still debatable as it has been correlated with numerous other formations (Kumar 1997; Kesari 2010). Above this formation, Early Permian fossil-bearing units are present, which has been regionally correlated with Bijni Formation in Uttarakhand, Sisne Formation in Nepal, Rangit Pebble Bed in Darjeeling, and Diuri Formation in Bhutan (Nandi 1975; Laskar and Roy Chowdhury 1977; Bhargava 2008). The basic character of these correlatable formations is their overall arenaceous composition, frequently associated with coaly units, and discontinuous horizons with rich near-coastal and marine macro-invertebrates. However, although many attempts have been made to raise a generalized stratigraphy of this region, no consensus has been obtained.
In this context, the present study attempts to perform the following objectives: (1) provide a concise review of the regional lithostratigraphic and biostratigraphic frameworks, which has been followed in the present study. Associated sedimentological aspects were also analyzed to further confirm the litho- and biostratigraphic characteristics of these marine units; (2) recently, during several fieldworks in the west of Siang river of Arunachal Pradesh, we collected several marine fossils (mostly brachiopods) from these formations. Integrating lithofacies and fossil distributions, we provide a detailed palaeoecological, palaeoenvironmental, and palaeobiogeographic accounts of these Early Permian fossils to identify their global affinities.
2 Material and methods
2.1 Stratigraphic and palaeontological accounts
Since there is no consensus regarding a generalized regional stratigraphy of the Permian rocks exposed in Siang and Subansiri districts, Arunachal Pradesh (figure 1), we first attempted to provide a general, concise stratigraphic review. Some of these units have rich marine fossils. Sedimentological details of the fossiliferous units and representative rock samples were collected for thin section studies to ascertain the stratigraphic position and for the palaeoenvironmental studies.

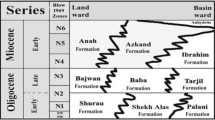
Comparative stratigraphic schemes of Arunachal Pradesh proposed by different researchers. Vertical lines temporally differentiating different members and groups are somewhat arbitrary.
Several field investigations were conducted to study the Permian rocks exposed in and around Rilu, Tatamori, Littemori, and Kimin, all in the west of Siang river, in between 2016 and 2018 (figure 2). Fossils have been collected from well-defined stratigraphic units of the Bomte Member of the Bichom Formation, and the Khelong/Bhareli Formation (figure 3). These fossils are hosted in a black carbonaceous, grey shale unit intercalated with thin sandstone bands with calcareous, ferruginous cherty nodules (figure 4). Other than this, a few nodules host brachiopod or other types of fossils inside it as a nucleus (figure 5). These nodules are similar to the ones recorded by Laskar in Ranga valley, confirming that the Early Permian brachiopod and molluscan fossils of the Siang and Subansiri districts belong to the Bomte Formation. In comparison, in Littemori, the fractured carbonaceous nodules have numerous mollusc fossils, dominantly gastropods, which are otherwise rare. In all cases, the state of preservation of fossils, whether they are present within the concretion or within the sediment, are always poor – the original shell materials have been removed during diagenesis and only the casts and some moulds are preserved (figures 5 and 6). Thin section study also indicates that concretions are fossiliferous with fossil shells partially or fully dissolved with faint traces of chamber walls that differentiates it from concretions. Because of the poor preservation and patchy distribution of fossils, instead of bulk samples, all identifiable shells or macroinvertebrates were collected. For the same reason, it was impossible to identify these up to species level and hence, we restricted our study up to the genus and family levels.

Distribution of Early Permian rocks along the Siang and Subansiri districts in Arunachal Pradesh. The map also shows fossil locations (star marked).

Stratigraphic column used in the present study. For details, see Kesari (2010) and the stratigraphic review discussed in the text.

Major fossiliferous rocks and related thin sections of Bichom Formation. (A–D) thin sections, (A) calcite rhombs of various sizes and shapes. In the groundmass, silty angular fragments of clasticsare present. Note, the presence of microfossil, foraminifera, (B) angular grains of quartz, feldspar, mica, and pyrites as opaque, mostly pyrites, (C) organic-rich shales with occasional lumpy phosphorites, (D) angular to sub-rounded grains of quartz and feldspar, and few mica flakes; (E, F) rocks with brachiopod fossil clusters from the Bichom Formation at Nina river, Tatamori, and Rilu, respectively. Scale bars for E and F = 2 cm.

Different types of concretions commonly present in the Bichom Formation from the study area. (A–B) Carbonaceous-phosphatic nodules with brachiopod fossils inside, at Rilu and Nina river, Tatamori, respectively, (C) unfossiliferous carbonaceous silty–shaly nodule showing concentric lamellae at Daring, (D) carbonate nodule with numerous moulds of small-sized molluscs and other shell fragments from Littemori.

Major macroinvertebrate fossils collected from the Early Permian locations in Arunachal Pradesh. (a–h) brachiopods, (a, b) Tivertonia sp., (c–f) Costatumulus sp., (g) Cyrtella sp., (h) Trigonotreta sp., (i) a broken specimen of the bivalve Aviculopecten sp., (j) Fenestellidae bryozoan fragment, (k) crinoid fragment, (l) conularid fragment, (m) remains of the plant Vertebraria sp., (n) an unidentified seed. (a–c), j, (l–m) from Tatamori, (d–i), k, n from Rilu.
The taxonomic validity of all previously reported taxa was checked using the Paleobiology database (www.paleobiobd.org/). Genera recorded in palaeontological literature with supporting taxonomic description were considered, and genera which did not have taxonomic information but were only recorded in stratigraphic reports are excluded. Finally, to validate the stratigraphic ages of these fossils, stratigraphic distributions of the previously-reported genera and species have been noted from the Paleobiology database. The ecological characteristics of these fossils have been listed from the Fossilworks website (www.fossilworks.com).
2.2 Paleobiogeographic accounts
In the present work, we have comprehensively studied the assemblage, using different multivariate statistical techniques, so that the faunal similarities of the Early Permian assemblage of the Arunachal Pradesh can be compared statistically with the other paleoprovinces within the Gondwanan and Palaeoequatorial realms. Global occurrences of the Early Permian genera have primarily been obtained from the Paleobiology database supplemented with other publications (Runnegar 1969; Waterhouse 1980, 1987; Shen 2018). Since the Indian record is poorly represented in the Paleobiology database, we used few additional publications to improve this database, the most important of these are Nakazawa et al. (1975) and Ghosh (2003). Also, because the unequal representation of occurrence data may introduce sampling-related bias in the similarity analyses, any genera for which at least two occurrences other than that from Arunachal Pradesh is not known, have not been considered, accordingly; Fredericksia, Neoschizodus, Mourlonopsis, Nucula, and Goniophora are deleted from the analysis. Finally, a total of 15 brachiopods and 15 molluscan genera are included for the analyses.
For the paleobiogeographic analyses, a taxonomic presence–absence matrix has been made to compare our taxa with the global database based on previously reported general as well as genera collected during our fieldwork (Supplementary table S1). Global occurrences of our listed genera have been noted for the following areas: Central India, Kashmir (North India), Nepal, Tibet, Pakistan, Afghanistan, West Asia (Oman and Iran), Pamir, China, Mongolia, East Australia, and New Zealand and West Australia and the locations were grouped based on their respective Early Permian palaeobiogeographic positions: (1) Central India; (2) North India–Nepal, together; (3) North-East India (Arunachal Pradesh); (4) Tibet–Pakistan; (5) Afghanistan–Pamir–West Asia, together; (6) Western Australia; (7) South America; (8) China–Mongolia; and (9) East Australia–New Zealand (figures 7 and 8). Among these, the first four areas come within the Himalayan Province; number 5 in the Cimmerian Province; number 6 in the Westralian Province, number 7 in the Andean Province; number 8 within the Cathaysian Province; and number 9 within the Austrazean Province (Shi and Archbold 1993; Shi and Grunt 2000; Shen et al. 2013; Shen 2018). Note, China–Mongolia is considered in both Cathaysian Province as well as in the North Transitional Zone of Shen et al. (2013) and lies in the Palaeoequatorial Realm. According to Shen et al. (2013), the Indoralian Province included all of Australia, Central India, North India, and the Himalayas, and therefore our stations from 1–4, 6, and 9 are in it. In other words, the Indoralian Province included the Himalayan, Westralian, and Austrazean Provinces. According to palaeolatitudinal position, the Himalayan Province was located in between the Gondwanan Realm and the Palequatorial Realm.

Palaeobiogeographic distribution of brachiopod and bivalve genera found during the Early Permian time interval. Note, Arunachal Pradesh (Arunachal) indicate strong affinity with the other Indoralian as well as with Himalayan and Gondwanan locations. Other locations: E. Austr-N. Zea = East Australia–New Zealand, W. Austr = West Australia, N. Ind-Nep = North India–Nepal, S.Amer = South America, Cent Ind = central India, Pak-Tib = Pakistan–Tibet, Chi-Mon = China-Mongolia, Afg-Pam-W.Asia = Afghanistan–Pamir–West Asia.

Non-matric Multidimensional Scaling (nMDS) plot showing cluster of nine palaeobiogeographic locations and their closeness with respect to our Arunachal Pradesh (Arunachal) assemblages. Note, our assemblages are plotted close to the other Indoralian as well with Himalayan and Gondwanan locations. In comparison, China–Mongolia (Chi-Mon) and Afghanistan–Pamir–West Asia (Afg-Pam-W.Asia) are plotted distinctly away. For other abbreviations, see figure 7.
Based on these data, taxonomic compositional similarities among these nine subprovinces are compared by using the hierarchical unweighted pair–group cluster analysis method (UPGMA). Two similarity coefficients – Jaccard and Dice – are used because these two coefficients are least affected by sampling biases and have commonly been used in previous studies involving palaeobiogeographic analysis using the Permian brachiopods (Shen et al. 2009, 2013; Ke et al. 2016). Along with this, the Non-metric Multidimensional Scaling (nMDS) was also used to visually analyze these clusters; we have used Jaccard similarity coefficients for this purpose. Stress values of the nMDS plots (both 2- and 3-dimensional) are used to identify the quality of the cluster plot. All cluster analyses are performed using PAST (Hammer et al. 2001).
3 Stratigraphy
3.1 Lithostratigraphy
The stratigraphic units lying above the Miri Formation and representing the Early Permian age (Acharyya et al. 1975; Kesari 2010), can be subdivided into the following units discussed below.
3.1.1 Bichom Formation
Acharyya et al. (1975) identified the presence of a pebble bed lying unconformably above the pre-Gondwana succession, which they named as Rangit Pebble Bed (equivalent to Rangit Boulder Bed: Kapoor and Maheshwari 1978), because of the overall lithological similarity with the Rangit Pebble-slate unit exposed at the Sikkim–Darjeeling area. However, later researchers did not consider this name and according to these new classification schemes, the stratigraphically equivalent rock units have been considered as Bichom Formation (Dhoundial et al. 1989; Kumar 1997; Bhusan 1999).
The Bichom Formation, exposed near Tatamori, Littemori, Rilu, and Daring in the West Siang district, was later subdivided into three members – Rilu, Bomte, and Sessa, in ascending order (Dhoundial et al. 1989; Kumar 1997; Kesari 2010). The lithology of the Rilu Member matches strongly with the Rangit Pebble Bed of Acharyya et al. (1975). For example, the Rilu Member is mainly diamictitic, along with sandstones, coloured shales, and grits (Singh 1983). Diamictites have angular and sub-angular clasts of quartzite, dolomites, and rock fragments of varied compositions including granite and gneiss. In comparison, the Rangit Pebble Bed is also diamictitic, along with siltstone, quartzite, limestone, etc. (Acharyya et al. 1975). Srivastava et al. (1987) treated this diamictite unit as a separate member (i.e., Diamictite Member) from the upper Rilu Member; these two together were assigned a new formation: Rangit Formation. However, Acharyya et al. (1975) reported numerous sedimentary structures associated with the Rangit Pebble Bed. Therefore, the stratigraphic equivalence needs to be confirmed from the floral and faunal assemblages. Acharyya et al. (1975) identified several bivalves, brachiopods, bryozoans, crinoid ossicles, and ichnotaxa from this member. The Rilu Member also has similar faunal assemblage (Singh 1983; Kesari 2010), suggesting that the Rilu Member and the Rangit Pebble Bed (therefore the Diamictite Member) may be stratigraphically equivalent (figure 3). The Rilu Member is present around Tatamori, Gensi, Rilu and Daring.
Above the Rilu Member conformably lies the Bomte Member and is represented by grey–black shale with calcareous and other types of nodules (Singh 1983; Kesari 2010). Our field and sedimentological observations also reveal that, in all locations (Littemori, Tatamori, Rilu, and Daring), the Bomte Member rocks are very fine-grained (i.e., shale) and thinly laminated, dark-grey to olive-green to greenish-grey in colour (figure 4). Locally, intermittent thin, lensoid bodies of siltstones are present, which are often calcareous and micaceous, and they alternate with the non-calcareous black or steel-grey shale. Late diagenetic calcite veins are observed as white streaks traversing the grey shales (figure 4). These units are also characterized by pyrite enrichment throughout the layers and show variable abundance. Rocks show shiny appearances under normal light and are very hard and compact. Spatially, these rocks frequently host oval to lenticular-shaped nodules of varied compositions (i.e., calcareous shale, chert, and phosphate) (figure 5).
Our observations further reveal, in thin section, the presence of silt lenses containing angular grains of quartz, feldspar and mica with opaques mostly framboidal pyrites was noticeable (figure 4). Silt fragments, 20–50 µm in size, consist of mostly angular quartz and feldspar with fewer amount of mica flakes and show reaction with the calcareous groundmass along the boundaries. Shales at places are organic-rich showing a dark appearance along with the presence of lumpy phosphorites (figure 4). It contains partings and fractures that are filled with carbonates. Partings are regular in appearance and parallel to each other while the fractures are irregular in nature and cross-cut each other as well as the partings. At few places calcite rhombs of various sizes and shapes are present. Importantly, foraminifera ranges in size from 1–1.5 mm with abundant test fragments scattered in the groundmass along with silty angular fragments of clastics (figure 4). The Bomte Member is highly fossiliferous with rich marine brachiopod and bivalve fauna, sometimes present inside the concretions (figures 4 and 5) (Acharyya et al. 1975; Singh 1983).
Above the Bomte Member, a grey to black tuffaceous shale unit with sandstone/quartzite lenses represents the Sessa Member (Dhoundial et al. 1989; Kumar 1997; Bhusan 1999; Kesari 2010). This member also hosts ill-preserved brachiopods and bryozoans (Kesari 2010). The Sessa Member is observed only after crossing Rilu towards Igo. A similar lithostratigraphic unit, known as the Sikki Abu Member (Singh 1983) also consists of pebbly mudstones with clasts of quartzite, sandstones, as well as limestone and igneous rocks. Unfortunately, Singh did not find any fossil from this member, therefore the stratigraphic equivalence of the Sikki Abu Member with the Sessa Member could not be confirmed. However, Srivastava et al. (1987) considered Siki Abu Member as part of the Diamictite Member of his Rangit Formation (figure 1).
These three members, together, or in parts, have been called as Garu Formation by Kapoor and Maheshwari (1978), Singh (1979, 1983) and Kumar (1997). Srivastava et al. (1987), however, placed this Garu Formation above the Rilu Member (figure 1). Because of the overall similarities, we think that the Garu Formation is synonymous with the Bichom Formation (Dhoundial et al. 1989; Kumar 1997; Bhusan 1999). The Garu Formation or the equivalent Bichom Formation appears to be stratigraphically equivalent to the Gensi Formation of Roy Chowdhury and Sinha (1984), because of exactly similar lithology and associated marine and continental floral and faunal associations.
3.1.2 Bhareli and Khelong Formations
Bhareli Formation conformably overlies the Bichom Formation, and represents an overall continental sedimentation (Kumar 1997). According to Acharyya et al. (1975), the Bhareli Formation is equivalent to the Damuda Group of Peninsular India. Acharyya et al. (1975) and Bhusan et al. (1989) identified another formation, the Khelong Formation. While the Khelong Formation has relatively coarser sediments – mostly sandstone – of different colour shades, sediments of the Bhareli Formation are mostly dominated by greenish-grey and black shales and slates of various colours, with occasional occurrences of silts and sandstones (Acharyya et al. 1975). Apart from this, however, they are almost similar: in both cases, shales and coaly units are sometimes present as lenses, and calcareous and pyritic nodules have occasionally been found from these two (Acharyya et al. 1975). Even more, these two formations are floristically similar (Laul et al. 1988; Bhusan et al. 1989). Acharyya et al. (1975) recorded marine faunas from the Khelong Formation, similar to that of the Bhareli Formation (appendix table A1). We therefore, have considered Khelong and Bhareli Formations as stratigraphically equivalent (Kesari 2010), which can be subdivided into two members – Upper and Lower. However, no formal litho- and biostratigraphic accounts are available for these two units.
Laul et al. (1988) recorded gradational contact between the ‘Rilu marine beds’ (Bichom Formation) and the sulphurous carbonaceous shale and siltstone with the coal lenses units (~Bhareli Formation) in the West Siang district. Along the Rilu–Igo road section, outcrops of dark grey sandstone intercalated with grey shale, lenticular coal beds and occasional tuffaceous material representing the Bhareli Formation have been observed by the present authors. The Bhareli Formation, along the Subansiri and Siang districts, shows some lithological and faunal variations and is identified by the typical presence of calcareous, ferruginous, and phosphatic nodules (figure 5).
As mentioned in the previous paragraph, Acharyya et al. (1975) have described Rilu Formation as a spatially different, but stratigraphically equivalent, of the Bhareli Formation. They also mention that in a hillock at Garu–Rilu road section, a gradational contact between the tuffaceous lithic wacke unit of Rangit Pebble Slate (~Rilu Member, Bichom Formation) grade into the sandstone and siltstone of the Rilu Formation. Near the Rilu temple, a polymictic conglomerate unit, dark greenish siltstone, and shale were also observed by the present authors. The frequent presence of pyrites and siderites are characteristics of the Rilu Formation (Acharyya et al. 1975). At Siang and Subansiri, lithounits host rich marine faunas, like brachiopods, gastropods, bivalves, crinoids, etc. (Chandra 1972; Jain and Das 1973) (appendix table A1), although the distribution is sporadic. Typical marine succession is exposed around Likhabali and extends towards Tatamori (Acharyya et al. 1974). The occasional presence of fresh water mollusc like Eurydesma sp. is also reported (Acharyya et al. 1974). The Rilu Formation was not used by the later researchers and is not entrenched in the literature. From the lithological and fossil descriptions, it is thought to be the equivalent of Bichom Formation.
3.1.3 Abor and Lichi Volcanics
Abor Volcanics is a series of basaltic and andesitic volcanic rocks, with associated volcano-sedimentary units (tuff, lapilli, etc.), which drove numerous attention (see Ali et al. 2012 and references therein). Although there are plenty of studies regarding its composition and distribution, its exact stratigraphic age is still elusive, ranging from Late Carboniferous/Early Permian to Early Eocene (Acharyya et al. 1975; Sengupta et al. 1996; Liebke et al. 2011; Ali et al. 2012) (figure 1). In terms of biostratigraphy, Tripathi et al. (1979, 1981) recorded Early Eocene foraminifers and plants associated with the Abor Volcanics. In sharp contrast, on the other hand, marine molluscs, as well as sporomorphs of the Gondwanan affinity was recorded in sediments associated with this volcanic unit by Sinha et al. (1986). Abor volcanism could have started in the Early Permian (Acharyya et al. 1975; Kesari 2010, and many others) and continued till the early Palaeogene. The rocks associated with the earlier stage of this volcanic event is also compared with the Lichi Volcanics (Kesari 2010).
3.2 Biostratigraphy
The first report of marine invertebrate fossils was provided by Diener (1905) when he described a collection made by JM Maclaren. The rich assemblage were represented by about ten species of brachiopods, reported from sandstones and limestones, although these fossils were never found from in-situ rocks. Similar rolled boulders with rich fossils (Laskar’s collection (B Laskar, a pioneer of studying geology of this area)) was also described by Sahni and Srivastava (1956). The first report of in-situ marine invertebrate came from Ranga valley by Lasker (Krishnan 1958). Later, brachiopod, as well as molluscan fauna were collected from a dark-coloured argillaceous unit by Chandra (1972) from Kimin, from the Subansiri district. Jain and Das (1973) collected additional fossils from the adjoining Kameng district.
Based on the fossils collected from carbonaceous shales and laminated sandstones exposed in the Subansiri district, Singh (1973) reassigned the age of these beds to the Middle to Upper Carboniferous and correlated these with the so-called ‘Gondwana Belt’ exposed at Darjeeling, northern Sikkim, and central Bhutan, as well as with those exposed at Kameng and Siang districts. Singh (1978a, b, c) and Singh and Archbold (1993) described several species of brachiopods in and around Siang. Finally, in 1983 (Singh 1983, 1987), he provided a review of collected fauna from those localities, which he assigned Early Permian (Asselian–Artinskian) age for the assemblage collected from a carbonaceous, shaly Formation, which he called the Garu Formation (figure 1). Acharyya et al. (1974, 1975) also identified the age as Permian. Apart from these macroinvertebrates, diverse palynomorphs and foraminifera have also been reported (Kumar 1997; Kalia et al. 2000).
A review of these macroinvertebrates reveals that the major fauna of the Bichom Formation is brachiopods and molluscs, whereas, the Bhareli/Khelong Formation has diverse plant leaf fossils (appendix table A1). The other megafossils include crinoids, ammonoids, and conularids. About 40 species within 30 genera of brachiopods have been reported; whereas, about 30 species within 20 molluscan genera are reported (appendix table A1). Plant fossils from the Khelong/Bhareli are represented by about 20 species of 10 genera. Out of all taxa, only ~50 were identified up to its species level. Of these 50 species, nearly 85% are taxonomically valid, although the taxonomic status of some of them could not be validated. Most of these literatures listing these taxa never provided taxonomic details or even a standard photograph to cross-check the taxonomic validity. Even, some taxa are completely misidentified: the gastropod genus Peruvispira and the ichnotaxa Oldhamia have been identified as brachiopods (Acharyya et al. 1975; Kesari 2010).
The main genera of brachiopods include Subansiria, Lissochonetes, Linoproductus, and Productus. In comparison, molluscs are always rare, although Eurydesma, Uraloceras, Bellerophon, and mytilids are more common (appendix table A1). The major genera of the floristically diverse Khelong/Bhareli Formation are Glosspoteris and Schizoneura.
In our own collection, 16 Early Permian genera under six families of brachiopods were distributed all over the localities covering the entire stratigraphic range. The dominant brachiopod genera identified in the present study are Costatumulus, Cyrtella, Tivertonia, Linoproductus, Trigonotreta, and Subansiria. The only bivalve genus recorded from the Early Permian is Aviculopecten. The genera have previously been reported from the area and therefore systematics detail are not provided.
In comparison, from the Khelong/Bhareli Formation, we have found plant (figure 6) remains from Rilu. The specimen has been tentatively identified as the characteristic Gondwana plant fossil Vertebraria sp. Acharyya et al. (1975) reported a sponge, Paraconularia sp. (their plate 2, figure A), which resembles this specimen. But Paraconularia is characterized by transverse moderately strong ribs, which are abruptly bent adaperturally and the midline on faces indicated only by slight deflection of ribs along it (Waterhouse 1979), which are not observed in the specimen figured by Acharyya et al. (1975). An unidentified seed has also been collected (figure 6). Apart from these brachiopod and molluscan fossils, bryozoans are also found to be present, representing two families, both reticulate colonial, Fenestellidae and Polyporidae; these bryozoans occur with the brachiopod fauna in the Bichom Formation, as observed by the previous authors (Acharyya et al. 1975; Kesari 2010). Also, crinoid stems and conularids are rarely found.
Among these taxa, many have long stratigraphic ranges, therefore not suitable for biostratigraphic analyses (appendix table A1). For example, the molluscan taxa Ptychomphalina sp. (Silurian–Triassic) and Myalina sp. (Ordovician–Triassic) have long stratigraphic ranges. Similarly, the brachiopod species, Lissochonetes carbonifera (Carboniferous–Triassic), Chonetes carbonifera (Silurian–Triassic), and the genus Chonetes (Silurian–Triassic) also have long stratigraphic ranges. However, at least 34 taxa (16 brachiopods, 16 molluscs, and two plants) have a strict Permian stratigraphic range, having high implications in biostratigraphy.
Based on these Permian taxa, it appears that at least 80% taxa are restricted to the Early Permian only, whereas, few others have a relatively longer range (i.e., Lower to Middle Permian range, or ranging the entire Permian) (appendix table A1). Based on these, we think that the lithounits from where these rocks have been collected have an Early Permian age, as stated by the previous authors (Archarya et al. 1974, 1975). Further exploration reveals that, among these Early Permian taxa, ~32% are only found from the Sakmarian, and the additional ~40% have stratigraphic ranges inclusive of this age. These observations reinforce the claim that this fossil assemblage indicates an Early Permian, mostly the Sakmarian age. Singh (1983, 1987) also, based on his brachiopod collections, assigned the age of these rocks an Early Permian (Asselian–Artinskian) age, which is inclusive of the Sakmarian age. Importantly, taxa form the stratigraphically above, Khelong/Bhareli Formation range from the Permian (Sakmarian) up to the Triassic. We conclude that rocks of the Bichom Formation represent tentatively the Early Permian (most likely Sakmarian) and the overlying Khelong/Bhareli Formation represent a relatively younger, but still Early Permian age.
4 Palaeoecology, palaeobiogeography, and palaeoenvironment of the Early Permian assemblages
4.1 Palaeoecology of the Early Permian fossils
Our collection from the Bomte Member of the Bichom Formation includes a total of about 65 brachiopods specimens which can be separated individually for identification. Other than these, there are many specimens which are clustered together and it was not possible to retrieve without destroying them and we have restrained from doing that. Among brachiopods, Costatumulus sp. (40% of all brachiopods), and Cyrtella sp. (~21% of all brachiopod specimens) were the two most dominant species, whereas, Trigonotreta sp. and Tivertonia tatamariensis were relatively less abundant. The other species are rare. In comparison, bivalves are represented by only one fragmented specimen and has been identified as ?Aviculopecten sp. Brachiopod fauna are sometimes associated with bryozoans, which belong to the two families, Fenestellidae and Polyporidae. Conularids and crinoid fragments are also present. We collected only one plant specimen and one tentative seed from the Khelong/Bhareli Formation, and no such ecological analysis was possible for this formation.
It is therefore evident that, in terms of dominance, brachiopods represent almost 100% of specimens collected. This typical brachiopod dominance over bivalves has also been reported from other Permian records from other parts of the world (Clapham and Bottjer 2007; Payne et al. 2014). This matches well with the established global pattern. However, this dominance pattern was not same throughout the Permian – the Early Permian was completely dominated by the brachiopods, but after that, bivalves and other molluscs started to increase their dominance globally (Clapham and Bottjer 2007). If an Early Permian assemblage is compared with a Late Permian assemblage, the Late Permian assemblage has a relatively large number of molluscs compared to its Early Permian counterpart, and in both cases brachiopods are the dominant component. This overall Permian pattern was followed globally across different basins, suggesting an ecological cum environmental changes happening during the late Palaeozoic (Clapham and Bottjer 2007) and other Early Permian data fits with this model. It should be noted that this is also observed in Siang valley: the proportional dominance of brachiopods over bivalves weakens a bit during the late Permian in Arunachal Pradesh. The Early Permian faunal assemblages collected from Dalbuing are still brachiopod dominant but the taxonomic and ecologic dominance of bivalves have started to increase (Mukherjee et al., in preparation). For this reason, it is of enormous importance to study the Permian brachiopod–bivalve dynamics in much finer time bins, i.e., at the Epoch level so that the aforementioned change in abundance can be documented across different ocean basin globally.
In terms of life modes of the Bichom fauna, similar to the global trend, most of the Early Permian brachiopod fauna were epifaunal free-lying forms attached by the spines (like productids including the chonetacians) or delthyrially attached forms like spiriferids (e.g., Neospirifer) matching well with the life habits of the Permian fauna. All were suspension feeders, as did the single bivalve species. Therefore, considering the prevalence of brachiopods, which are exclusively marine, it can be concluded that the Bichom fauna represents a normal marine ecology. The most abundant taxa, Costatumulus, has fine radial ribbing, costellae, and rugae at the outer shell surface (Singh and Archbold 1993), which resembles with Costatumulus described from the Upper Permian of South China (He et al. 2014). According to He et al. (2014), this morphology has been interpreted as morphological adaptations of deepwater adaptation to a highly stressed environment. The specimens observed by the present authors were all internal moulds and therefore shell microstructure could not be studied. Therefore, it cannot be ascertained if the presence of pseudopunctae is restricted in the late Permian species as an adaptation of environmental stress or it is a plesiomorphic character of the genus. Interestingly Costatumulus has maximum abundance in the area and the high density of spines could have aided in anchorage and protection from predation.
4.2 Palaeoenvironment of the Early Permian fossils
Acharyya et al. (1975), by using the sedimentological and general fossil occurrences, proposed that during the Early Permian the depositional milieu was a near-coastal, brackish water setup. However, their palaeoenvironmental observations and interpretations were preliminary, which have been revisited and reanalyzed here.
The faunal composition of the Bichom Formation fauna represents a normal marine autecology. Based on all fossils that have ever been collected from here, it can be said that these types of ecological assemblages are typical of the near-coastal marine habitats where substrates are mostly soft. The majority of these fauna is suspension feeding, suggesting an oxygen-rich condition, which is further supported by the presence of bryozoans. However, their ecology indicates a varied water depth, ranging from shallow to relatively deeper shelf condition: benthic invertebrates represents shallower condition, as do the bryozoans (usually live within 100 m of water depth), whereas crinoids are mostly representative of deeper water condition. Moreover, the presence of some morphological features (e.g., crowded pseudopunctae) on one of the most abundant brachiopod genus, Costatumulus, further supports this view (He et al. 2014). A similar interpretation has been made by the previous authors also: according to them, the palaeoenvironmental condition of the Bomte Member of the Bichom Formation indicates that the environment of deposition was from a relatively nearshore (marginal marine, swampy environment) to the ‘epineritic’ environment, in many cases depositing just below the wave base under normal salinity and 25–31°C temperature (Roy Chowdhury and Sinha 1984; Kumar 1997). In a stable shelf of low relief, perhaps a slight change in the sea level resulted in great lateral displacement of the shoreline and lithofacies and biofacies variation. The presence of small lensoidal bodies of coarser sediments within fine sedimentary units support this type of intermittent sea level fluctuations. Fine jet-black to yellowish-brown shale could further support this view.
Under these conditions, sediments were deposited at the nearshore environment with chemical conditions suitable for the formation of different types of nodules (Roy Chowdhury and Sinha 1984). The productids are also thought to be capable of living in a relatively higher energy conditions as the spines acted as stabilizers to reduce the scouring action (Mendonca et al. 2018). However, the associated sediments in many cases are dark-coloured shale, which suggests that the shells were transported from their normal marine habitat and were deposited in a nearby shallow, near-coastal, marginal-marine environment like back-swamp or lagoon, transitional brackish environment, and continental alluvial fans (Acharyya et al. 1975). The presence of coaly unit and secondary pyrites as revealed in thin section study further confirms this claim. Moreover, the presence of lumpy and disseminated phosphates and pyrite associated with disseminated organic matter indicates anoxic condition (figure 4). Later, due to widespread marine transgression, because of deglaciation (Singh 1987), the coastal marine regions became inundated; the sediments of Bhareli/Khelong Formation was deposited under a fluvial setting (Srivastava et al. 1987).
Climatologically, the litho- and the biofacies indicate that the Bichom Formation was deposited during the waning phase of a glacial marine influence as indicated by the presence of diamictites (Singh 1988). This diamictitic unit has been correlated with the Talchir Boulder Bed of the Peninsular India, which indicates a typical glacial boulder bed (Oldham 1887; Acharyya et al. 1975 and references therein). In fact, the Rilu Member has been correlated with the Peninsular Lower Karharbari Member, indicating that glacial retreat (op. cit.). The Costatumulus–Trigonotreta–Tomiopsis brachiopod assemblage along with the bivalve Eurydesma are characteristic cold water assemblage of the Sakmarian (Shen et al. 2013), which is seen as the dominant fauna in the Bichom Formation. This association along with Bandoproductus and Puncocyrtella is characteristic of the Himalayas, south and Central Tibet, Baoshan Sibamusu blocks of SE Asia (all being under cold water influence). However, with the melting of the Gondwana ice caps in the Late Sakmarian, the warmer conditions were setting in, at some regions as evidenced by the fusulinid fauna. The presence of a fusulinid genus Earlandia have been reported by Kalia et al. (2000) from the Bichom Formation (their Garu Formation). The presence of bryozoans in the Bichom Formation, which are characteristic of tropical waters, is also indicative of the warmer condition. Although so much has been said for the palaeoclimatic condition of the Bichom Formation, the same for the overlying Khelong/Bhareli Formation is poorly known, it probably represent a much warmer, but moisture-rich, phase, equivalent to the Upper Coal Measures in Darjeeling, West Bengal (Srivastava et al. 1987).
4.3 Palaeobiogeography of the Early Permian fossils
Three marine palaeobiogeographic realms, based on the palaeolatitudes of different biogeographic locations, have been recognized during the Permian: Gondwanan, Palaeoequatorial/Tethyan and Boreal; in addition, two transitional zones were also recognized: Southern and the Northern Transitional zone (Shi and Grunt 2000; Shen et al. 2013). For the present study, the faunal stations (i.e., locations from where the Early Permian fauna have been reported globally) were chosen from the Palaeoequatorial/Tethyan and Gondwanan realms, because of their proximity with Arunachal Pradesh during the Early Permian. For the same reason, the Boreal Realm, which was far apart from Arunachal Pradesh, was not considered for this analysis. The Early Permian palaeobiogeographic study by Shen et al. (2013) showed that the palaeo-position of the East Himalaya was the closest to the North Indian plate, and these two clusters together with Tibet, Kashmir, NW Himalaya, Pamir, and Karakorum (Pakistan). Though Kashmir, East Himalaya, and Peninsular India fauna of the Early Permian was placed within the Gondwana Realm by Shi and Grunt (2000), later Shen et al. (2013) put them in the Indoralian Province which belongs to the peri-Gondwanan realm. Shen et al. (2013) also showed that, by the Sakmarian, the Indoralian Province can be divided into the Austrazean Province and the Southern Transitional Zone. The Himalayan Province, proposed by Singh (1987, 1988), included faunas from the Himalayan regions with Peninsular India and later Singh and Archbold (1993) have linked the Garu fauna in Arunachal Pradesh with Afghanistan in the west to Australia in the East.
Our analyses reveal that the Early Permian macroinvertebrates show good clustering with respect to their respective palaeo-provinces (figure 7). Since the outcome of the analyses, based on Jaccard and Dice similarity coefficients are similar, we are hereby discussing the results based on the Jaccard coefficients (J) only. The faunal assemblage of Arunachal Pradesh shows the strongest resemblance with East Australia–New Zealand (J = 0.80), and is strongly related to West Australian (J = 0.73), and North India–Nepal (J = 0.70) faunal assemblages (figure 7). Similarities with Pakistan–Tibet (J = 0.67), South American (J = 0.60), and Central India (J = 0.57) are also moderately high. The Early Permian fauna from Arunachal Pradesh has low similarities with the assemblages from Afghanistan–Pamir–West Asia (J = 0.43) and China–Mongolia (J = 0.47). This clustering is also supported by the nMDS plot (k = 3; stress = 0.14), where our assemblages are plotted very close to East Australia–New Zealand (figure 8). North India–Nepal, South American, and Pakistan–Tibet are also very close to our assemblages. Notably, matching with the cluster plot, Afghanistan–Pamir–West Asia and China–Mongolia are plotted farthest from our assemblages (figure 8).
This pattern of paleobiogeographic similarities match well with the proposed biogeographic affinities of these palaeo-locations; the Indoralian Province of Shi and Archbold (1993) and Shen et al. (2013) is also observed in the cluster plot (figure 7). For example, locations within the Himalayan Province (i.e., Pakistan, Tibet, North India, Nepal, Arunachal Pradesh, and Central India) are clustered together, and these locations also show similarity with the Westralian and Austrazean provinces. Also, South America, being in the Andean province of the Gondwana Realm show close proximity with our assemblages and these general observations are also supported by previous authors. According to Acharyya et al. (1975), the Arunachal Pradesh fauna is similar to those of the peninsular Gondwana records (Srivastava et al. 1987). Singh and Archbold (1993) also established the Gondwanan affinity of their Garu fauna (presently the faunal assemblage of Bichom Formation as discussed at length previously) with Australia.
In comparison, palaeolatitudinally, during the Early Permian, these Himalayan clusters were wide apart from the Cimmerian blocks represented by Afghanistan, Pamir, and west Asian countries (e.g., Oman, Iran), and Cathaysian Province represented by China and Mongolia which were near the equator; for this reason, Arunachal Pradesh clusters to the farthest (figures 7 and 8). According to Angiolini et al. (2013) and Shen (2018), Afghanistan, Pamir, and west Asian part within the Cimmerian blocks broke apart from the Gondwanan block during the Early Permian and began their northward drift, which explains the lower similarities among them. Palaeobiogeographic studies of Shen et al. (2013), in fact, recognizes a supergroup – the Southern Transitional Zone – in the Sakmarian that have members situated in the Cimmerian blocks, Himalayan Province, Sibamusu terranes and Westralian Province and they further grouped Peninsular India, Kashmir and Himalayan regions with Australian stations and showed that the Indoaralian Province divided into the South Transitional Zone and the Austrazean Province in the late Sakmarian (Shen et al. 2013). Our study, which was mainly based on the regions from the Southern Transitional Zone of Shen et al. (2013) and Cathaysian Province from the North Transitional Zone (Angiolini et al. 2013), validates the existence of the Indoralian Province. Our results also show that the Arunachal fauna perhaps depicts the time just before the Late Sakmarian, when the Australian fauna was separating from the Himalayan Province which was within the Southern Transitional Zone. Interestingly one of the characteristic brachiopod genus of the province is Trigonotreta which has been found in moderate abundance in the study area. Kalia et al. (2000) based on multiple species of foraminifera from Tatamori, also had a similar palaeobiogeographic interpretation. Archbold (1983, 1987, 1993) also referred it as peri-Gondwanan assemblage.
References
Acharyya S K, Ghosh S C and Ghosh R N 1974 Biostratigraphic study of the continental and marine Arunachal Gondwana and new lights in the pre-Gondwana rocks of Arunachal Himalayas; Ind. Mineral. 28 103–108.
Acharyya S K, Ghosh S C, Ghosh R N and Shah S C 1975 The Continental Gondwana Group and associated marine sequence of Arunachal Pradesh (NEFA), Eastern Himalaya; Him. Geol. 5 60–82.
Archbold N W 1983 Permian marine invertebrate provinces of the Gondwanan Realm; Alcheringa 7 59–73.
Archbold N W 1987 South Western Pacific Permian and Triassic marine faunas: Their distribution and implications for terrane identification; Terrane Accret. Orogen. Belts 19 119–127.
Archbold N W 1993 Studies on western Australian Permian Brachiopods 11, new genera, species and records; Proc. Royal Soc. Victoria 105 1–29.
Angiolini L, Brunton H and Gaetani M 2005 Early Permian (Asselian) brachiopods from Karakorum (Pakistan) and their palaeobiogeographical significance; Palaeontol. 48 69–86.
Angiolini L, Gaetani M, Muttoni G, Stephenson M H and Zanchi A 2007 Tethyan oceanic currents and climate gradients 300 myr ago; Geology 35 1071–1074.
Angiolini L, Zanchi A, Zanchetta S, Nicora A and Giovanni V 2013 The Cimmerian geo puzzle: New data from South Pamir; Terra Nova 25 352–360.
Ali J R, Aitchison J C, Chik S Y, Baxter A T and Bryan S E 2012 Paleomagnetic data support Early Permian age for the Abor Volcanics in the lower Siang Valley, NE India: Significance for Gondwana-related break-up models; J. Asian Earth Sci. 50 105–115.
Bhargava O N 2008 Palaeozoic successions of the Indian plate; Mem Geol. Soc. India 74 209–244.
Bhusan S K, Banerjee D C, Bindal C M, Sharma S, Agrwal R K, Passayat R N and Srivastava J K 1989 Preliminary report on section measurement in parts of West Kameng and Lower Subansiri districts, Arunachal Pradesh; Unpubl. Rep., Geol. Surv. India.
Bhusan S K 1999 Reappraisal of the geology between the MBF and the MCT in western Arunachal Pradesh; In: Geological Studies in the Eastern Himalaya (ed.) Verma P K, Pilgrim books Pvt. Ltd., Delhi, pp. 161–176.
Chandra S 1972 New localities for marine beds in association with Arunachal Gondwanas; Soil Cult. 38 361–364.
Clapham M E and Bottjer D J 2007 Permian marine paleoecology and its implications for large-scale decoupling of brachiopod and bivalve abundance and diversity during the Lopingian (Late Permian); Palaeogeogr. Palaeoclimatol. Palaeoecol. 249 283–301.
Dhoundial D P, Kumar G, Singh S and Reddy K V S 1989 Geology of Arunachal Himalaya; Unpubl. Rep., Geol. Surv. India.
Diener C 1905 Notes on Anthracolithic fauna from the mouth of Subansiri gorge; Rec Geol. Surv. India 32 189–198.
Ghosh S K 2003 First record of marine bivalves from the Talchir formation of the Satpura Gondwana basin, India: Palaeobiogeographic implications; Gondwana Res. 6 312–320.
Hammer Ø, Harper D A and Ryan P D 2001 PAST: Paleontological statistics software package for education and data analysis; Palaeontol. Electronica 4 9.
He W H, Shi G R, Zhang Y, Yang T, Zhang K, Wu S, Niu Z and Zhang Z 2014 Changhsingian (latest Permian) deep-water brachiopod fauna from South China; J. System Palaeontol. 12 907–960.
Jain A K and Das A K 1973 On the discovery of Upper Palaozoic bryozoans from Kameng district, Arunachal Pradesh; Curr. Sci. 42 352–355.
Jain A K and Thakur V C 1975 Stratigraphic and tectonic significance of the Eastern Himalayan Gondwana belt with special reference to the Permo-Carboniferous Rangit Pebble Slate; Bull. Ind. Gondwanan Assoc. B2 8–9.
Jain A K and Thakur V C 1978 Abor Volcanics of the Arunachal Himalaya; J. Geol. Soc. Ind. 19 335–349.
Kalia P, Pande P K and Singh T 2000 Early Permian Foraminifera from Eastern Himalaya and their palaeobiogeographic significance; Alcheringa 24 207–227.
Kapoor H M and Maheshwari H K 1978 Early Permian palaeogeography of the Perigondwana in the Indian segment; Curr. Sci. 61 648–653.
Ke Y, Shen S, Shi G R, Fan J, Zhang H, Qiao L and Zeng Y 2016 Global brachiopod palaeobiogeographical evolution from Changhsingian (Late Permian) to Rhaetian (Late Triassic); Palaeogeogr. Palaeoclimatol. Palaeoecol. 448 4–25.
Kesari G K 2010 Geology and Mineral Resources of Arunachal Pradesh; Misc. Publ., Geol. Surv. India, 30, 54p.
Krishnan 1958 General Report of the Geological Survey of India for the year 1953; Rec. Geol. Surv. India, 87p.
Kumar G 1997 Geology of Arunachal Pradesh; Geological Society of India, Bangalore, 217p.
Laskar B 1959 On the Permo-Carboniferous of the foothills of the Assam Himalayas; India Min Res. Dev. Ser. 10 171–172.
Laskar B and Roy Chowdhury J 1977 Sedimentation pattern along the margin of the Gondwana continent, Arunachal Pradesh, North Eastern India; IVth Int. Gondwana Symp. Kolkata, India.
Laul V P, Mishra U K and Shrivastava S C 1988 Final report on Gondwana of Arunachal Pradesh; Unpubl. Rep., Geol. Surv. India.
Liebke U, Antolin B, Appel E, Basavaiah N, Mikes T, Dunkl I and Wemmer K 2011 Indication for clockwise rotation in the Siang window south of the eastern Himalayan syntaxis and new geochronological constraints for the area; In: Growth and Collapse of the Tibetan Plateau, (eds) Gloaguen R and Ratschbacher L; Geol. Soc. London, Spec. Pub., pp. 71–97.
Mendonca S E, Barclay K M, Schneider C L, Molinaro D J, Webb A E, Forcino F L and Leighton L R 2018 Analyzing trends in tropical Devonian brachiopod communities during environmental change in the Waterways Formation of northern Alberta; Palaeogeogr. Palaeoclimatol. Palaeoecol. 506 84–100.
Nakazawa K, Kapoor H M, Ishii K I, Bando Y, Okimura Y and Tokuoka T 1975 The upper Permian and the lower Triassic in Kashmir, India, Memoirs of the Faculty of Science, Kyoto University; Ser. Geo. Mineral. 42 1–106.
Nandi D R 1975 Geology of NEFA Himalaya; Geol. Surv. India Misc. Pub. 29 91–114.
Oldham R D 1887 Preliminary sketch of the geology of Simla and Jutogh; Rec. Geol. Surv. India 3 143–152.
Payne J L, Heim N A, Knope M L and McClain C R 2014 Metabolic dominance of bivalves predates brachiopod diversity decline by more than 150 million years; Proc. R. Soc. B 281 20133122.
Roy Chowdhury A K and Sinha J K 1984 Systematic geological mapping on the Gondwana rocks in Siang district Arunachal Pradesh; Unpubl. Rep., Geol. Surv. India.
Runnegar B N 1969 Permian fossils from the southern extremity of the Sydney Basin. In Stratigraphy Palaeontolology; Australian Natl. Univ. Press, Canberra, pp. 276–298.
Sahni M R and Srivastava J P 1956 Discovery of Eurydesma and Conularia in Eastern Himalaya and description of associated fauna; J. Pal. Soc. Ind. 17 50–54.
Sengupta S, Acharyya S K and De Smith J B 1996 Geochemical characteristics of the Abor volcanics zone, N.E. Himalaya, India, Nature of Early Eocene magmatism; J. Geol. Soc. London 153 695–704.
Shen S Z 2018 Global Permian brachiopod biostratigraphy: An overview Geol; Soc. London Spec. Pub. 450 289–320.
Shen S Z, Shi G R and Archbold N W 2003 A Wuchiapingian (late Permian) brachiopod fauna from an exotic block in the Indus–Tsangpo Suture zone, southern Tibet and its palaeobiogeographical and tectonic implication; Palaeontology 46 225–256.
Shen S Z, Xie J F and Zhang Hand Shi G R 2009 Roadian–Wordian (Guadalupian, Middle Permian) global palaeobiogeography of brachiopods; Glob. Planet. Change 65 166–181.
Shen S Z, Zhang H, Shi G R, Li W Z, Xie J F, Mu L and Fan J X 2013 Early Permian (Cisuralian) global brachiopod palaeobiogeography; Gondwana Res. 24 104–124.
Shi G R and Archbold N W 1993 Distribution of Asselian to Tastubian (Early Permian) circum-Pacific brachiopod faunas; In: Palaeontological studies in honour of Ken Campbell (ed.) Jell P A, Mem. Assoc. Australasian Palaeontol. 15 343–351.
Shi G R and Grunt T A 2000 Permian Gondwana–Boreal antitropicality with special reference to brachiopod faunas; Palaeogeogr. Palaeoclimatol. Palaeoecol. 155 239–263.
Singh T 1973 Note on Upper Palaeozoic fauna from Subansiri district, Arunachal Pradesh; Him. Geol. 3 401–410.
Singh T 1975 Discovery of Eurydesma from the Siang district Arunachal Pradesh; Bull. Indian Geologist’s Assoc. 8 90–91.
Singh T 1978a Lower Permian gastropods and bivalves from eastern Himalaya, India; In: Recent Researches in Geology, a Collection of Papers in Memory of Professor A.G. Jhingran, Hindusthan Publishing Corporation, Delhi, India, pp. 276–310.
Singh T 1978b A New Species of Spiriferoid genus ‘Subansiria’ from Subansiri District, Arunachal Pradesh; Contrib. Him. Geol. 1 162–164.
Singh T 1978c Brachiopods from Permian Formation of Siang district, Arunachal Pradesh; Contrib. Him. Geol. 1 171–188.
Singh T 1979 Palynostratigraphy of the Permian rocks of Siang district, Arunachal Pradesh. In Metamorphic rock sequences of the Eastern Himalaya; American Geophysical Union, USA, pp. 191–194.
Singh G 1983 On the stratigraphic correlation of Upper Tertiary of Arunachal Pradesh; Symp. Geol. Min. Res. of N.E. Him., Shillong, Geol. Soc. Ind. Misc. Pub. 43 82–84.
Singh T 1987 Paleoclimatic significance of fauna and mioflora of the Garu Formation (Permian), Arunachal Pradesh, eastern Himalaya; Gondwana Six: Stratigr., Sedimentol. Paleontol. 2 191–194.
Singh S 1988 The Permian brachiopod fauna of the Himalaya: Palaeobiogeographic implications; J. Geol. Soc. Ind. 33 1–6.
Singh T and Archbold N W 1993 Brachiopoda from the Early Permian of the eastern Himalaya; Alcheringa 17 55–75.
Sinha N K, Satsangi P P and Misra U K 1986 Palaentology of Permian and Eocene rocks of Siang district, Arunachal Pradesh; Rec. Geol. Surv. Ind. 114 51–60.
Srivastava S C, Prakash A and Singh T 1987 Permian palynofossils from the eastern Himalaya and their genetic relationship; Palaeobotany 36 326–338.
Tripathi C, Dungrakoti B D and Ghosh R N 1979 Note on discovery of nummulitics from Dihang Valley, Siang district, Arunachal Pradesh; Ind. Miner. 33 43–44.
Tripathi C, Gaur R K and Singh S 1981 A note on the occurrence of Nummulities in East Siang district, Arunachal Pradesh; Ind. Miner. 35 36–38.
Waterhouse J B 1979 New members of the Atomodesminae (Bivalvia) from the Permian of Australia and New Zealand; Dept. Geol. Univ. Queensland Paper 9 1–18.
Waterhouse J B 1980 Permian bivalves from New Zealand; J. Royal Soc. New Zealand 10 97–133.
Waterhouse J B 1987 Late Paleozoic mollusca and correlations from the south-east Bowen Basin, East Australia; Palaeontographic. Abteilung. A. Paläozoologie. Stratigraphie. 198 129–233.
Acknowledgements
The study is part of the GSI FSP 2016–2018 RP/CHQ/M-IV/2016/99 in collaboration with the University of Calcutta and the authors would like to thank the Director General GSI for granting permission to carry out the work. Sabyasachi Shome, GSI is also thanked for his advice and support during fieldwork. The Map and Cartography Division, North East Region Shillong, is also gratefully acknowledged for the help in the preparation of the map of the study area. SM would like to acknowledge University of Calcutta for infrastructural facility and partial funding from the RGT 2017 grant. Deepjay Sarkar helped in photography. Comments and suggestions from the Journal Editor and two anonymous reviewers are acknowledged.
Author information
Authors and Affiliations
Contributions
SM and DM identified the problem and designed the study; DM, SM, BI, and AR conducted fieldwork and collected specimens; SS and SM performed the lithological analyses; SM and DM performed all analyses and interpreted data; SM and DM wrote the manuscript with input from others.
Corresponding author
Additional information
Communicated by Pratul K Saraswati
Supplementary material pertaining to this article is available on the Journal of Earth System Science website (http://www.ias.ac.in/Journals/Journal_of_Earth_System_Science)
Supplementary Information
Below is the link to the electronic supplementary material.
Appendix
Appendix
Rights and permissions
About this article

Cite this article
Mondal, S., Mukherjee, D., Iangrai, B. et al. Early Permian macroinvertebrate assemblages from the Siang and Subansiri districts, Arunachal Pradesh: Implications on the regional stratigraphy, palaeoenvironment, palaeoecology, and palaeobiogeography. J Earth Syst Sci 130, 87 (2021). https://doi.org/10.1007/s12040-021-01569-3
Received:
Revised:
Accepted:
Published:
DOI: https://doi.org/10.1007/s12040-021-01569-3