
Abstract
The biorefinery technology aiming at protein extraction is rising and identification of suitable plant biomass input with valuable protein compounds for extraction is needed. Forage crops have been evaluated by the Cornell Net Carbohydrate and Protein System (CNCPS), and the result used as proxy of extractable protein in a biorefinery process. This serves as a helpful link between crop production and refinery output; however, the method has never been validated. Such validation is the main aim of this study. Five forage species—white clover, red clover, lucerne, perennial ryegrass, and tall fescue—were cut at four dates during spring and processed in a lab-scale refinery (screw press and subsequent protein precipitation from the green juice). The pulp fraction and the precipitated protein concentrate were both CNCPS analyzed to follow the initial crude protein (CP) plant input into these two fractions. Total recovery in concentrate was highest for the legumes, which points to an advantage of these species in protein extraction setups. High recovery of B1 and B2 (50% or higher for the grasses) in the pulp demonstrated a large proportion of soluble protein ending up in the fibrous pulp and shed light on the reason behind high feed quality of the pulp fraction. In conclusion, the existing tentative assumption of extractable protein being equal to CNCPS fractions of B1 and B2 and partly B3 was shown to be too simplified. The presented findings can improve crop species screening in terms of expected extractable protein yield.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
The increased demand for protein globally entails a growing awareness of sustainability of protein sources as well as increased focus on developing new sources [1, 2]. Green forage plants such as legumes and grasses have long been used as high-quality protein sources for ruminants, while monogastrics generally cannot efficiently utilize these protein sources [3, 4]. The nature of cultivation of perennial forage crops comprises potential improvement of crop rotation systems as well as environmental benefits in terms of high carbon sequestration and low nitrogen (N) leaching and need for pesticides as compared to for instance cereals [5, 6]. In order to process forage crop protein for wider utilization purposes, the biorefinery technology appears promising [7]. The biorefinery separates the protein from the plant fiber matrix into a green protein concentrate suited for monogastric digestion in qualities comparable to soybean protein and a fibrous pulp fraction, which has shown good results as ruminant feed [7, 8] or for biogas [9].
The full production chain from soil to biorefinery output products comprises the crop production, the refinery process technology, and the quality of the end products, each more or less isolated in their field of science [10,11,12]. Nevertheless, in order to optimize the overall output and efficiency, this knowledge needs to be linked. Considering the agricultural part, namely the biomass cultivation, an emerging effort is put into investigating the optimal choice of crop and management for maximized biomass and protein input to the biorefinery operation [6, 10]. These studies largely resemble previous studies examining forage crop protein concentration for ruminant feed quality [13,14,15]. The plant CP content could presumably be linked to biorefinery protein output by applying the Cornell Net Carbohydrate and Protein System (CNCPS), which is a widely applied and internationally recognized analysis [16, 17]. The method divides the crude protein (CP) into five fractions according to the type of N compound and degradability. The nomenclature of the CNCPS has changed over the years and according to the system concept described in 2008 [18], the CP fractions are divided into A, B1, B2, B3, and C, where especially B fractions are interesting as A is non-protein N and C is cell wall–bound N. The application of CNCPS analysis of the biomass may serve as a screening of biomass and thereby a link between crop production research and the final refinery protein output and quality. A suggested interpretation of the CNCPS protein fractions in biomass and potential protein extraction entails that B1 and B2 fractions are expected to be directly extractable whereas B3 is potentially extractable [6]. However, an inaccuracy exists regarding this assumption since no study previously validated this relationship.
The aim of this study was to examine the relationship between the forage crop biomass CP contents and CNCPS fractions and the biorefinery output in terms of pulp and protein concentrate, which were correspondingly analyzed by the CNCPS method. This was done for a variety of forage species cut on consecutive dates in the spring, hereby providing novel insights into the destiny of each of the CNCPS CP fractions in the biomass throughout the biorefinery process to the end products. We hypothesized that the B1 and B2 fractions would largely constitute the protein that distributed into the protein concentrate from biorefinery, and that the recovery of specific CNCPS fractions in pulp and protein concentrate would differ significantly between forage species of different plant functional groups (grasses and legumes).
Materials and Methods
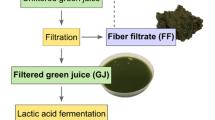
The overall experimental design includes a number of steps (Fig. 1), including a field trial (biomass source), lab-scale biorefinery processing, and chemical analyses of biomass input and refinery output according to the CNCPS method.

The overall experimental design showing the flow of plant samples from field trial to refinery output. The initial plant biomass as well as the pulp and protein concentrate refinery outputs is analyzed according to the CNCPS method into CP fractions (A, B1, B2, B3, and C) in order to follow the distribution of these fractions throughout the process. For the brown juice, the CP fractions of A, B1, B2, B3, and C are not measured but calculated. The refinery process includes the screw press and protein precipitation steps
Field Trial Design and Management
The samples examined in this study were taken in 2016 in second production year of a grassland experiment established at Aarhus University, Foulumgaard (56° 30′ N, 9° 35′ E). The soil is classified as a Typic Hapludult, according to the USDA Soil Taxonomy System with 7% clay, 10% silt, 81% sand, and 1.7% C in the topsoil (0–20 cm). The soil pH was 5.9 and concentrations of extractable P, exchangeable K, and exchangeable Mg were 36, 129, and 39 mg kg−1 soil, respectively.
The legume species were white clover (Trifolium repens L., cv. Silvester), red clover (Trifolium pratense L., cv. Suez), and lucerne (Medicago sativa L., cv. Creno) and the grass species were perennial ryegrass (Lolium perenne L., cv. Calvan 1) and tall fescue (Festuca arundinacea L., Tower) under sown in a spring barley (Hordeum vulgare L.) main crop in 2014. All seeds used in the trial were acquired from DLF, Seeds and Science (Roskilde, Denmark). The seed rate was 10 kg ha−1 for white clover and red clover, 20 kg ha−1 for lucerne, and 25 kg ha−1 for the grasses. Grasses were applied 140 kg N ha−1 in mineral fertilizer. Field plots (1.5 × 12 m) were arranged in a split plot design (harvest time as main plot and species as sub‐plot) with two replications.
The plots were harvested at 7-cm stubble height using a Haldrup plot harvester (J. Haldrup a/s, Løgstør, Denmark) at four dates in 2016 (May 17, May 24, May 31, and June 6). A representative sub‐sample of approximately 2–3 kg was taken manually from the harvested plant material, and stored at − 18 °C until further analysis.
The field trial was conducted in accordance to local legislation and no further permissions or licenses were required.
Protein Extraction and Precipitation
The plant material was thawed overnight prior to processing, which was done in a commercial lab-scale by using twin-screw press (Angelia 8500S, Angel Juicer, Seoul, South Korea) as previously described [4]. Processing resulted in separation of the plant material into a pulp and a juice. The juice was acidified to pH 4 using 12 M phosphoric acid resulting in precipitation of soluble proteins. The acidified juice was left overnight at 4 °C and centrifuged for 10 min at 2000g at 4 °C during which the precipitated protein was separated from the supernatant liquid (brown juice). All samples were freeze-dried and ground (0.5 mm) before further analysis.
Biorefinery Outputs
Each of the two refinery outputs (i.e., pulp and protein concentrate) was quantified and qualified by mass balances and the corresponding content of CNCPS CP fractions, whereas the supernatant output was calculated by difference. To reconcile the mass balance, truncation was used in order to match input and output in case of inconsistency, mainly caused by losses in the screw press. This was done relative to output product proportions. The truncation was performed twice during calculations. The first time was in relation to the DM distribution between outputs. The second time was performed to reconcile the output of the specific CNCPS CP fractions, so that for instance the summed B2 output in pulp and protein concentrate did not exceed the initial plant content. The result from this procedure was inevitably that the results on the supernatant part would be a combination of real material, process losses, and uncertainties in relation to calculations, why this output will be named supernatant/loss hereafter.
Analysis for Protein Fractions (CNCPS)
Sub‐samples of 200–300 g of biomass samples from the field trial were freeze-dried and 50–60 g of dried samples was collected and ground to 1-mm particle size in a Foss Cyclotec 1093 mill. Ground samples were stored in airtight bottles at room temperature until further analysis. Crude protein was divided into five fractions adhering to the CNCPS nomenclature and method described by Licitra et al. (1996) and Tylutki et al. (2008). Correspondingly, the analysis was performed on the pulp and protein concentrate output. Finally, all CP measurements were carried out using the Kjeldahl method with a nitrogen-to-CP conversion factor of 6.25. This study was conducted in connection to a parallel study investigating N recovery in pulp and protein concentrate, however, without considering the CNCPS fractions of neither refinery input nor output [19]. The plant samples were handled and identified by technician Margit Paulsen and sub-samples were deposited according to standard lab procedures.
Statistical Analyses
The statistical software used in this study was R [20]. Test of differences in recovered CP in pulp and extracted protein between species was performed by the lme mixed linear model in the nlme package [21] followed by a post hoc Tukey test (α = 0.05) using the glht function in the multcomp package [22]. Harvest date was included as a random factor. Transformation of data from recovered C fraction in extracted protein and B1, B2, and B3 fractions in the pulp was performed (square root, square root, log, and quartered, respectively).
Results
To increase robustness of the results in this first-of-its-kind study, we choose to collapse the four spring cut dates resulting in eight replicates per species. Hereby, the data represent more variation within plant physiological development and thus strengthen the reliability of concluded differences between species, which is one of the main aims of the study (see Electronic Supplementary Material for information disaggregated on cut dates).
The DM contents of the plant material ranged from 12.6 to 20.5% and CP content from 145 to 217 g kg−1 across the five species (Table 1, Fig. 2). The B2 fraction constituted the highest CP fraction content in all plant species across all cut dates followed by fraction A (Fig. 2, S1). The proportional order of B1, B3, and C alternated between species, although fraction C generally showed the lowest concentration. The relative distribution of CNCPS fractions of total CP for the five species (species showing lowest and highest values in parenthesis) ranged from 9.7 to 16.2% for B1 (perennial ryegrass; lucerne), from 40.6 to 48.6% for B2 (red clover; lucerne), from 4.7 to 17.9% for B3 (lucerne; perennial ryegrass), from 4.1 to 6.5% for C (white clover; red clover), and from 22.9 to 26.2% for A (perennial ryegrass; red clover) (Fig. 3).

CP content in plant biomass and the distribution of this CP into the three outputs: pulp, concentrate, and supernatant/loss divided on the five CNCPS fractions, for five forage species. Forage species are white clover (W.C.), red clover (R.C.), lucerne (Luc.), perennial ryegrass (Per.Ry.), and tall fescue (Tall Fe.). For example, the three black bars (B2 content) of the red clover pulp, red clover concentrate, and red clover supernatant/loss sum to the black bar representing the red clover plant. Individual bar colors (from beneath) represent B1, B2, B3, C, and A, according to the CNCPS method. The bars show results averaged across four spring cut dates. CP contents for the supernatant/loss fraction are calculated as difference. The order of fractions is changed to facilitate the overview of monogastric digestible CP fractions, which has been argued to be B1, B2, and partly B3

CNCPS fraction distribution of total crude protein in the plant biomass, pulp, protein concentrate, and supernatant/loss for five forage species. Individual bar colors (from beneath) represent B1, B2, B3, C, and A, according to the CNCPS method. Forage species are white clover (W.C.), red clover (R.C.), lucerne (Luc.), perennial ryegrass (Per.Ry.), and tall fescue (Tall Fe.). The bars show results averaged across four spring cut dates. The order of fractions is changed to facilitate the overview of monogastric digestible CP fractions, which has been argued to be B1, B2, and partly B3
The DM content of the pulp fractions ranged from 28.0 to 42.7% and CP content from 92 to 164 g kg−1 DM (Table 1). The CP recovery in the pulp ranged from 34 to 48% of plant CP across the four cut dates and five forage species (Table 2). The grasses showed significantly higher recovery as compared to red clover and lucerne and especially the recovery of the B3 fraction drove this difference. Of the soluble B1 and B2 CP fractions, between 8 and 37% and between 39 and 55% of B1 and B2, respectively, were recovered in the pulp. The relative distribution of CNCPS fractions of total CP in the pulp for the five species (species showing lowest and highest values in parenthesis) ranged from 0.6 to 7.6% for B1 (tall fescue; lucerne), from 46.0 to 59.1% for B2 (red clover; white clover), from 5.7 to 31.3% for B3 (lucerne; perennial ryegrass), from 2.3 to 6.0% for C (perennial ryegrass; lucerne), and from 15.3 to 22.0% for A (red clover; lucerne) (Fig. 3). This showed that B2 generally constituted the main proportion, although the B3 fraction tangented the B2 level on some cut dates for red clover and perennial ryegrass (see Fig. S1). As compared to the CNCPS fraction distribution in the plant, the pulp had relatively less or similar B1 and A content, and more or similar B2 and B3 content pointing at an overall increased proportion of soluble proteins in the pulp.
For the protein concentrate, the DM contents were from 15.3 to 18.3% and CP content from 266 to 336 g kg−1 DM (Table 1). Total CP recovery in the protein concentrate fraction in proportion of plant CP ranged from 23 to 39% with the legumes having significantly higher recovery than the grasses (Table 2). The driver behind this was mainly a systematically higher B2 recovery in the legumes (51 to 58%) as compared to the grasses (29 to 33%) (for recovery-dynamics across the four spring cut dates, see Fig. S1). The relative distribution between CNCPS fractions in the protein concentrate for the five species (species showing lowest and highest values in parenthesis) ranged from 7.0 to 10.7% for B1 (perennial ryegrass; red clover), from 58.4 to 69.7% for B2 (perennial ryegrass; white clover), from 0.9 to 18.4% for B3 (lucerne; perennial ryegrass), from 0.7 to 1.2% for C (lucerne; white clover), and from 12.2 to 19.6% for A (red clover; tall fescue) (Fig. 2). The summed B1 and B2 fractions comprised at least 65% of the concentrate CP revealing a general picture of relatively decreased A content and increased B2 content in the concentrate compared to the plant.
Higher or equal amounts of total CP were recovered in the pulp as compared to the protein concentrate (except for lucerne) (Fig. 2). The supernatant/loss held between 25.8 and 33.7% of plant CP (mean = 29.0%) and contained a mix of the five CP fractions, although no (or very low levels) B2 was recovered for the legumes and no B3 was recovered for the grasses.
The mass balance of the specific CNCPS CP revealed a systematic surplus in the summed B2 content in pulp and concentrate (mean = 32%; data not shown) as compared to the initial plant B2 input in the legumes, whereas a systematic surplus in the summed B3 content in pulp and concentrate (mean = 84%; data not shown) as compared to the initial plant B3 input in the grasses. This overload was reduced by the second step of the truncation procedure (see the Materials and Methods section “Biorefinery Outputs”).
Discussion
Recovery
The results revealed differences between species in CP recovery of some of the five CNCPS fractions as well as total CP recovery for both pulp and protein concentrate. Nevertheless, in the pulp, the only systematic differences found were higher proportion of B3 and total CP in the grasses compared to legumes (except white clover for total CP). Higher CP recovery in pulp from grasses compared to legumes was also shown by Stødkilde et al. (2020) using the Dumas procedure for overall CP determination. The high recovery of B1 and B2 (50% or higher for the grasses) in the pulp showed that soluble protein does not necessarily end up in the protein concentrate. The pulp contained from 57 to 72% water in the present study, and therefore it can be argued that solutes will naturally also distribute to the pulp output. The inability to perform a complete separation of soluble and insoluble protein during processing is in agreement with previous findings, where similarities between amino acid compositions of the pulp and protein concentrate suggest that the fiber-rich pulp contains a proportion of soluble proteins enclosed in the fiber network [4, 23]. These findings nuance the existing assumption that the plant B1, B2, and possibly B3 content represent a proxy for estimated extractable true protein [10].
Considering the CP recovery in the protein concentrate, the grasses differ from the legumes in terms of higher B3 recovery, except for red clover. Total recovery was systematically higher for the legumes, which points to an advantage of these species in protein extraction setups, although this has to be viewed in a broader perspective also including, e.g. biomass yields and environmental impact of cultivation. In addition, the quality as defined by the CNCPS fractions is also important, which will be treated further below. Stødkilde et al. (2020) came to the same conclusion regarding higher total CP recovery in the legumes and further found a significant correlation between total CP plant content and CP recovery in the concentrate, which assumingly represents the same pattern as shown by the data presented in this study. However, recovery percentages reported from this study are generally lower for both pulp and protein concentrates as compared to Stødkilde et al. (2020), which will be discussed in section “CP Fraction Shift During Processing”. Santamaría-Fernández et al. (2019) found CP recovery in the protein concentrate from red clover to be approximately 40%, which is roughly in line with the 35% found in this study for red clover, whereas they found recovery in timothy (a grass species) to be approximately 25%.
Quality of Pulp and Concentrate
It has previously been shown that the pulp has a feeding quality similar to the initial crop biomass, based on the CP content and feeding experiments [8, 23]. In this study, we found consistently less CP in the pulp as compared to pre-treated biomass. Stødkilde et al. (2019) reported CP content of the pulp to be on average 12.6% lower than CP plant content across the four investigated species—white clover, red clover, lucerne, and perennial ryegrass—whereas this study found a corresponding decrease of 33.3% as an average for the same four species. This could be explained (at least partly) by the truncation issue as discussed below, pointing at a shift of CP between CNCPS fractions during the process. Interestingly, the new knowledge on CP quality provided by this study confirms that the relative distribution between CNCPS fractions in the pulp resembles the one in the crop biomass when it comes to the important components of B1 and B2 for all species. The summed B1, B2, and B3 in the pulp were on similar levels as the crop biomass for all species. Damborg et al. (2020) reported total amino acid content in plant and pulp protein CP to be 76.8% and 82.5% of total CP, thereby showing an amino acid increase in pulp as compared to plant biomass, although that method does not clarify the distribution between A, B1, B2, B3, and C in the outputs. Our results shed light on the reason behind high feed quality of the pulp fraction in terms of the somewhat surprising CNCPS fraction distribution in the pulp.
Regarding the protein concentrate, the findings indicate that the quality is improved compared to the initial forage crop biomass, since the concentrate had a higher proportion of B2. This fraction containing soluble true protein is expected to have a higher digestibility than the cell wall–bound C fraction and would hence be more utilizable by the monogastric animal. The recovery of non-protein N (A fraction) in the protein concentrate can be compared to the previously reported results by Damborg et al. (2018) of 12%, whereas Stødkilde et al. (2019) reported values as low as 1% or less.
CP Fraction Shift During Processing
The proportion of plant CP ending up in the supernatant/loss was (averaged for each species) 25.8–33.7% (mean = 29.0%), whereas Stødkilde et al. (2020) found it to be 8.6 to 17.7% (mean = 12.2%) for the corresponding treatment (single screw pressed). These differences can be caused by increased uncertainty on CP contents when quantified as the summed content of the five CNCPS fractions as compared to the Dumas method for total N content. Moreover, the truncation used in this study is likely to play a role. When the summed CP contents of pulp and concentrate, e.g., for the B2 fraction, exceed the initial plant content, the relative reduction by truncation terminates this part of the total output without considering that another CNCPS fraction should be increased equivalently. This, so to speak, missing CP will reappear in the supernatant/loss fraction, since this is not measured but calculated as the difference between plant CP input and the summed pulp and concentrate CP output. Although we found the truncation as a necessary step in the analysis of this first-of-its-kind study, we also conclude that the unintended consequence is that the supernatant/loss fraction assumingly becomes inaccurate when calculated by difference. Future studies on this matter should include CNCPS analysis of the supernatant in order to improve determination of CP flow into the outputs. A tentative estimate of the significance of the second truncation (see the Materials and Methods section “Biorefinery Outputs”) was provided by omitting this, and hereby the supernatant recovery dropped to − 3 to 18% (mean = 13.1%; data not shown). The fact that the average for one of the species becomes a negative value reveals that there are some uncertainties related to the CNCPS method as compared to the Dumas method, assumingly due to the five independently quantified CP fractions instead of one (total N). However, the non-truncated CP recovery in the supernatant is much more in line with the values presented in Stødkilde et al. (2020). The presented data here indicate and raise an important question: whether or not some of the CP shifts from one CNCPS fraction to another during the biorefinery process. The reported surplus of B2 and B3 found in the mass balance reveals a pattern, which suggests that during the biorefinery process, the B2 fraction increases in the legumes whereas the B3 fraction increases in the grasses. The data does not point at which fractions are correspondingly decreased, which indicates that it could be a combination of more contributing fractions. This question needs further examination in order to achieve improved estimation of biorefinery CP output quality based on forage crop analyses.
Linking Forage Biomass CNCPS Analysis to Biorefinery Output
The above discussion underpins different aspects of the findings that influence the link between forage crop CP content and biorefinery output. Farmers will tend to maximize the B1 and B2 protein output per hectare and thereby choose the highest yielding forage crop, which might be unbeneficial in case this plant has a low recovery of B1 and B2 in the protein concentrate. Thus, the knowledge on CP recovery for a variety of crop species is important. The finding regarding that the protein concentrate contains other CP fractions than B1 and B2 (namely A, B3, and C) stresses the importance of qualifying the feed properties for these fractions to monogastrics. This will improve further optimization of forage crop choice for protein concentrate extraction. Finally, the possible shift between CNCPS fractions during the biorefinery process might change which crop is the most suitable for protein extraction, since the input could be different from the output. In addition, future studies should look more into cut dates and management, e.g., fertilization, and how this influences the distribution between CNCPS protein fractions.
In summary, the tentative assumption of extractable protein being equal to CNCPS fractions of B1 and B2 and partly B3 was shown to be too simplified. The CP recovery varies across forage crop species; however, only the B3 and total CP recovery showed systematic differences between legumes and grasses. The presented insight will potentially influence the balance between crop species in terms of expected maximized extractable protein yield, meaning that the forage species producing the largest crop CP yield per hectare may not be the overall most beneficial species. We point at the replication of this experiment to shed light on an even broader number of species, varieties, and cut dates—not only in the spring but also throughout the year - in order to secure further improvement for field experiment results to be related to realistic extractable protein yields.
Data Availability
Raw data can be accessed by contacting the corresponding author.
Code Availability
Not applicable
References
Boland, M. J., Rae, A. N., Vereijken, J. M., Meuwissen, M. P. M., Fischer, A. R. H., van Boekel, M. A. J. S., … Hendriks, W. H. (2013). The future supply of animal-derived protein for human consumption. Trends in Food Science and Technology. Elsevier. https://doi.org/10.1016/j.tifs.2012.07.002
Kristensen, L., Støier, S., Würtz, J., & Hinrichsen, L. (2014). Trends in meat science and technology: the future looks bright, but the journey will be long. Meat Science, 98(3), 322–329. https://doi.org/10.1016/j.meatsci.2014.06.023.
Buxton, D. R. (1996). Quality-related characteristics of forages as influenced by plant environment and agronomic factors. Animal Feed Science and Technology, 59(1–3 SPEC. ISS), 37–49. https://doi.org/10.1016/0377-8401(95)00885-3.
Stødkilde, L., Damborg, V. K., Jørgensen, H., Lærke, H. N., & Jensen, S. K. (2019). Digestibility of fractionated green biomass as protein source for monogastric animals. Animal, 13(9), 1817–1825. https://doi.org/10.1017/S1751731119000156.
Pugesgaard, S., Schelde, K., Larsen, S. U., Lærke, P. E., & Jørgensen, U. (2015). Comparing annual and perennial crops for bioenergy production - influence on nitrate leaching and energy balance. GCB Bioenergy, 7(5), 1136–1149. https://doi.org/10.1111/gcbb.12215.
Solati, Z., Jørgensen, U., Eriksen, J., & Søegaard, K. (2017). Dry matter yield, chemical composition and estimated extractable protein of legume and grass species during the spring growth. Journal of the Science of Food and Agriculture, 97(12), 3958–3966. https://doi.org/10.1002/jsfa.8258.
Kamm, B., Schönicke, P., & Hille, C. (2016). Green biorefinery - industrial implementation. Food Chemistry, 197(B), 1341–1345. https://doi.org/10.1016/j.foodchem.2015.11.088.
Damborg, V. K., Jensen, S. K., Weisbjerg, M. R., Adamsen, A. P., & Stødkilde, L. (2020). Screw-pressed fractions from green forages as animal feed: chemical composition and mass balances. Animal Feed Science and Technology, 261, 114401. https://doi.org/10.1016/j.anifeedsci.2020.114401
Santamaría-Fernández, M., Molinuevo-Salces, B., Lübeck, M., & Uellendahl, H. (2018). Biogas potential of green biomass after protein extraction in an organic biorefinery concept for feed, fuel and fertilizer production. Renewable Energy, 129, 769–775. https://doi.org/10.1016/j.renene.2017.03.012
Solati, Z., Manevski, K., Jørgensen, U., Labouriau, R., Shahbazi, S., & Lærke, P. E. (2018). Crude protein yield and theoretical extractable true protein of potential biorefinery feedstocks. Industrial Crops and Products, 115, 214–226. https://doi.org/10.1016/j.indcrop.2018.02.010
Stødkilde, L., Damborg, V. K., Jørgensen, H., Laerke, H. N., & Jensen, S. K. (2018). White clover fractions as protein source for monogastrics: dry matter digestibility and protein digestibility-corrected amino acid scores. Journal of the Science of Food and Agriculture, 98(7), 2557–2563. https://doi.org/10.1002/jsfa.8744.
Santamaria-Fernandez, M., Ambye-Jensen, M., Damborg, V. K., & Lübeck, M. (2019). Demonstration-scale protein recovery by lactic acid fermentation from grass clover – a single case of the production of protein concentrate and press cake silage for animal feeding trials. Biofuels, Bioproducts and Biorefining, 13(3), 502–513. https://doi.org/10.1002/bbb.1957.
Kleen, J., Taube, F., & Gierus, M. (2011). Agronomic performance and nutritive value of forage legumes in binary mixtures with perennial ryegrass under different defoliation systems. Journal of Agricultural Science, 149(1), 73–84. https://doi.org/10.1017/S0021859610000456.
Krawutschke, M., Kleen, J., Weiher, N., Loges, R., Taube, F., & Gierus, M. (2013). Changes in crude protein fractions of forage legumes during the spring growth and summer regrowth period. Journal of Agricultural Science, 151(1), 72–90. https://doi.org/10.1017/S002185961200024X.
Sleugh, B., Moore, K. J., George, J. R., & Brummer, E. C. (2000). Binary legume-grass mixtures improve forage yield, quality, and seasonal distribution. Agronomy Journal, 92(1), 24–29. https://doi.org/10.2134/agronj2000.92124x.
Licitra, G., Hernandez, T. M., & Van Soest, P. J. (1996). Standardization of procedures for nitrogen fractionation of ruminant feeds. Animal Feed Science and Technology, 57(4), 347–358. https://doi.org/10.1016/0377-8401(95)00837-3.
Sniffen, C. J., O’Connor, J. D., Van Soest, P. J., Fox, D. G., & Russell, J. B. (1992). A net carbohydrate and protein system for evaluating cattle diets: II. Carbohydrate and protein availability. Journal of Animal Science, 70(11), 3562–3577. https://doi.org/10.2527/1992.70113562x.
Tylutki, T. P., Fox, D. G., Durbal, V. M., Tedeschi, L. O., Russell, J. B., Van Amburgh, M. E., & Pell, A. N. (2008). Cornell Net Carbohydrate and Protein System: a model for precision feeding of dairy cattle. Animal Feed Science and Technology, 143(1–4), 174–202. https://doi.org/10.1016/j.anifeedsci.2007.05.010.
Stødkilde, L., Eriksen, J., & Jensen, S. K. (2020). Biorefined grasses and legumes – effect of species and spring cutting date on extractable protein yield. In Meeting the future demands for grassland production. Proceedings of the 28th General Meeting of the European Grassland Federation Helsinki, Finland. 19–22 October 2020. Helsinki, Finland
R Core Team. (2018). R: a language and environment for statistical computing. Vienna, Austria: R Foundation for Statistical Computing. Retrieved from https://www.r-project.org/
Pinheiro, J., Bates, D., DebRoy, S., Sarkar, D., & R Core Team. (2018). nlme: linear and nonlinear mixed effects models. R package version 3.1–137. Retrieved from https://cran.r-project.org/package=nlme
Hothorn, T., Bretz, F., & Westfall, P. (2008). Simultaneous inference in general parametric models. Biometrical Journal, 50(3), 346–363
Damborg, V. K., Stødkilde, L., Jensen, S. K., & Weisbjerg, M. R. (2018). Protein value and degradation characteristics of pulp fibre fractions from screw pressed grass, clover, and lucerne. Animal Feed Science and Technology, 244, 93–103. https://doi.org/10.1016/j.anifeedsci.2018.08.004
Santamaría-Fernández, M., Karkov Ytting, N., & Lübeck, M. (2019). Influence of the development stage of perennial forage crops for the recovery yields of extractable proteins using lactic acid fermentation. Journal of Cleaner Production, 218, 1055–1064. https://doi.org/10.1016/j.jclepro.2019.01.292
Acknowledgments
We gratefully acknowledge the help of lab technicians Margit Paulsen and Elsebeth Lyng Pedersen, Aarhus University Foulum.
Funding
The research was supported by the Biobase Green Protein project funded by Aarhus University and the Danish Ministry of Food, Agriculture, and Fisheries; and by the [Green Development and Demonstration Program] “Multiplant” project [grant number 34009-13-0678] and coordinated by the International Centre for Research in Organic Food Systems (ICROFS).
Author information
Authors and Affiliations
Contributions
Conceptualization: Lene Stødkilde, Søren Krogh Jensen, Jørgen Eriksen;
Methodology: Lene Stødkilde, Søren Krogh Jensen, Jørgen Eriksen;
Formal analysis and investigation: Lene Stødkilde, Henrik Thers;
Writing—original draft preparation: Henrik Thers;
Writing—review and editing: Henrik Thers, Lene Stødkilde, Søren Krogh Jensen, Jørgen Eriksen;
Funding acquisition: Lene Stødkilde, Søren Krogh Jensen, Jørgen Eriksen;
Resources: Lene Stødkilde, Søren Krogh Jensen, Jørgen Eriksen;
Supervision: Søren Krogh Jensen, Jørgen Eriksen
Corresponding author
Ethics declarations
Ethical Approval
Not applicable (No human participants or data were used in this study.)
Consent to Participate
Not applicable (No human participants or data were used in this study.)
Consent to Publish
Not applicable (No human participants or data were used in this study.)
Competing Interests
The authors declare that they have no competing interests.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Below is the link to the electronic supplementary material.
Rights and permissions
About this article

Cite this article
Thers, H., Stødkilde, L., Jensen, S.K. et al. Linking Protein Quality in Biorefinery Output to Forage Crop Crude Protein Input via the Cornell Net Carbohydrate and Protein System. Appl Biochem Biotechnol 193, 2471–2482 (2021). https://doi.org/10.1007/s12010-021-03496-x
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s12010-021-03496-x