
Abstract
The present experiment aims at evaluating chronic toxic effects of the combination of cadmium (Cd) and molybdenum (Mo) according to residual element contents, apoptosis gene expression, and ultrastructure and histopathology changes of caprine spleen. In total, 36 Boer goats were randomly divided into four groups with the equal number in each group. The control group was orally administered with deionized water while the experimental groups I, II, and III were administered with the equal quantity of CdCl2 (1 mg kg−1 BW) and (NH4)6·Mo7O24·4H2O including 15, 30, and 45 mg·Mo kg−1 BW, respectively. Three individuals from each group were treated with euthanasia on days 0, 25, and 50. The data showed that the content of splenic residual Mo and Cd increased (P < 0.05) in the experimental groups on days 25 and 50, while no significant difference was observed in the content of Cu. The apoptosis-related gene expression levels including Bcl-2, Bax, Caspase-3, Smac, and ceruloplasmin (CP) were also determined. Results showed that significant reductions were observed in Bcl-2 and CP expressions (P < 0.01), while Caspase-3 gene was up-regulated (P < 0.05). However, no significant difference was observed in Smac and Bax expressions. Furthermore, on day 50, spleen tissues were presented to observe ultrastructural changes in lesions by means of transmission electron microscopy, with fragmentized nucleus, vesiculation of cytoplasm, mitochondria hyperplasia, and increasing lysosomes included. In addition, histopathology results corroborated the toxicity by showing cell hemorrhage, thickening central arteries, and enhanced capsule thickness. To sum up, our study revealed that the combination of Cd and Mo could induce remarkable damage to the spleen of goats by promoting cell apoptosis in the mitochondrial pathway and affecting the deposition of Mo and Cd.
Similar content being viewed by others


Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
Molybdenum (Mo) is a metal of vital industrial importance and is extensively applied in the steel industry. Moreover, Mo is an essential trace element for many animals and plants and has already been confirmed as a critical component for xanthine oxidase, aldehyde, and sulfite oxidase [1]. However, excessive dietary Mo intake turned out to be toxic for many species including rabbits, mice, and sheep [2–4]. In 1981, high-level Mo in feeding stuff was first discovered to be responsible for the persistent diarrhea in cattle, with features such as red skin and white hair, in southern Jiangxi Province, China [5]. Mo is traditionally regarded as playing a causative role in the secondary deficiency of copper and being poisonous when transformed to thiomolybdates [6, 7]. In addition, an overdose of Mo intake has been reported to result in kidney and liver damage by means of cell apoptosis [8, 9].
Cadmium (Cd) is an environmental toxicant worldwide, with its existence primarily resulting from industrial and agricultural emissions [10]. Chronic Cd accumulation in vivo leads to liver and spleen disorders [11]. Cd-induced reactive oxygen species could trigger oxidative stress damage, and the latter could impair spleen immune function by causing splenocyte apoptosis [12]. Also, some findings confirmed that mitochondrial pathway activation was another possible means inducing apoptosis in Cd toxicity [13, 14].
Because of the higher-than-permitted plant emission and surplus mining industry [15], livestock’s Mo and Cd exposure was more common in Jiangxi Province than in other parts of China, and the mixed intoxication of Mo and Cd occurred more frequently in Jiangxi, China. However, the relationship between these two elements remains a controversy at the present time. On the one hand, Mo may provide protection against the highly concentrated heavy metal (e.g., Cd) toxicity [16] whose mechanism is somehow linked to Cd-metallothionein induction enhancement [17]. A combination of Mo and sulfur in feeding stuff has showed its efficiency on reducing Cd accumulation in sheep tissues [18]. On the other hand, some previous studies reported that Mo and Cd might have a synergistic effect on kidney and testicle tissues [19, 20]. The objective of the current study was to evaluate the effects of the combination of Mo and higher level of Cd on caprine spleen and to explore the mechanism relationship between the two elements by studying apoptosis-related genes on mRNA levels and histopathological changes in vivo.
Materials and Methods
Animals and Treatments
Thirty-six clinically healthy Boer goats aging between 5 and 6 months and weighing from 15 to 20 kg were purchased from an intensively commercial farm with vaccination and deinsectization in advance. The goats were randomly divided into four groups with each group containing nine animals. They were housed for 2 weeks under sanitary conditions for acclimation before the experiment commencement and provided with feed and water ad libitum. Goats in the control group were orally administered with corresponding quantitative deionized water, while goats in the treatment groups were orally administered with identical levels of CdCl2 (Cd 1 mg kg−1 BW) and varying doses of ammonium molybdate [(NH4)6·Mo7O24·4H2O] (Mo 15 mg kg−1 for group I, 30 mg kg−1 for group II, and 45 mg kg−1 for group III). Basic diet for goats met the standard nutritional requirements for goat breeding set by the National Research Council [21]. The content of the basic diet and the content of Mo and Cd in grass, water, and fodder are listed in Tables 1 and 2. The whole experiment span was 50 days. All animal care and experimental procedures were approved by the institutional ethics committee, and this study also complied with the criteria in Guide for the Care and Use of Laboratory Animals.
Sample Collection
A total of 12 goats were randomly selected from four groups (three individuals in each group). Before sampling, they were performed with euthanasia with an overdose intravenous injection of sodium pentobarbital (100 mg kg−1, Nembutal, Abbot Labs, IL, USA), and subsequently, spleen tissues were collected instantly from 12 goats in vivo on days 0, 25, and 50, respectively.
Splenic Mo, Cd, and Cu Level Determination
The trace elements including Mo, Cd, and copper (Cu) in spleen were measured by using an Agilent 240 AA atomic absorption spectrophotometer (Agilent, USA) after wet-ashing the samples. All analyses were carried out according to the manufacturer’s instructions.
RT-PCR for Determining Apoptosis-Related Genes Expression in Spleen
Total spleen RNA was extracted by using TRIzol reagent according to the manufacturer’s instructions (Invitrogen, California, USA). The first-strand complementary DNA (cDNA) was synthetized with Titanium® RT-PCR Kit (Clontech, CA, USA) by strictly following the manufacturer’s instructions. Expression of apoptosis-related genes, including Bax, Bcl-2 and Caspase-3, Smac, Ceruloplasmin (CP), and β-actin, was evaluated according to previously published RT-PCR assays [22]. Relative expression was calculated according to RT-PCR efficiency formula reported [23]. The reference genes (β-actin) worked as an internal control for normalization of the results.
Histopathological Examination
The spleen samples were fixed in 10 % neutral-buffered formalin and embedded in paraffin. Sections with 5-μm thickness were incised from each block and stained with hematoxylin and eosin (H&E). After that, the optical microscope was used to observe the histopathological results.
Transmission Electron Microscopy
Transmission electron microscopy (TEM) studies were performed based on the protocol as previously mentioned [24]. After removal of spleen samples, they were performed and observed under TEM Zeiss 900 (Zeiss, Germany). The ultrastructural pathological changes in splenocytes were compared with the counterparts of the control group.
Statistical Analysis
The experiments were performed in triplicate, and data were analyzed with the software Microsoft Excel 2007, OriginPro 9 (OriginLab Corporation, Northampton, MA, USA), SDS 2.4, and SPSS version 17.0 (SPSS Inc., Chicago, IL, USA). All experimental data were tested by the analysis of variance with significant differences between means determined by Duncan post hoc tests. The data were represented as mean ± SD. Differences were considered significant. Statistical significance level was set at a P value < 0.05.
Results
Mo, Cu, and Cd Content in Spleen
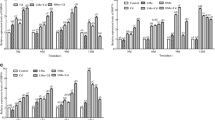
Splenic Mo, Cd, and Cu concentrations are shown in Fig. 1. Splenic Mo concentration was supposed to be positively connected with Mo’s intake. Three experimental groups showed increases on days 25 and 50, and they were statistically different among each other as administered Mo concentration increased. Cd content was boosted significantly (P < 0.05) in all experimental groups on day 50. Surprisingly, Cu concentrations in the four groups were not significantly different on days 25 and 50.

Residual content of Mo, Cd, and Cu in the splenocytes of goats. Capital letters stand for intra-group comparison and small letters stand for inter-group comparison. Different letters mean statistical difference (P < 0.05). It is the same below
The Relative mRNA Expression of Apoptosis-Related Genes
The data are illustrated in Fig. 2. Bcl-2 mRNA expression in group II significantly decreased (P < 0.05) on day 25 compared with that in the control group, and its expression reduced (P < 0.01) in all experimental groups on day 50. Furthermore, CP mRNA expression in spleen plunged on day 25 (P < 0.05) while that in all experimental groups significantly plummeted when compared with that in the control group on day 50 (P < 0.05). Interestingly, no significant difference was observed in the Bax and Smac expressions on day 25 and 50. Moreover, Caspase-3 mRNA expression in groups I and III were found to have been increased (P < 0.05) on day 50.

Determination of the mRNA expression of apoptosis-related genes in the goat spleen
Histopathological Results
No abnormal architecture was found in spleens from the control group, including regular central artery and capsule thickness (Fig. 3a, b). Samples in group I showed slight spleen lesions, with evident hemorrhage spots among splenocytes (Fig. 3c, d). In group II, spleens showed more serious degree of hemorrhage and thickening central artery layer (Fig. 3e, f). In addition, samples in group III showed an increase of capsule thickness and the most serious hemorrhage degree (Fig. 3g, h).

Histological results of the spleen in goats on day 50. Control group (a, b). Group I (c, d). Group II (e, f). Group III (g, h)
TEM Observation
Electron microscope revealed splenocytes’ ultrastructural changes induced by toxic effects of different levels of Mo and a certain level of Cd. The spleen cell’s integrative organelles in the control group are shown in Fig. 4a. On day 50, decreased organelle numbers, nucleus deformation, and some high-density electron bodies were observed in group I (Fig. 4b). Pathological lesions are more serious in group II fed by Cd and mediate level of Mo (30 mg kg−1) with apoptosis phenomenon including nuclear deformation and the vesiculation of mitochondrion whose cristaes and membranes also started to rupture and even dissolve (Fig. 4c). Figure 4d illustrated the effect of Cd with high level of Mo (45 mg kg−1) on splenocyte, which demonstrated a more noticeable cell apoptosis with fragmentized nucleus and vesiculation of cytoplasma. In addition, some mitochondria dissolved to a large extent, and mitochondria hyperplasia and increasing numbers of lysosomes were also presented.

Ultrastructural images in the spleen cell of experimental goats on day 50. a Control group. b Group I.c Group II. d Group III.(Note: N nuclei, Mito mitochondria, ER endoplasmic reticulum, Lyso lysosome)
Discussion
Affluent metal mineral resources in Jiangxi Province plus excessive mineral mining raised environmental problems and made livestock and birds under risky circumstances including Mo and Cd. High level of Mo in feeding stuff was first discovered to cause animals’ persistent diarrhea, and “Red Skin and White Hair” syndromes in southern Jiangxi Province turned out to be a result of the combination of Mo and Cd rather than that of Mo alone [5], which made our research meaningful for local animal breeding. In the present study, residual element contents, apoptosis gene expression, and TEM were determined in order to evaluate the relationship between Mo and Cd on spleen cell apoptosis. Residual Mo and Cd concentration in spleen were positively linked with experimental time and administered dose. Besides, residual Cu concentration and CP expression were determined for splenic Cu metabolism evaluation. CP is a Cu-containing glycoprotein whose main function is to transfer Cu to the blood, which means Cu is mainly used in the body [25]. Therefore, CP expression is used as a reliable indicator of Cu nutrition status in vivo. In the current study, CP expression decreased as the experimental time lasted. This could potentially be explained by the fact that an overtake of Mo may bound Cu in the Cu-Mo complex which is biologically unavailable in vivo [26], which shows available Cu absorption disturbance and Cu excretion disorder. However, the biological Cu-Mo antagonism was not self-evident according to the data. Thiomolybdates are supposed to form in the rumen by a reaction between sulfide and molybdate, binding with Cu or Cu proteins in the gastrointestinal tract and in the blood and tissues to render Cu unavailable for normal absorptive or enzymic functions [27]. Cd was reported to promote the complex with metallothionein [28]. Cd-metallothionein was assumed to play a potentially causative role in removing Cu from the Cu-Mo-S complex and changing into absorbable forms [22]. The results suggested that the combination of Mo and Cd could possibly impair the splenocyte function of Cu’s absorption and excretion; the data also indicated that the combination of Mo and Cd could initiate splenic antioxidant defense reduction and inhibit anti-oxidant indicators which also induce cell apoptosis.
Mitochondria play key roles in inducing apoptosis in ruminants’ cells [29]. Mitochondrial malfunction is regarded as an early sign of cell apoptosis. Mitochondria-induced apoptosis begins with upstream Caspase activation and is regulated through members of the Bcl-2 family. Bcl-2, an oncogene belonging to the anti-apoptotic family, was expected to regulate cell apoptosis at mitochondrial level, suppressing apoptotic death [30]. However, suppression of prosurvival Bcl-2 proteins is insufficient to kill cells in the absence of Bax [31]. Bax, a proapoptotic gene, was also believed to be involved in cell apoptosis by boosting the permeability of the outer mitochondrial membrane and promoting efflux of apoptotic proteins into the cytoplasm. The apoptotic cascade involving in the activation of Caspase-3 was considered a downstream event in the mitochondrial pathway [32]. In the current study, the toxicity of a constant Cd with varying levels of Mo significantly down-regulated Bcl-2 expression but promoted Caspase-3 expression, with Bax and Smac remaining relatively unchanged. As for the proapoptotic gene, no remarkable difference was observed among the four groups in Bax, while Bcl-2 decreased significantly. This is partly different from some previous reports saying dietary Mo could enhance Bax protein and decrease Bcl-2 protein presence [33]. However, this could be explicable if the ratio of Bcl-2/Bax constitutes a rheostat model for apoptosis susceptibility evaluation [34]. The inhibition of apoptosis and the ratio of Bcl-2/Bax are somehow linked with the mitochondrial permeability transition pores, which is indispensable for involved genes’ release and therefore affects mitochondrial pathway apoptosis process [35]. In addition, Caspase-3 expression was elevated to be in agreement with previous studies [36, 37]. No significant difference was found on Smac. This proapoptotic gene was reported to induce apoptosis through binding to inhibitors of apoptosis, relieving inhibitory effects and activating on Caspase-3 activity [29, 38]. This may indicate that the spleen cells under Mo and Cd stress tend to resist apoptosis caused by dietary Mo and Cd. In total, changes in Bcl-2 and Caspase-3 suggested that the damage could involve the mitochondrial pathway in vivo.
Besides, splenic ultrastructural observation suggested that different levels of Mo and Cd induced splenic morphological changes by means of apoptosis justified by nucleus deformation, vacuolation in cytoplasma and mitochondria, as well as increasing lysosomes in splenocyte. Because of the mitochondrial damage, a series of proteins were released into the cytoplasm accelerating apoptosis execution [29]. The histological results from the experimental group showed spleen lesions with cell hemorrhage, thickening central arteries, and enhanced capsule thickness. These pathological changes verified the results above showing that Mo may present a synergistic effect on Cd in terms of spleen toxicity.
Conclusion
In conclusion, the study revealed that the combination of Mo and Cd could aggravate spleen cell apoptosis through the mitochondrial pathway and induce pathological lesions whose mechanism is somehow linked with Mo, Cd, and Cu deposition in spleen.
References
Schwarz G, Mendel RR, Ribbe MW (2009) Molybdenum cofactors, enzymes and pathways. Nature 460(7257):839–847. doi:10.1038/nature08302
Arrington LR, Davis GK (1953) Molybdenum toxicity in the rabbit. J Nutr 51(2):295–304
Fairhall LT, Dunn RC, Sharpless NE, Prichard EA (1945) The toxicity of molybdenum. Public Health Bull 293:1–36
Hogan KG, Money DFL, White DA, Walker R (1971) Weight responses of young sheep to copper, and connective tissue lesions associated with the grazing of pastures of high molybdenum content. N Z J Agric Res 14(3):687–701. doi:10.1080/00288233.1971.10421662
Fan P, Wu ZL, Wang JY, Liu X, Dai QW, Pan ZJ, Lang YX, Hu YX, Liu TF, Liu YL (1981) A preliminary report on molybdenosis of cattle. J Jiangxi Agric Univ 01:5–7(in Chinese)
Frank A (2004) A review of the "mysterious" wasting disease in Swedish moose (alces alces L.) related to molybdenosis and disturbances in copper metabolism. Biol Trace Elem Res 102(1–3):143–159. doi:10.1385/bter:102:1-3:143
Nederbragt H (1980) The influence of molybdenum on the copper metabolism of the rat at different Cu levels of the diet. Br J Nutr 43(2):329–338
Zhuang Y, Liu P, Wang L, Luo J, Zhang C, Guo X, Hu G, Cao H (2015) Mitochondrial oxidative stress-induced hepatocyte apoptosis reflects increased molybdenum intake in caprine. Biol Trace Elem Res. doi:10.1007/s12011-015-0450-0
Gu X, Ali T, Chen R, Hu G, Zhuang Y, Luo J, Cao H, Han B (2015) In vivo studies of molybdenum-induced apoptosis in kidney cells of caprine. Biol Trace Elem Res 165(1):51–58. doi:10.1007/s12011-015-0238-2
Satarug S, Garrett SH, Sens MA, Sens DA (2010) Cadmium, environmental exposure, and health outcomes. Environ Health Perspect 118(2):182–190. doi:10.1289/ehp.0901234
Godowicz B (1988) Cadmium-induced liver and spleen damage in inbred mouse strains. Folia Biol 36(1–2):47–51
Pathak N, Khandelwal S (2006) Oxidative stress and apoptotic changes in murine splenocytes exposed to cadmium. Toxicology 220(1):26–36. doi:10.1016/j.tox.2005.11.027
Li M (2003) Cadmium directly induced the opening of membrane permeability pore of mitochondria which possibly involved in cadmium-triggered apoptosis. Toxicology 194(1–2):19–33. doi:10.1016/s0300-483x(03)00327-5
Liu S, Xu FP, Yang ZJ, Li M, Min YH, Li S (2014) Cadmium-induced injury and the ameliorative effects of selenium on chicken splenic lymphocytes: mechanisms of oxidative stress and apoptosis. Biol Trace Elem Res 160(3):340–351. doi:10.1007/s12011-014-0070-0
Dai QW, Zeng ZM, Wang JY, Wu ZL, Fan P (1993) A preliminary investigation on effect of mine in major metal factory on animal husbandry. Agri-Environmental Protect 12:124–126(in Chinese)
Rautio A, Kunnasranta M, Valtonen A, Ikonen M, Hyvarinen H, Holopainen IJ, Kukkonen JV (2010) Sex, age, and tissue specific accumulation of eight metals, arsenic, and selenium in the European hedgehog (Erinaceus europaeus). Arch Environ Contam Toxicol 59(4):642–651. doi:10.1007/s00244-010-9503-8
Yasuhiro Yamane MF, Li C, Koizumi T (1990) Protective effect of sodium molybdate against the acute toxicity of cadmium chloride. Toxicology 60(3):235–243
Smith GM, White CL (1997) A molybdenum–sulfur–cadmium interaction in sheep. Aust J Agric Res 48:147–154
Xia B, Cao H, Luo J, Liu P, Guo X, Hu G, Zhang C (2015) The Co-induced effects of molybdenum and cadmium on antioxidants and heat shock proteins in duck kidneys. Biol Trace Elem Res. doi:10.1007/s12011-015-0348-x
Xia B, Chen H, Hu G, Wang L, Cao H, Zhang C (2015) The Co-induced effects of molybdenum and cadmium on the trace elements and the mRNA expression levels of CP and MT in duck testicles. Biol Trace Elem Res. doi:10.1007/s12011-015-0410-8
NRC NRC (1981) Nutrient requirements of goat: Angora, dairy and meat goats in temperate and tropical countries. Nutrient requirements of domestic animals, number-15. National Academic Press, Washington DC, USA, pp:2–12
Gu X, Chen R, Hu G, Zhuang Y, Luo J, Zhang C, Guo X, Huang A, Cao H (2015) Cell apoptosis of caprine spleen induced by toxicity of cadmium with different levels of molybdenum. Environ Toxicol Pharmacol 40(1):49–56. doi:10.1016/j.etap.2015.05.001
Pfaffl MW, Horgan GW, Dempfle L (2002) Relative expression software tool (REST) for group-wise comparison and statistical analysis of relative expression results in real-time PCR. Nucleic Acids Res 30(9):e36
Antunez MI, Penco JM, Lozano JL, Macho AL, Climent V (2011) Influence of intestinal resections on biliary composition and liver ultrastructure. Clin Nutr 30(2):247–251. doi:10.1016/j.clnu.2010.09.002
Cantin A, Crystal RG (1985) Oxidants, antioxidants and the pathogenesis of emphysema. Eur J Respir Dis Suppl 139:7–17
RICHARD P, DOWDY GM (1967) A copper-molybdenum complex: its effects and movement in the piglet and sheep. J Nutr 95:197–201
White GMSaCL (1997) A molybdenum-sulfur-cadmium interaction in sheep. Aust J Agric Res 48:147–154
Godt J, Scheidig F, Grosse-Siestrup C, Esche V, Brandenburg P, Reich A, Groneberg DA (2006) The toxicity of cadmium and resulting hazards for human health. J Occup Med Toxicol 1:22. doi:10.1186/1745-6673-1-22
Wang C, Youle RJ (2009) The role of mitochondria in Apoptosis*. Annu Rev Genet 43:95–118. doi:10.1146/annurev-genet-102108-134850
Hockenbery DNG, Milliman C, Schreiber R, Korsmeyer SJ (1990) Bcl-2 is an inner mitochondrial membrane protein that blocks programmed cell death. Nature 348:334–336
Doerks T, Copley RR, Schultz J, Ponting CP, Bork P (2002) Systematic identification of novel protein domain families associated with nuclear functions. Genome Res 12(1):47–56
Korsnes MS, Hetland DL, Espenes A, Aune T (2006) Induction of apoptosis by YTX in myoblast cell lines via mitochondrial signalling transduction pathway. Toxicology in Vitro. Int J Published Associ BIBRA 20(8):1419–1426. doi:10.1016/j.tiv.2006.06.015
Xiao J, Cui HM, Yang F, Peng X, Cui Y (2011) Effect of dietary high molybdenum on the cell cycle and apoptosis of kidney in broilers. Biol Trace Elem Res 142(3):523–531. doi:10.1007/s12011-010-8772-4
Danial NN, Korsmeyer SJ (2004) Cell death: critical control points. Cell 116(2):205–219
Bagci EZ, Vodovotz Y, Billiar TR, Ermentrout GB, Bahar I (2006) Bistability in apoptosis: roles of bax, bcl-2, and mitochondrial permeability transition pores. Biophys J 90(5):1546–1559. doi:10.1529/biophysj.105.068122
Wang Y, Wu Y, Luo K, Liu Y, Zhou M, Yan S, Shi H, Cai Y (2013) The protective effects of selenium on cadmium-induced oxidative stress and apoptosis via mitochondria pathway in mice kidney. food and chemical toxicology. Int J Published Br Ind Biol Res Assoc 58:61–67. doi:10.1016/j.fct.2013.04.013
Lopez E, Figueroa S, Oset-Gasque MJ, Gonzalez MP (2003) Apoptosis and necrosis: two distinct events induced by cadmium in cortical neurons in culture. Br J Pharmacol 138(5):901–911. doi:10.1038/sj.bjp.0705111
Kashkar H, Haefs C, Shin H, Hamilton-Dutoit SJ, Salvesen GS, Kronke M, Jurgensmeier JM (2003) XIAP-mediated caspase inhibition in Hodgkin’s lymphoma-derived B cells. J Exp Med 198(2):341–347. doi:10.1084/jem.20021279
Acknowledgments
The research was supported by the Program of the National Natural Science Foundation (No. 31101863, Beijing, P.R. China), Training Plan for Young Scientists of Jiangxi Province (No. 2014BCB23040, Nanchang, P.R. China), and Department of Education of the Science Foundation of Jiangxi Province (No. GJJ14294, Nanchang, P.R. China) to Huabin Cao.
Author information
Authors and Affiliations
Corresponding authors
Ethics declarations
Conflict of Interest Statement
The authors declare that there are no conflicts of interest.
Additional information
Qingyang Xiao, Caiying Zhang, and Xiaolong Gu are the equal first authors.
All authors have read the manuscript and agreed to submit it in its current form for consideration for publication in the Journal
Rights and permissions
About this article

Cite this article
Xiao, Q., Zhang, C., Gu, X. et al. Varying Dietary Levels of Molybdenum Inducing Cell Apoptosis of Spleen Under Cadmium Stress in Caprine. Biol Trace Elem Res 172, 127–133 (2016). https://doi.org/10.1007/s12011-015-0565-3
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s12011-015-0565-3