
Abstract
Tuberculosis (TB) is an infectious disease caused by Mycobacterium tuberculosis (MTB) and considered as serious public health concern worldwide which kills approximately five thousand people every day. Therefore, TB drug development efforts are in gigantic need for identification of new potential chemical agents to eradicate TB from the society. The bacterial DNA gyrase B (GyrB) protein as an experimentally widely accepted effective drug target for the development of TB chemotherapeutics. In the present study, advanced pharmacoinformatics approaches were used to screen the Mcule database against the GyrB protein. Based on a number of chemometric parameters, five molecules were found to be crucial to inhibit the GyrB. A number of molecular binding interactions between the proposed inhibitors and important active site residues of GyrB were observed. The predicted drug-likeness properties of all molecules were indicated that compounds possess characteristics to be the drug-like candidates. The dynamic nature of each molecule was explored through the molecular dynamics (MD) simulation study. Various analyzing parameters from MD simulation trajectory have suggested rationality of the molecules to be potential GyrB inhibitor. Moreover, the binding free energy was calculated from the entire MD simulation trajectories highlighted greater binding free energy values for all newly identified compounds also substantiated the strong binding affection towards the GyrB in comparison to the novobiocin. Therefore, the proposed molecules might be considered as potential anti-TB chemical agents for future drug discovery purposes subjected to experimental validation.

Graphical Abstract
Similar content being viewed by others
Avoid common mistakes on your manuscript.
Introduction
The present treatment scenario is in the tremendous need of developing newer classes of anti-tubercular drugs in order to shorten the duration of therapy and as well as to combat the progression of mycobacterial resistance to existing antibacterial chemical entities [1, 2]. Drug-resistant tuberculosis (TB) is continuously rising worldwide and has become an important public health concern globally [3,4,5,6,7]. The causative agent of TB is Mycobacterium tuberculosis (MTB) which is an airborne pathogen transmitted through inhalation of contaminated droplets nuclei [8]. According to the World Health Organization (WHO) global TB report 2019, approximately 10 million peoples fell ill with TB infection in 2018, and among them, 484,000 people were found to be fallen ill with drug-resistant TB. Above all, the most important threatening fact was that about 1.5 million people died of TB in the year 2018. Since the introduction of four-drugs (isoniazid, rifampin, pyrazinamide, and either ethambutol or streptomycin) treatment regimens almost 40 years back [9, 10], now five decades have gone but drug development for TB treatment remained stagnant, except few new chemicals (such as bedaquiline, delamanid, and pretomanid) providing some hope by entering into the clinical trial phase III and can be a promising anti-tubercular drug in near future [10]. So, given the devastating fact of TB epidemics and the availability of the limited number of anti-tubercular drugs, it is now very obvious that extensive progressions are needed in the pipelines for the development of newer TB diagnostics methods, drugs, and vaccines at global or national levels.
DNA Gyrase is a bacterial enzyme and it is a member of the topoisomerase II families which plays a crucial role to stop the DNA supercoils formation during the viral DNA replication and transcription. The removal of supercoiling is done by the introduction of negative supercoils which permits the DNA to uphold appropriate topology throughout these functions. Due to its absence in the eukaryotic cells, the DNA Gyrase considered being an effective and crucial antibacterial drug target. It is already proved that DNA Gyrase enzyme inhibitors active against non-replicating Mycobacteria that might be crucial to remove the persistent organisms [11]. The DNA Gyrase is found in two subunits, DNA Gyrase A (GyrA) and DNA Gyrase B (GyrB) in heterodimer (A2B2) form for the fully functional enzyme. The GyrA involves breaking and reuniting of the DNA and GyrB controls the ATPase activity [12]. GyrA is a widely used and well-known antibacterial target and nowadays not considered as a novel drug target [13]. This subunit has been used for a long time as a target for the quinolone class of drugs and unfortunately develops mutations that lead to the development of MTB resistance [14]. Comparatively less known GyrB becomes an effective and novel drug target for bacterial infection [15,16,17,18,19,20]. To date, only a single GyrB inhibitor, novobiocin was approved by the FDA [21] which belongs to the aminocoumarins class of molecules and shows strong inhibition activity against the GyrB [16, 22]. Later, the novobiocin was withdrawn from the market due to adverse side effects and poor pharmacological behavior [16, 23]. Scientific communities across the world are trying to identify novel GyrB inhibitors [24,25,26,27,28,29,30,31,32] to combat the drug resistance of TB but no complete success made so far. In this scenario, there is demand of extreme efforts and investigation to identify therapeutically potential active GyrB inhibitors.
Herein, the present study was focused on computational identification and optimization of chemotherapeutic entities for TB by specifically targeting GyrB domain. Therefore, a multi steps virtual screening protocol was employed against Mcule database. In the virtual screening protocol, three-steps molecular docking study, such as high throughput virtual screening (HTVS), standard precision (SP), and extra precision (XP) docking was performed followed by binding energy calculation using Molecular Mechanics-Generalized Born Surface Area (MM-GBSA) [33] approach. Furthermore, molecular dynamic simulations were carried out to evaluate the interaction stability of the obtained molecules in complex with GyrB domain. Finally, the Molecular Mechanics Poisson-Boltzmann Surface Area (MM-PBSA) [34] approach was applied for determining binding free energies upon selective GyrB inhibitors.
Materials and Methods
Preparation of Chemical Library Dataset and DNA GyraseB Protein
A large chemical library, “Mcule Purchasable” (Building block) database (https://mcule.com/database/) consisting of more than two million compounds was downloaded in December 2019 for virtual screening purposes. Mcule database is comprised of high-quality compounds ready-to-use for drug discovery purposes. Diverse structural featured molecules are in the above database which contains crucial and effective functional groups that make the database suitably use for hit-to-lead identification and optimization. Therefore, the database can be readily used in fragment-based drug design (FBDD) and structure-based drug design (SBDD) approaches. In the current study, the database was used for the SBDD-based virtual screening against the DNA GyrB. The downloaded compounds (in .sdf format) were prepared using the LigPrep module of Maestro. In addition, redundant molecules were removed and the repaired the incorrect valency and finally saved in 3D coordinates. The novobiocin, a well known anti-tubercular drug targeting the DNA GyrB was considered for the comparison and assessment purpose of the study. The novobiocin was also prepared using the same parameters as mentioned above.
The crystal structure of the GyrB was obtained from the Research Collaboratory for Structural Bioinformatics-Protein Data Bank (RCSB-PDB) having PDB ID: 4B6C [35]. The resolution and R value of the selected protein were found to be 2.3 Å and 0.287, respectively. The receptor was consisting of two chains in the form of dimer and each chain having 234 amino acids. To speed up and simplify the calculation, only chain “A” was considered in the study. The “Protein Preparation Wizard” utility tool embedded in the Maestro user interface [36] was used to prepare the protein. Initially, the protein was preprocessed, in which the correct bond order assignment and hydrogen atoms were added. Missing side chains and loops were repaired using the Prime module [37]. The protein was minimized by the optimized potentials for liquid simulations_2005 (OPLS_2005) force field [38]. Thereafter, the prepared protein crystal structure was considered for the grid preparation for the molecular docking study using the “Receptor Grid Generation” panel. The positional information of bound co-crystal ligand was used to define the coordinate of the grid. The grid size of the box was considered such a way that the bound co-crystal ligand confined properly.
Virtual Screening Through Multi-Step Docking and MM-GBSA Estimation
The “Virtual Screening Workflow” (VSW) module of the Schrödinger-Meastro interface was used for virtually screened the large Mcule database against the GyrB target protein. Particularly, the VSW module is an extensive and rigorous protocol that sequentially executes three different docking programs viz. Glide-HTVS [39], SP [39], and XP [40] followed by the MM-GBSA [41] based binding free energy calculation. The workflow was designed in such a way that allows users to customize the parameter to choose in each stage, and results retained in one stage passed on to the next stage for further execution. The systematic search of conformational and orientation position of each ligand was explored through multi-steps molecular docking analyses as mentioned above. The prepared grid file was browsed under the “Receptors” tab in VSW for docking. Prepared entire compounds dataset was given as ligand input. In three stages of docking protocol employed in VSW, best 10% docked molecules in each stage were considered as input for the next step. In details, first of all, the entire set of molecules were docked in HTVS approach, and best 10% successful molecules were considered as input of SP module. On successful completion of SP docking, the best 10% molecules were kept for the input of XP docking. Finally, molecules from the above step were docked through the XP-module, and XP-descriptor information was saved for further binding interaction analyses. Followed by the above step, the binding free energy of the best 10% XP-docked molecules were calculated through Prime-MM-GBSA approach and further compared with same of novobiocin. Compounds having better XP-Gscore and binding energy in comparison to the novobiocin were further considered for in-silico pharmacokinetics analyses.
In-silico ADME and Drug-Likeness Prediction
For being an effective drug molecule, the essential chemical substance should reach at the active site with sufficient concentration and without any structural alteration. The pharmacokinetics parameters included absorption, distribution, metabolism, and excretion (ADME) can explain the biophysical/biochemical states of the molecule during traveling starts from intake to reach the target site. For this purpose, a number of physicochemical and drug-likeness properties of molecules from VSW were obtained through the SwissADME web server [42], available at http://www.swissadme.ch/. Pharmacokinetics properties included n-octanol and water (log Po/w) partition coefficient or lipophilicity, molar solubility in water, blood-brain barrier (BBB) permeability, skin permeation, human gastrointestinal absorption (HIA) capability etc., were explored in details.
Molecular Dynamics Simulation and Binding Energy Through MM-PBSA Approach
It is essential and critical to understand the behavior of any molecule in dynamic states obtained from the computational screening approach. For this purpose, the molecular dynamics simulation is a widely used and crucial tool to explore the dynamic nature of the molecule inside the protein cavity. Final proposed DNA GyrB molecules were considered for a 100 ns all-atoms MD simulation study. The MD simulation was carried out in the Gromacs 2018.2 software tool (http://www.gromacs.org/) available at the Lengau CHPC server, Cape Town, South Africa. In MD simulation, the time step was kept 2 fs with constant pressure and temperature of 1 atm and 300 K, respectively. The topology of the protein was generated using the all-atoms CHARMM36 forcefield. The online tool, SwissParam [42] was used to generate the ligand topology. To conduct the simulation, a system of the cubic-shaped box was generated surrounding the protein-ligand complex having a diameter of 10 Å from the center of the system. The TIP3P water model was used to solvate the system prior to simulation. A sufficient number of positive (Na+) or negative (Cl−) ions was used to neutralize each system. Each system was minimized to relieve any high energy strains in the system. Further, the system was equilibrated of 10,000 steps through the steepest descent algorithm. The long-range interactions such as van der Waals and electrostatic cut-off were used to 0.9 and 1.4 nm respectively. The trajectory information from MD simulation was saved in 1 ps intervals. A number of crucial parameters such as root-mean-square deviation (RMSD), root-mean-square fluctuation (RMSF), and radius of gyration (Rg) were calculated from the trajectory to explore the dynamic stability of the protein-ligand complex. The binding affection of each ligand towards the DNA GyrB was assessed by calculation of the binding free energy through the MM-PBSA approach with the help of using g_mmpbsa utility tool [43]. The procedure of the MM-PBSA approach can be found in one of the previous publications by our research group [44].
Results and Discussion
Virtual Screening Analysis of Large Chemical Library Database
Virtual screening (VS) is an important and widely used computational drug discovery platform for screening against a specific target. With the help of advanced computational approaches, a large dataset of small molecules can be searched to find out effective therapeutic hit molecules. VS become extremely popular to the drug discovery community for reaching towards a higher number of active hits with fewer efforts and cost as well as no animal sacrifice. The structure-based virtual screening (SBVS) is one of the most popular screening methods in which one or more molecular docking studies can be performed to filter out the promising molecules from a large molecular dataset. In the current study, three levels of molecular docking study were performed through the “VSW” available in the Maestro Suite [36]. The workflow of the present study is given in Fig. 1. Prior to VSW, the molecular docking protocol was validated by the self-docking approach [45]. In this method, the bound co-crystal ligand was re-drawn and docked in the same receptor site. The best-docked pose was superimposed on the original co-crystal bound ligand and their RMSD calculated based on the atomic alignment. It is reported that RMSD value less than 2.0 Å, between re-docked best pose and co-crystal pose, successfully validated the employed molecular docking protocol [45]. In the current study, the RMSD was found to be 1.12 Å which clearly validated the used molecular docking protocol.

Workflow of the screening of promising DNA GyraseB inhibitors
Particularly, a large set of 2,018,271 diverse molecules under the purchasable building block were downloaded from the Mcule database. The entire set was considered for three levels of molecular docking one by one in order to execute HTVS, SP, and XP-docking. In each step, best 10% molecules were considered. Finally, about two thousand best molecules remained, and those were further considered for binding free energy calculation through the MM-GBSA approach. In similar fashion, the XP-Glide score and MM-GBSA-based binding free energy of novobiocin-GyrB complex were generated and found to be − 9.21 and − 37.582 Kcal/mol, respectively. The above two values were considered as the threshold for further reduction of chemical space. Molecules having XP-Glide score and binding free energy values higher than novobiocin were removed for further analysis. By asserting the above screening criteria, a total of 356 molecules were retained and considered for further analyses. A number of pharmacokinetics and drug-likeness properties including Lipinski’s rule of five (LoF) and Veber’s rule were assessed. A total of 28 molecules were found to have acceptable pharmacokinetics profile and drug-likeness characteristics. Finally, based on binding interaction analyses and synthetic accessibility values, five molecules (T1–T5) were identified as promising GyrB inhibitors. The two-dimensional schematic representation of the finally considered molecules is given in Fig. 2.

Two-dimensional representation of proposed DNA GyraseB inhibitors
Molecular Binding Interactions Analysis
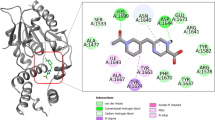
For evaluation of the molecular docking poses of protein-ligand complexes, the lowest XP-GlideScore (Gscore) followed by the lowest MM-GBSA score (i.e., more negative values) were checked, selected, and investigated. Initially for the standard inhibitor of GyrB, the best docking pose and energy score of novobiocin were retrieved and considered to investigate the docking interaction profile. The best docking poses of all identified compounds (T1–T5) including the novobiocin obtained through Glide XP-docking were critically checked and displayed in Fig. 3. The Glide XP docking score of identified compounds T1, T2, T3, T4, T5, and novobiocin was found to be − 10.45, − 10.21, − 9.97, − 10.81, − 9.46, and − 9.21 Kcal/mol, respectively. On the other hand, the MM-GBSA value of the docked complexes (i.e., compounds T1, T2, T3, T4, T5, and novobiocin with GyrB) was revealed as − 69.26, − 65.17, − 62.43, − 68.62, − 61.49, and − 37.58 Kcal/mol, respectively. In order to infer the biological or molecular interaction significance of identified ligands, those bind with active site residues of GyrB, different types of molecular binding interaction patterns were explored. The T1 was observed to interact with three residues (Glu56, Asp79, and Gln102) through hydrogen bonding (H-bond), mostly with their side-chain atoms. In addition, T1 was also formed hydrophobic contacts with residues Asn52, Ile84, Val99, and Val123. Moreover, T1 also links the carbonyl-group of Glu56 via a water molecule. From the surface view presentation (Fig. S1 in supplementary data), it can also be observed that T1 deeply buried inside the active site cavity of GyrB. It can be seen that (Fig. 3) T2 was formed hydrogen bond interaction with Asp79 and hydrophobic interactions with Pro85 and Val99. Exploration of the molecular binding interaction between T5 and GyrB was revealed the involvement of residue Asp79 in H-bond interaction (distance measured as 2.11 Å), whereas several other residues such as Asn52, Arg82, Ile84, Pro85, Gln102, and Val123 observed to participate in hydrophobic interactions with T5. Two residues, Glu56 and Arg141 of GyrB were also formed water bridge interactions with T5. The close-up view (Fig. S1 in Supplementary file) also highlighted the absolute deep buried-ness of T5 at the cavity of the GyrB protein. Interestingly, the water-mediated interaction with residue Arg141 was also reported by Shirude et al. through binding interaction with some aminopyrazinamides [11]. Likewise T1, two residues (Glu56 and Asp79) were observed to interact with the T4 through H-bond interactions. The residue, Glu56, was not only formed H-bond interaction but also established a water bridge connection with T4. The docking interaction has also revealed some important residues (Asn52, Ile84, Pro85, Val99, and Val123) participation in the formation of hydrophobic contacts with T4. As T4, almost similar binding interaction patterns were obtained for T3 except for an additional residue (Gln102) found to form hydrophobic interaction with T3 (Fig. 3). For T3 and T4, such alike or similar binding patterns were probably obtained due to similar structural conformer they hold. The 3D surface view binding mode of each molecule was checked and given in Fig. S1 (Supplementary file). It can be seen that all identified molecules perfectly enclosed within a deeply buried receptor cavity of GyrB. Fig. S1 (Supplementary file) indicates that all molecules represent a relatively better surface area than the novobiocin to hold their structural or interaction integrity inside the active site GyrB pocket.

Binding interaction profile of DNA GyraseB inhibitors
Comparative Analysis of Molecular Interaction Profiles
A number of research articles were explored to analyze binding interactions comparison with the current outcomes. Maharaj and Soliman [26] performed pharmacophore-based virtual screening and proposed two important GyrB inhibitors. Their study revealed that the molecules interacted with residues Asn52, Asp79, Gly83, and Ser169. A similar interaction pattern was also found for all the proposed molecules such as Asn52 and Asp79 potentially interacting with T1, T2, T3, T4, and T5. In another study, Kamsi et al. [24], performed a 3D-quantitative structure-activity relationship (QSAR) and MD simulation study for exploring the structural requirement of GyrB inhibitor compounds. They have reported that Asp79, Arg82, and Arg141 were found to be critical amino acid residues for binding interactions. Observation in the current study also substantiated by H-bond interaction with Asp79 to all molecules. Moreover, Arg82 was found to interact with T5. Wang et al. [46] performed 3D-QSAR, molecular docking, MD simulations to explore potential GyrB inhibitors. In the molecular docking study, they have found Asp79 crucial amino residues in binding interactions. As mentioned earlier that all proposed molecules (T1–T5) were found to form a strong interaction profile with residue Asp79. In one of our previous published works [19], pharmacophore-based virtual screening was carried out to find potent GyrB inhibitors. Three promising molecules were proposed with critical interacting amino acids like Asn52, Glu56, Arg82, Pro85, His89, Asp97, Val99, and Gln102. In the present molecular docking study of the proposed molecules was found to interact with all the above amino residues. The above observations undoubtedly adjudged and validate the potentiality of the proposed GyrB inhibitors.
Pharmacokinetics and Drug-Likeness Assessment
The pharmacokinetics characteristics of the proposed molecules (Fig. 2) were explored through, SwissADME (http://www.swissadme.ch/) and given in Table 1. Not a single molecule was found to violate the LoF and Veber’s rule. The molecular weight was found to be in the range of 310 to 360 g/mol. The molar refractivity of all molecules was revealed below 100. The topological polar surface area (TPSA) is an important parameter that describes the oral activeness of the molecules. It is illustrated that molecules having TPSA less than 130Å2 are orally active. The TPSA was found to be 103.04, 116.67, 73.58, 92.04, and 93.81 Å2 for T1, T2, T3, T4, and T5, respectively. Hence, the above data was clearly suggested that all molecules orally active in nature. It was also found that all molecules soluble in nature and highly absorbable in the gastrointestinal (GI) tract. The complexity of synthesis can be explored by the synthetic accessibility (SA) parameter. The value of SA from 1 to 10 describes the easiest to difficult for chemical synthesis. From Table 1, it can be seen that SA of all molecules exerted as less than 4.32 which also indicates that proposed inhibitors are not difficult to synthesize in the laboratory.
The bioavailability was explored through the radar plot and given in Fig. S1 (Supplementary file). The area under the pink color expressed the different features included unsaturation (INSATU), insolubility (INSOLU), hydrophobicity (LIPO), rotatable bonds (FLEXI), molecular weight (SIZE), and polar surface area (POLAR). The recommended range of LIPOPHILICITY, i.e., XLOGP3 is between − 0.7 and + 5.0, SIZE, i.e., MW should be < 500 g/mol, POLARITY: TPSA should lie between 20 and 130 Å2, INSOLU should be in the range of 0 to 6, INSATU should be within 1, and FLEX should be less than 9 rotatable bonds. Table 1 explained that compounds T1, T2, T3, T4, and T5 possess adequate drug-likeness properties for showing GyrB inhibitory action biologically.
In order to explore two important characteristics such as human intestinal absorption (HIA) and BBB, the boiled-egg figure were derived and it is given in Fig. S2 (Supplementary file). In Fig. S2, BBB penetration is designated by the yolk areas (yellow region) and the white part (albumin) signifies the HIA absorption. Both areas are mutually exclusive. The substrate (PGP+) and non-substrate (PGP−) permeability glycoprotein (PGP) also can be analyzed from the boiled-egg model. The substrate characteristic is denoted by the blue dots, and it predicts the effluation from the CNS by the P-glycoprotein. The ‘PGP−’ can be identified by the red dots and it signifies not to be effluated from the CNS by P−glycoprotein. From Fig. S2, it can be seen that T1, T2, and T3 were belonged to ‘PGP−’ whereas, T4 and T5 under ‘PGP+’ category. From the above analysis of the several physicochemical and pharmacokinetics properties, it is certainly justified that all the GyrB inhibitors are potential enough to show drug-like characteristics.
Molecular Dynamics Simulation
The dynamic behavior of the proposed molecules inside the GyrB was explored through all-atoms 100 ns MD simulation study. To analyze the dynamic nature and stability of the ligand-bound with GyrB, a number of parameters including RMSD, the radius of gyration (Rg), and RMSF were calculated from the entire MD-simulated trajectories.
The RMSD parameter from the MD trajectory can explain the deviation of the protein backbone bound with small molecules. Average, maximum, and minimum RMSD values were recorded and these are given in Table 2. The RMSD of each frame was plotted and it is given in Fig. 4. The mean RMSD of GyrB was found to be 0.217, 0.203, 0.144, 0.166, 0.192, and 0.206 nm bound with compounds T1, T2, T3, T4, T5, and novobiocin, respectively. Fig. 4 clearly explained that all the systems were consistent in maintaining the molecular interaction profile throughout the simulation run and most importantly not shown fluctuation beyond 0.301 nm which is an absolutely acceptable range to judge the protein-ligand complex interaction stability. However, the backbone atoms of GyrB was found to fluctuate a little bit higher when bound with compound T5, as compared to others. From Table 2 and Fig. 4, it was clear that the backbone of GyrB bound with proposed molecules remained stable during the simulation.

RMSD vs. time of MD simulation for the identified inhibitors (T1–T5 and novobiocin) complex with GyrB
Individual amino acid residues are extremely crucial to explore the relative stability of the protein-ligand complex in the MD simulation study. The fluctuation of each and every amino acid residue was explored through the RMSF and it is given in Fig. 5. The average, maximum, and minimum RMSF were recorded and values are given in Table 2. The difference between maximum and average, and, average and minimum can give an idea about the fluctuation of amino acid residues in the MD simulation. The difference between maximum and average, and, average and minimum were found to be 0.611 and 0.092 nm; 0.901 and 0.081 nm; 0.288 and 0.064 nm; 0.315 and 0.081 nm; 0.513 and 0.075 nm; and 0.434 and 0.083 nm in case of GyrB bound with T1, T2, T3, T4, T5, and novobiocin, respectively. The observed low values undoubtedly explained that there was no significant fluctuation in any amino acid residues during the simulation. However, it can be noted that amino acid residues between 82 and 100 were found higher fluctuation due to lack of substantial number bonding and non-bonding interactions with ligand and other parts of the protein.

RMSF of individual amino acid residues of GyrB complex with identified inhibitors and novobiocin
The rigidity of protein-ligand complexes was explored through the Rg parameter. The Rg value of each frame was plotted against the time of the simulation and it is given in Fig. 6. Maximum, minimum, and average Rg values were also calculated and these are given in Table 2. Average Rg value was found to be 1.642, 1.645, 1.633, 1.634, 1.633, and 1.642 nm for GyrB bound with T1, T2, T3, T4, T5, and novobiocin, respectively. It can be noted that the highest and lowest Rg value was found to be 1.686 and 1.595 nm in the case of T2 and T5, respectively. Hence, the low difference between the maximum and minimum Rg value (0.082 nm) clearly explained that all systems were remained rigid during dynamic conditions and hence stability not disturbed for the proposed molecules when interacted with GyrB. The above observation of RMSD, RMSF, and Rg parameters obtained from the MD simulation trajectory was clearly indicated that protein-ligand complexes remained stable in dynamic states, and proposed GyrB inhibitors retained inside the receptor cavity.

The radius of gyration (Rg) of DNA GyrB complex with proposed GyrB inhibitors
Binding Energy Through MM-PBSA Approach
The binding affinity of the proposed inhibitors and novobiocin towards the GyrB was explored through binding free energy (ΔGbind) analyses obtained using through MM-PBSA approach from the entire MD simulation trajectory. The binding free energy calculated through the MM-PBSA approach is thought to be more accurate in comparison to the binding energy obtained in the molecular docking study. Binding free energy of each frame was calculated and it is plotted against time and given in Fig. 7. Maximum, minimum, and average binding free energy were also recorded and these are provided in Table 3. It can be seen that binding free energy of all GyrB inhibitors except T2 was found much higher in comparison to the novobiocin which undoubtedly suggested the potentiality of the molecules.

Binding free energy of DNA GyrB inhibitors
Average binding free energy was found to be − 1181.110, − 198.482, − 1237.240, − 1268.580, − 1240.650, and − 243.769 kJ/mol for T1, T2, T3, T4, T5, and novobiocin, respectively. From Fig. 7, it can be observed that ΔGbind of all frames remained consistent throughout the simulation. Furthermore, it was also noted that the Coulomb or electrostatic interaction (ΔGCoulomb) and van der Waals interaction energy (ΔGvdW) contributed majorly to the ΔGbind values. Above all, such strong binding free energy profiles of the proposed inhibitors undoubtedly indicated the potentiality of the molecules being promising DNA GyrB inhibitors.
Future Prospects
The application of computational power and algorithm in drug discovery research became a pivotal approach. MD simulation, molecular docking, and binding free energy calculation through MM-PBSA are widely used to identify potential compounds for a specific target. Irrespective of the extremely high use of computational drug discovery, there is a need to check the potentiality of the final GyrB inhibitors through a number of experimental validation approaches. The thermal shift assay is one of the important approaches can be used to explore further affinity. The kinetic study can be sued to assess the binding and unbinding mechanism of the molecules. The therapeutic efficacy of the proposed molecules can be improved based on the experimental assessments.
Conclusions
The current work was aimed to find out important potential DNA GyrB inhibitors for therapeutic application in TB. To achieve the aim, a set of more than two million compounds from the Mcule database was considered for the structure-based virtual screening. The crystal structure of GyrB was used as a receptor in the virtual screening workflow. About three hundred and fifty best molecules were obtained through a multiple molecular docking workflow such as HTVS, SP, and XP followed by a binding energy assessment through the MM-GBSA approach. In-silico pharmacokinetics assessment followed by binding interaction analyses were carried out. Finally, five best molecules were proposed as potential DNA GyrB inhibitors. Molecular docking explained that all proposed molecules were formed a number of strong binding interactions with the catalytic amino residues of GyrB. The drug-likeness was also assessed and found that all molecules possessed drug-like characteristics. The stability of the molecules inside the receptor cavity in dynamic states was checked through a 100 ns time span of MD simulation study. RMSD, RMSF, and Rg parameters from the MD simulation trajectory were clearly explained that proposed GyrB inhibitors efficient enough to retain inside the receptor in dynamic states. The high binding free energy of all proposed molecules in comparison to the novobiocin was substantiated by the potentiality of the proposed molecules. Therefore, GyrB inhibitors found through a structure-based screening approach might be important molecules for the therapeutic application of TB infection.
Availability of Data and Material
Not applicable.
References
Hoagland, D. T., Liu, J., Lee, R. B., & Lee, R. E. (2016). New agents for the treatment of drug-resistant mycobacterium tuberculosis. Advanced Drug Delivery Reviews, 102, 55–72.
Sotgiu, G., Centis, R., D'Ambrosio, L., & Migliori, G. B. (2015). Tuberculosis treatment and drug regimens. Cold Spring Harbor Perspectives in Medicine, 5(5), a017822.
Pourakbari, B., Mamishi, S., Mohammadzadeh, M., & Mahmoudi, S. (2016). First-line anti-tubercular drug resistance of mycobacterium tuberculosis in IRAN: a systematic review. Frontiers in Microbiology, 7, 1139.
Wu, X., Yang, J., Tan, G., Liu, H., Liu, Y., Guo, Y., Gao, R., Wan, B., & Yu, F. (2019). Drug resistance characteristics of mycobacterium tuberculosis isolates from patients with tuberculosis to 12 antituberculous drugs in China. Frontiers in Cellular and Infection Microbiology, 9, 1.
Dookie, N., Rambaran, S., Padayatchi, N., Mahomed, S., & Naidoo, K. (2018). Evolution of drug resistance in mycobacterium tuberculosis: a review on the molecular determinants of resistance and implications for personalized care. Journal of Antimicrobial Chemotherapy, 73(5), 1138–1151.
Ghajavand, H., Kargarpour Kamakoli, M., Khanipour, S., Pourazar Dizaji, S., Masoumi, M., Rahimi Jamnani, F., Fateh, A., Yaseri, M., Siadat, S. D., & Vaziri, F. (2019). Scrutinizing the drug resistance mechanism of multi- and extensively-drug resistant mycobacterium tuberculosis: mutations versus efflux pumps. Antimicrobial Resistance & Infection Control, 8(1), 70.
Munir, A., Kumar, N., Ramalingam, S. B., Tamilzhalagan, S., Shanmugam, S. K., Palaniappan, A. N., Nair, D., Priyadarshini, P., Natarajan, M., Tripathy, S., Ranganathan, U. D., Peacock, S. J., Parkhill, J., Blundell, T. L., & Malhotra, S. (2019). Identification and characterization of genetic determinants of isoniazid and rifampicin resistance in mycobacterium tuberculosis in Southern India. Scientific Reports, 9(1), 10283.
Marimani, M., Ahmad, A., & Duse, A. (2018). The role of epigenetics, bacterial and host factors in progression of mycobacterium tuberculosis infection. Tuberculosis (Edinburgh, Scotland), 113, 200–214.
Jangam, C. S., Bhowmick, S., Chorge, R. D., Bharatrao, L. D., Patil, P. C., Chikhale, R. V., AlFaris, N. A., Altamimi, J. z., Wabaidur, S. M., & Islam, M. A. (2019). Pharmacoinformatics-based identification of anti-bacterial catalase-peroxidase enzyme inhibitors. Computational Biology and Chemistry, 83, 107136.
Bahuguna, A., & Rawat, D. S. (2020). An overview of new antitubercular drugs, drug candidates, and their targets. Medicinal Research Reviews, 40(1), 263–292.
Shirude, P. S., Madhavapeddi, P., Tucker, J. A., Murugan, K., Patil, V., Basavarajappa, H., Raichurkar, A. V., Humnabadkar, V., Hussein, S., Sharma, S., Ramya, V. K., Narayan, C. B., Balganesh, T. S., & Sambandamurthy, V. K. (2013). Aminopyrazinamides: novel and specific GyrB inhibitors that kill replicating and nonreplicating mycobacterium tuberculosis. ACS Chemical Biology, 8(3), 519–523.
Champoux, J. J. (2001). DNA topoisomerases: structure, function, and mechanism. Annual Review of Biochemistry, 70(1), 369–413.
Mdluli, K., & Ma, Z. (2007). Mycobacterium tuberculosis DNA gyrase as a target for drug discovery. Infectious Disorders Drug Targets, 7(2), 159–168.
van Doorn, H. R., An, D. D., de Jong, M. D., Lan, N. T., Hoa, D. V., Quy, H. T., Chau, N. V., Duy, P. M., Tho, D. Q., Chinh, N. T., Farrar, J. J., & Caws, M. (2008). Fluoroquinolone resistance detection in mycobacterium tuberculosis with locked nucleic acid probe real-time PCR. The International Journal of Tuberculosis and Lung Disease, 12(7), 736–742.
Aboul-Fadl, T., Abdel-Aziz, H. A., Abdel-Hamid, M. K., Elsaman, T., Thanassi, J., & Pucci, M. J. (2011). Schiff bases of indoline-2,3-dione: potential novel inhibitors of mycobacterium tuberculosis (Mtb) DNA gyrase. Molecules, 16(9), 7864–7879.
Brvar, M., Perdih, A., Hodnik, V., Renko, M., Anderluh, G., Jerala, R., & Solmajer, T. (2012). In silico discovery and biophysical evaluation of novel 5-(2-hydroxybenzylidene) rhodanine inhibitors of DNA gyrase B. Bioorganic & Medicinal Chemistry, 20(8), 2572–2580.
Brvar, M., Perdih, A., Oblak, M., Masic, L. P., & Solmajer, T. (2010). In silico discovery of 2-amino-4-(2,4-dihydroxyphenyl)thiazoles as novel inhibitors of DNA gyrase B. Bioorganic & Medicinal Chemistry Letters, 20(3), 958–962.
Azam, M. A., & Thathan, J. (2017). Pharmacophore generation, atom-based 3D-QSAR and molecular dynamics simulation analyses of pyridine-3-carboxamide-6-yl-urea analogues as potential gyrase B inhibitors. SAR and QSAR in Environmental Research, 28(4), 275–296.
Islam, M. A., & Pillay, T. S. (2017). Identification of promising DNA GyrB inhibitors for tuberculosis using pharmacophore-based virtual screening, molecular docking and molecular dynamics studies. Chemical Biology & Drug Design, 90(2), 282–296.
Islam, M. A., & Pillay, T. S. (2019). Identification of promising anti-DNA gyrase antibacterial compounds using de novo design, molecular docking and molecular dynamics studies. Journal of Biomolecular Structure & Dynamics, 1–12. https://doi.org/10.1080/07391102.2019.1617785.
Jeankumar, V. U., Renuka, J., Santosh, P., Soni, V., Sridevi, J. P., Suryadevara, P., Yogeeswari, P., & Sriram, D. (2013). Thiazole-aminopiperidine hybrid analogues: design and synthesis of novel mycobacterium tuberculosis GyrB inhibitors. European Journal of Medicinal Chemistry, 70, 143–153.
Kumar Muthyala, M. K., Jamullamudi, R. N., Sangeeta, G. P. V., & Kurre, P. N. (2018). Identification of N-Benzylated Indole Mannich bases as potential anti TB agents by using computational studies and molecular hybridization technique. Current Computer-Aided Drug Design, 14(3), 200–206.
Chopra, S., Matsuyama, K., Tran, T., Malerich, J. P., Wan, B., Franzblau, S. G., Lun, S., Guo, H., Maiga, M. C., Bishai, W. R., & Madrid, P. B. (2012). Evaluation of gyrase B as a drug target in mycobacterium tuberculosis. The Journal of Antimicrobial Chemotherapy, 67(2), 415–421.
Kamsri, P., Punkvang, A., Hannongbua, S., Suttisintong, K., Kittakoop, P., Spencer, J., Mulholland, A. J., & Pungpo, P. (2019). In silico study directed towards identification of the key structural features of GyrB inhibitors targeting MTB DNA gyrase: HQSAR, CoMSIA and molecular dynamics simulations. SAR and QSAR in Environmental Research, 30(11), 775–800.
Tari, L. W., Trzoss, M., Bensen, D. C., Li, X., Chen, Z., Lam, T., Zhang, J., Creighton, C. J., Cunningham, M. L., Kwan, B., Stidham, M., Shaw, K. J., Lightstone, F. C., Wong, S. E., Nguyen, T. B., Nix, J., & Finn, J. (2013). Pyrrolopyrimidine inhibitors of DNA gyrase B (GyrB) and topoisomerase IV (ParE). Part I: structure guided discovery and optimization of dual targeting agents with potent, broad-spectrum enzymatic activity. Bioorganic & Medicinal Chemistry Letters, 23(5), 1529–1536.
Maharaj, Y., & Soliman, M. E. (2013). Identification of novel gyrase B inhibitors as potential anti-TB drugs: homology modelling, hybrid virtual screening and molecular dynamics simulations. Chemical Biology & Drug Design, 82(2), 205–215.
Saxena, S., Renuka, J., Yogeeswari, P., & Sriram, D. (2014). Discovery of novel mycobacterial DNA Gyrase B inhibitors: in silico and in vitro biological evaluation. Molecular Informatics, 33(9), 597–609.
Kale, M. G., Raichurkar, A., Hameed, P. S., Waterson, D., McKinney, D., Manjunatha, M. R., Kranthi, U., Koushik, K., Jena, L., Shinde, V., Rudrapatna, S., Barde, S., Humnabadkar, V., Madhavapeddi, P., Basavarajappa, H., Ghosh, A., Ramya, V. K., Guptha, S., Sharma, S., Vachaspati, P., Kumar, K. N., Giridhar, J., Reddy, J., Panduga, V., Ganguly, S., Ahuja, V., Gaonkar, S., Kumar, C. N., Ogg, D., Tucker, J. A., Boriack-Sjodin, P. A., de Sousa, S. M., Sambandamurthy, V. K., & Ghorpade, S. R. (2013). Thiazolopyridine ureas as novel antitubercular agents acting through inhibition of DNA Gyrase B. Journal of Medicinal Chemistry, 56(21), 8834–8848.
Barancokova, M., Kikelj, D., & Ilas, J. (2018). Recent progress in the discovery and development of DNA gyrase B inhibitors. Future Medicinal Chemistry, 10(10), 1207–1227.
Durcik, M., Tammela, P., Barancokova, M., Tomasic, T., Ilas, J., Kikelj, D., & Zidar, N. (2018). Synthesis and evaluation of N-Phenylpyrrolamides as DNA Gyrase B inhibitors. ChemMedChem, 13(2), 186–198.
Zidar, N., Macut, H., Tomasic, T., Peterlin Masic, L., Ilas, J., Zega, A., Tammela, P., & Kikelj, D. (2019). New N-phenyl-4,5-dibromopyrrolamides as DNA gyrase B inhibitors. Medchemcomm, 10(6), 1007–1017.
Durcik, M., Lovison, D., Skok, Z., Durante Cruz, C., Tammela, P., Tomasic, T., Benedetto Tiz, D., Draskovits, G., Nyerges, A., Pal, C., Ilas, J., Peterlin Masic, L., Kikelj, D., & Zidar, N. (2018). New N-phenylpyrrolamide DNA gyrase B inhibitors: optimization of efficacy and antibacterial activity. European Journal of Medicinal Chemistry, 154, 117–132.
Zhang, X., Perez-Sanchez, H., & Lightstone, F. C. (2017). A comprehensive docking and MM/GBSA rescoring study of ligand recognition upon binding antithrombin. Current Topics in Medicinal Chemistry, 17(14), 1631–1639.
Wang, C., Greene, D., Xiao, L., Qi, R., & Luo, R. (2017). Recent developments and applications of the MMPBSA method. Frontiers in Molecular Biosciences, 4, 87.
Willems, A. R., Tahlan, K., Taguchi, T., Zhang, K., Lee, Z. Z., Ichinose, K., Junop, M. S., & Nodwell, J. R. (2008). Crystal structures of the Streptomyces coelicolor TetR-like protein ActR alone and in complex with actinorhodin or the actinorhodin biosynthetic precursor (S)-DNPA. Journal of Molecular Biology, 376(5), 1377–1387.
(2018). Schrödinger Release 2018-4: Maestro, Schrödinger,, LLC, New York.
(2018). Schrödinger Release 2018-4: Prime, Schrödinger, LLC, New York, NY.
William, L., Jorgensen, D. S., & Maxwell, J. T. R. (1996). Development and testing of the OPLS all-atom force field on conformational energetics and properties of organic liquids. Journal of the American Chemical Society, 118, 11225–11236.
Halgren, T. A., Murphy, R. B., Friesner, R. A., Beard, H. S., Frye, L. L., Pollard, W. T., & Banks, J. L. (2004). Glide: a new approach for rapid, accurate docking and scoring. 2. Enrichment factors in database screening. Journal of Medicinal Chemistry, 47(7), 1750–1759.
Friesner, R. A., Murphy, R. B., Repasky, M. P., Frye, L. L., Greenwood, J. R., Halgren, T. A., Sanschagrin, P. C., & Mainz, D. T. (2006). Extra precision glide: docking and scoring incorporating a model of hydrophobic enclosure for protein-ligand complexes. Journal of Medicinal Chemistry, 49(21), 6177–6196.
Genheden, S., & Ryde, U. (2015). The MM/PBSA and MM/GBSA methods to estimate ligand-binding affinities. Expert Opinion on Drug Discovery, 10(5), 449–461.
Daina, A., Michielin, O., & Zoete, V. (2017). SwissADME: a free web tool to evaluate pharmacokinetics, drug-likeness and medicinal chemistry friendliness of small molecules. Scientific Reports, 7(1), 42717.
Kumari, R., Kumar, R., Open Source Drug Discovery, C., & Lynn, A. (2014). g_mmpbsa--a GROMACS tool for high-throughput MM-PBSA calculations. Journal of Chemical Information and Modeling, 54(7), 1951–1962.
Islam, M. A., & Pillay, T. S. (2019). Beta-secretase inhibitors for Alzheimer’s disease: identification using pharmacoinformatics. Journal of Biomolecular Structure & Dynamics, 37(2), 503–522.
Taha, M. O., Habash, M., Al-Hadidi, Z., Al-Bakri, A., Younis, K., & Sisan, S. (2011). Docking-based comparative intermolecular contacts analysis as new 3-D QSAR concept for validating docking studies and in silico screening: NMT and GP inhibitors as case studies. Journal of Chemical Information and Modeling, 51(3), 647–669.
Wang, J., Zhao, C., Tu, J., Yang, H., Zhang, X., Lv, W., & Zhai, H. (2019). Design of novel quinoline-aminopiperidine derivatives as mycobacterium tuberculosis (MTB) GyrB inhibitors: an in silico study. Journal of Biomolecular Structure & Dynamics, 37(11), 2913–2925.
Acknowledgments
The authors would like to extend their sincere appreciation to the Researchers Supporting Project for funding this work through Research Number (RSP-2020/138), King Saud University, Riyadh, Saudi Arabia.
Funding
The authors would like to extend their sincere appreciation to the Researchers Supporting Project for funding this work through Research Number (RSP-2019/138), King Saud University, Riyadh, Saudi Arabia.
Author information
Authors and Affiliations
Contributions
Pranjali Mahadeo Tambe: data curation, investigation and methodology; Shovonlal Bhowmick: research conceptualization, investigation and methodology, supervison, writing—review and editing; Sushil K. Chaudhary: research conceptualization, writing—review and editing; Mohammad Rizwan Khan: writing—review and editing; Saikh M Wabaidur: writing—review and editing; Mohd. Muddassir: research conceptualization, writing—review and editing; Preeti Chunarkar Patil: research conceptualization, writing—review and editing; Md Ataul Islam: research conceptualization, investigation and methodology, supervison, writing—review and editing
Corresponding author
Ethics declarations
Conflict of Interest
The authors declare that they have no competing interests.
Code Availability
Not applicable.
Computational Resource
The CHPC (www.chpc.ac.za), Cape Town, South Africa is thankfully acknowledged for computational resources and tools.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Electronic supplementary material
ESM 1
(DOCX 329 kb).
Rights and permissions
About this article

Cite this article
Tambe, P.M., Bhowmick, S., Chaudhary, S.K. et al. Structure-Based Screening of DNA GyraseB Inhibitors for Therapeutic Applications in Tuberculosis: a Pharmacoinformatics Study. Appl Biochem Biotechnol 192, 1107–1123 (2020). https://doi.org/10.1007/s12010-020-03374-y
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s12010-020-03374-y