
Abstract
The synthesis of selenocysteine and its incorporation into selenoproteins require serine during the action of seryl-tRNA synthetase. In view of this, we conducted this study to explore the effects of dietary serine supplementation on selenoprotein transcription and selenoenzyme activity in pigs. A total of 35 crossbred barrows (28 days old) were randomly assigned to five treatment groups. During the 42-day growth experiment, pigs were fed either a basal diet with no supplemented serine or diets supplemented with 0.25%, 0.5%, 0.75%, or 1% serine. The results showed that serine supplementation had no effect on the selenium content in the serum, skeletal muscle, and kidney of pigs. However, dietary supplementation with 0.5% serine significantly increased the selenium content in the liver. Diets supplemented with different levels of serine significantly increased the gene expression of glutathione peroxidase 1 (Gpx1), Gpx2, thioredoxin reductase 1 (Txnrd1), Txnrd2, and selenoprotein P (Sepp1) in the skeletal muscle and liver of pigs. Moreover, pigs supplemented with 0.5% serine had the highest selenoprotein P concentration and glutathione peroxidase (GPx) and thioredoxin reductase (TrxR) activities in the skeletal muscle, which were significantly higher than those in the control pigs. Additionally, pigs supplemented with 0.25% serine had the highest GPx and TrxR activities in the liver, which were significantly higher than those in the control pigs. In conclusion, dietary serine supplementation could improve selenoprotein transcription and selenoenzyme activity in pigs, with the appropriate concentrations of serine to be included in the diet being 0.25% or 0.5%.
Similar content being viewed by others
Avoid common mistakes on your manuscript.
Introduction
Selenium (Se) is considered an essential micronutrient for mammalian health. Se plays critical roles in a wide range of physiological processes, including the modulation and function of the antioxidant and the immune systems [1]. Several sources of Se have been commonly supplemented in livestock diets for improving growth performance, antioxidant and immune functions, and meat quality [2]. However, the bioavailability of Se, particularly for the inorganic form, is very low. Only approximately 1–2% of consumed inorganic Se is absorbed by the body [3]. Additionally, dietary Se is only beneficial for the animals within a narrow range, while an overdose of Se can result in selenosis. For example, a recent study found that dietary Se over 5 ppm was toxic to pigs [4]. Consequently, strategies to improve Se bioavailability are in high demand.
Se exerts most of its biological function through selenoproteins [5]. Selenocysteine, which is located in enzyme-active sites, is essential for selenoprotein activity [6]. The biosynthesis of selenocysteine and its insertion into selenoproteins, which is catalyzed by seryl-tRNA synthetase, require serine as an essential substrate [7]. Therefore, serine is needed for selenoprotein synthesis. A previous study demonstrated that serine exerted combined effects with selenocompounds, such as sodium selenite, selenomethionine, and selenomethylselenocysteine, to improve the expression of selenoprotein P (SelP) and glutathione peroxidase (GPx) in the liver based on in vitro experiments [8]. Additionally, with the increased intake of dietary serine, the blood Se content was increased in rats [9]. However, to our knowledge, little research has been conducted to investigate the possible synergistic effects of serine with Se in pigs. Consequently, the current study was performed to explore the effects of dietary serine supplementation on the Se content of tissue, selenoprotein expression, and selenoenzyme activity in pigs.
Materials and Methods
Experimental Design and Sample Collection
A total of 35 crossbred barrows (Duroc × Landrace × Yorkshire) were used in the experiment. After weanling at the age of 21 days, the pigs were fed a basal diet for 1 week. Then, the animals were randomly assigned into five treatment groups with seven replicates in each group. During the 42-day growth experiment, pigs were fed either a basal diet with no supplemented serine (CONT) or diets supplemented with 0.25%, 0.5%, 0.75%, or 1% serine, respectively. The basal diets were formulated to meet the National Research Council (NRC, 2012) recommendation (Table 1). Pigs were allowed ad libitum access to feed and water throughout the experimental period. The experimental protocol was approved by the Protocol Management and Review Committee of the Institute of Subtropical Agriculture, Chinese Academy of Science, and pigs were cared for and slaughtered according to the guidelines of the Institute of Subtropical Agriculture on Animal Care (Changsha, China). At the end of the experiment, blood samples were immediately collected and centrifuged at 2500 g at 4 °C for 10 min to obtain serum samples. Immediately after slaughtering, longissimus dorsi muscle, liver, and kidney samples were collected and stored at − 80 °C for further analysis.
Serum and Tissue Se Content Analysis
Serum and tissue samples were pretreated according to the Chinese National Standard (GB 5009.268—2016). Briefly, 0.5 g tissue or 0.5 mL serum was mixed with 8 mL nitric acid in 50-mL digestion tubes and kept at 20 ± 2 °C overnight. Then, the mixture was maintained at 120 °C for 5 min, 150 °C for 10 min, and 190 °C for 20 min, with a 5-min heat-up period in a microwave digestion system. Subsequently, the diluted samples were filtered through a 0.45-μm filter for the determination of Se content on Agilent 7900 ICP-MS (Agilent Technologies, Santa Clara, CA, USA) as previously described [10].
Measurement of Selenoproteins Activities and Antioxidant-Related Parameters
Concentrations of Se1P and malondialdehyde (MDA), as well as activities of GPx, thioredoxin reductase (TrxR), superoxide dismutase (SOD), and total antioxidant capacity (T-AOC), were measured using commercially available kits (Meimian, Jiangsu Yutong Biological Technology Co., Ltd., Nanjing, China) according to the manufacturer’s instructions.
Quantitative Real-Time PCR
Total RNA was isolated from skeletal muscle and liver samples with the TRIZOL reagents (Invitrogen, Carlsbad, CA, USA). To amplify cDNA fragments, we used primers as previously described (Table 2) [11]. A housekeeping gene (β-actin) was used to normalize target gene transcript levels. Real-time PCR was performed as previously described [12]. Briefly, 1 μL cDNA template was added with 3 μL deionized H2O, 5 μL SYBR Green mix, 0.2 μL Rox, and 0.4 μL each of forward and reverse primers. Relative mRNA expression was quantitated by the comparative Ct value method.
Statistical Analysis
All statistical analyses were performed by one-way ANOVA using the general linear model procedures and a mixed procedure (PROC MIXED) of the SAS software version 9.2 (SAS Institute Inc., Cary, NC, USA). Data are expressed as least squares means ± pooled SEM. Probability values < 0.05 were taken to indicate statistical significance.
Results
Effects of Dietary Serine Supplementation on Tissue Se Content in Pigs
As shown in Table 3, serine supplementation had no effect on the Se content in the serum, skeletal muscle, and kidney of pigs. Pigs fed a diet supplemented with 0.5% serine had the highest Se content in the liver, which was significantly higher than those of pigs fed the control diet and pigs fed diets supplemented with 0.75% or 1% serine.
Effects of Dietary Serine Supplementation on Selenoprotein Gene Expression in the Skeletal Muscle and Liver of Pigs
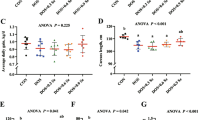
According to Fig. 1, dietary serine supplementation significantly increased the expression of Gpx1, Gpx2, Txnrd1, Txnrd2, and Sepp1 genes in the skeletal muscle and liver of pigs, while it had no significant effect on the expression of Selm and Sepn1. Dietary serine supplementation also significantly increased the expression of Gpx3 in the liver. There were no significant differences in the expression of these selenoprotein genes in the skeletal muscle and liver among the pigs fed diets supplemented with different levels of serine.

Effects of dietary serine supplementation on selenoprotein gene expression in the skeletal muscle and liver of pigs. Relative mRNA expression of selenoprotein in the skeletal muscle (a) and liver (b). Data were expressed as mean ± SEM, n = 7. *p < 0.05
Effects of Dietary Serine Supplementation on the Selenoprotein Concentration and Activity in the Skeletal Muscle and Liver of Pigs
As shown in Table 4, pigs fed diets supplemented with 0.25% or 0.5% serine had a significantly higher TrxR activity than did control pigs and pigs fed diets supplemented with 0.75% or 1% serine. However, there were no significant differences in the TrxR activity in the skeletal muscle among the control pigs and pigs fed diets supplemented with 0.75% or 1% serine. Pigs fed diets supplemented with 0.25% or 0.5% serine had a significantly higher GPx activity than did control pigs, while there were no significant differences in the GPx activity in the skeletal muscle among the control pigs and pigs fed diets supplemented with 0.75% or 1% serine. Pigs fed diets supplemented with different levels of serine had significantly higher SE1P concentrations in the skeletal muscle than did control pigs.
Pigs fed diets supplemented with 0.25% serine had a significantly higher TrxR activity than did control pigs, while there were no significant differences in the TrxR activity in the liver among the control pigs and pigs fed diets supplemented with 0.5%, 0.75%, or 1% serine. Pigs fed diets supplemented with 0.25%, 0.5%, or 0.75% serine had a significantly higher GPx activity and SE1P concentration than did control pigs, while there were no significant differences in the GPx activity and SE1P concentration in the liver among the control pigs and pigs fed diet supplemented with 1% serine.
Effects of Dietary Serine Supplementation on the Oxidative Status in the Skeletal Muscle and Liver of Pigs
As shown in Table 5, pigs fed diets supplemented with different levels of serine had a significantly lower MDA concentration and higher T-AOC activity in the skeletal muscle than did control pigs. Pigs fed diets supplemented with 0.5% or 0.75% serine had a significantly higher SOD activity than did control pigs and pigs fed diet supplemented with 0.25% serine. However, pigs fed diets supplemented with 0.25% or 1% serine had a significantly higher SOD activity in the skeletal muscle than did control pigs.
Pigs fed diets supplemented with different levels of serine had significantly higher T-AOC and SOD activities in the liver than did control pigs. Pigs fed diets supplemented with 0.5%, 0.75%, or 1% serine had a significantly lower MDA concentration in the liver than did control pigs and pigs fed diet supplemented with 0.25% serine. Additionally, pigs fed diet supplemented with 0.25% serine had a significantly lower MDA concentration in the liver than did control pigs.
Discussion
Se is an important micronutrient in the diet and is essential for health. Se deficiency can cause many kinds of diseases, while an overdose of Se can result in selenosis [13]. This suggests that it is crucial to improve Se bioavailability. Most research has focused on exploring new organic Se and comparing the bioactivity, transportation, and deposition with inorganic Se, with the organic sources usually having a higher Se bioavailability. Excluding the dependency upon speciation, Se bioavailability could be directly affected by dietary factors. A clinical study suggested that a balanced dietary structure could significantly improve the Se status of children [14]. Moreover, protein intake could also improve the absorption and deposition of Se [14]. Based on these factors, dietary protein or certain amino acids might affect the Se status of the body.
A previous study investigated the effects of dietary serine supplementation on the blood Se content in rats [9]. They found that serine and glycine deficiency increased the blood Se level in pregnant rats, while dietary serine supplementation increased the blood Se level in offspring rats. However, we found that dietary serine had no effects on the Se content in the serum, skeletal muscle, and kidney, while the Se content in the liver was significantly increased by dietary supplementation with 0.5% serine. These results are different from those of a previous study, as they found positive effects of dietary serine on the Se content [9]. There are many differences between the two experiments that may have caused these inconsistent results. Firstly, we used young growing pigs, while the authors of this previous study used pregnant rats and their offspring. This indicates that the effects of serine on the Se status may vary among species. Secondly, the experiment on rats lasted 2 months, while our experiment lasted 6 weeks. Finally, the rats were previously treated with a serine- and glycine-deficient diet for 30 days. Collectively, species, treatment period, and nutritional status are possible factors that influence the effects of serine on the Se status.
Serine participates in the transcription and translation processes of selenocysteine, which is essential for the activity of selenoproteins. We first explored the effects of serine on selenoprotein gene expression in pigs. Expectedly, the gene expression of selenoproteins, such as Gpx1, Gpx2, Txnrd1, Txnrd2, and Sepp1, were significantly increased in the skeletal muscle and liver, while the gene expression of other selenoproteins, such as Selm and Sepn1, were not changed. Subsequently, our results show that the increase in gene expression was consistent with the increased activities or concentrations of related selenoproteins, which is partially in line with the results of a previous in vitro study [8]. The previous study differed in that serine in combination with various kinds of selenocompounds increased Se1P and GPx concentrations in liver cells. However, they suggested that Se1P and GPx concentrations increased with the increased addition of serine in the culture medium of liver cells. In contrast, we found that pigs fed a diet supplemented with 0.5% serine had the highest Se1P concentration and GPx and TrxR activities. With 0.05% serine, the Se1P concentration and GPx and TrxR activities deceased with the increase in the dietary serine concentration. These results suggest that with the increase in serine addition, changes in the selenoprotein activity are different between in vivo and in vitro studies.
Although serine is considered a nonessential amino acid, it has been demonstrated to have many important functions. Importantly, recent studies have proved that dietary serine supplementation could exert antioxidative effects in rodents and pigs [15,16,17,18,19]. In these studies, one of the major mechanisms is that serine improved glutathione synthesis. Our results also indicated that dietary serine supplementation decreased oxidative stress in young growing pigs. Since the activities of GPx (an important antioxidant enzyme and an indicator of antioxidative status) and TrxR (an enzyme for maintaining intracellular redox status) were increased by serine supplementation, this proves that serine also increases the antioxidant capacity by improving the activities of selenoproteins, such as glutathione peroxidase and thioredoxin reductases.
In conclusion, our results suggest that dietary serine supplementation could improve transcription of selenoproteins and activities of selenoenzymes, although it did not affect the Se content in growing pigs. Our findings provide a nutritional strategy to prevent selenosis caused by the overdose of Se application in livestock production.
References
Kieliszek M, Blazejak S (2016) Current knowledge on the importance of selenium in food for living organisms: a review. Molecules 21(5). https://doi.org/10.3390/molecules21050609
Zhang K, Zhao Q, Zhan T, Han Y, Tang C, Zhang J (2019) Effect of different selenium sources on growth performance, tissue selenium content, meat quality, and selenoprotein gene expression in finishing pigs. Biol Trace Elem Res:1–9. https://doi.org/10.1007/s12011-019-01949-3
Castaneda-Ovando A, Segovia-Cruz JA, Flores-Aguilar JF, Rodriguez-Serrano GM, Salazar-Pereda V, Ramirez-Godinez J, Contreras-Lopez E, Jaimez-Ordaz J, Gonzalez-Olivares LG (2019) Serine-enriched minimal medium enhances conversion of selenium into selenocysteine by Streptococcus thermophilus. J Dairy Sci 102(8):6781–6789. https://doi.org/10.3168/jds.2019-16365
Kim YY, Mahan DC (2001) Comparative effects of high dietary levels of organic and inorganic selenium on selenium toxicity of growing-finishing pigs. J Anim Sci 79(4):942–948. https://doi.org/10.2527/2001.794942x
Labunskyy VM, Hatfield DL, Gladyshev VN (2014) Selenoproteins: molecular pathways and physiological roles. Physiol Rev 94(3):739–777. https://doi.org/10.1152/physrev.00039.2013
Kryukov GV, Castellano S, Novoselov SV, Lobanov AV, Zehtab O, Guigo R, Gladyshev VN (2003) Characterization of mammalian selenoproteomes. Science 300(5624):1439–1443. https://doi.org/10.1126/science.1083516
Amberg R, Mizutani T, Wu XQ, Gross HJ (1996) Selenocysteine synthesis in mammalia: an identity switch from tRNASerto tRNASec. J Mol Biol 263(1):8–19. https://doi.org/10.1006/jmbi.1996.0552
Wang Q, Sun LC, Liu YQ, Lu JX, Han F, Huang ZW (2016) The synergistic effect of serine with selenocompounds on the expression of SelP and GPx in HepG2 cells. Biol Trace Elem Res 173(2):291–296. https://doi.org/10.1007/s12011-016-0665-8
Han F, Liu Y, Wang Q, Huang Z (2019) Effects on chromium and selenium in blood of rats to different serine intakes. Environ Chem 38(4):957–960
Huang D, Hu Q, Fang S, Feng J (2016) Dosage effect of zinc glycine chelate on zinc metabolism and gene expression of zinc transporter in intestinal segments on rat. Biol Trace Elem Res 171(2):363–370. https://doi.org/10.1007/s12011-015-0535-9
Lu Z, Wang P, Teng T, Shi B, Shan A, Lei XG (2019) Effects of dietary selenium deficiency or excess on selenoprotein gene expression in the spleen tissue of pigs. Animals (Basel) 9(12). https://doi.org/10.3390/ani9121122
Huang D, Zhuo Z, Fang S, Yue M, Feng J (2016) Different zinc sources have diverse impacts on gene expression of zinc absorption related transporters in intestinal porcine epithelial cells. Biol Trace Elem Res 173(2):325–332. https://doi.org/10.1007/s12011-016-0655-x
Mistry HD, Broughton Pipkin F, Redman CW, Poston L (2012) Selenium in reproductive health. Am J Obstet Gynecol 206(1):21–30. https://doi.org/10.1016/j.ajog.2011.07.034
Ning YJ, Wang X, Ren L, Guo X (2013) Effects of dietary factors on selenium levels of children to prevent Kashin-Beck disease during a high-prevalence period in an endemic area: a cohort study. Biol Trace Elem Res 153(1–3):58–68. https://doi.org/10.1007/s12011-013-9651-6
Zhou X, He L, Zuo S, Zhang Y, Wan D, Long C, Huang P, Wu X, Wu C, Liu G, Yin Y (2018) Serine prevented high-fat diet-induced oxidative stress by activating AMPK and epigenetically modulating the expression of glutathione synthesis-related genes. Biochim Biophys Acta Mol basis Dis 1864(2):488–498. https://doi.org/10.1016/j.bbadis.2017.11.009
Zhou X, He L, Wu C, Zhang Y, Wu X, Yin Y (2017) Serine alleviates oxidative stress via supporting glutathione synthesis and methionine cycle in mice. Mol Nutr Food Res 61(11). https://doi.org/10.1002/mnfr.201700262
Zhou X, Zhang H, He L, Wu X, Yin Y (2018) Long-term l-serine administration reduces food intake and improves oxidative stress and Sirt1/NFkappaB signaling in the hypothalamus of aging mice. Front Endocrinol (Lausanne) 9:476. https://doi.org/10.3389/fendo.2018.00476
Zhou X, Zhang Y, Wu X, Wan D, Yin Y (2018) Effects of dietary serine supplementation on intestinal integrity, inflammation and oxidative status in early-weaned piglets. Cell Physiol Biochem 48(3):993–1002. https://doi.org/10.1159/000491967
He LQ, Long J, Zhou XH, Liu YH, Li TJ, Wu X (2020) Serine is required for the maintenance of redox balance and proliferation in the intestine under oxidative stress. FASEB J 34:4702–4717. https://doi.org/10.1096/fj.201902690R
Funding
This study was funded by the Open Fund of Key Laboratory of Agro-ecological Processes in Subtropical Region, Chinese Academy of Sciences (ISA2018204), “Huxiang Young Talents Plan” Project of Hunan Province (2019RS2046), Hunan Provincial Science and Technology Department (2018TP1031), and Earmarked Fund for China Agriculture Research System (CARS-35).
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
The study was conducted according to the principles of the animal welfare committee of the Institute of Subtropical Agriculture, Chinese Academy of Sciences and was approved by the animal welfare committee of the Institute of Subtropical Agriculture, Chinese Academy of Sciences.
Conflict of Interest
The authors declare that they have no conflict of interest.
Informed Consent
Informed consent was obtained from all individual participants included in the study.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
About this article

Cite this article
Long, J., Liu, Y., Zhou, X. et al. Dietary Serine Supplementation Regulates Selenoprotein Transcription and Selenoenzyme Activity in Pigs. Biol Trace Elem Res 199, 148–153 (2021). https://doi.org/10.1007/s12011-020-02117-8
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s12011-020-02117-8