
Abstract
The objective of this study was to determine how different sources of Zn, Mn, and Cu in the feed without and with phytase affect prececal myo-inositol hexakisphosphate (InsP6) breakdown to myo-inositol (MI), prececal P digestibility, bone mineralization, and expression of mineral transporters in the jejunum of broiler chickens. A total of 896 male broiler chicks (Cobb 500) were distributed to 7 diets with 8 replicate pens (16 birds per floor pen). Experimental diets were fed from day 0 to 28. Diets were without or with phytase supplementation (0 or 750 FTU/kg) and were supplemented with three different trace mineral sources (TMS: sulfates, oxides, or chelates) containing 100 mg/kg Zn, 100 mg/kg Mn, and 125 mg/kg Cu. Prececal InsP6 disappearance and P digestibility were affected by interaction (phytase × TMS: P ≤ 0.010). In diets without phytase supplementation, prececal InsP6 disappearance and P digestibility were greater (P ≤ 0.001) in birds fed chelated minerals than in birds fed sulfates or oxides. However, no differences were observed between TMS in diets with phytase supplementation. Ileal MI concentration was increased by exogenous phytase but differed depending on TMS (phytase × TMS: P ≤ 0.050). Tibia ash concentration as well as Zn and Mn concentration in tibia ash were increased by phytase supplementation (P < 0.010), but the Cu concentration in tibia ash was not (P > 0.050). Gene expression of the assayed mineral transporters in the jejunum was not affected by diet (P > 0.050), except for Zn transporter 5 (phytase × TMS: P = 0.024). In conclusion, the tested TMS had minor effects on endogenous phytate degradation in the digestive tract of broiler chickens. However, in phytase-supplemented diets, the choice of TMS was not relevant to phytate degradation under the conditions of this study.
Similar content being viewed by others

Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
Trace minerals (TM) such as Zn, Mn, and Cu are essential in animal nutrition due to their involvement in hormone and enzymatic systems. They are also components of proteins and play a role in the immune system [1]. Consequently, TM impact growth, bone development, and enzyme structures, among other things. If their supply is below the requirement, adverse effects on bone development and growth can be observed [1, 2]. Because plant-based feedstuffs commonly used in broiler diets do not contain adequate concentrations of available Zn, Mn, and Cu for the bird, their diets are usually supplemented with exogenous sources of those TM. According to the National Research Council [3], supply recommendations for broilers are 40 mg/kg Zn, 60 mg/kg Mn, and 8 mg/kg Cu in complete diets. However, TM concentrations used by the industry are often higher because they are used to promote growth performance, as in the case of Cu [4], or because they are supplemented with large safety margins to account for dietary antagonists such as phytate. However, the oversupply of trace minerals leads to elevated concentrations of trace minerals in the manure and is potentially harmful to the environment.
Phytate is the salt form of phytic acid (myo-inositol 1,2,3,4,5,6-hexakisphosphate (InsP6)) and the primary P source in feedstuffs derived from plant seeds. In non-ruminant animals, the phytate-P availability is limited and varies among species [5]. Phosphatases and phytases (myo-inositol hexaphosphate phosphohydrolases) are required for the dephosphorylation of phytate. Although broilers can degrade relevant amounts of phytate from diets with low mineral P and Ca contents by endogenous mucosal and microbial phosphatases [6], industry-type poultry diets are commonly supplemented with exogenous phytases to increase the digestibility of phytate-P further.
In vitro studies have shown that divalent cations can inhibit the phosphate release from InsP6 by exogenous phytases depending on the concentration and source of (trace) minerals [7, 8]. The inhibition could be due to reduced accessibility of the phytate molecule, because divalent cations such as Zn, Mn, and Cu can form insoluble complexes with phytate [9, 10]. The affinity of cations to form complexes with phytate was ranked as follows: Cu2+ > Zn2+ > Co2+ > Mn2+ > Fe3+ > Ca2+ [11]. Different trace mineral sources (TMS) can differ in their dissolution behavior, which might affect their potential to interact with phytate, with faster dissolving sources having a greater potential to interact than slower dissolving sources [12]. Indeed, it was observed in vitro that Cu sulfate reduced phytate solubility and phosphate release to a greater extent than Cu oxide, which may be due to the faster dissolution of Cu sulfate as opposed to Cu oxide [4]. Chelated TMS may have a lower interaction potential than inorganic sources because chelated TM are bound to a ligand that may prevent the mineral from interactions with other nutrients such as phytate. Therefore, sulfates, oxides, and chelates were selected as TMS in the present study and analyzed for their interaction potential with phytate. In order to investigate potential interactions, InsP6 and its degradation products were analyzed and InsP6 disappearance was calculated.
The first aim of this study was to investigate how different sources of Zn, Mn, and Cu, at levels commonly used by the industry, affect prececal phytate degradation, ileal concentration of partially dephosphorylated inositol phosphates (InsPx) and myo-inositol (MI), and prececal P digestibility. The second aim was to investigate whether TMS and exogenous phytase affect trace mineral concentration in the bone, expression of mineral transporters in the jejunum, and expression of genes related to protein synthesis and degradation in the breast muscle and the liver. We hypothesized that first, the impairment of phytate degradation by Zn, Mn, and Cu supplementation depends on TMS, and second, phytase supplementation affects Zn, Mn, and Cu concentration in the bone.
Materials and Methods
Birds and Housing
The trial was conducted at the Poultry Science Research Complex of the University of Georgia (Athens, GA, USA) following the approved protocol (IACUC number: A2021-06–006) of the Institutional Animal Care and Use Committee of the University of Georgia. A total of 896 day-old male broilers (Cobb 500) were used in a 28-day experiment. Birds were allocated to 56 floor pens (16 birds per pen) with wood shavings. Even weight distribution across all treatments and blocks was ensured when placing the birds on day 0. Seven treatments were arranged in a randomized complete block design with 8 replicate pens per treatment. Throughout the trial, tap water and feed were provided for ad libitum consumption. The barn was continuously illuminated for the first 3 days; from day 3, the period of darkness was gradually increased to 8 h per day. The room temperature was continuously decreased from 34 to 22°C. Health status of the birds was monitored daily.
Experimental Diets
Experimental diets were corn-soybean meal based (Table 1) and were fed in 2 phases, starter and grower. The starter diets were fed in mash form (day 0 to 12), and the grower diets were fed as pellets (3 mm diameter, day 12 to 28). Diets were formulated to meet or exceed all nutrient requirements according to the National Research Council [3] except non-phytate P (Table 3). In the grower diets, titanium dioxide was added as an indigestible marker (5 g/kg). In a 2 × 3 + 1 factorial arrangement, 7 experimental diets were prepared. The experimental factors were phytase supplementation (0 (PHY–) or 750 FTU/kg (PHY+) declared activity of an E. coli–derived 6-phytase (Quantum®Blue, AB Vista, UK) and TMS. The TMS was a combination of Zn, Mn, and Cu (100, 100, 125 mg/kg feed) supplemented either as hydrated sulfates (Eastern Minerals, Inc. Henderson, NC, USA), oxides (Animine, France), or chelates (Availa® (Availa Zn, 12%; Availa Mn, 8%; Availa Cu, 10%), Zinpro, MN, USA). Levels of Zn and Mn were chosen according to the recommendation of the breeding company [13]. The high levels of Cu were chosen to represent Cu levels used in non-European countries to promote growth performance [4]. A phytase-supplemented diet containing supplemented sulfates to reach the National Research Council [3] recommended levels of trace minerals (15 mg/kg Zn, 40 mg/kg Mn, 7.5 mg/kg Cu) was used as an additional treatment to test the effect of trace mineral concentration. The chosen phytase supplementation level of 750 FTU/kg allowed for a clear phytase effect while staying below the possible maximum InsP6 degradation. To prepare the starter diets, all non-variable ingredients were mixed to obtain a basal diet. The starter basal diet was divided into 7 parts and supplemented with 4 g/kg of the corresponding trace mineral premix (Table 2) and quantity of phytase. For the grower diets, a premix with all non-variable mineral ingredients, vitamins, and free amino acids was prepared and split into 7 parts. Corn, soybean meal, soybean oil, and premix were supplemented with 4 g/kg of the corresponding trace mineral premix (Table 2) and quantity of phytase. Since TM premixes differed in Ca concentrations, limestone was added accordingly on top to achieve the same Ca concentration in all treatments. Representative samples were taken for each treatment and ground through a 0.5-mm sieve for chemical analyses (Retsch, ZM 200, Haan, Germany).
Differences between calculated and analyzed Zn and Mn concentrations did not exceed 22% (Table 3). For Cu, differences did not exceed 25%, and for Ca, the maximal deviation was 12%. Differences between calculated and analyzed P concentrations did not exceed 10%. Maximal deviation of InsP6-P concentration did not exceed 20%. The maximal deviation of calculated and analyzed phytase activity values was 14% in the starter phase and 21% in the grower phase.
Experimental Procedures
For the determination of average daily feed intake (ADFI), average daily gain (ADG), and feed conversion ratio (FCR), broilers and feed were weighed on a pen basis on days 0, 12, and 28. Calculations were corrected for mortality by accounting for bird days. On day 28, birds were euthanized by carbon dioxide exposure. Ileal digesta was collected from 8 birds per pen by flushing with distilled water, pooled on a pen basis, and immediately put on dry ice. The ileum was defined as the distal two thirds of the segment between Meckel’s diverticulum and 2-cm anterior to the ileo-ceco-colonic junction. The left foot and tibiotarsus (tibia) were collected from 2 randomly selected birds per pen and frozen until further processing. Jejunal tissue (middle section), pectoralis muscle tissue, and liver tissue were collected from one randomly selected bird per pen for gene expression analysis. The tissues for gene expression analysis were snap-frozen in liquid nitrogen and stored at − 80°C until further processing.
Chemical Analyses
Ileal digesta was freeze-dried (Labconco 780601010, Labconco Corporation, Kansas City, MO, USA) and pulverized. Feed and freeze-dried ileal digesta samples were oven-dried at 100°C for 24 h following AOAC Method 934.01 [14] to determine dry matter (DM). For mineral analysis, samples of feed and ileal digesta were ashed in a muffle furnace (6 h, 550°C) with subsequent acid digestion (25% HCl) of the ash using AOAC Method 968.08 [14] and were analyzed for minerals following EPA Method 200.7 [15] by inductively coupled plasma-optical emission spectroscopy (ICP-OES, Spectro Arcos FHS16, Germany). Titanium was determined as described by Short et al. [16]. In brief, ashed samples were digested with 7.4 M sulfuric acid, hydrogen peroxide (30%) was added, and absorbance was measured in a spectrophotometer (Spectronic 200, Thermo Scientific, Waltham, MA, US) at 410 nm.
Tibiae were thawed and adhering soft tissue, fibula bones, and cartilage caps were manually removed. At the joint (articulatio intertarsalis), feet were detached and used completely below the joint including tissues, claws, and skins. Tibiae and feet were cleaned with distilled water, carefully patted, and dried for 48 h (tibiae) and 72 h (feet) at 100°C in a convection oven (VWR International, Radnor, PA, USA). Subsequently, the bone samples were cooled in a desiccator and weighed. The dried bones were ashed for 24 h (tibiae) or 48 h (feet) at 600°C in a muffle furnace (Lindenberg/Blue M, Thermo Scientific, Waltham, MA, USA). Ashed tibiae and feet were weighed and ground to 0.5 mm (Retsch, ZM 200, Haan, Germany). Minerals analysis of tibia ash was conducted as described for feed and ileal digesta.
The InsP3-6 isomers were determined in feed and ileal digesta using the method of Zeller et al. [17] with slight modifications described by Sommerfeld et al. [18]. To determine myo-inositol concentrations, digesta and feed were processed, and measurements were performed using an Agilent 5977A gas chromatograph/mass spectrometer (Waldbronn, Germany) as described in Sommerfeld et al. [19].
An ELISA assay was used to determine phytase activity in feed samples (AB Vista, Plantation, FL, USA), and results were subsequently converted to FTU per kilogram.
Quantitative Real-Time PCR Analysis
Quantitative real-time PCR was used to analyze gene expression of selected intestinal mineral transporters and protein synthesis and degradation in pectoralis muscle and liver tissue. Tissues (approximately 3 mm × 3 mm) were homogenized in QIAzol®Lysis Reagent (QIAGEN, Hilden, Germany). Total RNA was extracted according to the manufacturer’s instructions. Before converting extracted RNA to cDNA in a 96-well PCR system (Applied Biosystems™ Veriti™ Thermal Cycler) using a high-capacity cDNA reverse transcription kit (Thermo Fisher Scientific, Waltham, MA, USA), RNA quantity and quality were measured by use of synergy HTX multi-mode reader (BioTek Instruments, Winooski, VT, USA) and diluted accordingly. After cDNA conversion, the cDNA was diluted, and by using reaction master mix iTaq Universal SYBR® Green Supermix (Bio-Rad Laboratories, Hercules, CA, USA) real-time PCR reaction was performed in a CFX96™ Real-Time System (Bio-Rad Laboratories, Hercules, CA, USA). The PCR conditions were as follows: 95°C for 5 min; 40 cycles of 95°C for 10 s, annealing temperature for 45 s. The annealing temperature was adapted according to the primer. All samples were run in duplicate, and the 2(−ΔΔCt) method [20] was used for data analysis. The low-sulfates treatment was used as a control for the ΔΔCt normalization. All primers and their accession numbers are shown in Table 4.
Calculations and Statistical Analysis
On a pen basis, growth performance (ADG, ADFI, FCR) was calculated for the entire trial duration (day 0 to 28) and was corrected for mortality. Digesta samples and bones were pooled on a pen basis, and the pen was considered the experimental unit. The bird was considered the experimental unit for gene expression as tissues were obtained from individual birds.
The prececal InsP6 disappearance and prececal P and Ca digestibility were calculated based on analyzed concentrations of Ti, P, Ca, and InsP6 in feed and digesta, using the following equation:
where \({\text{y}}\left({\text{X}}\right)\) is the disappearance or digestibility of \({\text{X}}\) in % and \({\text{X}}\) is the concentration of P, Ca, or InsP6 in feed and digesta.
Data were checked with ANOVA residual diagnostic plots for normal distribution and variance homogeneity and analyzed in a nested 2-factorial analysis of variance using the GLIMMIX procedure of the software package SAS (version 9.4, SAS Institute Inc., Cary, NC); means were separated using t test. The following model was chosen:
where \({\text{y}}_{\text{ijkl}}\) is the response variable, \(\mu\) is the overall mean, \({\text{a}}\) represents the fixed effect of the ith treatment group (\({\text{i}}\) = low-sulfates or others), \({\text{b}}\) represents the fixed effect of the jth phytase supplementation level within the \(\textit{i}\)th treatment group (\({\text{j}}\) = 0 or 750 FTU/kg), \({\text{c}}\) represents the fixed effect of the kth trace mineral source within the \(\textit{i}\)th treatment group (\({\text{k}}\) = sulfates or oxides or chelates), \({\text{bc}}\) is the corresponding interaction of the jth phytase supplementation level and the kth trace mineral source within the ith treatment group, \(\delta\) is the random effect of lth block (\({\text{l}}\) = 1–8), and \({\varepsilon }_{\text{ijkl}}\) is the residual error. Statistical significance was declared at P ≤ 0.050.
Results
Ileal InsPx and myo-inositol concentrations
Ileal concentrations of InsP6 were affected by PHY × TMS interaction (P = 0.047, Table 5). The diets without phytase supplementation containing sulfates or oxides had higher ileal InsP6 concentrations than the diets containing chelates. Phytase supplementation decreased ileal InsP6 concentrations to a similar level regardless of TMS used. The Ins(1,2,4,5,6)P5 concentrations in sulfate diets were higher than in chelate diets (P < 0.021). Phytase supplementation increased Ins(1,2,3,4,5)P5 concentration (P < 0.001). The Ins(1,2,3,4,6)P5 isomer occurred at a quantifiable level only in diets without exogenous phytase. The concentrations of Ins(1,2,5,6)P4 and Ins(1,2,3,4)P4 were higher in diets with exogenous phytase than without (P < 0.001). Phytase supplementation increased InsP3x concentrations (P < 0.001). The concentrations of InsP3x were highest in birds fed the low-sulfates diet (P < 0.001). The PHY × TMS interaction affected ileal MI concentrations (P = 0.050). Ileal MI concentrations were higher in diets with exogenous phytase than without. Without exogenous phytase, feeding oxides resulted in a higher MI concentration than sulfates, whereas with exogenous phytase, feeding sulfates resulted in a higher MI concentration than feeding chelates.
Prececal InsP6 Disappearance and Mineral Digestibility
Prececal InsP6 disappearance was affected by PHY × TMS interaction (P < 0.001, Table 6). In the absence of exogenous phytase, birds fed chelates showed a higher prececal InsP6 disappearance than those fed oxides or sulfates. InsP6 disappearance was increased by phytase supplementation, without differences among the TMS. The InsP6 disappearance was similar between birds receiving the low-sulfates diet and those receiving the PHY+ diets. The PHY × TMS interaction affected prececal P digestibility (P = 0.003). Whereas the prececal digestibility of P in diets without exogenous phytase varied depending on TMS used, the TMS had no effect on prececal P digestibility in phytase-supplemented diets. Prececal P digestibility of birds fed the low-sulfates diet was lower than for birds receiving other diets (P < 0.001). Prececal digestibility of Ca was influenced by PHY × TMS interaction (P = 0.009). Whereas birds fed sulfates or chelates had similar prececal Ca digestibility whether phytase was supplemented or not, birds fed oxides had higher Ca digestibility in phytase-supplemented diets. Birds receiving the low-sulfate diet had lower prececal Ca digestibility than birds receiving other diets (P < 0.001).
Bone Ash
Phytase supplementation increased tibia and foot ash quantity and concentration (P < 0.050, Table 7). Tibia ash concentration was higher when birds received the low-sulfate diet compared with the other diets (P = 0.007). Foot ash concentration was higher in birds receiving chelates than sulfates (P = 0.038). Diet did not affect concentrations of P, Ca, and Cu in tibia ash. Interaction of PHY × TMS affected Zn concentration in tibia ash (P = 0.034). The concentration of Zn was increased in sulfate and oxide diets by phytase supplementation but remained at similar and low levels in diets with chelates, whether phytase was supplemented or not. The Mn concentration in tibia ash was lower in birds receiving the low-sulfate diet than in birds receiving the other diets and was increased by phytase supplementation (P < 0.001).
Gene Expression
Jejunum
Gene expression of Zn transporter 5 (ZnT5) in the jejunum was affected by PHY × TMS interaction (P = 0.024, Table 8). Whereas the expression of ZnT5 remained at similar levels in diets with oxides and chelated TM whether phytase was supplemented or not, the expression was increased in sulfate diets when phytase was supplemented. Gene expression of Zn transporter 10 (ZnT10) and of phosphate transporter (NaPi-IIb) was not affected by diet (P > 0.050). Expression of phosphate transporter (PiT-2) tended to be affected by phytase supplementation (P = 0.080). The gene expression of Cu transporter 1 (CTR1) tended to be affected by TMS (P = 0.058).
Pectoralis Muscle and Liver
In the pectoralis muscle, expression of EIF4EBP1 (eukaryotic translation initiation factor 4E binding protein 1) and mTOR (mechanistic target of rapamycin) was not affected by diet (P > 0.050, Table 9). Expression of Trim63 (Tripartite motif containing 63) and FBXO32 (F-box only protein 32) was increased in the low-sulfate diet compared to the other diets (P < 0.050).
In the liver, expression of mTOR and S6kinase was not affected by diet (P > 0.050, Table 9). Supplementation of phytase increased the expression of EIF4EBP1 (P = 0.033). The expression of Trim63 was not affected by diet (P > 0.050). Phytase supplementation tended to affect the expression of FBXO32 and PRKAB2 (P < 0.100).
Performance Traits
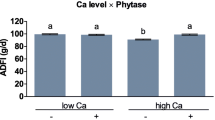
At the beginning of the experiment (day 0), the average body weight of broiler chickens was 45 g. During the experiment, 43 birds (4.8%) died, but mortality was unrelated to treatment. Birds fed the low-sulfate diet had higher ADG, ADFI, and lower FCR than birds fed other diets (P < 0.050, Table 10). Birds fed sulfates or oxides tended to have higher ADG (P = 0.057) and had lower FCR (P = 0.009) than birds receiving chelates.
Discussion
The first objective was to investigate how different TMS affect prececal phytate breakdown to MI and the second objective was to investigate whether TMS and exogenous phytase affect trace mineral concentration in the bone.
Effects of Trace Mineral Source on Phytate Breakdown
The InsP6 disappearance was higher for chelates than for oxides or sulfates in the PHY− diets, partially confirming the first hypothesis of this trial that the TMS affects InsP6 disappearance. This supports the assumption that chelated trace minerals are less prone to interact with InsP6 than inorganic trace minerals [21,22,23]. This could be because chelates might dissociate slower than sulfates and oxides, leaving less time for the formation of complexes of divalent cations and phytate. Another hypothesis was proposed by Wedekind et al. [23], who suggested that the ligands of chelates may compete with phytate for its ability to bind Zn. In contrast, Schlegel et al. [24] suggested that native (plant-derived) InsP6 does not interact with supplemental Zn and that chelation of inorganic Zn sources is not necessary [25]. Differences in the behavior of chelated sources could be a reason why divergent results were found in the literature. Chelated sources differ in their chelation strength, and this determines whether the chelation remains stable in different pH conditions [26].
In PHY+ diets, however, no differences in InsP6 disappearance were observed between TMS in the present experiment. This contradicts an in vitro study by Santos et al. [8], which showed that when Na-phytate was used, different sources (for example, glycinates and sulfates) of Zn, Cu, and Mn differed in their inhibitory effect on exogenous phytase efficacy. A possible explanation for these differences could be differences in substrates. The reactivity of Na-phytate and native phytate (derived from corn and soybean in the present trial) is different and could be higher for Na-phytate than native phytate [27]. In addition, the order in which the reactions or interactions of divalent cations and phytate occur may be different in vitro and in vivo. Whereas in vitro all reactions occur at the same time, the interactions in vivo occur in different sections of the gastrointestinal tract to different extents due to varying conditions. Another difference is that in the present study, the effects of Zn, Cu, and Mn were tested in combination, whereas in the in vitro study by Santos et al. [8], individual TM were tested.
A major part of phytate degradation by exogenous phytase occurs in the proximal digestive tract [17] resulting in a lower potential or possibility of strong complex formation of InsP6 with minerals in the small intestine, where pH is favorable for stable phytate-mineral-complex formation [28]. The lower interactive potential could be the reason why no differences were observed in the prececal InsP6 disappearance between the TMS in the present experiment upon adding exogenous phytase. Although the concentration of partially dephosphorylated InsPx increased in the small intestine in phytase-supplemented diets, the overall interactive potential is reduced because the binding strength between InsPx and (divalent) cations decreases with decreasing degree of phosphorylation [29].
Complete dephosphorylation of InsP6 leads to the release of MI. Consistent with previous studies [19, 30, 31], the ileal MI concentration was increased by exogenous phytase in the present study. To our knowledge, there is no information in the literature on the release of MI when different TMS are used. In the present study, ileal MI concentrations differed depending on TMS used: whereas in PHY− diets, the highest MI concentration was observed for oxides, in PHY+ diets, the highest MI concentration was found for sulfates. The supplementation of chelates resulted in the lowest MI concentrations. A significant difference in the ileal MI concentration was also found between the phytase-supplemented sulfate diet and the low-sulfate diet. The lower concentration of MI coincided with a higher accumulation of Ins(1,2,5,6)P4 and InsP3x in the low-sulfate diet. This could indicate lower activity of endogenous phosphatases in this diet. The Zn concentration in the low-sulfate diet may not have been sufficient for the endogenous phosphatases to develop their full potential since Zn is a cofactor of phosphatases [32]. This fits with the hypothesis suggested by Philippi et al. [33] that low dietary Zn concentrations may not be sufficient to achieve full activity of endogenous phosphatases. However, it has to be kept in mind that MI absorption by the chicken intestine is barely understood [34], and it cannot be ruled out that TMS affected MI transporters.
As expected, prececal P digestibility was increased by exogenous phytase. Except for the low-sulfate diet, P digestibility followed the same pattern as the InsP6 disappearance. Potential reasons why the prececal P digestibility of the low-sulfate diet was on a similar level as the PHY− diets could not be found. The chemical composition of this diet gave no indication as to why the prececal P digestibility was much lower than expected. In part, the low prececal P digestibility could be due to a lower concentration of titanium dioxide in the ileal digesta in the low-sulfates diet than in the other diets (data not shown). Still, the reasons why the titanium dioxide concentration was lower could not be identified. The prececal P digestibility of the low-sulfate diet should be interpreted with caution because it is inconsistent with the bone results.
In birds, active intestinal phosphate transport is mainly performed by Na-dependent phosphate transporters such as the NaPi-IIb [35]. In the present study, the expression of NaPi-IIb and PiT-2 was not affected by TMS or exogenous phytase or their interaction. As the investigation of gene expression on mRNA level or protein level does not allow us to conclude about the activity of a transporter, both increased transporter activity and increased paracellular uptake could be the mediators for increased phosphate absorption in PHY+ diets.
Prececal Ca digestibility was similar for chelate and sulfate diets whether or not phytase was supplemented. Yenice et al. [36] observed increased serum Ca concentrations in laying hens fed a chelated compared to an inorganic trace mineral mixture. The authors suggested that using chelated trace minerals reduces the amount of free ions in the small intestine, leaving fewer free ions available to form insoluble complexes. Still, because significant amounts of Ca can be excreted via the urine, the prececal Ca digestibility and serum Ca concentrations are difficult to interpret. Contrary to the hypothesis of Yenice et al. [36], Bertolo et al. [37] hypothesized that Ca2+ and Zn2+ may use the same non-specific transporter in the brush border membrane, leading to competition between Ca2+ and Zn2+. Yet, in the present study, Ca digestibility was significantly lower in the low-sulfates diet (15 mg/kg Zn supplementation) than in the sulfate diet (100 mg/kg Zn supplementation).
In consistency with other studies [38, 39], phytase supplementation increased the quantity (g/bone) of tibia and foot ash and concentration (g/g) of tibia and foot ash in the present study. Foot ash concentration was higher in the chelate diets than in the sulfate diets, reflecting the higher prececal InsP6 disappearance and P digestibility of the PHY–chelate diet compared to the PHY–sulfate diet. However, the main effect of TMS was not significant for tibia ash. It cannot be excluded that the composition of the soft tissue, which is also included in the ashing of the foot, changes depending on the TMS.
Augspurger et al. [40] found that tibia ash concentration was reduced in broilers when very high concentrations of Zn (800 mg/kg) were added to a phytase-supplemented diet compared with phytase supplementation alone. Mohanna and Nys [41] and Gosh et al. [42] observed no effect of the dietary Mn concentration (40 or 100 mg/kg Mn) on tibia ash quantity or concentration. Furthermore, high dietary Cu concentrations (200 mg/kg Cu) did not result in changes in the quantity or concentration of tibia ash in the study by Augspurger et al. [40]. Also, in a study by Banks et al. [43], high supplementations of Cu (up to 250 mg/kg) as Cu sulfate did not affect tibia ash concentrations. In the present study, no effects of trace mineral concentrations were observed on tibia ash quantity or concentration. However, it should be noted that these comparisons are confounded by differences in study duration, Zn concentration (800 vs. 100 mg), Mn concentration (40 vs. 100 mg), and Cu concentration (200/250 vs. 125 mg) and by the simultaneous supplementation of Zn, Cu, and Mn in the present study as opposed to changes of single minerals.
Effects of Phytase and Trace Mineral Source on Bone Trace Mineral Concentration and Performance
Zinc concentration in tibia ash was increased by phytase supplementation in sulfate and oxide diets. This has been observed before and is probably due to the release of Zn associated with InsP6 degradation [44]. Phytase supplementation did not increase the Zn concentration in tibia ash in the diet with chelated TMS in the present trial. Pang and Applegate [45] found that a high Cu concentration (250 mg/kg) supplemented as Cu chelate reduced Zn solubility in duodenum and jejunum. In contrast, the supplementation of Cu sulfate did not lead to reduced Zn solubility, supporting the results of the present study. Yet, no explanation was provided as to why Cu sulfate had no lowering effect on Zn solubility despite the antagonism between Cu and Zn [46]. Mohanna and Nys [41] observed increased Mn concentrations in tibia ash due to the combined supplementation of Mn sulfate and Zn sulfate. This is consistent with the higher Mn concentration in tibia ash in the sulfate diet compared to the low-sulfate diet. However, unlike in the present trial, phytase supplementation did not affect Mn concentration in tibia ash in a 21-day trial [41]. Singh et al. [47] observed an increasing effect of exogenous phytase on tibia Mn concentration on day 42 but not on day 21. When tibia ash is used as a trait to evaluate P availability, it has been shown to be more sensitive in a 4-week assay compared to a 10-day assay [48]. This could be one reason why differences were observed at different time points. In the present experiment, no differences in Mn concentration of tibia ash attributable to TMS were detected when supplemented at the same level. This confirms the results of Li et al. [49], where differences in the Mn concentration of tibia ash were only found due to different Mn supplementation levels but remained the same whether chelated or inorganic Mn sources were used. Consistent with observations by Nguyen et al. [50] in a study with 35-day-old broilers, Cu concentration in the present experiment was not affected by the dietary Cu concentration or the Cu source used.
In contrast to results from Bortoluzzi et al. [51], no consistent association between Zn supplementation and gene expression level of ZnT5 was found in the present study. In agreement with Hu et al. [52], the expression of ZnT10 mRNA level in the present experiment tended to be upwardly expressed in the diets with high TM supplementation compared to the low-sulfates diet. In addition, no differences were observed between chelate and sulfate diets with regards to mRNA level of ZnT10 in the present study or in the study by Hu et al. [52]. However, Hu et al. [52] observed increased expression of ZnT10 at the protein level in birds receiving a chelated Zn source compared to birds receiving ZnSO4. Of note, Buccitelli and Selbach [53] reviewed mRNA-level and protein-level measurements and concluded that both give valuable insights but have limitations in their interpretability in terms of active biological function of proteins. The gene expression on mRNA-level is only a “construction plan” for protein synthesis, and the protein-level simply shows that the respective mRNA was available and the protein was synthesized. Furthermore, the expression of CTR1 was analyzed at the mRNA level in the present experiment. Similar as in an experiment by Meng et al. [54], no significant differences were found in the expression of CTR1 due to TMS in the present trial. However, the comparison in the study by Meng et al. [54] was confounded by different Mn concentrations (100 mg/kg Mn as sulfate vs. 50 mg/kg Mn as chelate). Because CTR1 can be affected by Cu, Zn, and Mn [54, 55], interpretation is difficult.
The expression of genes related to protein synthesis and degradation in the pectoralis muscle and liver was analyzed to investigate whether the observed differences in growth performance correspond to changes in gene expression. Olukosi et al. [56] found an indication that differences in growth performance between Zn- and Cu-hydroxychloride compared with Zn- and Cu-sulfate could be partially explained by differences in mRNA expression of a gene in pectoralis muscle related to protein degradation. However, genes in the pectoralis muscle and liver analyzed in the present study could not indicate as to why growth performance differed when using different TMS. Among the genes analyzed, only mRNA expression of EIF4EBP1 in the liver was significantly affected by diet. The higher mRNA expression of this gene in the PHY+ diets compared with PHY− diets coincided with a numerically higher ADG of PHY+ diets.
Conclusions
In conclusion, the prececal InsP6 degradation by endogenous phosphatases (PHY− treatments) was slightly higher when chelates were used compared to sulfates or oxides, partially confirming the first hypothesis that TMS affects prececal InsP6 degradation. Adding exogenous phytase (750 FTU/kg) removed the differences between different TMS in InsP6 disappearance, probably due to a lower potential for complex formation. Since the vast majority of broiler diets is commonly supplemented with phytase, the choice of TMS is likely negligible in terms of phytate degradation.
The second hypothesis that phytase supplementation affects trace mineral concentration in the bone was confirmed for Zn and Mn but not for Cu. Concentrations of Zn and Mn in tibia ash were increased by exogenous phytase, although the supply was above NRC supply recommendations. The concentration of Cu in tibia ash was not affected by exogenous phytase or diet in general. Future studies should examine tissues other than bone to evaluate Cu accumulation in the bird.
Data availability
Data are not publicly available but are available from the corresponding author on reasonable request.
References
Zeigler TR, Leach RM, Norris LC, Scott ML (1961) Zinc requirement of the chick: factors affecting requirement. Poult Sci 40:1584–1593. https://doi.org/10.3382/ps.0401584
Suttle NF (2022) Mineral nutrition of livestock, 5th edn. CABI, Wallingford
National Research Council (1994) Nutrient requirements of poultry, 9th edn. National Academies Press, Washington, DC
Hamdi M, Solà D, Franco R, Durosoy S, Roméo A, Pérez JF (2018) Including copper sulphate or dicopper oxide in the diet of broiler chickens affects performance and copper content in the liver. Anim Feed Sci Technol 237:89–97. https://doi.org/10.1016/j.anifeedsci.2018.01.014
Rodehutscord M, Sommerfeld V, Kühn I, Bedford MR (2022) Phytases Potential and limits of phytate destruction in the digestive tract of pigs and poultry. In: Bedford et al (eds) Enzymes in farm animal nutrition, 3rd edn. CABI, Wallingford, pp 124–152
Rodehutscord M, Rosenfelder P (2016) Update on phytate degradation pattern in the gastrointestinal tract of pigs and broiler chickens. In: Walk et al (eds) Phytate destruction-consequences for precision animal nutrition. Wageningen Academic Publishers, Wageningen, pp 15–28
Tamim NM, Angel R (2003) Phytate phosphorus hydrolysis as influenced by dietary calcium and micro-mineral source in broiler diets. J Agric Food Chem 51:4687–4693. https://doi.org/10.1021/jf034122x
Santos T, Connolly C, Murphy R (2015) Trace element inhibition of phytase activity. Biol Trace Elem Res 163:255–265. https://doi.org/10.1007/s12011-014-0161-y
Maddaiah VT, Kurnick AA, Reid BL (1964) Phytic acid studies. Proc Soc Exp Biol Med 115:391–393. https://doi.org/10.3181/00379727-115-28922
Champagne E, Hinojosa O (1987) Independent and mutual interactions of copper(II) and zinc(II) ions with phytic acid. J Inorg Biochem 30:15–33. https://doi.org/10.1016/0162-0134(87)80041-7
Reddy NR, Sathe SK, Salunkhe DK (1982) Phytates in legumes and cereals. Adv Food Res 28:1–92. https://doi.org/10.1016/S0065-2628(08)60110-X
Cardoso D, Romeo A, Durosoy S, Meme N, Chevalier Y, Narcy A (2021) Effect of the physicochemical properties of zinc oxide sources on their bioavailability in broilers. Br Poult Sci 62:846–851. https://doi.org/10.1080/00071668.2021.1940862
Cobb (2022) Cobb500 Broiler. Performance & nutrition supplement. https://www.cobb-vantress.com/assets/5a88f2e793/Broiler-Performance-Nutrition-Supplement.pdf. Accessed 2022-04-20
AOAC (2006) Official methods of analysis of AOAC international, 18th edn. AOAC International, Arlington, VA
USEPA (1994) Method 200.7: Determination of metals and trace elements in water and wastes by inductively coupled plasma-atomic emission spectrometry. Revision 4.4, EMMC Version 1994. Revision 4.4, EMMC Version 1994. United States Environmental Protection Agency, Cincinnati, OH
Short FJ, Gorton P, Wiseman J, Boorman KN (1996) Determination of titanium dioxide added as an inert marker in chicken digestibility studies. Anim Feed Sci Technol 59:215–221. https://doi.org/10.1016/0377-8401(95)00916-7
Zeller E, Schollenberger M, Kühn I, Rodehutscord M (2015) Hydrolysis of phytate and formation of inositol phosphate isomers without or with supplemented phytases in different segments of the digestive tract of broilers. J Nutr Sci 4:e1. https://doi.org/10.1017/jns.2014.62
Sommerfeld V, Schollenberger M, Kühn I, Rodehutscord M (2018) Interactive effects of phosphorus, calcium, and phytase supplements on products of phytate degradation in the digestive tract of broiler chickens. Poult Sci 97:1177–1188. https://doi.org/10.3382/ps/pex404
Sommerfeld V, Künzel S, Schollenberger M, Kühn I, Rodehutscord M (2018) Influence of phytase or myo-inositol supplements on performance and phytate degradation products in the crop, ileum, and blood of broiler chickens. Poult Sci 97:920–929. https://doi.org/10.3382/ps/pex390
Livak KJ, Schmittgen TD (2001) Analysis of relative gene expression data using real-time quantitative PCR and the 2− ΔΔCT method. Methods 25:402–408. https://doi.org/10.1006/meth.2001.1262
Burrell AL, Dozier WA, Davis AJ, Compton MM, Freeman ME, Vendrell PF, Ward TL (2004) Responses of broilers to dietary zinc concentrations and sources in relation to environmental implications. Br Poult Sci 45:255–263. https://doi.org/10.1080/00071660410001715867
Świątkiewicz S, Koreleski J, Zhong D (2001) The bioavailability of zinc from inorganic and organic sources in broiler chickens as affected by addition of phytase. J Anim Feed Sci 10:317–28. https://doi.org/10.22358/jafs/67987/2001
Wedekind KJ, Hortin AE, Baker DH (1992) Methodology for assessing zinc bioavailability: efficacy estimates for zinc-methionine, zinc sulfate, and zinc oxide. J Anim Sci 70:178–187. https://doi.org/10.2527/1992.701178x
Schlegel P, Nys Y, Jondreville C (2010) Zinc availability and digestive zinc solubility in piglets and broilers fed diets varying in their phytate contents, phytase activity and supplemented zinc source. Animal 4:200–209. https://doi.org/10.1017/S1751731109990978
Schlegel P, Sauvant D, Jondreville C (2013) Bioavailability of zinc sources and their interaction with phytates in broilers and piglets. Animal 7:47–59. https://doi.org/10.1017/S1751731112001000
Byrne L, Hynes MJ, Connolly CD, Murphy RA (2021) Influence of the chelation process on the stability of organic trace mineral supplements used in animal nutrition. Animals 11:1730. https://doi.org/10.3390/ani11061730
Fordyce EJ, Forbes RM, Robbins KR, Erdman JW (1987) Phytate×calcium/zinc molar ratios: are they predictive of zinc bioavailability? J Food Sci 52:440–444. https://doi.org/10.1111/j.1365-2621.1987.tb06634.x
Nolan KB, Duffin PA, McWeeny DJ (1987) Effects of phytate on mineral bioavailability.in vitro studies on Mg2+, Ca2+, Fe3+, Cu2+ and Zn2+ (also Cd2+) solubilities in the presence of phytate. J Sci Food Agric 40:79–85. https://doi.org/10.1002/jsfa.2740400110
Persson H, Türk M, Nyman M, Sandberg AS (1998) Binding of Cu2+, Zn2+, and Cd2+ to inositol tri-, tetra-, penta-, and hexaphosphates. J Agric Food Chem 46:3194–3200. https://doi.org/10.1021/jf971055w
Walk CL, Olukosi OA (2019) Influence of graded concentrations of phytase in high-phytate diets on growth performance, apparent ileal amino acid digestibility, and phytate concentration in broilers from hatch to 28 D post-hatch. Poult Sci 98:3884–3893. https://doi.org/10.3382/ps/pez106
Novotny M, Sommerfeld V, Krieg J, Kühn I, Huber K, Rodehutscord M (2023) Comparison of mucosal phosphatase activity, phytate degradation, and nutrient digestibility in 3-week-old turkeys and broilers at different dietary levels of phosphorus and phytase. Poult Sci 102:102457. https://doi.org/10.1016/j.psj.2022.102457
McCall KA, Huang C-c, Fierke CA (2000) Function and mechanism of zinc metalloenzymes. J Nutr 130:1437S-1446S. https://doi.org/10.1093/jn/130.5.1437S
Philippi H, Sommerfeld V, Olukosi OA, Windisch W, Monteiro A, Rodehutscord M (2023) Effect of dietary zinc source, zinc concentration, and exogenous phytase on intestinal phytate degradation products, bone mineralization, and zinc status of broiler chickens. Poult Sci 102:103160. https://doi.org/10.1016/j.psj.2023.103160
Gonzalez-Uarquin F, Rodehutscord M, Huber K (2020) Myo-inositol: its metabolism and potential implications for poultry nutrition-a review. Poult Sci 99:893–905. https://doi.org/10.1016/j.psj.2019.10.014
Huber K, Zeller E, Rodehutscord M (2015) Modulation of small intestinal phosphate transporter by dietary supplements of mineral phosphorus and phytase in broilers. Poult Sci 94:1009–1017. https://doi.org/10.3382/ps/pev065
Yenice E, Mızrak C, Gültekin M, Atik Z, Tunca M (2015) Effects of organic and inorganic forms of manganese, zinc, copper, and chromium on bioavailability of these minerals and calcium in late-phase laying hens. Biol Trace Elem Res 167:300–307. https://doi.org/10.1007/s12011-015-0313-8
Bertolo RFP, Bettger WJ, Atkinson SA (2001) Calcium competes with zinc for a channel mechanism on the brush border membrane of piglet intestine. J Nutr Biochem 12:66–72. https://doi.org/10.1016/S0955-2863(00)00125-X
Künzel S, Borda-Molina D, Kraft R, Sommerfeld V, Kühn I, Camarinha-Silva A, Rodehutscord M (2019) Impact of coccidiostat and phytase supplementation on gut microbiota composition and phytate degradation in broiler chickens. Animal microbiome 1:1–14. https://doi.org/10.1186/s42523-019-0006-2
Künzel S, Borda-Molina D, Zuber T, Hartung J, Siegert W, Feuerstein D, Camarinha-Silva A, Rodehutscord M (2021) Relative phytase efficacy values as affected by response traits, including ileal microbiota composition. Poult Sci 100:101133. https://doi.org/10.1016/j.psj.2021.101133
Augspurger NR, Spencer JD, Webel DM, Baker DH (2004) Pharmacological zinc levels reduce the phosphorus-releasing efficacy of phytase in young pigs and chickens. J Anim Sci 82:1732–1739. https://doi.org/10.2527/2004.8261732x
Mohanna C, Nys Y (1999) Changes in zinc and manganese availability in broiler chicks induced by vegetal and microbial phytases. Anim Feed Sci Technol 77:241–253. https://doi.org/10.1016/S0377-8401(98)00254-5
Ghosh A, Mandal GP, Roy A, Patra AK (2016) Effects of supplementation of manganese with or without phytase on growth performance, carcass traits, muscle and tibia composition, and immunity in broiler chickens. Livest Sci 191:80–85. https://doi.org/10.1016/j.livsci.2016.07.014
Banks KM, Thompson KL, Jaynes P, Applegate TJ (2004) The effects of copper on the efficacy of phytase, growth, and phosphorus retention in broiler chicks. Poult Sci 83:1335–1341. https://doi.org/10.1093/ps/83.8.1335
Philippi H, Sommerfeld V, Windisch W, Olukosi OA, Monteiro A, Rodehutscord M (2023) Interactions of zinc with phytate and phytase in the digestive tract of poultry and pigs: a review. J Sci Food Agric 103:7333–7342. https://doi.org/10.1002/jsfa.12879
Pang Y, Applegate TJ (2007) Effects of dietary copper supplementation and copper source on digesta pH, calcium, zinc, and copper complex size in the gastrointestinal tract of the broiler chicken. Poult Sci 86:531–537. https://doi.org/10.1093/ps/86.3.531
Starcher BC (1969) Studies on the mechanism of copper absorption in the chick. J Nutr 97:321–326. https://doi.org/10.1093/jn/97.3.321
Singh A, Walk CL, Ghosh TK, Bedford HS (2013) Effect of a novel microbial phytase on production performance and tibia mineral concentration in broiler chickens given low-calcium diets. Br Poult Sci 54:206–215. https://doi.org/10.1080/00071668.2013.775403
Ammerman CB, Douglas CR, Davis GK, Harms RH (1961) Comparison of phosphorus availability assay techniques for chicks. Poult Sci 40:548–553. https://doi.org/10.3382/ps.0400548
Li SF, Luo XG, Lu L, Crenshaw TD, Bu YQ, Liu B, Kuang X, Shao GZ, Yu SX (2005) Bioavailability of organic manganese sources in broilers fed high dietary calcium. Anim Feed Sci Technol 123:703–715. https://doi.org/10.1016/j.anifeedsci.2005.04.052
Nguyen HTT, Morgan N, Roberts JR, Swick RA, Toghyani M (2020) Copper hydroxychloride is more efficacious than copper sulfate in improving broiler chicken’s growth performance, both at nutritional and growth-promoting levels. Poult Sci 99:6964–6973. https://doi.org/10.1016/j.psj.2020.09.053
Bortoluzzi C, Vieira BS, Lumpkins B, Mathis GF, King WD, Graugnard D, Dawson KA, Applegate TJ (2019) Can dietary zinc diminish the impact of necrotic enteritis on growth performance of broiler chickens by modulating the intestinal immune-system and microbiota? Poult Sci 98:3181–3193. https://doi.org/10.3382/ps/pez045
Hu Y, Huang Y, Wang C, Zhang W, Qu Y, Li D, Wu W, Gao F, Zhu L, Wu B, Zhang L, Cui X, Li T, Geng Y, Liao X, Luo X (2023) The organic zinc with moderate chelation strength enhances the expression of related transporters in the jejunum and ileum of broilers. Poult Sci 102:102477. https://doi.org/10.1016/j.psj.2023.102477
Buccitelli C, Selbach M (2020) mRNAs, proteins and the emerging principles of gene expression control. Nat Rev Genet 21:630–644. https://doi.org/10.1038/s41576-020-0258-4
Meng T, Gao L, Xie C, Xiang Y, Huang Y, Zhang Y, Wu X (2021) Manganese methionine hydroxy analog chelated affects growth performance, trace element deposition and expression of related transporters of broilers. Animal Nutr 7:481–487. https://doi.org/10.1016/j.aninu.2020.09.005
Santos TS, Augusto KVZ, Han Y, Sartori MMP, Denadai JC, Santos CT, Sobral NC, Roça RO, Sartori JR (2021) High levels of copper and zinc supplementation in broiler diets on growth performance, carcass traits and apparent ileal mineral absorption. Br Poult Sci 62:579–588. https://doi.org/10.1080/00071668.2021.1887453
Olukosi OA, van Kuijk SJA, Han Y (2019) Sulfate and hydroxychloride trace minerals in poultry diets–comparative effects on egg production and quality in laying hens, and growth performance and oxidative stress response in broilers. Poult Sci 98:4961–4971. https://doi.org/10.3382/ps/pez261
Acknowledgements
We thank Lindsey Rackett, Derell Hardman, Shravani Veluri, Mohammad Pilevar, Yang Lin, Adeleye Ajao, and Iyabo Oluseyifunmi for their assistance in conducting the experiment and chemical analyses. We also thank the laboratory team of the Animal Nutrition Department for the analysis of inositol phosphates and myo-inositol.
Funding
Open Access funding enabled and organized by Projekt DEAL. This study was funded by the Animine (Annecy, France).
Author information
Authors and Affiliations
Contributions
All authors contributed to the study design. Hanna Philippi and Oluyinka A. Olukosi were responsible for data collection and statistical analysis. The first draft of the manuscript was written by Hanna Philippi, and all authors commented on previous versions of the manuscript. All authors read and approved the final version of the manuscript.
Corresponding author
Ethics declarations
Ethics Approval
The Institutional Animal Care and Use Committee of the University of Georgia approved the protocol of the present trial (IACUC number: A2021-06–006).
Competing Interests
Alessandra Monteiro is an employee of Animine (FR). The authors declare no conflict of interest.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Philippi, H., Sommerfeld, V., Monteiro, A. et al. Impact of Trace Mineral Source and Phytase Supplementation on Prececal Phytate Degradation and Mineral Digestibility, Bone Mineralization, and Tissue Gene Expression in Broiler Chickens. Biol Trace Elem Res (2024). https://doi.org/10.1007/s12011-024-04076-w
Received:
Accepted:
Published:
DOI: https://doi.org/10.1007/s12011-024-04076-w