
Abstract
Epileptic encephalopathies account for a large proportion of the intractable early-onset epilepsies and are characterized by frequent seizures and poor developmental outcome. The epileptic encephalopathies can be loosely divided into two related groups of named syndromes. The first comprises epilepsies where continuous EEG changes directly result in cognitive and developmental dysfunction. The second includes patients where cognitive impairment is present at seizure onset and is due to the underlying etiology but the epileptic activity may then worsen the cognitive abilities over time. Recent, large-scale exome studies have begun to establish the genetic architecture of the epileptic encephalopathies, resulting in a re-consideration of the boundaries of these named syndromes. The emergence of this genetic architecture has lead to three main pathophysiological concepts to provide a mechanistic framework for these disorders. In this article, we will review the classic syndromes, the most significant genetic findings, and relate both to the pathophysiological understanding of epileptic encephalopathies.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Epilepsies are a common neurological disorder in infancy, childhood, and adolescence with a prevalence of approximately 0.5% [1]. Within the broad spectrum of childhood epilepsies, epileptic encephalopathies account for a large proportion of the severe, intractable early-onset seizure disorders, characterized by frequent and intractable seizures and poor developmental outcome. According to the International League Against Epilepsy (ILAE), epileptic encephalopathies are defined as conditions where “the epileptic activity itself may contribute to severe cognitive and behavioral impairments above and beyond what might be expected from the underlying pathology alone (e.g. cortical malformation), and that these can worsen over time” [2]. Developmental stagnation or regression is a key feature of epileptic encephalopathies. The underlying mechanisms of the seizures and cognitive disabilities are poorly understood, but the cognitive issues are believed to be a result of uncontrolled excitation of neuronal networks by ongoing abnormal EEG activity or the underlying etiology. The concept implied in this definition is that additional seizure activity worsens the clinical course of either an acquired (e.g., stroke, hypoxic-ischemic encephalopathy) or genetic cause (SCN1A, STXBP1) in the developing brain, which is presumed to be the underlying cause of the epilepsy. Thus, in many conditions, it is challenging to differentiate whether ongoing epileptic activity worsens cognitive dysfunction or whether progressive disturbance of brain function is due to the underlying etiology of the primary condition itself. This has led to some confusion with regard to the proper use of the concept of epileptic encephalopathy [3].
To accommodate both points of view, we suggest that epileptic encephalopathies can be divided into two groups. The first group is epilepsies where continuous EEG changes directly result in cognitive and developmental dysfunction such as epileptic encephalopathy with continuous spike-wave during slow-wave sleep, Landau-Kleffner syndrome, and childhood absence epilepsy with spike-wave stupor. These can be considered “true” or primary epileptic encephalopathies where epileptic activity directly leads to cognitive worsening and behavioral impairment in patients with previously normal development. The second group includes patients where cognitive impairment is present at seizure onset and cognitive issues are due to the underlying etiology, which alters proper neuronal network development. Epileptic activity may then worsen intellectual abilities over time and ultimately lead to a worse neurologic outcome. This group comprises a spectrum of age-related epileptic encephalopathies that have been defined by electro-clinical characteristics. These include Ohtahara syndrome, early myoclonic encephalopathy, and some patients with infantile spasms, Dravet syndrome, and Lennox-Gastaut syndrome.
The age-related electro-clinical syndromes do not match perfectly the clinical presentation of all children who present with a severe early-onset epileptic encephalopathy. Some of these clinical phenotypes can now be conceptualized as genetic entities (e.g., KCNQ2 encephalopathy) in which mutations in a gene correlate with certain phenotypic characteristics that, however, do not coincide with a specific electro-clinical syndrome. Recently, high-throughput sequencing methods such as whole-exome sequencing have uncovered an expanding number of causative genes for epileptic encephalopathies (see Table 1 for a list of the more prevalent genes). Large-scale exome studies have led to a massive gene discovery with over 100 genes identified to date. This new genetic information challenges the traditional concept of epileptic encephalopathies as patients can have clinical presentations that range from static to degenerative, blurring a clear distinction between primary epileptic encephalopathies or secondary encephalopathic aspects of neurodevelopmental disorders. In the current review, we will delineate the known clinical syndromes associated with epileptic encephalopathies and describe significant genetic findings linked to each electro-clinical syndrome. The emerging genetic architecture has led to a better understanding of the pathophysiology of epileptic encephalopathy syndromes, in particular related to ion channel disruption, synaptic transmission, and interneuron dysfunction which will then be briefly discussed to place the classic epileptic encephalopathies into a mechanistic and genetic framework.
Ohtahara Syndrome
Ohtahara syndrome or early infantile epileptic encephalopathy with suppression-burst pattern was first described in 1976. Ohtahara syndrome is an early-onset epileptic encephalopathy with clinical manifestation generally within first 3 months of life [4, 5]. Frequent clustering of tonic seizures or epileptic spasms is the clinical hallmark of Ohtahara syndrome and an interictal suppression-burst pattern is typical [4, 6]. The suppression-burst pattern consists of frequent high voltage bursts and almost flat suppression phases. Seizures are often resistant to antiepileptic treatment, and the developmental outcome is poor [4,5,6]. The etiology of Ohtahara syndrome is heterogeneous. Prior to the discovery of underlying genetic etiologies, structural brain lesions were identified as one of the most common underlying etiologies [6]. Treatment options are very limited as seizures are usually intractable; however, hormone treatment with ACTH can be effective in a small subset of patients [7].
With the advent of massively parallel sequencing techniques, growing evidence suggests that pathogenic mutations in three genes (STXBP1, KCNQ2, and SCN2A) account for a significant proportion of the non-lesional individuals with Ohtahara syndrome. Saitsu and collaborators were the first ones to identify mutations in STXBP1, encoding the syntaxin binding protein 1, in patients with Ohtahara syndrome [8]. Within the following years, the phenotypic spectrum of STXBP1 mutations expanded rapidly with increasing evidence for its role in Ohtahara syndrome [9,10,11,12,13,14]. Current mutation frequencies in STXBP1 account for 20% of individuals with Ohtahara syndrome [14].
Mutations in KCNQ2, encoding a potassium channel subunit, were originally reported in patients with benign familial neonatal seizures (BFNS) [15, 16]. More recent, mutations in KCNQ2 were found in up to 10% of patients with neonatal epileptic encephalopathy that share features with Ohtahara syndrome [17]. Interestingly, most of the genetic alterations associated with Ohtahara syndrome are de novo missense mutations whereas the BFNS mutations are deletions or loss of function mutations [15, 16, 18, 19]. In contrast to mutations in STXBP1, there is rarely a transition to other electro-clinical syndromes such as West syndrome [19]. Two thirds of individuals with pathogenic variants in KCNQ2 become seizure-free. Unfortunately, despite being seizure-free, nearly all patients with KCNQ2 encephalopathy have some degree of intellectual disability [19].
More recently, Nakamura and collaborators performed WES in patients with early-onset epileptic encephalopathy (EOEE) including 67 cases with Ohtahara syndrome [20]. The study revealed de novo SCN2A mutations in approximately 10% of the study participants [20]. Mutations in SCN2A were originally reported in patients with benign familial neonatal-infantile seizures (BFNIS) hinting towards an interesting parallel between KCNQ2 and SCN2A as both cause a benign neonatal seizure syndrome and an early-onset epileptic encephalopathy [21, 22].
Early Myoclonic Encephalopathy
In addition to Ohtahara syndrome, early myoclonic encephalopathy (EME) is a second electro-clinical epilepsy syndrome that manifests itself within the neonatal period or within the first 3 months of life [23, 24]. Erratic myoclonus is the most common seizure type, but partial motor seizures and tonic spasms can be found with evolution of the EME. Similar to Ohtahara syndrome, suppression-burst pattern is the characteristic EEG feature [6]. However, compared to Ohtahara syndrome, the suppression-burst pattern usually persists into childhood and appears to be more evident during sleep. Some patients show a transient period of hypsarrythmia in infancy [6]. Ohtahara and collaborators observed progressive diffuse brain atrophy in all of their patients which the authors interpreted as an underlying progressive disease process [6]. Generally, the long-term prognosis of early myoclonic encephalopathy is poor, and there are usually no effective treatment options.
The current genetic understanding of EME is limited. It has been suggested that EME is more frequently associated with metabolic disorders, even though a large proportion of patients do not have an identified etiology. Brain malformations appear to be less frequent compared to Ohtahara syndrome [6, 25]. Considering the metabolic impact on the pathogenesis of EME, the discovery of SLC25A22 as an underlying causative genetic etiology in several patients is intriguing [26]. SLC25A22 codes for a solute carrier implicated in glutamate transport into mitochondria. Furthermore, alterations in SLC25A22 may impact neuronal excitability. However, as with many other genes implicated in the etiology of EME, SLC25A22 only accounts for a small proportion of EME [26]. Recently, several novel genes including PIGA and SIK1 have been associated with EME [27, 28].
West Syndrome
West syndrome or infantile spasms syndrome was the first epilepsy syndrome that carried the basic notion of an epileptic encephalopathy when it was first described by the general practitioner W.J. West from Tunbridge, England, in his own son [29]. Dr. West already described many of the clinical features that would later give rise to the basic concept of an epileptic encephalopathy, namely developmental regression and epileptic spasms. West syndrome is characterized by an onset in the first year of life and is defined as the triad of hypsarrhythmia on EEG, infantile spasm seizures, and arrest of and/or regression of psychomotor development. Hormone treatment can initially sufficiently control spasms and improve developmental outcome in patients with no identified underlying etiology [30]. In patients with West syndrome resulting in an epileptic encephalopathy refractory to treatment, underlying etiologies can be identified in a significant subset of patients, which can be broken down into structural/metabolic and primary genetic etiologies. Within the group of patients with lesional etiologies, hypoxic-ischemic encephalopathy and tuberous sclerosis complex are amongst the most common causes [31].
The genetic etiologies for West syndrome are heterogeneous and probably account for 15–20% of individuals with this condition [32••]. Modern massive parallel sequencing technologies have uncovered a plethora of underlying genetic etiologies for West syndrome, which frequently overlap with other epileptic encephalopathies. SCN1A, SCN2A, STXBP1, and DNM1 are among the most frequently observed genes [32••, 33••]. Some genes, in contrast may be more specific to infantile spasms. For example, one of the most common genetic causes of infantile spasms are alterations in the CDKL5 gene coding the ubiquitously expressed cyclin-dependent kinase-like 5, also termed serine-threonine kinase 9 [34]. Prior to the transition to West syndrome, most female patients with CDKL5 mutations develop early-onset epilepsy with generalized tonic-clonic or focal seizures [35, 36] and some patients show stereotypical hand movements resembling Rett syndrome (i.e., Hanefeld variant of Rett syndrome) [37]. Another prominent West syndrome gene is ARX where children with and without brain malformations still develop infantile spasms, suggesting something unique about this gene leading to spasms [38,39,40].
Dravet Syndrome
One well-defined epileptic encephalopathy with a relatively homogeneous genetic architecture is Dravet syndrome, previously referred to as severe myoclonic epilepsy of infancy (SMEI). Classically, seizures in Dravet syndrome start in the first year of life, as unilateral clonic or generalized tonic-clonic febrile seizures that are often prolonged [41]. As Dravet syndrome progresses, myoclonic seizures, afebrile focal seizures, and atypical absences may occur. Status epilepticus, commonly encountered between 1 and 4 years, is usually clinically challenging [42]. Generalized spike-waves and polyspike waves are the most frequent EEG findings [42]. Patients with Dravet syndrome often have significant developmental delay and cognitive dysfunction with most patients developing unremarkably in their first year of life, before cognitive functions plateau or decline in the second year of life [41]. Despite the severity of the seizure, there seems to be no significant correlation between seizure type or frequency and the long-term cognitive outcome [43]. With the European Medicines Agency approval of Stiripentol for Dravet syndrome in the early 2000s, there now is an effective therapeutic option [7].
Mutations in SCN1A can be found in 70–80% of patients with Dravet syndrome [44, 45]. SCN1A mutations may even be more common, as a significant number of mutations may have been missed previously [46]. SCN1A encodes a neuronal voltage-gated sodium channel that is highly expressed in the brain and is involved in action potential initiation and neuronal excitability. SCN1A mutations are associated with other phenotypes such as generalized epilepsy with febrile seizures plus (GEFS+). There is a poor genotype-phenotype correlation with SCN1A variants, even though more severe phenotypes are associated with de novo truncation or deletion mutations resulting in haploinsufficiency [42, 44, 45, 47, 48]. Missense mutations, however, are the most common SCN1A mutation type seen in patients with Dravet syndrome [48]. Nevertheless, there is no mutational hotspot and well over 1000 different known mutations have been reported in SCN1A [49•]. Among patients with Dravet syndrome, negative for mutations in SCN1A, various other genes including PCDH19, GABRG2, CHD2, HCN1, STXBP1, and GABRA1 have been implicated [13,51,52,, 50–53]. Given the clinically distinctive Dravet syndrome phenotype, genetic testing for SCN1A is a useful diagnostic tool as it can alter treatment regimens and prevent a further prolonged diagnostic odyssey [54].
Lennox-Gastaut Syndrome
Lennox-Gastaut syndrome has onset in early childhood (3-6 years) and is characterized by the triad of tonic seizures, the EEG patterns of spike- and slow-wave activity, and generalized paroxysmal fast activity [55, 56]. Besides multiple other seizure types, there is significant cognitive comorbidity that may often only become prominent as the child ages. Different levels of cognitive impairment are present in LGS patients, and global developmental delay/intellectual disability is present at onset in most cases [57, 58]. Nonconvulsive status epilepticus (NCSE) is seen in 50–75% of patients with LGS, defined as persistent epileptiform EEG abnormalities with no apparent motor activity [59]. NCSE is frequently underdiagnosed and undertreated due to the lack of an observable motor component. However, NCSE presumably contributes significantly towards the cognitive impairment in LGS, even though it is often difficult to distinguish periods with excessive epileptiform activity from the baseline EEG features [59]. Seizures in LGS are often intractable and difficult to treat with antiepileptic medication. With respect to the role of LGS in the setting of other epilepsy syndromes, 40–60% of the cases evolve from West syndrome, while up to one third of patients develop LGS without a prior history of epilepsy [56, 60].
The etiology of LGS is heterogeneous. In addition to a lesional and metabolic etiology, pathogenic de novo variants in ALG13, CHD2, DNM1, GABRB3, GRIN2B, and SCN1 have been identified in a small subset of cases [32••, 33••, 61]. In fact, the proportion of patients without an identified etiology appears to be higher in the LGS group than in other epileptic encephalopathies, suggesting an area of need in genetic diagnostics. Further gene discovery studies are necessary to expand our understanding of the etiologies of LGS.
Childhood Absence Epilepsy with Spike-Wave Stupor
Genetic generalized epilepsies (GGE) account for about 20–30% of all epilepsies [62]. A subset of GGE patients can develop frequent episodes of nonconvulsive status epilepticus (NCSE) defined as ongoing epileptic activity longer than 30 min or lack of return to baseline during this period [63]. During and after NCSE, there is a dramatic change in alertness and the diagnosis of NCSE requires continuous generalized spike-waves on EEG [64]. Acute change in performance in a child with ongoing epilepsy may be an indicator for NCSE, and its transient nature is essential for the differentiation from epileptic encephalopathies [65]. NCSE evolves from regular absence attacks displaying the typical 3Hz spike-and-wave pattern and, thus, may be considered a prolonged absence seizure. Treatment for NCSE is oral or intravenous benzodiazepines, and generally no anesthetic coma is needed.
Many of the genes known to cause GGE were initially identified by systematic analysis of large families with mild dominant epilepsies prior to the advent of massive parallel sequencing. In 2001, Wallace et al. associated a mutation in GABRG2, encoding a GABAA receptor subunit, with childhood absence epilepsy (CAE) and febrile seizures [66]. This was the first disruption of an ion channel gene reported in patients with CAE. In the following years, the role of GABRG2 as candidate gene for GGE was confirmed and other ion channel genes such as CACNA1A and GABRA1 were identified [50,68,69,70,, 67–71]. No gene for sporadic GGE has been identified. Mutations in SLC2A1 coding for a glucose transporter at the blood-brain barrier have been found to be associated with early-onset absence epilepsy (EOAE) [72,73,74] but also other atypical GGE subtypes including myoclonic astatic epilepsy (MAE) [75, 76]. While the frequency of GLUT1 variants in GGE is assumed to be very low, identification of these rare cases is thought to be relevant as the ketogenic diet may offer a causal treatment choice for these patients [75].
Also, up to 3% of patients with generalized epilepsy are known to carry microdeletions including microdeletions at 15q13.3, 16p13.11, and 15q11.2 [77,78,79,80]. While these microdeletions may also be found in unaffected carriers, they are significantly associated with GGE and some microdeletions such as 15q13.3 and 16p13.11 may be particularly be prominent in patients with atypical GGE subtypes that often have features of epileptic encephalopathies [80, 81].
Epilepsy-Aphasia Syndromes
Landau-Kleffner syndrome (LKS) and epileptic encephalopathy with continuous spike-wave during slow-wave sleep (CSWS) are two closely related epileptic encephalopathies with onset in childhood. They represent the severe end of a spectrum of focal epilepsy syndromes including rolandic epilepsy among others that can be conceptualized as epilepsy-aphasia syndromes (EAS). LKS and CSWS share similar clinical and EEG features as seizures, developmental and language regression, and interictal paroxysmal EEG abnormalities increased during sleep. The characteristic EEG findings are centro-temporal sharp-slow wave complexes that can be bilaterally independent or bi-synchronous. When the epileptiform activity becomes constant during sleep, different authors have used the term electrical status epilepticus during sleep (ESES) or CSWS [82, 83]. The spike-and-wave pattern during non-REM sleep is typical for CSWS. Landau-Kleffner syndrome or acquired epileptic aphasia is characterized by several different seizure types, early regression of language, and bi-temporal but predominantly frontotemporal spike-wave pattern during non-REM sleep [84]. Treatment-wise, there is no established consensus on the ideal treatment as options are limited.
Recent genetic findings suggest a genetic etiology in a small proportion of patients with epilepsy-aphasia syndromes. GRIN2A, encoding a subunit of the NMDA receptor, has emerged most prominently as an underlying cause of this group of conditions [85••, 86••, 87••]. The NMDA receptor is a neurotransmitter-gated ion channel involved in glutamatergic transmission in the central nervous system. Mutations in GRIN2A have been implicated in various types of focal epilepsy syndromes including rolandic epilepsy, atypical benign partial epilepsy (ABPE), LKS, and CSWS accounting for up to 7.5% of patients [85••].
Shared Mechanisms of the Epileptic Encephalopathies
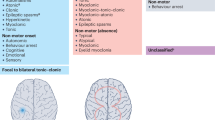
Epilepsy is caused by malfunction in hypersynchronous discharges within local or diffuse neuronal networks. The current literature has not been able to fully explain the underlying mechanisms of the epileptic encephalopathies and mechanisms leading to the common electroencephalographic patterns including suppression-burst, hypsarrythmia, and continuous spike-wave activity during slow-wave sleep. The emerging genetic etiologies are starting to provide a framework for the underlying mechanisms (see Table 1 and Fig. 1). While the current pathophysiological understanding is still very limited, three main pathophysiological mechanistic concepts have emerged throughout the past decade. These include ion channels dysfunction, deficits in synaptic transmission, and impairment in interneuron development (see Fig. 1 for an illustration of these issues).

Genes and mechanisms implicated in epileptic encephalopathies. Examples of established genes for epileptic encephalopathies that can disrupt synaptic transmission, increase or decrease neuron excitability, and influence cellular homeostasis and metabolism
Channelopathies
The channelopathy concept has been applied to multiple neurological and cardiac disorders and dates back to the work of Louis Ptáček on hyperkalemic periodic paralysis [88, 89]. Ion channels control the influx and efflux of ions and are considered the main regulator of cellular electrophysiological homeostasis (Fig. 1). Several genes coding for ion channels have been implicated in the etiology of the epileptic encephalopathies. Dravet syndrome due to pathogenic variants in SCN1A is an archetypal epileptic encephalopathy with a predominant genetic cause. The mechanisms that link SCN1A to epilepsy are poorly understood. Disruptions of an ion channel important for neuron excitability would conceptually result in decreased excitability. Thus, the loss of SCN1A, an excitatory sodium channel, leading to increased excitability seems counterintuitive. Yu et al. suggested a failure of excitability of hippocampal GABAergic inhibitory neurons as possible pathophysiological mechanism in a Scn1a-mouse model that may be sufficient to cause in epilepsy patients with Dravet syndrome [90]. Interestingly, the sodium current in excitatory pyramidal cells, however, seemed unchanged, and the loss of SCN1A is compensated by SCN3A in pyramidal cells [90, 91] but not in interneurons resulting in hyperexcitability. More recently, this theory is questioned by observations in human-induced pluripotent stem cells (iPSCs), where increased sodium currents in bipolar- and pyramidal-shaped neurons and evidence for hyperexcitability were identified [92]. In conclusion, a failure of inhibition in neuronal networks, as a mechanism linking ion channel dysfunction to epilepsy, has been suggested as a key mechanism in the pathogenesis of epilepsy disrupting the equilibrium of excitation and inhibition in the brain. Nonetheless, further work is needed to understand the mechanisms behind genetic alterations in genes coding for ion channels and human epilepsy.
Synaptic Transmission Dysfunction
Several different genes disrupting synaptic transmission have been implicated in human epilepsy (Fig. 1). STXBP1 is the predominant gene and was first identified in Ohtahara syndrome in 2008 [8]. The protein encoded by the STXBP1 gene is involved in vesicular neurotransmitter secretion and presynaptic vesicular docking and plays a regulatory role in synaptic exocytosis [93,94,95,96]. STXBP1 is an interesting example how disorders of synaptic transmission require a different approach to unravel the underlying pathophysiology of epileptic encephalopathies. Another well-established gene for epileptic encephalopathies is DNM1 [32••, 33••]. Functional studies on DNM1, a gene involved in synaptic endocytosis and neurotransmitter release, pointed towards inhibitory neuronal dysfunction as an underlying mechanism for seizures [97]. This hypothesis is supported by the observation that endocytotic defects in primary neuron cultures are activity dependent and that inhibitory neurons are uniquely prone to perturbation of endocytic proteins [98]. In addition, it has been shown in vitro that identified DNM1 mutations inhibit endocytosis, indicating that pathogenic variants in patients result in functionally relevant changes [99•]. As with STXBP1, the conceptual understanding of DNM1-related epileptic encephalopathies as disorders of synaptic transmission is still in its infancy requires further studies.
Interneuronopathy
In 2005, Kato and Dobyns introduced the term “interneuronopathy” for developmental disorders that are accompanied by disrupted migration and loss of GABAergic interneurons [100]. The concept is based on the observation that both radial and non-radial migration are crucial for brain development [101,102,103]. ARX-related X-linked lissencephaly with abnormal genitalia (XLAG) was the prototypic disorder for this concept, as impaired tangential migration and a loss of interneurons were described [102, 104]. ARX consists of a paired-class homeodomain and four polyalanine repeats and functions as a transcription factor that regulates brain development and maturation mainly as a transcriptional repressor [105,106,107]. A reliable genotype-phenotype correlation has been suggested for mutations in ARX depending on their location within the gene. While mutations in the highly conserved regions of the ARX gene account for severe phenotypes including brain malformations, expansions in the second polyalanine tract have been associated with non-malformation phenotypes [40, 108, 109]. Within the non-malformation phenotypes, intellectual disability and epilepsy are the predominant clinical findings. The length of polyalanine tract expansions influences the degree of transcriptional repression, and defects in interneuron development have been associated with polyalanine tract expansions [105, 110]. Furthermore, Arx knockout in mice leads to a malfunction in maturation and migration of GABAergic in the embryonic and the neocortex [102]. This is particularly relevant as the embryonic ganglionic eminence is an interneuron reservoir in the developing brain with a high degree of ARX expression [101]. The general concept underlying the pathogenesis in ARX-related disorders suggests that interneurons loss and abnormal segregation due to disruption in tangential migration results in epilepsy [100]. In the postnatal brain, ARX is exclusively expressed in GABAergic interneurons. However, the role of interneuronal ARX function in the post-natal brain remains elusive. It has been hypothesized that postnatal ARX expression may at least partially account for some of the clinical features seen in patients with epileptic encephalopathies.
Conclusions
In this review, we have discussed the clinical features of the most prominent epileptic encephalopathies and have attempted to contrast the known clinical presentations with the expanding genetic data that exists in the field. The concepts of channelopathies, synaptic transmission defects, and interneuronopathies likely cover only a subset within the pathogenesis of epileptic encephalopathies, but they provide a useful scaffold to explain some of the relevant mechanisms.
Within the past 15 years, the genetic findings and the molecular understanding have had a dramatic impact on patient care. It has become clear that for a subset of epileptic encephalopathies genetic testing can be beneficial. Positive genetics testing has multiple benefits including providing closure for families, providing counseling for reproductive decision-making, and ultimately to guide treatment (i.e., to not use sodium channel blocking drugs for SCN1A mutations) and provide a prognosis [111]. A general approach for the epileptic encephalopathies is to start with a gene panel, which has been shown to be a suitable diagnostic tool to screen for known epilepsy genes [112]. As de novo mutations are identified more often and mosaicism is becoming increasingly important, many patients will profit from diagnostic trio exome sequencing. Dravet syndrome is a good example on how early diagnostic genetic testing can improve therapeutic decisions by avoiding unnecessary aggravation due to sodium channel blockers, which will ultimately result in better patient care. However, lack of reliable correlation for some genes such as STXBP1 makes interpretation of genetic data challenging [14]. With the era of large-scale genomic studies, clinicians and researchers have come one step closer to offering precision medicine to patients. Thus, we suggest interdisciplinary efforts between medical geneticists, genetic counselors, basic researches, and child neurologists in the care of patients with epileptic encephalopathies.
References
Papers of particular interest, published recently, have been highlighted as: • Of importance •• Of major importance
Waaler PE, Blom BH, Skeidsvoll H, Mykletun A. Prevalence, classification, and severity of epilepsy in children in western Norway. Epilepsia. 2000;41:802–10.
Berg AT et al. Revised terminology and concepts for organization of seizures and epilepsies: report of the ILAE Commission on Classification and Terminology, 2005-2009. Epilepsia. 2010;51:676–85.
Howell KB, Harvey AS, Archer JS. Epileptic encephalopathy: use and misuse of a clinically and conceptually important concept. Epilepsia. 2016;57:343–7.
Ohtahara S, Ohtsuka Y, Oka E. Epileptic encephalopathies in early infancy. Indian J Pediatr. 1997;64:603–12.
Yamatogi Y, Ohtahara S. Early-infantile epileptic encephalopathy with suppression-bursts, Ohtahara syndrome; its overview referring to our 16 cases. Brain Dev. 2002;24:13–23.
Ohtahara S, Yamatogi Y. Ohtahara syndrome: with special reference to its developmental aspects for differentiating from early myoclonic encephalopathy. Epilepsy Res. 2006;70:2–5.
Wilmshurst JM et al. Summary of recommendations for the management of infantile seizures: Task Force Report for the ILAE Commission of Pediatrics. Epilepsia. 2015;56:1185–97.
Saitsu H et al. De novo mutations in the gene encoding STXBP1 (MUNC18-1) cause early infantile epileptic encephalopathy. Nat Genet. 2008;40:782–8.
Deprez L et al. Clinical spectrum of early-onset epileptic encephalopathies associated with STXBP1 mutations. Neurology. 2010;75:1159–65.
Saitsu H et al. STXBP1 mutations in early infantile epileptic encephalopathy with suppression-burst pattern. Epilepsia. 2010;51:2397–405.
Mignot C et al. STXBP1-related encephalopathy presenting as infantile spasms and generalized tremor in three patients. Epilepsia. 2011;52:1820–7.
Milh M et al. Epileptic and nonepileptic features in patients with early onset epileptic encephalopathy and STXBP1 mutations. Epilepsia. 2011;52:1828–34.
Carvill GL et al. GABRA1 and STXBP1: novel genetic causes of Dravet syndrome. Neurology. 2014;82:1245–53.
Stamberger H et al. STXBP1 encephalopathy A neurodevelopmental disorder including epilepsy. Neurology. 2016;86:954–62.
Biervert C. A potassium channel mutation in neonatal human epilepsy. Science. 1998;279:403–6.
Singh N et al. A novel potassium channel gene, KCNQ2, is mutated in an inherited epilepsy of newborns. Nat Genet. 1998;18:231–6.
Weckhuysen S et al. Extending the KCNQ2 encephalopathy spectrum: clinical and neuroimaging findings in 17 patients. Neurology. 2013;81:1697–703.
Saitsu H et al. Whole exome sequencing identifies KCNQ2 mutations in Ohtahara syndrome. Ann Neurol. 2012;72:298.
Kato M et al. Clinical spectrum of early onset epileptic encephalopathies caused by KCNQ2 mutation. Epilepsia. 2013;54:1282–7.
Nakamura K et al. Clinical spectrum of SCN2A mutations expanding to Ohtahara syndrome. Neurology. 2013;81:992–8.
Heron SE et al. Sodium-channel defects in benign familial neonatal-infantile seizures. Lancet. 2002;360:851–2.
Berkovic SF et al. Benign familial neonatal-infantile seizures: characterization of a new sodium channelopathy. Ann Neurol. 2004;55:550–7.
Aicardi J, Goutieres F. [Neonatal myoclonic encephalopathy (author’s transl)]. Rev Electroencephalogr Neurophysiol Clin. 1978;8:99–101.
Dalla Bernardina B et al. Early myoclonic epileptic encephalopathy (E.M.E.E.). Eur J Pediatr. 1983;140:248–52.
Ohtahara S, Yamatogi Y. Epileptic encephalopathies in early infancy with suppression-burst. J Clin Neurophysiol. 2003;20:398–407.
Molinari F et al. Impaired mitochondrial glutamate transport in autosomal recessive neonatal myoclonic epilepsy. Am J Hum Genet. 2005;76:334–9.
Kato M et al. PIGA mutations cause early-onset epileptic encephalopathies and distinctive features. Neurology. 2014;82:1587–96.
Hansen J et al. De novo mutations in SIK1 cause a spectrum of developmental epilepsies. Am J Hum Genet. 2015;96:682–90.
Eling P, Renier WO, Pomper J, Baram TZ. The mystery of the Doctor’s son, or the riddle of West syndrome. Neurology. 2002;58:953–5.
Lux AL et al. The United Kingdom Infantile Spasms Study (UKISS) comparing hormone treatment with vigabatrin on developmental and epilepsy outcomes to age 14 months: a multicentre randomised trial. Lancet Neurol. 2005;4:712–7.
Osborne JP et al. The underlying etiology of infantile spasms (West syndrome): information from the United Kingdom Infantile Spasms Study (UKISS) on contemporary causes and their classification. Epilepsia. 2010;51:2168–74.
•• Allen AS et al. De novo mutations in epileptic encephalopathies. Nature. 2013;501:217–21. Collaborative large-scale exome study revealing the signicance of de novo mutations in epileptic encephalopathies.
•• EuroEPINOMICS-RES Consortium, E. P. & Genome Project, E. C. De novo mutations in synaptic transmission genes including DNM1 cause epileptic encephalopathies. Am J Hum Genet. 2014;95:360–70. Collaborative large-scale exome study revealing the signicance of de novo mutations in epileptic encephalopathies.
Kalscheuer VM et al. Disruption of the serine/threonine kinase 9 gene causes severe X-linked infantile spasms and mental retardation. Am J Hum Genet. 2003;72:1401–11.
Klein KM et al. A distinctive seizure type in patients with CDKL5 mutations: hypermotor-tonic-spasms sequence. Neurology. 2011;76:1436–8.
Jähn J et al. CDKL5 mutations as a cause of severe epilepsy in infancy: clinical and electroencephalographic long-term course in 4 patients. J Child Neurol. 2013;28:937–41.
Evans JC et al. Early onset seizures and Rett-like features associated with mutations in CDKL5. Eur J Hum Genet. 2005;13:1113–20.
Strømme P et al. Mutations in the human ortholog of Aristaless cause X-linked mental retardation and epilepsy. Nat Genet. 2002;30:441–5.
Kato M, Das S, Petras K, Sawaishi Y, Dobyns WB. Polyalanine expansion of ARX associated with cryptogenic West syndrome. Neurology. 2003;61:267–76.
Kato M et al. Mutations of ARX are associated with striking pleiotropy and consistent genotype-phenotype correlation. Hum Mutat. 2004;23:147–59.
Dravet C. The core Dravet syndrome phenotype. Epilepsia. 2011;52:3–9.
Bender AC, Morse RP, Scott RC, Holmes GL, Lenck-Santini PP. SCN1A mutations in Dravet syndrome: impact of interneuron dysfunction on neural networks and cognitive outcome. Epilepsy Behav. 2012;23:177–86.
Akiyama M, Kobayashi K, Yoshinaga H, Ohtsuka Y. A long-term follow-up study of Dravet syndrome up to adulthood. 2010;51:1043–52.
Claes L et al. De novo mutations in the sodium-channel gene SCN1A cause severe myoclonic epilepsy of infancy. Am J Hum Genet. 2001;68:1327–32.
Sugawara T et al. Frequent mutations of SCN1A in severe myoclonic epilepsy in infancy. Neurology. 2002;58:1122–4.
Djémié T et al. Pitfalls in genetic testing: the story of missed SCN1A mutations. Mol Genet Genomic Med. 2016;1–8. doi:10.1002/mgg3.217.
Mulley JC et al. SCN1A mutations and epilepsy. Hum Mutat. 2005;25:535–42.
Depienne C et al. Spectrum of SCN1A gene mutations associated with Dravet syndrome: analysis of 333 patients. J Med Genet. 2009;46:183–91.
• Meng H et al. The SCN1A mutation database: updating information and analysis of the relationships among genotype, functional alteration, and phenotype. Hum Mutat. 2015;36:573–80. This is the most up-to-date listing and consideration of mutations and phenotypes in SCN1A related epilepsy, which is the most common genetic casue of the EEs.
Harkin LA et al. Truncation of the GABA A-receptor g2 subunit in a family with generalized epilepsy with febrile seizures plus. 2002;530–6.
Depienne C et al. Sporadic infantile epileptic encephalopathy caused by mutations in PCDH19 resembles Dravet syndrome but mainly affects females. PLoS Genet. 2009;5:e1000381.
Suls A et al. De novo loss-of-function mutations in CHD2 cause a fever-sensitive myoclonic epileptic encephalopathy sharing features with Dravet syndrome. Am J Hum Genet. 2013;93:967–75.
Nava C et al. De novo mutations in HCN1 cause early infantile epileptic encephalopathy. Nat Genet. 2014;46:640–5.
Hirose S et al. SCN1A testing for epilepsy: application in clinical practice. Epilepsia. 2013;54:946–52.
Gastaut H et al. Childhood epileptic encephalopathy with diffuse slow spike-waves (otherwise known as ‘petit-mal variant’) or Lennox syndrome. Epilepsia. 1966;7:85–138.
Arzimanoglou A et al. Lennox-Gastaut syndrome: a consensus approach on diagnosis, assessment, management, and trial methodology. Lancet Neurol. 2009;8:82–93.
Kaminska A et al. Delineation of cryptogenic Lennox-Gastaut syndrome and myoclonic astatic epilepsy using multiple correspondence analysis. Epilepsy Res. 1999;36:15–29.
Filippini M, Boni A, Dazzani G, Guerra A, Gobbi G. Neuropsychological findings: myoclonic astatic epilepsy (MAE) and Lennox-Gastaut syndrome (LGS). Epilepsia. 2006;47:56–9.
Hoffmann-Riem M et al. Nonconvulsive status epilepticus—a possible cause of mental retardation in patients with Lennox-Gastaut syndrome. Neuropediatrics. 2000;31:169–74.
Zupanc ML. Clinical evaluation and diagnosis of severe epilepsy syndromes of early childhood. J Child Neurol. 2009;24:6S–14S.
Lund C, Brodtkorb E, Øye AM, Røsby O, Selmer KK. CHD2 mutations in Lennox-Gastaut syndrome. Epilepsy Behav. 2014;33:18–21.
Jallon P, Loiseau P. Newly diagnosed unprovoked epileptic seizures: presentation at diagnosis in CAROLE Study. 2001;42:464–75.
DeLorenzo RJ et al. A prospective, population-based epidemiologic study of status epilepticus in Richmond, Virginia. Neurology. 1996;46:1029–35.
Wasterlain CG, Fujikawa DG, Penix L, Sankar R. Pathophysiological mechanisms of brain damage from status epilepticus. Epilepsia. 1993;34 Suppl 1:S37–53.
Walker MC. Diagnosis and treatment of nonconvulsive status epilepticus. CNS Drugs. 2001;15:931–9.
Wallace RH et al. Mutant GABA(A) receptor gamma2-subunit in childhood absence epilepsy and febrile seizures. Nat Genet. 2001;28:49–52.
Jouvenceau A et al. Early report human epilepsy associated with dysfunction of the brain P/Q-type calcium channel. Lancet. 2001;358:801–7.
Cossette P et al. Mutation of GABRA1 in an autosomal dominant form of juvenile myoclonic epilepsy. Nat Genet. 2002;31:184–9.
Lu J et al. Mutation screen of the GABAA receptor gamma 2 subunit gene in Chinese patients with childhood absence epilepsy. Neurosci Lett. 2002;332:75–8.
Imbrici P et al. Dysfunction of the brain calcium channel Cav2.1 in absence epilepsy and episodic ataxia. Brain. 2004;127:2682–92.
Maljevic S et al. A mutation in the GABAA receptor α1-subunit is associated with absence epilepsy. Ann Neurol. 2006;59:983–7.
Suls A et al. Early-onset absence epilepsy caused by mutations in the glucose transporter GLUT1. Ann Neurol. 2009;66:415–9.
Arsov T et al. Early onset absence epilepsy: 1 in 10 cases is caused by GLUT1 deficiency. Epilepsia. 2012;53:e204–7.
Muhle H et al. The role of SLC2A1 in early onset and childhood absence epilepsies. Epilepsy Res. 2013;105:229–33.
Mullen SA et al. Glucose transporter 1 deficiency as a treatable cause of myoclonic astatic epilepsy. Arch Neurol. 2011;68:1152–5.
Larsen J et al. The role of SLC2A1 mutations in myoclonic astatic epilepsy and absence epilepsy, and the estimated frequency of GLUT1 deficiency syndrome. Epilepsia. 2015;56:e203–8.
Helbig I et al. 15Q13.3 microdeletions increase risk of idiopathic generalized epilepsy. Nat Genet. 2009;41:160–2.
Dibbens LM et al. Familial and sporadic 15q13.3 microdeletions in idiopathic generalized epilepsy: precedent for disorders with complex inheritance. Hum Mol Genet. 2009;18:3626–31.
de Kovel CGF et al. Recurrent microdeletions at 15q11.2 and 16p13.11 predispose to idiopathic generalized epilepsies. Brain. 2010;133:23–32.
Mullen SA et al. Copy number variants are frequent in genetic generalized epilepsy with intellectual disability. Neurology. 2013;81:1507–14.
Helbig I et al. Structural genomic variation in childhood epilepsies with complex phenotypes. Eur J Hum Genet. 2014;22:896–901.
Patry G, Lyagoubi S, Tassinari CA. Subclinical ‘electrical status epilepticus’ induced by sleep in children. Arch Neurol. 1971;24:242–52.
Doose H, Neubauer B, Carlson G. Children with benign focal sharp waves. Neuropdatrics. 1996;227–41.
Landau WM, Kleffner FR. Syndrome of acquired aphasia with convulsive disorder in children. Neurology. 1957;7:8–1241.
•• Lemke JR et al. Mutations in GRIN2A cause idiopathic focal epilepsy with rolandic spikes. Nat Genet. 2013;45:1067–72. One of three studies published in the same issue of Nature Genetics highlighting the importance of mutations in GRIN2A as gene for epilepsy-aphasia syndromes.
•• Carvill GL et al. GRIN2A mutations cause epilepsy-aphasia spectrum disorders. Nat Genet. 2013;45:1073–6. One of three studies published in the same issue of Nature Genetics highlighting the importance of mutations in GRIN2A as gene for epilepsy-aphasia syndromes.
•• Lesca G et al. GRIN2A mutations in acquired epileptic aphasia and related childhood focal epilepsies and encephalopathies with speech and language dysfunction. Nat Genet. 2013;45:1061–6. One of three studies published in the same issue of Nature Genetics highlighting the importance of mutations in GRIN2A as gene for epilepsy-aphasia syndromes.
Ptáček LJ et al. Identification of a mutation in the gene causing hyperkalemic periodic paralysis. Cell. 1991;67:1021–7.
Ptáček LJ. Channelopathies: ion channel disorders of muscle as a paradigm for paroxysmal disorders of the nervous system. Neuromuscul Disord. 1997;7:250–5.
Yu FH et al. Reduced sodium current in GABAergic interneurons in a mouse model of severe myoclonic epilepsy in infancy. Nat Neurosci. 2006;9:1142–9.
Catterall WA, Kalume F, Oakley JC. NaV1.1 channels and epilepsy. J Physiol. 2010;588:1849–59.
Liu Y et al. Dravet syndrome patient-derived neurons suggest a novel epilepsy mechanism. Ann Neurol. 2013;74:128–39.
Swanson DA, Steel JM, Valle D. Identification and characterization of the human ortholog of rat STXBP1, a protein implicated in vesicle trafficking and neurotransmitter release. Genomics. 1998;48:373–6.
Verhage M et al. Synaptic assembly of the brain in the absence of neurotransmitter secretion. 2000;287:864–9.
Weimer RM et al. Defects in synaptic vesicle docking in unc-18 mutants. Nat Neurosci. 2003;6:1023–30.
Chen Y et al. Association between genetic variation of CACNA1H and childhood absence epilepsy. Ann Neurol. 2003;54:239–43.
Ferguson SM et al. A selective activity-dependent requirement for dynamin 1 in synaptic vesicle endocytosis. Science. 2007;316:570–4.
Hayashi et al. Cell- and stimulus-dependent heterogeneity of synaptic vesicle endocytic recycling mechanisms revealed by studies of dynamin 1-null neurons. Proc Natl Acad Sci U S A. 2008;105:2175–80.
• Dhindsa RS et al. Epileptic encephalopathy-causing mutations in DNM1 impair synaptic vesicle endocytosis. Neurol Genet. 2015;1:1–9. This is a nice recent example of how to determine the underlying pathophysiological deficiencies in specific genetic EEs.
Kato M, Dobyns WB. X-linked lissencephaly with abnormal genitalia as a tangential migration disorder causing intractable epilepsy: proposal for a new term, ‘interneuronopathy’. J Child Neurol. 2005;20:392–7.
Anderson SA, Eisenstat DD, Shi L, Rubenstein JL. Interneuron migration from basal forebrain to neocortex: dependence on Dlx genes. Science. 1997;278:474–6.
Kitamura K et al. Mutation of ARX causes abnormal development of forebrain and testes in mice and X-linked lissencephaly with abnormal genitalia in humans. Nat Genet. 2002;32:359–69.
Kato M, Dobyns WB. Lissencephaly and the molecular basis of neuronal migration. Hum Mol Genet. 2003;12 Spec No:R89–96.
Bonneau D et al. X-linked lissencephaly with absent corpus callosum and ambiguous genitalia (XLAG): clinical, magnetic resonance imaging, and neuropathological findings. Ann Neurol. 2002;51:340–9.
McKenzie O et al. Aristaless-related homeobox gene, the gene responsible for West syndrome and related disorders, is a Groucho/transducin-like enhancer of split dependent transcriptional repressor. Neuroscience. 2007;146:236–47.
Fulp CT et al. Identification of Arx transcriptional targets in the developing basal forebrain. Hum Mol Genet. 2008;17:3740–60.
Colasante G et al. ARX regulates cortical intermediate progenitor cell expansion and upper layer neuron formation through repression of Cdkn1c. Cereb Cortex. 2015;25:322–35.
Sherr EH. The ARX story (epilepsy, mental retardation, autism, and cerebral malformations): one gene leads to many phenotypes. Curr Opin Pediatr. 2003;15:567–71.
Friocourt. Mutations in ARX result in several defects involving GABAergic neurons. Front Cell Neurosci. 2010;4:1–11.
Nasrallah MP et al. Differential effects of a polyalanine tract expansion in Arx on neural development and gene expression. Hum Mol Genet. 2012;21:1090–8.
Helbig KL et al. Genetic risk perception and reproductive decision making among people with epilepsy. Epilepsia. 2010;51:1874–7.
Lemke JR et al. Targeted next generation sequencing as a diagnostic tool in epileptic disorders. Epilepsia. 2012;53:1387–98.
Acknowledgements
Markus von Deimling was supported by funds of the Stiftung zur Förderung der medizinischen Forschung of the University of Kiel and by a grant from the German Research Foundation (DFG, HE5415/5-1, HE5415/6-1). Ingo Helbig was supported by intramural funds of the University of Kiel, by a grant from the German Research Foundation (HE5415/3-1) within the EuroEPINOMICS framework of the European Science Foundation, and additional grants of the German Research Foundation (DFG, HE5415/5-1, HE 5415/6-1), German Ministry for Education and Research (01DH12033, MAR 10/012), and grant by the German chapter of the International League against Epilepsy (DGfE).
Eric D. Marsh was supported by NIH/NINDS - R01 NS082761-01. Dr. Marsh has also received a grant from GW Pharma, advisory board fees from Stanley Brothers Social Enterprises, and support from Neuren Pharmaceuticals.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of Interest
None of the authors have anything to declare related to this review
Human and Animal Rights and Informed Consent
This article does not contain any studies with human or animal subjects performed by any of the authors.
Additional information
This article is part of the Topical Collection on Pediatric Neurology
Rights and permissions
About this article

Cite this article
von Deimling, M., Helbig, I. & Marsh, E.D. Epileptic Encephalopathies—Clinical Syndromes and Pathophysiological Concepts. Curr Neurol Neurosci Rep 17, 10 (2017). https://doi.org/10.1007/s11910-017-0720-7
Published:
DOI: https://doi.org/10.1007/s11910-017-0720-7