
Abstract
A stable culture environment is the key for optimal growth and metabolic activity of microorganisms, especially in marine species, and intermittent feeding during DHA production using Schizochytrium sp. generates an unstable culture environment. To investigate the effect of unstable culture environment on the cells’ physiological status and DHA synthesis, fermentations with different feeding strategies were performed on the lab scale. The intermittent feeding strategy caused fluctuations of substrate concentration and osmotic pressure, which had a negative effect on cell division and product synthesis. The physiological status and metabolic level of Schizochytrium sp. were relatively stable under a continuous feeding strategy with a relatively stable substrate concentration of 20–25 g/L, which was beneficial for the efficient transformation of substrate, leading to an improvement of DHA productivity. This strategy was further applied to pilot scale, whereby the DHA content, DHA productivity, convert ratio of glucose to lipid and DHA reached 55.02%, 320.17 mg/(L·h), 24.35%, and 13.40%, respectively. This study therefore provides an efficient strategy for ensuring a stable culture environment for the production of DHA and similar metabolites.

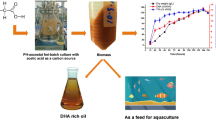
Graphical Abstract
Similar content being viewed by others

Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
Long-chain polyunsaturated fatty acids (LC-PUFAs) are critical components of the plasma membrane and play significant roles in improving physiological functions of humans [1,2,3]. Especially docosahexaenoic acid (DHA, C22:6), which is the most representative omega-3 LC-PUFA, has received worldwide attention due to its health benefits to both infants and adults [4, 5]. There have been many clinical studies on DHA supplementation, which showed that this LC-PUFA is important for the normal development of the visual and nervous system tissues in infants and young children, while also reducing the risk of cardiovascular disease, alleviating depressive symptoms, and contributing to immune function in adults [6, 7]. Consequently, DHA is widely used as a dietary health supplement. Marine fish used to be the main source of DHA, but the safety, availability, and sustainability of fish oil have been compromised by increasingly severe marine pollution and overfishing [8, 9].
Research has shown that DHA is produced directly by marine microorganisms and phytoplankton, while marine fish accumulate significant amounts of DHA by consuming microalgae [7, 10]. The microalga Schizochytrium sp. has been the subject of a substantial amount of research due to its high DHA content and growth rate and is thus considered as a potentially commercially viable host for DHA production [11, 12]. Consequently, the use of Schizochytrium sp. fermentation to produce DHA-rich oil has become a hot topic in the scientific community [7]. The development and optimization of fermentation processes is one of the key problems in production improvement. A stable culture environment is especially important for the growth and productivity of cultured microorganisms. In the DHA production processes by Schizochytrium sp., the optimal conditions for cell proliferation, lipid accumulation, and DHA synthesis are not the same, with different pH, temperature, and oxygen preferences for each [13, 14]. Diverse fermentation strategies have been developed aiming to improve the biomass and the content of DHA in total lipids [9, 15]. A large number of DO-control strategies (dissolved oxygen: DO) have been investigated, which were usually based on the stage characteristics of fermentations using Schizochytrium sp., and also achieved good results [1, 16]. Ganuza et al. developed a NH4-pH-auxostat fed-batch system for Schizochytrium sp., and they achieved the fastest possible growth rate in the stage of cell reproduction [17]. Hu et al. proposed a novel osmotic pressure control strategy for Schizochytrium sp. and obtained a maximum cell dry weight of 56.95 g/L as well as the highest DHA content in total lipid of 55.21% [13]. Zeng et al. also employed a temperature shift strategy for efficient DHA production by Schizochytrium sp., reaching a DHA percentage of up to 51.98% (in total fatty acids) with a DHA production of 6.05% (in dry cell weight) [18]. In addition, there were a number of studies on the effect of nutrients such as nitrogen, sulfur, and microelements on DHA production [19,20,21]. In spite of the efforts that have been made previously, no study has been developed for the effect of nonhomogeneous culture environment on DHA production.
The stability of the culture environment is essential for microbial fermentations, especially in large-scale bioreactors. This includes a stable nutrition supply, osmotic pressure, pH, temperature etc. The cells’ transcriptional and metabolic activity will be changed when they are cultured in a fluctuating environment. Moreover, the yield and quality of target products can also be affected [22,23,24]. Most industrial processes for the production of DHA-rich oil rely on fed-batch fermentation operations. In intermittent feeding strategies, the substrate concentration is kept above 15 g/L and can reach as high as 100 g/L [25]. In the stage of lipid accumulation, the fermentation parameters including pH, DO, etc., reach the stabilization phase, and only the substrate concentration remains in a state of constant fluctuation [22, 26]. It has been reported that a high substrate concentration can result in substrate inhibition which can negatively affect the cell growth and lipid accumulation, while a low substrate concentration may prolong the fermentation period and decrease DHA productivity, so the high and low substrate concentrations are bad for cell growth and DHA accumulation [9]. Li et al. reported that high substrate concentrations can inhibit the activity of important enzymes such as glycerol kinase and glycerol-3-phosphate dehydrogenase [27]. Moreover, the addition of high substrate concentrations was reported to increase the osmolarity of the medium [9, 28]. Thus, fluctuations of substrate concentration can also lead to constant changes in osmotic pressure, which in turn has a significant effect on cell proliferation, and cellular osmotic adjustment is also a highly important biological process in marine microorganisms [13]. Therefore, if the cells repeatedly enter environments of high or low osmotic pressure and insufficient or overabundant substrate, these fluctuations can have a significant impact on DHA production. To our best knowledge, no research has been done to study the effect of a fluctuating substrate supply on DHA production.
In the present work, we aimed to investigate the effects of environments with different substrate supply and osmotic pressure on the cells’ physiological status and changes of DHA synthesis and lipid components during the fermentation. Two types of feeding strategies were tested in a 50-L bioreactor—an intermittent and a continuous feeding strategy. Based on this, a rational feeding strategy was developed. Finally, pilot-scale fermentations were developed using the optimal feeding strategy with a controlled substrate concentration and osmotic pressure environment to verify its potential for industrial production. This approach will hopefully provide guidance for the large-scale production of DHA and similar products.
Materials and Methods
Microorganism
The Schizochytrium sp. strain HX-308 (CCTCC M209059) used in this study was obtained from the China Center for Type Culture Collection and maintained in 20% (v/v) glycerol at − 80 °C [1, 25].
Culture Conditions
The seed medium was composed of glucose (50 g/L) and yeast extract (0.4 g/L) in artificial seawater [26]. The fermentation broth consisted of glucose (70 g/L) and monosodium glutamate (20 g/L) in artificial seawater.
For lab-scale fermentations, the culture stock preserved in glycerol was transferred into a 250-mL baffled shake flask with 50 mL medium and cultivated for 24 h at 30 °C. After culturing for three generations, the culture (1%, v/v) was inoculated to a 10-L seed bioreactor (Shanghai Guoqiang, China) with 7 L liquid volume and cultivated for 24 h at 30 °C. Then, the mature seed (10%, v/v) was used to inoculate a 50-L fermenter (Shanghai Guoqiang, China) with 35 L medium and cultured at 200 rpm 1.0 vvm and 30 °C for 120 h. The pH and DO were monitored using a sensor electrode (Mettler Toledo, Switzerland). The bioreactor was equipped with an on-line gas analysis device (Shanghai Shunyuhengping, China) which monitored the concentration of O2 and CO2 in the inlet and exhaust. Furthermore, real-time data collection software was used to monitor the oxygen uptake rate (OUR) and respiratory quotient (RQ) on-line [29]. Additionally, every 12 h, 100 mL samples were taken and measure the relevant parameters, such as the concentrations of glucose and nitrogen, biomass, lipid content, etc.
To investigate the effect of different feeding strategies and substrate concentrations, an intermittent and a continuous feeding strategy was established. In the intermittent feeding strategy, the 800-g/L glucose solution was fed into the bioreactor to keep the substrate concentration above 15 g/L, while the highest substrate concentration could reach up to 100 g/L. For the continuous feeding strategy, which can provide a stable culture environment, three different substrate concentration ranges (15–20 g/L, 20–25 g/L, and 25–30 g/L) were designed and executed in three independent fermentation processes. All experiments were carried out in duplicate.
The pilot-scale fermentations were performed in a 500-L bioreactor. The seed culture (10%, v/v) was cultivated for two generations (24 h for each generation) in the 10- and 50-L seed bioreactors, respectively. The matured second-stage seed (10%, v/v) was used to inoculate the 500-L bioreactor, which contained 350 L of medium. The substrate feedback control was used for the optimized feeding strategy during the fermentation process. The other fermentation parameters were the same as for the lab-scale fermentation.
Analytical Methods
The measurements of biomass, total lipid content, fatty acid composition, and fatty acid methyl ester (FAMEs) preparation were the same as in our previous studies [21, 30]. The glucose and glutamate concentrations were measured enzymatically using an SBA-40C bioanalyzer (Institute of Biology, Shandong Academy of Sciences, China).
The osmotic pressure in the medium was calculated using the equation P = cRT, where P is osmotic pressure of the solution, c is the total ion concentration (for non-electrolyte substances such as glucose) or molecular concentration (for electrolytes such as inorganic salts), R is the gas constant (R = 8.315 L·kPa/(K·mol)), and T is the absolute temperature of the solution, as described in our previous studies [13].
Results and Discussion
Effect of Intermittent and Continuous Feeding Strategies on the DHA Fermentation
As shown in Fig. 1 and Table 1, there were significant differences in each parameter of DHA production, such as substrate assimilation, biomass growth, and lipid and DHA synthesis, when using the different intermittent and continuous feeding strategies.

Fermentation profiles showing the growth characteristics of Schizochytrium sp. HX-308. cultured using an intermittent feeding strategy (Control, black squares), and continuous feeding strategies with controlled glucose concentrations of 15–20, 20–25, and 25–30 g/L (red downward triangles, green circles, blue upright triangles, respectively). a Glucose feeding (g/L). b Monosodium glutamate consumption, MGS (g/L). c Biomass (g/L). d Total lipids (g/L)
The glucose concentration profiles of fermentations using each feeding strategy are shown in Fig. 1a. The glucose concentration in the intermittent feeding strategy underwent large fluctuations, reaching up to 74 g/L and falling down to 6 g/L, which may have led to a nonhomogeneous culture environment. The cells’ transcriptional and metabolic activity will be changed when exposed to these fluctuations of the culture environment. Moreover, the yield and quality of target products could also be affected [23, 24]. By contrast, the glucose concentration in each continuous feeding strategy was relatively constant. The amounts of glucose consumed by the four continuously fed cultures were respectively 11.81, 9.32, 10.27, and 10.15 kg (Table 1). The amount of glucose consumed was the highest in the intermittent feeding strategy, which demonstrated that Schizochytrium sp. has a higher substrate assimilation capacity and improved energy metabolism in a stable culture environment.
However, nitrogen consumption was only marginally influenced by these feeding strategies, and the time of nitrogen exhaustion in each feeding strategy was almost the same (Fig. 1b). Nitrogen limitation activates lipid accumulation in Schizochytrium sp., which means that the cells enter the phase of active lipid accumulation at the same time in each strategy. Moreover, the initial lipid content in the biomass was also practically the same at 5%. With the continuing fermentation process, differences of lipid content appeared. The highest lipid content (68.74 g/L) was obtained using the continuous feeding strategy with a glucose concentration of 20–25 g/L, and the lowest lipid content (45.43 g/L) was obtained using the continuous feeding strategy with a glucose concentration at 15–20 g/L. The lipid contents in intermittent feeding strategy and continuous feeding strategy with a glucose concentration of 25–30 g/L were 61.03 and 65.58 g/L. The change trend of biomass reflected the lipid content in the intermittent and continuous feeding strategies, as shown in Fig. 1c, d.
Interestingly, the highest biomass (117.58 g/L), lipid content (68.74 g/L), and DHA (54.68%) content were all achieved using the continuous feeding strategy with a glucose concentration of 20–25 g/L (Table 1). The lowest fermentation yield was obtained using the intermittent feeding strategy and continuous feeding strategy with a glucose concentration of 15–20 g/L. There seems to be a marginal value of glucose concentration, so that lipid synthesis and cell growth were inhibited when the glucose concentration was below or above this value. Notably, the fluctuations of the culture environment in the intermittent feeding strategy appear to have caused significant damage to the cells, so that the DHA content, DHA productivity, and conversion ratio of glucose to lipid and DHA were only 50.81%, 258.41 mg/(L·h), 18.09%, and 9.19%, respectively. By contrast, the highest DHA productivity (313.23 mg/(L·h)), conversion ratio of glucose to lipid (23.43%), and DHA (12.81%) were obtained using the continuous feeding strategy with a glucose concentration of 20–25 g/L. These values represent increases of 21.21, 29.52, and 39.39% over the intermittent feeding strategy. Thus, there was considerable substrate waste in the intermittent feeding strategy. These results also demonstrated that a proper substrate concentration and stable culture environment are beneficial for the efficient use of glucose and the efficient synthesis of oil and DHA. At the same time, the cells may have been subjected to potential damage caused by an unsuitable substrate concentration and fluctuations of the culture environment [31]. Hu et al. also came to an analogous conclusion in environments with different osmotic pressure [13].
Effects of the Different Feeding Strategies on Bioprocess Parameters
Procedure parameters were recorded during all the fermentations, as shown in Fig. 2. The DO showed a continuous decreasing trend until reaching zero, and subsequently, it remained at a constant low level (Fig. 2a). Thus, all four cultures entered a DO-limited state. Moreover, the time when DO reached zero was almost at the same time in all culture modes, which indicates that all the cultures had similar oxygen supply and demand levels. A similar tendency was evident in real-time change of pH for these four cultures, and the pH reached and maintained a constant level at a similar time in subsequent fermentation phase (Fig. 2b). This phenomenon was also analogous to what we observed in our earlier study and reflected the sequential metabolic changes during the utilization of the various classes of nitrogen sources in the broth [32, 33]. These observations imply that the cells had a similar physiological status before the cultures entered the DO- and nitrogen-limited state.

Time course profiles of fermentation utilizing Schizochytrium sp. HX-308 in fed-batch mode in conjunction with an intermittent feeding strategy (control, black squares), and continuous feeding strategies with substrate concentrations of 15–20, 20–25, and 25–30 g/L (red downward triangles, green circles, and blue upward triangles, respectively). a Dissolved oxygen. DO (%). b pH. c Respiratory quotient, RQ. d Osmotic pressure (kPa)
In addition, RQ, as a critical physiological parameter, was also investigated. It increased rapidly with the decrease of DO until the culture reached the DO-limited state and subsequently remained in a dynamic equilibrium, as shown in Fig. 2c. However, the RQ value of the cultures with different feeding strategies reached different levels during the final stages, even though the fermentations had the same oxygen supply levels. Thus, there was a difference in cell metabolism among the four different fermentations, and the RQ profiles were in agreement with the DHA content in total lipids. In Schizochytrium sp., the PUFAs are synthesized through the non-oxygen-requiring PKS pathway. Consequently, an increase of PUFA synthesis reduces the consumption of O2 and increases the release of CO2, increasing the RQ value [1, 34, 35]. On the contrary, a lower RQ value (i.e., RQ ≈ 1.0) signified that the Kreb’s cycle is more active and substrate is preferentially transformed into H2O and CO2 [36]. The highest RQ value was obtained using the continuous feeding strategy with a substrate concentration of 20–25 g/L, and the lowest value was achieved in the continuous feeding strategy with a substrate concentration of 15–20 g/L. The DHA contents in total lipids obtained using these two strategies were 54.68 and 49.32%, respectively, which further confirmed the relationship between RQ and DHA content in total lipids.
The substrate assimilation, cell growth, lipid synthesis, DHA production, and process parameters showed significant differences, which were likely caused by the differences of glucose concentration and osmotic pressure during the fermentation (Fig. 2d). The osmotic pressure was subjected to large fluctuations in the intermittent feeding strategy due to the large swings of glucose concentration, while it was relatively constant in each of the continuous feeding strategies. The genetic, transcriptional, and metabolic levels of cells change when they are cultured under these fluctuation culture conditions. In non-growing cells, an increase in external osmolarity will cause loss of water from the cell, whereas a decrease in external osmolarity will cause uptake of water. To reduce or eliminate these osmotically induced changes in the amount of cellular water, many osmotically stressed growing cells exhibit active responses in which the amounts of various cellular solutes are changed by biosynthesis or transport [37]. Moreover, the yield and quality of target products could also be influenced. A certain turgor pressure of the cell is one of the necessary conditions for cell division and synthesis of products [38]. Thus, the stable glucose concentration that was kept at 20–25 g/L could provide a proper osmotic pressure and suitable substrate concentration for cell proliferation and products accumulated during the fermentation. Under these conditions, the DHA content in total lipids and the conversion ratio of glucose to DHA reached their highest values of 54.68 and 12.81%, respectively. However, the opposite was true when the glucose concentration was maintained at 15–20 g/L. Therefore, this continuous feeding strategy based on osmotic pressure can be suggested as an effective method for improving DHA production. Subsequently, the lipid components were also investigated to reveal the interrelation between substrate concentrations/osmotic pressure and lipid metabolism in the different feeding strategies.
Changes of Fatty Acid Composition in the Different Feeding Strategies
The fatty acid composition analysis showed that more than 90% of the total lipids extracted from Schizochytrium sp. were composed of DHA, DPA (docosapentenoic acid; C22:5), tetradecanoic acid (C14:1), myristic acid (C14:0), hexadecanoic acid (C16:0), and stearic acid (C18:0). In this organism, polyunsaturated fatty acids, such as DHA and DPA, are mainly synthesized by the PKS pathway, while the FAS pathway mainly synthesized the monounsaturated and saturated fatty acids, such as C14:1, C14:0, C16:0, and C18:0 [1, 39]. Therefore, the relative activity of the PKS and FAS pathways is reflected in the content of PUFAs and saturated or monounsaturated fatty acids in total lipids [1]. The abundance of polyunsaturated fatty acids was also highly consistent with the RQ profiles of each feeding strategy. This was in agreement with our previous study, which has shown that high RQ values indicate a high relative activity of the PKS pathway [1, 40]. The content of monounsaturated and saturated fatty acids constantly decreased during the fermentation, while the content of PUFAs, and especially DHA, constantly increased simultaneously (Table. 2). Importantly, PUFAs content in the continuous feeding strategy with a substrate concentration of 20–25 g/L was consistently above that of the other strategies, which means that the PKS pathway was more active with this feeding strategy. However, this strategy also produced the lowest content of DHA with a substrate concentration at 15–20 g/L. Interestingly, the content of squalene showed an apparent fluctuation in the intermittent feeding strategy. These differences were most likely caused by the drastic fluctuation of glucose concentration and osmotic pressure. The water activity in the broth is changed by the fluctuation of substrate concentration, which affects the cells’ metabolic activity and the composition of the cytoplasm. Moreover, the cells can autolyze when exposed to an extremely unsuitable osmotic pressure. Hence, the cells must be cultured at a proper osmotic pressure and substrate concentration. Akimoto et al. reported that optimal osmotic pressure can promote cell grown and the synthesis of EPA, another important PUFA, similar to DHA [41]. Thus, the continuous feeding strategy with a substrate concentration of 20–25 g/L was able to provide a proper osmotic pressure for DHA synthesis and increased the relative activity of the PKS pathway. The phenomenon of fluctuating environments is constantly to appear in industrial-scale fermentation process, which implies that osmotic pressure may be an underestimated factor in the fermentation industry [22].
Scale-Up of Fermentation Using the Optimized Feeding Strategy in the Pilot-Scale Bioreactor
The results of preliminary lab-scale fermentations showed that maximum values of biomass, lipid content, DHA content of lipids, DHA productivity, and conversion ratio of glucose to total lipids and DHA were obtained using the continuous feeding strategy with a glucose concentration of 20–25 g/L. To verify the validity of continuous feeding strategy in the scale-up of DHA production, the optimal strategy and intermittent feeding strategies were consequently tested in a 500-L pilot-scale bioreactor.
The results obtained in the 500-L bioreactor using the intermittent and continuous feeding strategies were compared in Table 3. A biomass of 122.37 g/L, lipid content of 69.83 g/L, and DHA productivity of 320.17 mg/(L·h) were achieved using the optimized continuous feeding strategy with a glucose concentration of 20–25 g/L, which represent increases of 25.53, 18.52, and 37.68% over the intermittent feeding strategy, respectively. The DHA content in total lipids increased to 55.02% using the continuous feeding strategy (20–25 g/L). This greatly exceeded the minimum level required for marketing (≥ 35%) while also showing an increase of 13.92% beyond the intermittent feeding strategy. At the same time, the maximal conversion ratios of glucose to total lipids and DHA achieved using this process were 24.35 and 13.40%, which was respectively 38.43 and 43.62% higher than what was obtained using the intermittent feeding strategy. The results thus clearly indicate that the continuous feeding strategy with a glucose concentration of 20–25 g/L provided a stable glucose supply and culture environment, which prevented harmful fluctuations of the cells’ transcriptional and metabolic activity. This finding has the potential to effectively promote the industrialization of DHA fermentation and improve the yield of both DHA and total lipids in microalgae. Therefore, this research offers a significant improvement over preceding process arrangements and will hopefully also be applicable to the industrialization of other biological products in the future.
Conclusions
The effect of a stable substrate supply and osmotic pressure environment on the cells’ physiological status, DHA synthesis, and lipid composition was studied. In lab-scale fermentations, the continuous feeding strategy provided the most suitable culture environment for DHA production. The optimized strategy was further tested in pilot-scale fermentation. The DHA content, DHA productivity, conversion ratio of glucose to total lipids, and DHA reached 55.02%, 320.17 mg/(L·h), 24.35%, and 13.40%, respectively, representing increases of 37.68, 13.92, 38.43, and 43.62% over the intermittent feeding strategy. The results clearly indicate that this strategy has the potential to be applied in the industry.
References
Guo, D. S., Ji, X. J., Ren, L. J., Li, G. L., Yin, F. W., & Huang, H. (2016). Development of a real-time bioprocess monitoring method for docosahexaenoic acid production by Schizochytrium sp. Bioresource Technology, 216, 422–427.
Ji, X. J., Mo, K. Q., Ren, L. J., Li, G. L., Huang, J. Z., & Huang, H. (2015). Genome sequence of Schizochytrium sp. CCTCC M209059, an effective producer of docosahexaenoic acid-rich lipids. Genome Announcements, 3, e00819–e00815.
Safdar, W., Zan, X., Shamoon, M., Sharif, H. R., Mukama, O., Tang, X., & Song, Y. (2017). Effects of twenty standard amino acids on biochemical constituents, docosahexaenoic acid production and metabolic activity changes of Crypthecodinium cohnii. Bioresource Technology, 238, 738–743.
Ren, L. J., Sun, X. M., Ji, X. J., Chen, S. L., Guo, D. S., & Huang, H. (2017). Enhancement of docosahexaenoic acid synthesis by manipulation of antioxidant capacity and prevention of oxidative damage in Schizochytrium sp. Bioresource Technology, 223, 141–148.
Sakarika, M., & Kornaros, M. (2017). Kinetics of growth and lipids accumulation in Chlorella vulgaris during batch heterotrophic cultivation: effect of different nutrient limitation strategies. Bioresource Technology, 243, 356–365.
Guo, D. S., Ji, X. J., Ren, L. J., Li, G. L., Sun, X. M., & Huang, H. (2017). Development of a scale-up strategy for fermentative production of docosahexaenoic acid by Schizochytrium sp. Chemical Engineering Science, 176, 600–608.
Cui, G. Z., Ma, Z., Liu, Y. J., Feng, Y., Sun, Z., Cheng, Y., Song, X., & Cui, Q. (2016). Overexpression of glucose-6-phosphate dehydrogenase enhanced the polyunsaturated fatty acid composition of Aurantiochytrium sp. SD116. Algal Research, 19, 138–145.
Ji, X. J., Ren, L. J., & Huang, H. (2015). Omega-3 biotechnology: a green and sustainable process for omega-3 fatty acids production. Frontiers in Bioengineering and Biotechnology, 3, 158.
Singh, D., Barrow, C. J., Puri, M., Tuli, D. K., & Mathur, A. S. (2016). Combination of calcium and magnesium ions prevents substrate inhibition and promotes biomass and lipid production in thraustochytrids under higher glycerol concentration. Algal Research, 15, 202–209.
Furlan, V. J., Maus, V., Batista, I., & Bandarra, N. M. (2017). Production of docosahexaenoic acid by Aurantiochytrium sp. ATCC PRA-276. Brazilian Journal of Microbiology, 48(2), 359–365.
Ren, L. J., Hu, X. C., Zhao, X. Y., Chen, S. L., Wu, Y., Li, D., Yu, Y. D., Geng, L. J., Ji, X. J., & Huang, H. (2017). Transcriptomic analysis of the regulation of lipid fraction migration and fatty acid biosynthesis in Schizochytrium sp. Scientific Reports, 7(2017), 3562.
Ren, L. J., Zhuang, X. Y., Chen, S. L., Ji, X. J., & Huang, H. (2015). Introduction of omega-3 desaturase obviously changed the fatty acid profile and sterol content of Schizochytrium sp. Journal of Food Agriculture and Environment, 63, 9770–9776.
Hu, X. C., Ren, L. J., Chen, S. L., Zhang, L., Ji, X. J., & Huang, H. (2015). The roles of different salts and a novel osmotic pressure control strategy for improvement of DHA production by Schizochytrium sp. Bioprocess and Biosystems Engineering, 38, 2129–2136.
Lian, M., Huang, H., Ren, L. J., Ji, X. J., Zhu, J., & Jin, L. (2010). Increase of docosahexaenoic acid production by Schizochytrium sp. through mutagenesis and enzyme assay. Applied Biochemistry and Biotechnology, 162(4), 935–941.
Steinrücken, P., Erga, S. R., Mjøs, S. A., Kleivdal, H., & Prestegard, S. K. (2017). Bioprospecting North Atlantic microalgae with fast growth and high polyunsaturated fatty acid (PUFA) content for microalgae-based technologies. Algal Research, 26, 392–401.
Schörken, U., & Kempers, P. (2009). Lipid biotechnology: industrially relevant production processes. European Journal of Lipid Science and Technology, 111, 627–645.
Ganuza, E., Anderson, A. J., & Ratledge, C. (2008). High-cell-density cultivation of Schizochytrium sp. in an ammonium/pH-auxostat fed-batch system. Biotechnology Letters, 30(2008), 1559–1564.
Zeng, Y., Ji, X. J., Lian, M., Ren, L. J., Jin, L. J., Ouyang, P. K., & Huang, H. (2011). Development of a temperature shift strategy for efficient docosahexaenoic acid production by a marine fungoid protist, Schizochytrium sp. HX-308. Applied Biochemistry and Biotechnology, 164(3), 249–255.
Liu, T., Li, Y., Liu, F., & Wang, C. (2016). The enhanced lipid accumulation in oleaginous microalga by the potential continuous nitrogen-limitation (CNL) strategy. Bioresource Technology, 203, 150–159.
Ling, X., Guo, J., Liu, X., Zhang, X., Wang, N., Lu, Y., & Ng, I. S. (2015). Impact of carbon and nitrogen feeding strategy on high production of biomass and docosahexaenoic acid (DHA) by Schizochytrium sp. LU310. Bioresource Technology, 184, 139–147.
Sun, L. N., Ren, L. J., Zhuang, X. Y., Ji, X. J., Yan, J. C., & Huang, H. (2014). Differential effects of nutrient limitations on biochemical constituents and docosahexaenoic acid production of Schizochytrium sp. Bioresource Technology, 159, 199–206.
Guo, D. S., Ji, X. J., Ren, L. J., Li, G. L., & Huang, H. (2017). Improving docosahexaenoic acid production by Schizochytrium sp. using a newly designed high-oxygen-supply bioreactor. AICHE Journal, 63, 4278–4286.
Duan, S., Yuan, G., Zhao, Y., Ni, W., Luo, H., Shi, Z., & Liu, F. (2013). Simulation of computational fluid dynamics and comparison of cephalosporin C fermentation performance with different impeller combinations. Korean Journal of Chemical Engineering, 30, 1097–1104.
Zou, X., Xia, J. Y., Chu, J., Zhuang, Y. P., & Zhang, S. L. (2012). Real-time fluid dynamics investigation and physiological response for erythromycin fermentation scale-up from 50 L to 132 m3 fermenter. Bioprocess and Biosystems Engineering, 35(5), 789–800.
Qu, L., Ren, L. J., & Huang, H. (2013). Scale-up of docosahexaenoic acid production in fed-batch fermentation by Schizochytrium sp. based on volumetric oxygen-transfer coefficient. Biochemical Engineering Journal, 77, 82–87.
Zhao, X. Y., Ren, L. J., Guo, D. S., Wu, W. J., Ji, X. J., & Huang, H. (2016). CFD investigation of Schizochytrium sp. impeller configurations on cell growth and docosahexaenoic acid synthesis. Bioprocess and Biosystems Engineering, 39(8), 1297–1304.
Li, J., Liu, R., Chang, G., Li, X., Chang, M., Liu, Y., Jin, Q., & Wang, X. (2015). A strategy for the highly efficient production of docosahexaenoic acid by Aurantiochytrium limacinum SR21 using glucose and glycerol as the mixed carbon sources. Bioresource Technology, 177, 51–57.
Yan, J., Cheng, R., Lin, X., You, S., Li, K., Rong, H., & Ma, Y. (2013). Overexpression of acetyl-CoA synthetase increased the biomass and fatty acid proportion in microalga Schizochytrium. Applied Microbiology and Biotechnology, 97(5), 1933–1939.
Zhang, S. L., Chu, J., & Zhuang, Y. P. (2004). A multi-scale study of industrial fermentation processes and their optimization. Advances in Biochemical Engineering/Biotechnology, 87, 97–150.
Sun, X. M., Ren, L. J., Ji, X. J., Chen, S. L., Guo, D. S., & Huang, H. (2016). Adaptive evolution of Schizochytrium sp. by continuous high oxygen stimulations to enhance docosahexaenoic acid synthesis. Bioresource Technology, 211, 374–381.
Liu, L., Xu, Q., Li, Y., Shi, Z., Zhu, Y., Du, G., & Chen, J. (2007). Enhancement of pyruvate production by osmotic-tolerant mutant of Torulopsis glabrata. Biotechnology and Bioengineering, 97, 825.
Ren, L. J., Ji, X. J., Huang, H., Qu, L., Feng, Y., Tong, Q. Q., & Ouyang, P. K. (2010). Development of a stepwise aeration control strategy for efficient docosahexaenoic acid production by Schizochytrium sp. Applied Microbiology and Biotechnology, 87(5), 1649–1656.
Wu, S. T., Yu, S. T., & Lin, L. P. (2005). Effect of culture conditions on docosahexaenoic acid production by Schizochytrium sp. S31. Process Biochemistry, 40, 3103–3108.
Ratledge, C. (2014). The role of malic enzyme as the provider of NADPH in oleaginous microorganisms: a reappraisal and unsolved problems. Biotechnology Letters, 36(8), 1557–1568.
Hauvermale, A., Kuner, J., Rosenzweig, B., Guerra, D., Diltz, S., & Metz, J. G. (2006). Fatty acid production in Schizochytrium sp.: involvement of a polyunsaturated fatty acid synthase and a type I fatty acid synthase. Lipids, 41, 739.
Xiong, Z. Q., Guo, M. J., & Guo, Y. X. (2010). RQ feedback control for simultaneous improvement of GSH yield and GSH content in Saccharomyces cerevisiae T65. Enzyme and Microbial Technology, 46, 598–602.
Record, M. T., Courtenay, E. S., Cayley, D. S., & Guttman, H. J. (1998). Responses of E. coli to osmotic stress: Large changes in amounts of cytoplasmic solutes and water. Trends in Biochemical Sciences, 23(4), 143–148.
Hohmann, S. (2002). Osmotic stress signaling and osmoadaptation in yeasts. Microbiology and Molecular Biology Reviews, 66(2), 300–372.
Metz, J. G., Roessler, P., Facciotti, D., Levering, C., Dittrich, F., Lassner, M., Valentine, R., Lardizabal, K., Domergue, F., & Yamada, A. (2001). Production of polyunsaturated fatty acids by polyketide synthases in both prokaryotes and eukaryotes. Science, 293(5528), 290–293.
Chang, G. F., Wu, J., Jiang, C., Tian, G., Wu, Q., Chang, M., & Wang, X. (2014). The relationship of oxygen uptake rate and k(L)a with rheological properties in high cell density cultivation of docosahexaenoic acid by Schizochytrium sp. S31. Bioresource Technology, 152, 234–240.
Akimoto, M., Ishii, T., Yamagaki, K., Ohtaguchi, K., Koide, K., & Yazawa, K. (1991). Metal salts requisite for the production of eicosapentaenoic acid by a marine bacterium isolated from mackerel intestines. Journal of the American Oil Chemists Society, 68, 504–508.
Funding
This work was financially supported by the National Key Research and Development Program of China (No. 2018YFC1604104), the National Natural Science Foundation of China (Nos. 21376002, 21476111, and 21606192), the Jiangsu Provincial Natural Science Foundation (No. BK20131405), and the Priority Academic Program Development of Jiangsu Higher Education Institutions.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of Interest
The authors declare that they have no conflict of interest.
Ethical Statement
This article does not contain any studies with human participants or animals performed by any of the authors.
Informed Consent
Informed consent was obtained from all individual participants included in the study.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Highlights
• Intermittent feeding caused fluctuation of substrate content and osmotic stress.
• The Physiological of Schizochytrium sp in unstable culture environment was studied.
• Fatty acid metabolism in Schizochytrium sp at different osmotic stress was studied.
• The DHA efficient production was realized by a continuous feeding strategy.
Rights and permissions
About this article

Cite this article
Guo, DS., Tong, LL., Ji, XJ. et al. Development of a Strategy to Improve the Stability of Culture Environment for Docosahexaenoic Acid Fermentation by Schizochytrium sp.. Appl Biochem Biotechnol 192, 881–894 (2020). https://doi.org/10.1007/s12010-020-03298-7
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s12010-020-03298-7