
Abstract
The ventromedial hypothalamic nucleus-ventrolateral part (VMNvl) is an estradiol-sensitive structure that controls sex-specific behavior. Electrical reactivity of VMNvl neurons to hypoglycemia infers that cellular energy stability is monitored there. Current research investigated the hypothesis that estradiol elicits sex-dimorphic patterns of VMNvl metabolic sensor activation and gluco-regulatory neurotransmission during hypoglycemia. Rostral-, middle-, and caudal-VMNvl tissue was separately micropunch-dissected from letrozole (Lz)- or vehicle-injected male and estradiol- or vehicle-implanted ovariectomized (OVX) female rats for Western blot analysis of total and phosphorylated 5′-AMP-activated protein kinase (AMPK) protein expression and gluco-stimulatory [neuronal nitric oxide synthase (nNOS); steroidogenic factor-1 (SF1) or -inhibitory (glutamate decarboxylase65/67 (GAD)] transmitter marker proteins after sc insulin (INS) or vehicle injection. In both sexes, hypoglycemic up-regulation of phosphoAMPK was estradiol-dependent in rostral and middle, but not caudal VMNvl. AMPK activity remained elevated after recovery from hypoglycemia over the rostro-caudal VMNvl in female, but only in the rostral segment in male. In each sex, hypoglycemia correspondingly augmented or suppressed nNOS profiles in rostral and middle versus caudal VMNvl; these segmental responses persisted longer in female. Rostral and middle segment SF1 protein was inhibited by estradiol-independent mechanisms in hypoglycemic males, but increased by estradiol-reliant mechanisms in female. After INS injection, GAD expression was inhibited in the male rostral VMNvl without estradiol involvement, but this hormone was required for broader suppression of this profile in the female. Neuroanatomical variability of VMNvl metabolic transmitter reactivity to hypoglycemia underscores the existence of functionally different subgroups in that structure. The regional distribution and estradiol sensitivity of hypoglycemia-sensitive VMNvl neurons of each neurochemical phenotype evidently vary between sexes.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
The ventromedial hypothalamic nucleus (VMN) is a principal element of the sex-dimorphic neural network that is established by estrogen receptor (ER) imprinting during the ‘critical period’ of brain differentiation (Simerly 2002). In the adult, estrogen receptors (ERs) are prominently expressed in the ventrolateral region of the VMN (VMNvl) (Yang et al. 2013), a structure that is implicated in sex-specific control of critical physiological and behavioral functions, including sexual and maternal behaviors, physical activity, and metabolic homeostasis (Musatov et al. 2007; Yang et al. 2013; Martínez de Morentin et al. 2014; Correa et al. 2015; Hashikawa et al. 2017). Cellular energy balance is continuously monitored in the VMN and a limited number of other brain loci by specialized metabolic-sensory neurons that adjust synaptic firing when substrate fuel supply deviates from the normal range (Oomura et al. 1969; Ashford et al. 1990; Adachi et al. 1995, 1997; Silver and Erecińka, 1998). This dynamic readout, along with endocrine signals that are surrogates for peripheral energy reserves, provides critical input to VMN control of glucostasis. The ovarian steroid estradiol acts on VMN substrates to regulate blood glucose profiles (Nedungadi and Briski 2012; Liu et al. 2013). VMNvl neurons that exhibit decreased or elevated synaptic activity in response to glucose deficiency (Cotero and Routh, 2009; Santiago et al. 2016) may thus be direct or indirect targets of estradiol modulatory action.
The ultra-sensitive, evolutionarily-conserved energy gauge 5′-AMP-activated protein kinase (AMPK) is activated by phosphorylation when the cellular AMP: ATP ratio increases (Hardie et al. 2016; Lin and Hardie 2018). Mediobasal hypothalamic AMPK activation is required for maximum glucose counter-regulatory responses to insulin-induced hypoglycemia (Han et al. 2005; McCrimmon et al. 2008). Neurotransmitter effectors of ventromedial hypothalamic energy insufficiency include nitric oxide (NO) and steroidogenic factor-1 (SF1) or γ-aminobutyric acid (GABA), which respectively elevate versus suppress counter-regulatory hormone secretion (Chan et al. 2006; Fioramonti et al. 2010; Garfield et al. 2014; Routh et al. 2014; Meek et al. 2016). VMN nitrergic, SF1, and GABAergic neuron function may be directly impacted by glucoprivation as these cell types express AMPK (Ibrahim et al. 2020; Briski and Mandal 2020). AMPK gluco-regulatory signaling likely originates in the VMN as insulin-induced hypoglycemia increases AMPK phosphorylation in local NO and GABA neurons (Briski et al. 2020; Ibrahim et al. 2020). Estradiol evidently controls metabolic sensory signaling by these neurotransmitter cell types in a sex-specific manner, as ER-alpha (ERα) and ER-beta (ERβ) impose differential control of VMN expression profiles of neuronal nitric oxide synthase (nNOS) and glutamate decarboxylase65/67 (GAD), marker proteins for nitric oxide and GABA transmission, in hypoglycemic male versus female rats (Mahmood et al. 2018). VMN NO and GABA neurons are each a likely target for estradiol action as both cell populations express ERα and -β, as well as the transmembrane G protein-coupled estrogen receptor GPER (Uddin et al. 2019).
Previous Western blot studies performed in our laboratory and by others did not analyze VMNvl tissue apart from other, e.g. central and dorsomedial regions of the VMN. Thus, there remains a need to clarify, for each sex, the role of estradiol in hypoglycemia effects on metabolic sensor and gluco-regulatory neurotransmitter activity in this critical sex-dimorphic substructure. The current project utilized high-resolution microdissection/high-sensitivity molecular analytical techniques to address the hypothesis that in each sex, estradiol regulation of basal and/or hypoglycemia patterns of sensor activation and nNOS, SF1, and/or GAD protein expression may vary over the rostro-caudal length of the VMNvl, and that estradiol-mediated sex differences in expression profiles of one or more target proteins may occur at distinctive neuroanatomical levels of the VMNvl.
Materials and Methods
Animals:
Adult male and female Sprague Dawley were housed 2–3 per cage, according to sex, under a 14-h light/10-h dark cycle (lights on at 05.00 h). Animals had ad-libitum access to standard laboratory rat chow and water, and were acclimated to daily handling. All surgical and experimental protocols were conducted in accordance with NIH guidelines for care and use of laboratory animals, under approval by the ULM Institutional Animal Care and Use Committee. Five days prior to experimentation, female rats were bilaterally ovariectomized (OVX) under ketamine/xylazine (0.1 mL/100 g bw; 90 mg ketamine: 10 mg xylazine/mL; Henry Schein Inc. Melville, NY) anesthesia. After surgery, rats were injected subcutaneously (sc) with ketoprofen (1 mg/kg bw) and intramuscularly with enrofloxacin (10 mg/0.1 mL), treated by topical application of 0.25% bupivacaine to closed incisions, then transferred to individual cages. A validated hormone replacement paradigm was implemented in female subjects for the purpose of standardizing plasma estradiol levels at metestrus-like levels, in order to avoid variability due to variation in endogenous circulating hormone secretion over the estrous cycle (Briski et al. 2001). On study day 1, randomly assigned female rats were implanted with a sc silastic capsule (i.d. 0.062 in./o.d. 0.125 in.; 10 mm/100 g bw) containing 30 μg 17β estradiol-3-benzoate/mL safflower oil or oil alone, under isoflurane anesthesia (5%—induction; 2.5%—maintenance). On study days 1–5, testes-intact male rats were randomly injected sc with the aromatase inhibitor letrozole (Lz; 1 mg/kg bw) or the vehicle 0.9% saline (SAL) containing 5.0% Tween-30 and 5.0% ethanol (Kokras et al. 2018); this Lz dosage reduces circulating estradiol levels by more than 70% (Bhatnagar et al. 1993). On day 6, animals of each sex were injected sc at 09.00 h with neutral protamine Hagedorn insulin (INS; 10.0 U/kg bw) or sterile diluent (V), then sacrificed by microwave fixation (1.45 s; In Vivo Microwave Fixation System, 5kW; Stoelting Co. Wood Dale, IL) at the following time points (Fig. 1a): V-injected controls: time zero (n = 4 V- and n = 4 Lz-pretreated males; n = 4 estradiol- and n = 4 oil-implanted females); INS-injected animals: + 1.0 h (n = 4 V- and n = 4 Lz-pretreated males; n = 4 estradiol- and n = 4 oil-implanted females), + 3.5 h (n = 4 V- and n = 4 Lz-pretreated males; n = 4 estradiol- and n = 4 oil-implanted females), or + 7.0 h (n = 4 V- and n = 4 Lz-pretreated males; n = 4 estradiol- and n = 4 oil-implanted females). Each brain was snap-frozen in a liquid nitrogen-cooled isopentane and stored at − 80 °C. Plasma was stored at − 20 °C for glucose measurement.

Experimental design and ventromedial hypothalamic nucleus-ventrolateral part (VMNvl) micropunch dissection. As shown in a, groups of male were pretreated on study days 1–5 by serial once-per-day subcutaneous (sc) injection of vehicle (V) or letrozole (Lz), while ovariectomized (OVX) female rats were implanted sc with a silastic capsule containing estradiol or the vehicle safflower oil alone over the same time interval. On day 6, groups of male V- or Lz-pretreated rats and groups of female estradiol- or oil-implanted OVX animals were injected sc at 09.00 hr (time zero;to) with vehicle (V; sterile diluent; sacrificed at to) or insulin (INS; 10.0 U neutral protamine Hagedorn insulin/kg bw; sacrificed at 1.0, 3.5, or 7.0 h after injection). For each sex, final treatment groups consisted of n = 4 rats each. In b, the rectangle in the Panel A brain map (− 2.85 mm posterior to bregma) depicts mediobasal hypothalamus and is enlarged (Panel B) to illustrate the location of VMN in that region. The blue circle denotes positioning of a 0.50-mm diameter circular micropunch tool over the VMNvl for selective harvesting of this substructure. Abbreviations in Panel B: ARH arcuate hypothalamic nucleus (n.), DMHa,p anterior, posterior dorsomedial hypothalamic n., fx fornix, LHA lateral hypothalamic area, ME median eminence, PVi intermediate periventricular hypothalamic n., VMHc,dm,vl central,dorsomedial,ventormlateral ventromedial hypothalamic n., TU tuberal n., V3 third ventricle
VMN Tissue Micropunch Dissection and Western Blot Analysis:
Each brain was cut into consecutive 10-μm-thick frozen sections through the VMN over the distance of − 1.80 to − 3.3 mm posterior to bregma. For each animal, bilateral micropunches of VMNvl tissue were taken using a calibrated 0.5 mm hollow punch tool (prod. no. 57401; Stoelting Co., Kiel, WI; Fig. 1b) from sections cut at rostral (− 1.8 to − 2.3 mm), middle (− 2.3 to − 2.8), and caudal (− 2.8 to − 3.3 mm) VMNvl regions, and pooled within each neuroanatomical segment in lysis buffer (2.0% sodium dodecyl sulfate, 0.05M dithiothreitol, 10.0% glycerol, 1.0 mM EDTA (Ibrahim et al. 2019) for Western blot analysis. Accuracy of use of micropunch methodology for collection of distinctive hypothalamic loci of interest, including the VMN, has been validated by marker protein expression (Mandal et al. 2017, 2018). In each treatment group, tissue lysate aliquots from individual subjects were combined within rostral middle, and caudal VMNvl levels to create for each target protein, at minimum, triplicate sample pools for each target protein per region. Pooled tissue samples were separated in Bio-Rad TGX 10% stain-free gels (Bio-Rad, Hercules, CA); after electrophoresis, gels were UV light-activated (1 min) in a Bio-Rad ChemiDoc TM Touch Imaging System (Ibrahim et al. 2019) to quantify total protein content in each lane. After protein transblotting, 0.45-μm PVDF-Plus membranes (prod. no. 121639; Data Support Co. Panorama City, CA) were blocked with Tris-buffer saline containing 0.1% Tween-20 and 2.0% bovine serum albumin, then incubated between 36–42 h (4 °C) with primary rabbit polyclonal antisera raised against AMPKα1/2 (AMPK; prod. no. 2532S; 1:2000; Cell Signaling Technology, Inc. Danvers, MA), phosphoAMPKα1/2 (pAMPK; prod. no. 2535S; 1:2000; Cell Signaling Technol.), nNOS (prod. no. NBP1-39681, 1:2000; Novus Biologicals, LLC, Littleton, CO), SF1 (prod. no. PA5-41967, 1:2000; Invitrogen/ThermoFisherScientific, Carlsbad, CA), or GAD (prod. no. ABN904, 1:10000; Millipore Sigma, Burlington, MA). Membranes were then incubated with goat anti-rabbit horseradish peroxidase-labeled secondary antibodies (1:5,000; prod. no. NEF812001EA; PerkinElmer, Waltham, MA), followed by SuperSignal West Femto maximum sensitivity chemiluminescence substrate (prod. no. 34096; ThermoFisherScientific, Waltham, MA). Automated membrane buffer washes and blocking and antibody incubations were performed in a Freedom Rocker™ Blotbot. Protein band optical density (O.D.) measures were normalized to total in-lane protein using Image Lab™ 6.0.0 software (Bio-Rad). Precision plus protein molecular weight dual color standards (prod. no. 161-0374, Bio-Rad) were included in each Western blot analysis.
Statistics:
Mean normalized VMNvl segmental protein O.D. and plasma glucose measures were evaluated within each sex by two-way analysis of variance and Student-Newman-Keuls post-hoc test. Differences of p < 0.05 were considered significant. In each figure, statistical differences between specific pairs of treatment groups are denoted as follows: *p < 0.05; **p < 0.01; ***p < 0.001; ****p < 0.0001.
Results
Rostral, middle, and caudal VMNvl tissue was microdissected independently from Lz- or V-pretreated male (Fig. 2a–c) and estradiol- or oil-implanted OVX female (Fig. 2d–f) rats after INS (patterned bars) or V (solid bars) injection for Western blot analysis of total AMPKα1/2 protein expression. In males, Lz pretreatment elevated baseline AMPK expression in the middle, but not rostral or caudal VMNvl. INS injection significantly decreased rostral VMNvl AMPK levels at +1, 3.5, and 7 h post-injection in both V- (white bars) and Lz- (gray bars) pretreated animals (Fig. 2a; F(7,24) = 10.82; p < 0.001). In this sex, hypoglycemic augmentation of middle VMNvl AMPK content (Fig. 2b; F(7,16) = 10.50; p < 0.0001) at + 1 h was prevented by Lz pretreatment. In females, INS respectively increased or decreased AMPK protein expression in rostral (Fig. 2d; F(7,24) = 4.48; p = 0.001) or caudal (Fig. 2f; F = 4.58; p = 0.0023) VMNvl at single post-injection time points in estradiol- (gray bars), but not oil- (white bars) implanted rats. Data show that in each sex, hypoglycemia caused differential changes in total AMPK protein levels between distinct neuroanatomical VMNvl segments. Hypoglycemic patterns of VMNvl AMPK expression in male rats were estradiol-dependent or -independent, depending upon rostro-caudal segment.

Effects of sc INS onjection on ventromedial hypothalamic nucleus-ventrolateral part (VMNvl) 5′-AMP- activated protein kinase (AMPK) protein expression in V- or Lz-pretreated male and estradiol- or oil-implanted OVX female rats. VMNvl tissue was bilaterally micropunch-dissected over predetermined rostro-caudal levels from groups of male and female rats after sc vehicle (V; sterile diluent) or INS (10.0 U neutral protamine Hagedorn insulin/kg bw) injection. Data depict mean normalized rostral (left-hand column; a male, d female), middle (middle column; b male, e female), and caudal (right-hand column; c male, f female) VMNvl AMPK protein optical density (O.D.) measures ± S.E.M. for groups of male (top row) and female (bottom row) rats. Groups of male rats (n = 4/group) were pretreated by serial daily sc injection of V- (gray bars) or Lz (white bars) prior to sacrifice after V injection (solid bars) or at 1.0 (diagonal-striped bars), 3.5 (cross-hatched bars), or 7.0 (stippled bars) h after INS injection. Groups of OVX estradiol- (gray bars) or oil (white bars)-implanted female rats (n = 4/group) were sacrificed at identical time points after V or INS treatment. *p < 0.05; **p < 0.01; ***p < 0.001; ****p < 0.0001
Effects of hypoglycemia on VMNvl pAMPKα1/2 protein expression were examined in Lz- or V-injected male (Fig. 3a–c) and estradiol- or oil-implanted OVX female (Fig. 3d–f) rats. INS elevated rostral VMNvl pAMPK profiles in V-pretreated male (Fig. 3a; F(7,24) = 11.39; p < 0.001) and estradiol-implanted OVX female (Fig. 3d; F(7,24) = 8.28; p < 0.0001) at each post-injection time point, e.g., + 1, 3.5, and 7 h. In V-pretreated males, INS injection also augmented pAMPK content in the middle (+ 1 h; Fig. 3b; F(7,16) = 9.01; p < 0.0001) and caudal (+ 3.5 h; Fig. 3c; F = 37.91; p < 0.0001) VMNvl; these responses were respectively averted or unaffected by Lz pretreatment. Estradiol-, but not oil-implanted female rats exhibited up-regulated middle VMNvl pAMPK expression at + 1, 3.5, and 7 h; Fig. 3e; F(7,32) = 3.77; p = 0.004). OVX rats treated with estradiol or oil exhibited augmented pAMPK levels in caudal VMNvl at + 3.5 h; Fig. 3f; F = 9.70; p < 0.0001). Outcomes show that pAMPK expression was up-regulated over the duration of hypoglycemia in the rostral VMNvl in each sex as well as middle and caudal VMNvl segments in females, and that this protracted amplification of sensor activity is mitigated by experimental manipulation of systemic estradiol in the rostral VMNvl in both sexes as well as middle female VMN. Lz pretreatment elevated baseline pAMPK content in rostral and middle VMNvl in males rats, whereas oil-implanted OVX females exhibited higher pAMPK expression in each rostro-caudal level of the VMNvl compared with estradiol-treated animals.

VMNvl rostrocaudal segment-specific patterns of phospho-AMPK (pAMPK) expression after V- versus INS-injection to male and female rats. Data show mean normalized rostral (left-hand column; a male, d female), middle (middle column; b male, e female), and caudal (right-hand column; c male, f female) VMNvl pAMPK O.D. measures ± S.E.M. for groups of male (top row) and female (bottom row) rats. Groups of male rats (n = 4/group) were pretreated by serial daily sc injection of V- (gray bars) or Lz (white bars) prior to sacrifice after V injection (solid bars) or at 1.0 (diagonal-striped bars), 3.5 (cross-hatched bars), or 7.0 (stippled bars) h after INS injection. Groups of OVX estradiol- (gray bars) or oil (white bars)-implanted female rats (n = 4/group) were sacrificed at identical time points after V or INS treatment. *p < 0.05; **p < 0.01; ***p < 0.001; ****p < 0.0001
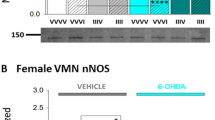
Figure 4 depicts effects of INS injection to Lz- versus V-pretreated male and estradiol- versus oil-implanted OVX female rats on regional patterns of nNOS protein expression in the VMNvl. V-pretreated male and estradiol-implanted OVX female rats exhibited up- [(rostral; male: Fig. 4a, F(7,24) = 6.58, p < 0.001; female: Fig. 4c, F(7,24) = 3.60; p = 0.004) (middle; male: Fig. 4b, F(7,16) = 12.99, p < 0.0001; female: Fig. 4e, F(7,32) = 7.21, p < 0.001)] or down- (caudal; male: Fig, 4c, F = 13.56, p < 0.0001; female: Fig. 4f, F = 10.77; p < 0.0001) regulated VMNvl nNOS protein profiles in response to INS injection. In males, Lz pretreatment prevented these nNOS responses in each VMNvl level, with the exception of the rostral segment. Compared with males, females exhibited comparatively more prolonged adjustments in nNOS expression due to hypoglycemia at each rostro-caudal level. Data show that in each sex, hypoglycemia causes segment-specific bi-directional, e.g., stimulatory versus inhibitory changes in nNOS expression, which are detectable over longer post-INS injection intervals in female versus male. Effects of hypoglycemia on this protein were estradiol-dependent throughout the female VMNvl, whereas in males, estradiol regulation of this protein response varies among rostro-caudal levels. Oil-implanted euglycemic controls female rats exhibited elevated middle VMNvl or diminished caudal VMNvl tissue nNOS levels compared with estradiol-implanted control group.

Effects of INS injection on VMNvl expression of neuronal nitric oxide synthase (nNOS), marker protein for the gluco-stimulatory transmitter nitric oxide, in V- or Lz-pretreated male and estradiol- or oil-implanted OVX female rats. Data show mean normalized rostral (left-hand column; a male, d female), middle (middle column; b male, e female), and caudal (right-hand column; c male, f female) VMNvl nNOS O.D. measures ± S.E.M. for groups of male (top row) and female (bottom row) rats. In each sex, bar colors differentiate pretreatments before sc V or INS injection; animals were sacrificed after V injection (solid bars) or at 1.0 (diagonal-striped bars), 3.5 (cross-hatched bars), or 7.0 (stippled bars) h after INS injection. *p < 0.05; **p < 0.01; ***p < 0.001; ****p < 0.0001
Effects of Lz- versus V-pretreatment of males and estradiol- versus oil-implantation of OVX female rats on VMNvl SF1 protein responses to hypoglycemia are shown in Fig. 5. V-pretreated hypoglycemic males exhibited decreased or elevated SF1 expression at rostral (Fig. 5a; F(7,24) = 20.50; p < 0.001) and middle (Fig. 5b; F(7,16) = 6.61; p = 0.001) versus caudal (Fig. 5c; F = 12.16; p < 0.0001) levels of the VMNvl, respectively. In male rats, Lz pretreatment blocked INS effects on SF1 profiles at all (rostral VMNvl) or a subset (middle and caudal VMNvl) of post-injection time points. Estradiol-, but not oil-implanted OVX females showed up-regulated SF1 protein expression in the rostral (Fig. 5d; F(7,24) = 7.78; p < 0.0001) and middle (Fig. 5e; F(7,24) = 8.02; p < 0.0001) VMNvl. Results indicate that male rats exhibit divergent changes in VMNvl SF expression over the rostro-caudal length of the VMNvl in response to hypoglycemia, whereas females show up-regulated protein profiles in rostral and middle segments only. Oil-implanted euglycemic controls female rats exhibited decreased middle VMNvl SF1 expression compared with estradiol-implanted control group.

Effects of INS injection on VMNvl expression of the gluco-stimulatory neurotransmitter steroidogenic factor-1 (SF1) in V- or Lz-pretreated male and estradiol- or oil-implanted OVX female rats. Data show mean normalized rostral (left-hand column; a male, d female), middle (middle column; b male, d female), and caudal (right-hand column; c male, f female) VMNvl SF1 O.D. values ± S.E.M. for male (top row) and female (bottom row) rat treatment groups. In each sex, bar colors indicate specific pretreatments ahead of sc V or INS injection; animals were sacrificed after V injection (solid bars) or at 1.0 (diagonal-striped bars), 3.5 (cross-hatched bars), or 7.0 (stippled bars) h after INS injection. *p < 0.05; **p < 0.01; ***p < 0.001; ****p < 0.0001
VMNvl regional GAD protein responses to INS injection in Lz- versus V-pretreated male and estradiol- versus oil-implanted female rats are presented in Fig. 6. Hypoglycemia inhibited GAD expression in the rostral VMNvl in male rats, irrespective of pretreatment (Fig. 6a; F(7,24) = 19.60; p < 0.001), but did not modify this protein profile in other VMNvl segments in this sex. In contrast, GAD levels were suppressed in hypoglycemic estradiol-, but not oil-implanted OVX females in both rostral (Fig. 6c; F(7,24) = 4.42; p = 0.001) and middle (Fig. 6e; F(7,32) = 3.61; p = 0.006) levels of the VMNvl. Results show that hypoglycemia inhibits GAD protein expression by estradiol-independent versus–dependent mechanisms in male and female rats, respectively, and that this negative response is limited to the rostral VMNvl in males versus both rostral and middle VMNvl in females. Baseline GAD protein expression was significantly elevated in Lz- versus V-pretreated euglycemic controls.

Effects of INS injection on VMNvl expression of Glutamate Decarboxylase65/67 (GAD), marker protein for the gluco-inhibitory neurotransmitter γ-aminobutyric acid (GABA), in V- or Lz-pretreated male and estradiol- or oil-implanted OVX female rats. Data show mean normalized rostral (left-hand column; a male, d female), middle (middle column; b male, d female), and caudal (right-hand column; c male, f female) VMNvl GAD O.D. measures ± S.E.M. for male (top row) and female (bottom row) rat treatment groups. In each sex, bar colors denote specific pretreatments prior to sc V or INS injection; animals were sacrificed after V injection (solid bars) or at 1.0 (diagonal-striped bars), 3.5 (cross-hatched bars), or 7.0 (stippled bars) h after INS injection. *p < 0.05; **p < 0.01; ***p < 0.001; ****p < 0.0001
Results in Fig, 7 show that plasma glucose concentrations were significantly decreased below control values in male F(7,42)= 25.68; p = 0.006) and female F(7,42)= 21.55; p < 0.0001) rats at both 1 and 3.5 h after INS injection. In both sexes, circulating glucose levels were not different between V controls and animals sacrificed 7 h after INS injection. Post-INS plasma glycemic profiles were equivalent in Lz- versus V-pretreated male and in estradiol- versus oil-implanted OVX females.

Effects of INS injection on plasma glucose concentrations in V- or Lz-pretreated male and eestradiol- or oil-implanted OVX female rats. Figures 7 a and b depicts mean circulating glucose levels in male and female rats, respectively. As depicted in a, groups of male rats (n = 4/group) were pretreated by serial daily sc injection of V- (white bars) or Lz (gray bars), as indicated at the x axis, before V (solid bars) or INS (patterned bars) injection. INS-injected animals were sacrificed at 1.0 (diagonal-striped bars), 3.5 (cross-hatched bars), or 7.0 (stippled bars) h after treatment. Figure 7 b shows data from groups of OVX estradiol- (E; gray bars) or oil (O; white bars)-implanted female rats (n = 4/group) were sacrificed at identical time points after V or INS treatment. *p < 0.05 compared with V-injected controls; **p < 0.05 compared with V-pretreated, V-injected male rats
Discussion
The VMNvl is an estradiol-sensitive structure that is a plausible source of metabolic-sensory input to the neural gluco-regulatory network. Herein, high neuroanatomical-resolution microdissection tools were used to address the premise that VMNvl energy sensor and metabolic transmitter marker protein responses to hypoglycemia are sex-dimorphic owing to estradiol regulation. In male rats, hypoglycemic patterns of total AMPK expression involved estradiol-independent or -dependent mechanisms, depending upon rostrocaudal level, whereas the protein response in females required estradiol signaling. pAMPK profiles were up-regulated in each VMNvl segment during as well as after hypoglycemia in female rats, whereas this temporal pattern occurred only in rostral VMNvl in male rats. Results provide novel evidence for region-specific stimulatory (rostral and middle VMNvl) or inhibitory (caudal VMNvl) nNOS protein responses to hypoglycemia in each sex. Male and female hypoglycemic rats exhibited divergent adjustments in SF1 profiles at each VMNvl level, as expression was decreased (males) or increased (females) in rostral and middle VMNvl, but enhanced (male) or unchanged (female) in the caudal segment. Present outcomes show that hypoglycemic down-regulation of GAD expression in males is restricted to the rostral VMNvl, but occurs in both rostral and middle segments in the female. Interestingly, neurotransmitter protein responses to hypoglycemia were estradiol-dependent in each segment, excepting the male rostral VMNvl. Present outcomes provide an invaluable neuroanatomical map that can expedite investigation of the respective role of functionally heterogenous subpopulations of distinctive metabolic neurotransmitter cell types in the VMNvl in neural regulation of glucostasis in each sex.
Hypoglycemic male rats exhibited a reduction in total AMPK protein expression in the rostral VMNvl, yet this protein profile remained unchanged in other segments; the implications of this regional inhibitory response are clear. A possible physiological consequence of down-regulated AMPK profiles is augmentation of the cellular pAMPK/AMPK ratio, i.e., enzyme specific activity. Alternatively, decreased AMPK availability could, depending upon extent of decline, eventually limit enzyme mass available for activation by phosphorylation. Phosphorylation is a rapid post-translational modification that generates an appropriate acute response to hypoglycemia, whereas decrements in total AMPK protein levels could conceivably serve as a more protracted adaptive response. This response in the male is evidently unassociated with estradiol action as it occurred in Lz- as well as V-pretreated animals. The rostral VMNvl is typified by lengthy, estradiol-dependent hypoglycemic up-regulation of pAMPK expression in each sex; this protein was also increased over a similar temporal pattern in middle and caudal VMNvl segments in the female. A plausible explanation is that sensors functioning in these rostro-caudal segments may, in part, monitor the time-course of hypoglycemia per se as opposed to magnitude of cellular-level sequelae of diminished circulating glucose. The unique observation here that pAMPK expression persisted in those locations despite recovery from hypoglycemia may indicate either that neuronal energy stability is not immediately restored, or alternatively, that up-regulation of this protein could reflect afferent neurochemical regulation or depletion of brain energy reserve levels rather than cellular metabolic balance. It should be noted that averaged values for protein expression within a tissue sample mass, as reported here, may likely obscure differences in magnitude and/or direction of change in AMPK activity between individual neurotransmitter cell types present in that substructure. Previous studies showed that VMN nNOS, SF1, and GAD neurons express hypoglycemia-sensitive AMPK (Ibrahim et al. 2020). Here, correlated longitudinal patterns of expression of pAMPK versus SF1 in the rostral VMNvl imply that this transmitter population may be responsive to cellular energy imbalance during and after hypoglycemia. A similar point can be made for extended changes in marker protein expression in the female middle (nNOS, SF1, GAD) and caudal (GAD) VMNvl. Additional work is warranted to determine, for each nerve cell type, whether pAMPK regulates transmitter marker protein expression over all or part of the 7-h time frame investigated here. Hypoglycemic up-regulation of pAMPK expression in rostral and middle VMNvl was totally prevented by Lz treatment of males or OVX of females, which infers that sensor activation by this metabolic stress is subject to continuous control by estradiol-dependent mechanisms. In each sex, up-regulated pAMPK profiles in the caudal VMNvl coincide with diminished nNOS content. However, it may be unlikely that these protein responses are linked by causal association, as nNOS expression is a surrogate for gluco-stimulatory nitric oxide transmission, which is elevated in response to energy deficiency. Current evidence that this signal is suppressed in the caudal VMNvl suggests that positive energy balance may be maintained in this particular segment despite elevated pAMPK expression.
Present studies provide novel proof that in each sex, hypoglycemia elicits bi-directional, segment-specific changes in VMNvl nNOS expression. In male and female, this protein profile is up-regulated in rostral and middle VMNvl, but down-regulated in the caudal segment. In females, nNOS levels were consistently elevated (middle VMNvl) or decreased (caudal VMNvl) between 1 and 7 h after INS treatment; males exhibited similar changes in this protein profile in each segment, but at fewer time points. Attenuation of these patterns of response in oil-implanted OVX females or Lz-injected males infers that they are controlled by estradiol. Notably, in the female, middle and caudal VMNvl nNOS protein levels remained elevated or decreased, respectively, at 7 h post-injection, despite restoration of euglycemia. It remains to be determined if continued changes in gluco-stimulatory NO transmission in these segments reflect lingering effects of hypoglycemia on metabolic balance in these neurons, or, rather, persistent input by hormonal or neurochemical stimuli. Critical questions that emerge from current work concern the role of middle versus caudal VMNvl nitrergic neurons in each sex in glucostasis, the identity of ER(s) that mediate estradiol control of hypoglycemic patterns of nNOS expression in distinct rostro-caudal segments, and clarification of hypoglycemia-associated metabolic cues that trigger segment-specific nNOS responses.
While previous studies showed that nNOS content in VMN tissue samples encompassing dorsomedial, central, and ventrolateral parts is increased during hypoglycemia (Napit et al. 2019; Ali et al. 2019), current outcomes evidently identify neuroanatomically distinct populations of hypoglycemia-stimulated versus hypoglycemia-inhibited nitrergic neurons in the VMNvl, It would be informative to learn if and how afferent and efferent connectivity of nitrergic neurons may vary among rostro-caudal VMNvl regions in each sex, including input, via polysynaptic pathways, by AMPK-expressing elements of cell type to CNS gluco-regulatory motor structures. However, microinjection tools capable of accurate, selective high-spatial resolution anterograde/retrograde tracer delivery to the VMNvl are not currently available. The catecholamine neurotransmitter norepinephrine (NE) conveys information from hindbrain metabolic sensors to the hypothalamus (Briski and Patil 2005; Briski and Mandal 2019, 2020), and regulates VMN nNOS expression in the male and female VMN (Ibrahim et al. 2019; Mahmood et al. 2019). Nitrergic neurons obtained by laser-catapult microdissection over the rostro-caudal length of the VMN, without regard for substructure boundaries, express ERα, ERβ, and G-protein coupled ER-1 (GPER/GPR30) proteins and several adrenergic receptor (AR) variants (Uddin et al. 2019). That work showed that ERα and –β impose differentially regulate patterns of GPER and β1-AR protein expression during hypoglycemia, and that adjustments in nitrergic nerve cell ER and AR expression may render these neurons more sensitive to both estradiol and NE in males, but more or less sensitive to estradiol and NE, respectively, in females. Extrapolation of those findings to the present work is not feasible. Further effort is thus warranted to characterize, in each sex, ER regulation of NO nerve cell receptivity to NE and other gluco-regulatory stimuli over the rostro-caudal length of the VMNvl. Supply of lactate, the oxidizable, trafficable end-product of astrocyte glycolysis, is a critical measure of VMN metabolic balance, as exogenous lactate infusion to the mediobasal hypothalamus attenuates counter-regulatory hormone responses to hypoglycemia (Borg et al. 2003). It remains unclear, for either sex, if this monitored metabolite affects VMNvl nitrergic neurotransmission over the entire or partial length of the VMN. There is also a need for insight on whether hypoglycemic patterns of lactate release by astrocytes and/or NO nerve cell monocarboxylate transporter expression differs between male and female in segments where lactate-sensitive nitrergic neurons reside.
Present findings show that hypoglycemic male and female rats exhibited opposite adjustments in SF1 protein expression in the rostral and middle VMNvl, as this protein profile was diminished in both regions in males, but was instead amplified in those same sites in the female. In the same animals, caudal VMNvl SF1 protein levels were up-regulated in INS-injected males, but were unaffected by INS administration in females. Outcomes here support the notion that gluco-stimulatory SF1 neurons may reside in different VMNvl segments in male versus female. Since caudal VMNvl pAMPK levels were correspondingly refractory to or elevated by hypoglycemia in male and female, local SF1 responses to this metabolic stress may not reflect metabolic status. In males, it is unclear if down-regulated neurotransmitter expression in the rostral and middle VMNvl points to the existence of a distinctive hypoglycemia-inhibited SF1 neuron population (which lacks gluco-stimulatory functionality), or alternatively, reflects a state of enhanced energy stability in those regions, despite increased pAMPK expression. Notably, in each sex, hypoglycemia-associated patterns of VMNvl SF1 expression largely persisted after normal plasma glucose levels were regained. These findings support the need for insight on the metabolic cues and neurochemical inputs that shape SF1 signaling during and after exposure to hypoglycemia. Present data also show that hypoglycemic regulation of SF1 expression primarily involves estradiol-independent mechanisms in the male, but involves estradiol in the female.
Current results show that hypoglycemia was exclusively inhibitory to VMNvl GAD expression, in contrast to bi-directional impacts on nNOS and SF1, as discussed above, and suppression of GAD occurred only in the rostral VMNvl in males, but was observed more broadly, e.g., in both rostral and middle VMNvl in the female. These findings infer that the origin of gluco-inhibitory GABAergic signaling in the VMNvl is not identical in each sex. In the rostral VMNvl, time points corresponding to reductions in GAD protein varied between hypoglycemic males versus females; therefore, GABAergic neurons in that site may respond to a different array of cues over time according to sex. Hypoglycemic regulation of VMNvl GAD expression was predominantly estradiol-independent in males, but estradiol-reliant in females. NE regulates VMN GAD expression in the male and female VMN (Ibrahim et al. 2019; Mahmood et al. 2019). VMN GABA neurons collected without discrimination for substructure boundaries exhibit sex-dimorphic ERα and ERβ – dependent alpha1-AR, alpha2-AR, and ERα protein expression during hypoglycemia (Uddin et al. 2019). Those studies suggest that sex-specific effects of hypoglycemia on GABA nerve cell ER and AR protein expression may likely diminish sensitivity of these cells to both estradiol and NE profiles in males, but may cause divergent, e.g., up- versus down-regulated receptivity to estradiol and NE, respectively, in the female. Additional studies are needed to determine if and how metabolic cues, e.g., substrate fuels and glucose-regulatory hormones, estradiol, and gluco-regulatory neurochemical inputs affect GABA transmission in the rostral VMNvl in males versus rostral and middle segments in the female, and to determine how those stimuli impact functionality of downstream brain gluco-regulatory network elements.
Table 1 provides a summary of effects of hypoglycemia on target proteins across rostro-caudal VMNvl segments in male and female rats. Data provide unique evidence for protracted AMPK activation, e.g., after recovery from hypoglycemia, in this structure, and show that this activity pattern occurs only in the rostral VMNvl segment in males, but over the length of the VMNvl in female. The rostral VMNvl is characterized by estradiol-dependent up-regulation of pAMPK, but estradiol-independent suppression of total AMPK protein in males. Sex differences in the ratio of activated versus total AMPK (and resultant effects on enzyme specific activity) in individual gluco-regulatory neuron populations investigated here may be a factor in divergent hypoglycemic patterns of marker protein expression. Up-regulation of nNOS at fewer time points in the male as well as a decline in gluco-stimulatory SF1 signaling in that sex may reflect a more positive energy state in males, despite augmented pAMPK expression. Identification of the rostral VMNvl as the sole location where GAD expression is down-regulated in hypoglycemic males suggests Down-regulated GAD profiles in the rostral VMNvl In middle and caudal VMNvl levels, AMPK activation was detected at a single time point in hypoglycemic males, but occurred throughout as well as after hypoglycemia in females. These temporal patterns of sensor activation correlate closely with transmitter protein expression in these regions. Unique among VMNvl segments, the caudal level is characterized by an apparent lack of association between hypoglycemic patterns of AMPK activation and metabolic transmitter protein expression.
In conclusion, new evidence described here for functional heterogeneity of VMNvl metabolic neurotransmitter cell types underscores the need for high-neuroanatomical specificity in future work for accurate discernment of the role of these neurochemicals in monitoring cellular energy stability during and after recovery from hypoglycemia, and their involvement in neural control of counter-regulatory outflow during this metabolic stress . The VMNvl is implicated in the control of several vital reproductive behaviors. Neural command of reproduction and metabolism is tightly linked, plausibly by neurotransmitters that signal within interconnected pathways that impose synchronized adjustments in these functions in response to loss of neuro-metabolic stability. Further research is needed to investigate whether hypoglycemia-sensitive VMNvl neurotransmitters identified here provide such a functional connection between metabolic and reproduction-relevant pathways.
Abbreviations
- AMPK:
-
5′-AMP-activated protein kinase
- ER:
-
Estrogen receptor
- ERα:
-
Estrogen receptor-alpha
- ERβ:
-
Estrogen receptor-beta
- GABA:
-
γ-Aminobutyric acid
- GAD:
-
Glutamate decarboxylase65/67
- INS:
-
Insulin
- IIH:
-
Insulin-induced hypoglycemia
- LZ:
-
Letrozole
- nNOS:
-
Neuronal nitric oxide synthase
- OVX:
-
Ovariectomy
- pAMPK:
-
PhosphoAMPK
- SF1:
-
Steroidogenic factor-1
- VMN:
-
Ventromedial hypothalamic nucleus
- VMNvl:
-
Ventromedial hypothalamic nucleus–ventrolateral part
References
Adachi A, Kobashi M, Funahashi M (1997) Glucose sensors in the medulla oblongata. Jpn. J. Physiol. 47(Suppl 1):S37–S38
Adachi A, Kobashi M, Funahashi M (1995) Glucose-responsive neurons in the brainstem. Obes. Res. 3(Suppl 5):735S-740S
Ali MH, Napit PR, Mahmood ASMH, Bheemanapally K, Alhamami HN, Uddin MM, Mandal SK, Ibrahim MMH, Briski KP (2019) Hindbrain estrogen receptor regulation of ventromedial hypothalamic glycogen metabolism and glucoregulatory transmitter expression in the hypoglycemic male rat. Neuroscience 409:253–260
Ashford MLJ, Boden PR, Treherne JM (1990) Glucose-induced excitation of hypothalamic neurons is mediated by ATP-sensitive K+ channels. Pfugers Arch. 415:479–483
Bhatnagar AS, Batzl C, Häusler A, Nogués V (1993) The role of estrogen in the feedback regulation of follicle-stimulating hormone secretion in the female rat. J. Steroid Biochem. Mol. Biol. 47:161–166
Borg MA, Tamborlane WV, Shulman GI, Sherwin RS (2003) Local lactate perfusion of the ventromedial hypothalamus suppresses hypoglycemic counterregulation. Diabetes 52:663–666
Briski KP, Mandal SK (2019) Hindbrain lactoprivic regulation of hypothalamic neuron transactivation and glucoregulatory neurotransmitter expression: Impact of antecedent insulin-induced hypoglycemia. Neuropeptides. https://doi.org/10.1016/j.npep.2019.101962
Briski KP, Mandal SK (2020) Hindbrain metabolic deficiency regulates ventromedial hypothalamic nucleus glycogen metabolism and glucose-regulatory signaling. Acta Neurobiol. Exper. 80:57–65
Briski KP, Patil GD (2005) Induction of Fos immunoreactivity labeling in forebrain metabolic loci by caudal fourth ventricular administration of the monocarboxyate transporter inhibitor, α-cyano-4-hydroxycinnamic acid. Neuroendocrinology 82:49–57
Briski KP, Marshall ES, Sylvester PW (2001) Effects of estradiol on glucoprivic transactivation of catecholaminergic neurons in the female rat caudal hindbrain. Neuroendocrinology 73: 369-377
Briski KP, Mandal SK, Bheemanapally K, Ibrahim MMH (2020) Effects of acute versus recurrent insulin-induced hypoglycemia on ventromedial hypothalamic nucleus metabolic-sensory neuron AMPK activity: impact of alpha1-adrenergic receptor signaling. Brain Res. Bull. 157:41–50
Chan O, Zhu W, Ding Y, McCrimmon RJ, Sherwin RS (2006) Blockade of GABA(A) receptors in the ventromedial hypothalamus further stimulates glucagon and sympathoadrenal but not the hypothalamo-pituitary-adrenal response to hypoglycemia. Diabetes 55:1080–1087
Correa SM, Newstrom DW, Warne JP, Flandin P, Cheung CC, Lin-Moore AT, Pierce AA, Xu AW, Rubenstein JL, Ingraham HA (2015) An estrogen-responsive module in the ventromedial hypothalamus selectively drives sex-specific activity in females. Cell Rep. 10:62–74
Cotero VE, Routh VH (2009) Insulin blunts the response of glucose-excited neurons in the ventrolateral-ventromedial hypothalamic nucleus to decreased glucose. Amer. J. Physiol. Endocrinol. Metab. 296:E1101–E1109
Fioramonti X, Marsollier N, Song Z, Fakira KA, Patel RM, Brown S, Duparc T, Pica-Mendez A, Sanders NM, Knauf C, Valet P, McCrimmon RJ, Beuve A, Magnan C, Routh VH (2010) Ventromedial hypothalamic nitric oxide production is necessary for hypoglycemia detection and counterregulation. Diabetes 59:519–528
Garfield AS, Shah BP, Madara JC, Burke LK, Patterson CM, Flak J, Neve RL, Evans ML, Lowell BB, Myers MG Jr, Heisler LK (2014) A parabrachial-hypothalamic cholecystokinin neurocircuit controls counterregulatory responses to hypoglycemia. Cell Metab. 20:1030–1037
Han SM, Namkoong C, Jang PG, Park IS, Hong SW, Katakami H, Chun S, Kim SW, Park JY, Lee KU, Kim MS (2005) Hypothalamic AMP-activated protein kinase mediates counter-regulatory responses to hypoglycaemia in rats. Diabetologia 48:2170–2178
Hardie DG, Schaffer BE, Brunet A (2016) AMPK: an energy-sensing pathway with multiple inputs and outputs. Trends Cell Biol. 26:190–201
Hashikawa K, Hashikawa Y, Tremblay R, Zhang J, Feng JE, Sabol A, Piper WT, Lee H, Rudy B, Lin D (2017) Esr1 + cells in the ventromedial hypothalamus control female aggression. Nat Neurosci. 20:1580–1590
Ibrahim MMH, Bheemanapally K, Alhamami HN, Briski KP (2020) Effects of intracerebroventricular glycogen phosphorylase inhibitor CP-316,819 infusion on hypothalamic glycogen content and metabolic neuron AMPK activity and neurotransmitter expression in the male rat. J. Mol. Neurosci. 70:647–658
Ibrahim MMH, Alhamami HN, Briski KP (2019) Norepinephrine regulation of ventromedial hypothalamic nucleus metabolic transmitter biomarker and astrocyte enzyme and receptor expression: impact of 5’-AMP-activated protein kinase. Brain Research 1711:48–57
Kokras N, Pastromas N, Papasava D, de Bournonville C. Cornil CA, Dalla C (2018) Sex differences in behavioral and neurochemical effects of gonadectomy and aromatase inhibition in rats. Psychoneuroendcrinol. 87: 93-107
Lin SC, Hardie DG (2018) AMPK: sensing glucose as well as cellular energy status. Cell Metab. 27:299–313
Liu J, Bisschop PH, Eggels L, Foppen E, Ackermans MT, Zhou JN, Fliers E, Kalsbeek A (2013) Intrahypothalamic estradiol regulates glucose metabolism via the sympathetic nervous system in female rats. Diabetes 62:435–443
López M (2018) Hypothalamic AMPK and energy balance. Eur J Clin Invest. 48:e12996
Mahmood ASMH, Bheemanapally K, Mandal SK, Ibrahim MMH, Briski KP (2019) Norepinephrine control of ventromedial hypothalamic nucleus glucoregulatory neurotransmitter expression in the female rat: role monocarboxylate transporter function. Mol. Cell. Neurosci. 95:51–58
Mahmood ASMH, Uddin MM, Mandal SK, Ibrahim MMH, Alhamami HN, Briski KP (2018) Sex differences in forebrain estrogen receptor regulation of hypoglycemic patterns of counter-regulatory hormone secretion and ventromedial hypothalamic nucleus gluco-regulatory neurotransmitter and astrocyte glycogen metabolic enzyme expression. Neuropeptides 72:65–74
Mandal SK, Shrestha PK, Alenazi FSH, Shakya M, Alhamami HN, Briski KP (2017) Role of hindbrain adenosine 5’-monophosphate-activated protein kinase (AMPK) in hypothalamic AMPK and metabolic neuropeptide adaptation to recurring insulin-induced hypoglycemia in the male rat. Neuropeptides 66:25–35
Mandal SK, Shrestha PK, Alenazi FSH, Shakya M, Alhamami HN, Briski KP (2018) Effects of estradiol on lactoprivic signaling of the hindbrain upon the contraregulatory hormonal response and metabolic neuropeptide synthesis in hypoglycemic female rats. Neuropeptides 70(70):37–46
Martínez de Morentin PB, González-García I, Martins L, Lage R, Fernández-Mallo D, Martínez-Sánchez N, Ruíz-Pino F, Liu J, Morgan DA, Pinilla L, Gallego R, Saha AK, Kalsbeek A, Fliers E, Bisschop PH, Diéguez C, Nogueiras R, Rahmouni K, Tena-Sempere M, López M (2014) Estradiol regulates brown adipose tissue thermogenesis via hypothalamic AMPK. Cell Metab. 20: 41-53
McCrimmon RJ, Shaw M, Fan X, Cheng H, Ding Y, Vella MC, Zhou L, McNay EC, Sherwin RS (2008) Key role for AMP-activated protein kinase in the ventromedial hypothalamus in regulating counterregulatory hormone responses to acute hypoglycemia. Diabetes 57:444–450
Meek TH, Nelson JT, Matsen ME, Dorfman MD, Guyenet SJ, Damian V, Allison MB, Scarlett JM, Nguyen HT, Thaler JP, Olson DP, Myers MG Jr, Schwartz MW, Morton GJ (2016) Functional identification of a neurocircuity regulating blood glucose. Proc. Natl. Acad. Sci. 113:E2073–E2082
Musatov S, Chen W, Pfaff DW, Mobbs CV, Yang XJ, Clegg DJ, Kaplitt MG, Ogawa S (2007) Silencing of estrogen receptor alpha in the ventromedial nucleus of hypothalamus leads to metabolic syndrome. Proc. Natl. Acad. Sci. 104:2501–2506
Napit PR, Ali MH, Shakya M, Mandal SK, Bheemanapally K, Mahmood ASMH, Ibrahim MMH, Briski KP (2019) Hindbrain estrogen receptor regulation of ventromedial hypothalamic glycogen metabolism and glucoregulatory transmitter expression in the hypoglycemic female rat. Neuroscience 411:211–221
Nedungadi TP, Briski KP (2012) Site-specific effects of intracranial estradiol administration on recurrent insulin-induced hypoglycemia in ovariectomized female rats. Neuroendocrinology 96:311–323
Oomura Y, Ono H, Ooyama H, Wayner MJ (1969) Glucose and osmosensitive neurons of the rat hypothalamus. Nature 222:282–284
Pimentel GD, Ropelle ER, Rocha GZ, Carvalheira JBC (2013) The role of neuronal AMPK as a mediator of nutritional regulation of food intake and energy homeostasis. Metabolism 62:171–178
Routh VH, Hao L, Santiago AM, Sheng Z, Zhou C (2014) Hypothalamic glucose sensing: making ends meet. Front. Syst. Neurosci. 8:236. https://doi.org/10.3389/fnsys.2014.00236
Santiago AM, Clegg DJ, Routh VH (2016) Estrogens modulate ventrolateral ventromedial hypothalamic glucose-inhibited neurons. Mol. Metab. 5:823–833
Silver IA, Erecińska M (1998) Glucose-induced intracellular ion changes in sugar-sensitive hypothalamic neurons. J. Neurophysiol. 79:1733–1745
Simerly, RB (2002) Wired for reproduction: organization and development of sexually dimorphic circuits in the mammalian forebrain. Annu Rev Neurosci. 25 507–536
Uddin MM, Mahmood ASMH, Ibrahim MMH, Briski, KP (2019) Sex dimorphic estrogen receptor regulation of ventromedial hypothalamic nucleus glucoregulatory neuron adrenergic receptor expression in hypoglycemic male and female rats. Brain Res. https://doi.org/10.1016/j.brainres.2019.146311
Xue B, Kahn BB (2006) AMPK integrates nutrient and hormonal signals to regulate food intake and energy balance through effects in the hypothalamus and peripheral tissues. J. Physiol. 574:73–83
Yang CF, Chiang MC, Gray DC, Prabhakaran M, Alvarado M, Juntti SA, Unger EK, Wells JA, Shah NM (2013) Sexually dimorphic neurons in the ventromedial hypothalamus govern mating in both sexes and aggression in males. Cell 153:896–909
Funding
NIH DK-109382
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of Interest
The authors declare that they have no conflict of interest.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
About this article

Cite this article
Alshamrani, A.A., Bheemanapally, K., Alhamyani, A. et al. Sex-dimorphic Rostro-caudal Patterns of 5′-AMP-activated Protein Kinase Activation and Glucoregulatory Transmitter Marker Protein Expression in the Ventrolateral Ventromedial Hypothalamic Nucleus (VMNvl) in Hypoglycemic Male and Female Rats: Impact of Estradiol. J Mol Neurosci 71, 1082–1094 (2021). https://doi.org/10.1007/s12031-020-01730-5
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s12031-020-01730-5