
Abstract
Development of an ideal process for reduction of food phytates using microbial phytases is a demanding task by all food and feed industries all over the world. Phytase production by Bacillus subtilis subsp. subtilis JJBS250 isolated from soil sample was optimized in submerged fermentation using statistical tools. Among all the culture variables tested, sucrose, sodium phytate and Tween-80 were identified as the most significant variables using the Placket–Burman design. Further optimization of these variables resulted in a 6.79-fold improvement in phytase production (7170 U/L) as compared to unoptimized medium. Supplementation of microbial phytases (fungal and bacterial) resulted in improved bioavailability of nutritional components with the concomitant liberation of inorganic phosphorus, reducing sugar, soluble protein and amino acids, thus mitigating anti-nutritional properties of phytic acid.
Similar content being viewed by others


Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
Phytic acid, the organic form of phosphorus present in plant-derived food is not available to monogastric animals due to lack of an insufficient level of phytases in their digestive tracts, therefore, undigested phytic acid excreted into the environment and leads to phosphorus pollution [1–3]. Phytic acid chelates different mineral ions, thus making them unavailable and precipitates several proteins and enzymes, thus inhibiting their activities and digestion, therefore, acting as an anti-nutritional factor [4, 5]. Phytases hydrolyze phytic acid in a stepwise manner and release inorganic phosphorus and myo-inositol phosphate derivatives making bioavailability of phosphorus for absorption [5, 6]. Thus, the supplementation of phytase in animal feedstuffs is only the solution to mitigate both the phosphorus pollution and anti-nutritional behaviour of plant phytate.
On the basis of catalytic function and structure, four different classes of phytases have been proposed [2]. These four classes include histidine acid phosphatases (HAP), β-propeller phytases (BPP), purple acid phytases (PAP) and cysteine phosphatases (CP) [5]. Most of the bacterial phytases belonging to the Bacillus spp. are grouped under β-propeller phytases, alkaline in nature and metal dependent, whereas phytase from Sporotrichum thermophile belongs to histidine acid phosphatases and are characterized by conserved active site hepta-peptide motif RHGXRXP at the N-terminal end and active dipeptide motif is HD at the C-terminal end, acidic in nature and metal independent [3–5].
Several physical and chemical factors affect the phytase production during microbial fermentation which need to be optimized. A conventional method like one variable at a time approach is very laborious, expensive and time consuming. This method delimits the interaction of several factors in a single time and resulting into the low enzyme production [7–9]. To overcome these problems, several researchers used response surface methodology/statistical approach to screen and optimize several factors for phytase production. This method is efficient to study the effect of several factors and their interaction at a time which leads to decrease the cost of enzyme production and increase the yield of enzyme production [10–15].
The present investigation is made to evaluate the screen important factors and improve the phytase production by Bacillus subtilis subsp. subtilis JJBS250 by using the Plackett–Burman design followed by response surface methodology (RSM) in submerged fermentation. Microbial phytases have different potential application in ameliorating food nutrition; therefore, a process has been developed using B. subtilis and S. thermophile phytases in efficient dephytinization of animal feed.
Materials and Methods
Microbial Culture and Enzyme Production
A potent phytase producer B. subtilis subsp. subtilis JJBS250 was isolated from soil samples and identified on the basis of morphological features and biochemical and molecular level studies. The medium containing (g/L) sodium phytate 0.03, calcium chloride 2, sucrose 20, magnesium sulphate 0.5, potassium chloride 0.5, ferrous sulphate 0.01, manganese sulphate 0.01, yeast extract 20 (pH 7.0) was employed for phytase production and culture was incubated at 37 °C and 200 rpm for 48 h. The fermented broth was centrifuged at 10,000 rpm at 4 °C and cell free supernatant was assayed for phytase activity. The nucleotide sequences for 16S rDNA has been submitted in GenBank under accession no KC609372.1.
The thermophilic mould, S. thermophile BJTLR50 syn. Myceliophthora thermophila isolated from a soil sample was grown in optimized medium at 45 °C and 250 rpm for phytase production as described by Singh and Satyanarayana [11]. The cell-free culture filtrate was analysed for phytase activity.
Phytase Assay
Phytase assay was carried out by determining the amount of inorganic phosphorus liberated from 1 mM sodium phytate in 1 M tris-HCl buffer (pH 7.0) containing 1 mM CaCl2 at 50 °C for 10 min as described by Fiske and Subbarow [16], while the assay conditions for the fungal phytase were the same as described by Kumari et al. [14]. One unit of phytase is the amount of enzyme required for the liberation of 1 nmol of inorganic phosphorus sec− 1 under standard conditions.
Statistical Optimization of Culture Conditions for Phytase Production by B. subtilis subsp. subtilis JJBS250
Culture parameters for phytase production by B. subtilis were optimized by using the Plackett–Burman design followed by RSM.
The important physico-chemical parameters were selected using the Plackett–Burman design (Table 1) [17]. According to Plackett–Burman, total number of experiments carried out is n + 1, where n is the number of variables, each studied at two (high and low) levels. The effect of each variable is calculated by the difference between the average of the results at high and low level of that variable determined by following equation:
Where E(Xi) represents the effect of the test variable and Pi+ and Pi− are the value of phytase production obtained at high and low level, respectively. N represents the number of experiments carried out during the study.
Three independent variables, sucrose (A), sodium phytate (B) and Tween-80 (C) were chosen to study their effect on phytase production using central composite design (CCD) of RSM in synthetic medium (g L−1: yeast extract 5.0, MgSO4 0.5, CaCl2 0.5) at pH 5.0 and an incubation period of 24 h. These variables were studied at five (−α, −1, 0, +1, +α) different levels (Table 2). The average phytase production was taken as the response (Y) in each set of experiments. The analysis of the data was carried out using the following equation:
Where Y represents the phytase production, β0 is intercept, β1, β2, β3, and β4 are the linear coefficients; β11, β22, β33, and β44 are the squared coefficients; β12, β13, β23, and β34 are the interaction coefficients and A, B, C, A2, B2, C2, AB, AC, and BC are the selected variables and their interaction. A total number of 20 experiments in the design were analysed to study their interaction and to generate three-dimensional plots using ‘Design Expert 6.0.7’ software.
Process Development for Amelioration of Food and Feed Nutrition Using Microbial Phytases
Partial purification of microbial phytases were done by lyophilization, dialysis followed by ion exchange chromatographies. Five grammes of animal feed samples (wheat flour, corn flour and soybean flour) were mixed with 0.1 M tris-HCl buffer (pH 7.0) and incubated at a temperature of 50 °C and 100 rpm with 20 U partially purified phytases from thermophilic fungus (S. thermophile) and bacterium (B. subtilis subsp. subtilis JJBS250) separately and in combination for 24 h. Samples were withdrawn after a definite time and analysed for estimating inorganic phosphorus, phytic acid, soluble proteins and reducing sugars as described by Singh and Satyanarayana [18]. The liberation of amino acids was determined by calorimetric method (http://www.eng.umd.edu/~nsw/ench485/lab3a.htm). Amount and liberation of protein was also confirmed by SDS-PAGE analysis. Substrate and enzyme controls were also analysed separately.
Results and Discussion
Process Optimization for Enhanced Phytase Production in Submerged Fermentation
To enhance the phytase production, it is very essential to screen the entire medium component using the Plackett–Burman design (Table 1). Marked variations from 10 to 7100 U/L phytase production were reported, showing the role of medium optimization for enhanced phytase production. A p value less than 0.05 indicated that the model terms are significant. From the Plackett–Burman design study, phytase production was significantly affected by sucrose, sodium phytate and Tween-80. Mittal et al. [19] found orange peel bran, sucrose and ammonium dihydrogen phosphate and pH affecting phytase production for Klebsiella sp. DB3, whereas in case of S. thermophile peptone, starch, sodium phytate and Tween-80 were identified as important variables by Singh and Satyanarayana [11]. Wheat bran, pH, FeSO4 and CaCl2 were the important variables affecting the phytase production by Nocardia sp. MB 36 [17].
The experimental details for the variables selected for phytase production using RSM is given in Table 3. Analysis of variance (ANOVA) was used for the analysis of experimental data that calculated the following equation for the high phyatse production under the optimized condition:
Where Y is enzyme response (U/L), and A, B, C, A2, B2, C2, AB, AC, BC, A,3 B3 and ABC are variables.
The value of coefficient of determination (R 2) for phytase production is 0.9973 revealing 99.73 % variation in the model. This value should be closer to one for predicting better response for phytase production. A strong relationship between the experimental and predicted values of phytase production was shown by high value (0.9919) of correlation coefficient, i.e. predicted R 2 The F value (183.99), adequate precision (43.578) and p value (<0.0001) obtained in the ANOVA analysis showed that the model is significant.
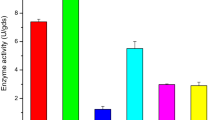
Response surface graphs displayed the interactions among the variables and their optimal levels for enhanced production of phytase by B. subtilis subsp. subtilis JJBS250 (Fig. 1a–c). Maximum phytase production of 7170 U/L was recorded with a predicted response of 7056.17 U/L, showing strong correlation between predicted and experimental responses. The optimum levels of variables were as sucrose 1.0 %, sodium phytate 0.4 % and Tween-80 2.0 %.

Response surface graph showing the effect and interaction of a sodium phytate and sucrose, b Tween-80 and sucrose and c Tween-80 and sodium phytate
Under optimal medium through the Placket-Burman design, B. subtilis produced 7170 U/L phytase, which is 6.79-fold higher than optimized medium by ‘one variable at a time’ approach (1056 U/L). Rani et al. [20] recorded a 7.95-fold enhancement in phytase production by using statistical approach in Rhizopus oryzae in comparison to unoptimized medium. Several studies have reported enhanced phytase production by different microorganisms like 1.3-fold by Aspergillus fumigatus NF171 [13], 5.25-fold by Klebsiella sp. DB3 [19], 3.73-fold by S. thermophile [11], 10.83-fold by Thermomyces lanuginosus [21], 17.5-fold by Hypocrea lixii SURT01 [22], 3.35-fold by Aspergillus oryzae [15] due to statistical optimization.
The phytase gene from Bacillus sp. was isolated and overexpressed in Escherichia coli resulted into 171.7 U/mL of recombinant phytase in batch cultivation. In the presence of inducers like lactose and isopropyl β-D-1-thiogalactopyranoside, recombinant phytase production reached up to 1183.57 and 5451 U/mL, respectively, in fed batch fermentation [23]. However, the level of recombinant phytase production from Bacillus phytase overexpressed in E. coli was resulted up to only 71.49 U/mL [24]. The phytase gene from Bacillus sp. expressed in Pichia pastoris resulted into 13.67 U/mL of recombinant phytase production with methanol induction in shake flask fermentation [25]. Similarly, the phytase gene from B. subtilis US417 was expressed in P. pastoris and recombinant strain produced 716.81 and 3784 U/mL in shake flask and high cell density fermentation, respectively [26].
Development of an Ideal Process for Dephytinization of Food Phytate Using Microbial Phytases
The phytic acid of food ingredients was hydrolyzed by microbial phytase efficiently at 50 °C that led to the liberation of inorganic phosphorus, reducing sugars, soluble protein and amino acids in a time-dependent manner (Table 4). Among all food samples, the liberation of inorganic phosphate was reported maximum in soybean flour (39.52 mg/g) followed by maize flour (31.94 mg/g) and wheat flour (16.58 mg/g) by combined effect of B. subtilis and S. thermophile phytases after an incubation period of 24 h. Similarly, concomitant liberation of soluble proteins, reducing sugars and amino acid liberation were reported high with cumulatively effect of both phytases (Table 4). Liberation of protein was confirmed by SDS-PAGE analysis which displayed the higher level of proteins released after a 24-h time interval in relative to control sample (0 h) [Fig. 2]. The content of phytic acid was also reduced significantly in all the treated flours (Data not shown). In comparison to Bacillus phytase, dephytinization of different flours was lower in case of S. thermophile phytase as it showed optimal activity at pH 5.0 [14], whereas Bacillus phytase showed optimal activity at pH 7.0 (Data not shown). Similarly, dephytinization of infant cereals by phytase from Bifidobacterium spp. resulted into increased levels of myo-inositol triphosphate and reduced the phytate contents [27], whereas sesame oil cake [18] and soymilk [28] were also hydrolyzed by the phytase of S. thermophile with concomitant liberation of inorganic phosphate, reducing sugars and soluble proteins. Phytase from Humicola nigrescens efficiently dephytinized the wheat and gramme flours with releases of soluble protein and inorganic phosphorus [29]. Sapna and Singh [30] reported the dephytinization of wheat bran with A. oryzae phytase. Sapna and Singh [15] also observed the hydrolysis of different flours (wheat, gramme and pearl millet) by the A. oryzae phytase at different temperatures (30 and 50 °C) and resulted into maximum dephytinization at 50 °C which is the optimal activity temperature of the phytase. Therefore, Microbial phytase treatment reduced the anti-nutritional factor, i.e. phytic acid with concomitant amelioration in nutritional components and thus making them bioavailable to the monogastric animals.

SDS-PAGE showing liberation of soluble proteins from food phytates treated by phytases from B. subtilis (lanes 1 and 2 after 0 and 24 h, respectively), both B. subtilis and S. thermophile (lanes 3 and 4 after 0 and 24 h, respectively) and S. thermophile (lanes 5 and 6 after 0 and 24 h, respectively). Arrow represents the prominent bands showing liberation of more proteins in comparison to control
Conclusions
Statistical approaches for the optimization of several parameters affecting phytase production by B. subtilis subsp. subtilis JJBS250 isolated from soil sample have been proved efficient and resulted in overall 6.79-fold higher production. An economical and ideal process was developed for dephytinization of food phytates that resulted in improvement of nutritional quality. Therefore, these microbial phytases could be suitable as additives in food and feed industries for ameliorating the nutrition and for reducing phytates in an eco-friendly and economical way.
References
Vohra, A., & Satyanarayana, T. (2003). Phytases: microbial sources, production, purification and potential biotechnological applications. Critical Reviews in Biotechnology, 23, 29–60.
Vats, P., & Banerjee, U. C. (2004). Production studies and catalytic properties of phytases (myo-inositolhexakisphosphate phosphohydrolases): an overview. Enzyme and Microbial Technology, 35, 3–14.
Singh, B., & Satyanarayana, T. (2011). Phytases from themophilic molds: their production, characteristics and multifarious applications. Process Biochemistry, 46, 1391–1398.
Singh, B., Kunze, G., & Satyanarayana, T. (2011). Developments in biochemical aspects and biotechnological applications of microbial phytases. Biotechnology and Molecular Biology Reviews, 6, 69–87.
Jinender, J., Sapna, & Singh, B. (2016). Characteristics and biotechnological applications of bacterial phytases. Process Biochemistry, 51, 159–169.
Singh, B., & Satyanarayana, T. (2015). Fungal phytases: characteristics and amelioration of nutritional quality and growth of non-ruminants. Journal of Animal Physiology and Animal Nutrition, 99, 646–660.
Vohra, A., & Satyanarayana, T. (2002). Statistical optimization of the medium components by response surface methodology to enhance phytase production by Pichia anomala. Process Biochemistry, 7, 999–1004.
Bogar, B., Szakacs, G., Pandey, A., Abdulhameed, S., Linden, J. C., & Tengerdy, R. P. (2003). Production of phytase by Mucor racemosus in solid-state fermentation. Biotechnology Progress, 19, 312–319.
Sonia, D., & Namita, S. R. (2009). Optimization of growth parameters of phytase producing fungus using RSM. Journal of Scientific and Industrial Research, 68, 955–959.
Singh, B., & Satyanarayana, T. (2006a). A marked enhancement in phytase production by a thermophilic mould Sporotrichum thermophile using statistical designs in a cost-effective cane molasses medium. Journal of Applied Microbiology, 101, 344–352.
Singh, B., & Satyanarayana, T. (2008a). Improved phytase production by a thermophilic mould Sporotrichum thermophile in submerged fermentation due to statistical optimization. Bioresource Technology, 99, 824–830.
Bajaj, B. K., & Wani, M. A. (2011). Enhanced phytase production from Nocardia sp. MB 36 using agro-residues as substrates: potential application for animal feed production. Engineering in Life Sciences, 6, 620–628.
Gangoliya, S. S., Gupta, R. K., & Singh, N. K. (2015). Phytase production through response surface methodology and molecular characterization of Aspergillus fumigatus NF191. Indian Journal of Experimental Biology, 53, 350–355.
Kumari, A., Satyanarayana, T., & Singh, B. (2015). Mixed substrate fermentation for enhanced phytase production by thermophilic mould Sporotrichum thermophile and its application in beneficiation of poultry feed. Applied Biochemistry and Biotechnology. doi:10.1007/s12010-015-1868-8.
Sapna, & Singh, B. (2015). Biocatalytic potential of protease-resistant phytase of Aspergillus oryzae SBS50 in ameliorating food nutrition. Biocatalysis and Biotransformation, 33, 167–174.
Fiske, C. H., & Subbarow, Y. P. (1925). The colorimetric determination of phosphorus. The Journal of Biological Chemistry, 66, 375–400.
Plackett, R. L., & Burman, J. P. (1946). The design of optimum multi-factor experiments. Biometrika, 33, 305–325.
Singh, B., & Satyanarayana, T. (2006b). Phytase production by thermophilic mold Sporotrichum thermophile in solid-state fermentation and its application in dephytinization of sesame oil cake. Applied Biochemistry and Biotechnology, 133, 239–250.
Mittal, A., Singh, G., Goyal, V., Yadav, A., & Aggarwal, N. K. (2011). Optimization of medium components for phytase production on orange peel flour by Klebsiella sp. DB3 using response surface methodology. Innovative Romanian Food Biotechnology, 9, 35–44.
Rani, R., Arora, S., Kumar, S., & Ghosh, S. (2013). Optimization of medium components for the production of phytase by R. oryzae using statistical approaches. Journal of Bioremediation & Biodegradation. doi:10.4172/2155-6199.S18-003.
Berikten, D., & Kivanc, M. (2014). Optimization of solid-state fermentation for phytase production by Thermomyces lanuginosus using response surface methodology. Preparative Biochemistry and Biotechnology, 44, 834–848.
Thyagarajan, R., Namasivayam, S. K. R., & Kumar, G. K. (2014). Optimization of medium components for phytase production by Hypocrea lixii SURT01 using response surface methodology. Journal of Pure and Applied Microbiology, 8, 2485–2490.
Tran, T. T., Mamo, G., Mattiasson, B., & Hatti-Kaul, R. (2010). A thermostable phytase from Bacillus sp. MD2: cloning, expression and high-level production in Escherichia coli. Journal of Industrial Microbiology and Biotechnology, 37, 279–287.
Rao, D. E. C. S., Rao, K. V., & Reddy, V. D. (2008). Cloning and expression of Bacillus phytase gene (phy) in Escherichia coli and recovery of active enzyme from the inclusion bodies. Journal of Applied Microbiology, 105, 1128–1137.
Guerrero-Olazaran, M., Rodriguez-Blanco, L., Carreon-Trevino, J. G., Gallegos-Lopez, J. A., & Viader-Salvado, J. M. (2010). Expression of a Bacillus phytase C gene in Pichia pastoris and properties of the recombinant enzyme. Applied and Environmental Microbiology, 76, 5601–5608.
Hmida-Sayari, A., Elgharbi, F., Farhat, A., Rekik, H., Blondeau, K., & Bejar, S. (2014). Overexpression and biochemical characterization of a thermostable phytase from Bacillus subtilis US417 in Pichia pastoris. Molecular Biotechnology, 56, 839–848.
Sanz-Penella, J. M., Frontela, C., Ros, G., Martinez, C., Monedero, V., & Haros, M. (2012). Application of bifidobacterial phytases in infant cereals: effect on phytate contents and mineral dialyzability. Journal of Agricultural and Food Chemistry, 60, 11787–11792.
Singh, B., & Satyanarayana, T. (2008b). Phytase production by a thermophilic mould Sporotrichum thermophile in solid state fermentation and its potential applications. Bioresource Technology, 99, 2824–2830.
Bala, A., Sapna, Jain, J., Kumari, A., & Singh, B. (2014). Production of an extracellular phytase from a thermophilic mould Humicola nigrescens in solid state fermentation and its application in dephytinization. Biocatalysis and Agricultural Biotechnology, 3, 259–264.
Sapna, & Singh, B. (2014). Phytase production by Aspergillus oryzae in solid-state fermentation and its applicability in dephytinization of wheat bran. Applied Biochemistry and Biotechnology, 173, 1885–1895.
Acknowledgments
Authors are highly thankful to the Department of Science and Technology (No. SR/FT/LS-95/2010), New Delhi, India for providing the financial assistance during this research work.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article

Cite this article
Jain, J., Singh, B. Phytase Production and Development of an Ideal Dephytinization Process for Amelioration of Food Nutrition Using Microbial Phytases. Appl Biochem Biotechnol 181, 1485–1495 (2017). https://doi.org/10.1007/s12010-016-2297-z
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s12010-016-2297-z