
Abstract
Polymorphisms in plants are main factors that determine the diversity of associated animal communities and their population dynamics. Typically, Persicaria lapathifolia var. lapathifolia (Polygonaceae) has no trichomes on leaf surfaces (glabrous type), but a hairy type does sometimes occur. Based on a cultivation experiment, the presence or absence of trichomes is clarified to be under genetic control. To reveal the defensive function of trichomes against herbivores, laboratory experiments were conducted using a major herbivore, Galerucella grisescens (Coleoptera: Chrysomelidae). In both choice and no-choice feeding tests, the glabrous type was significantly more consumed by G. grisescens adults, while the hairy type was not consumed. In the hairy leaf treatment, larval duration tended to become longer, the adult body weight became significantly lower, and adults laid significantly more eggs than in the glabrous leaf treatment. Hairy leaves contained significantly more total phenolics and condensed tannins than glabrous leaves, suggesting that the hairy type allocates more resources for physical and chemical defence. Because no significant differences in leaf consumption were detected in the feeding experiment using powdered host leaves, G. grisescens seems to have adapted to the chemical defences of P. lapathifolia var. lapathifolia. These results clearly indicate that leaf trichomes of P. lapathifolia var. lapathifolia effectively act as a physical defence against G. grisescens.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Plants are consumed by many different herbivores; thus, they have various anti-herbivore defences. Direct resistance includes physical defences, such as trichomes and/or leaf toughness, and chemical defences, such as alkaloids and tannins (Howe and Westley 1988; Walters 2011). Trichomes, typically occurring on stems and leaves, have various functions, such as the prevention of photoinhibition (Benzing 1976), protection against desiccation (Hess et al. 1974), and physical defence against herbivores (e.g. Zvereva et al. 1998; Reymond et al. 2004; Handley et al. 2005; Sletvold et al. 2010; Yamawo et al. 2012; Sato and Kudoh 2015). Glandular trichomes may evoke toxic and disruptive effects against various herbivores (Kruidhof et al. 2012). Non-glandular trichomes frequently increase plant resistance to insect damage by providing an effective barrier that prevents small insects, such as young instar larvae, from moving and contacting the plant surface (Southwood 1986). The effects depend largely on the size of herbivores (Agrawal and Spiller 2004; Yamawo et al. 2012). Trichomes may also deter females from oviposition; however, ovipositing females may prefer a hairy leaf surface (Hawthorne et al. 1992; Heinz and Zalom 1995). In many cases, plants possessing dense trichomes have been found to be resistant to herbivores; glabrous forms occasionally appear to be more resistant to some insects (e.g. Agrawal and Spiller 2004; Sato and Kudoh 2015). Although many researchers have studied the effects of trichomes as a direct defence against herbivores, few studies have demonstrated the effects throughout the feeding stages (e.g. larvae and adults) of herbivorous insects (Sato et al. 2014).
In this study, we focused on trichome dimorphism in Persicaria lapathifolia (L.) Delarbre var. lapathifolia (Polygonaceae) (PLL, hereafter). This species generally has no trichomes on leaf surfaces (glabrous type) in many parts of Japan, but individuals bearing dense trichomes (hairy type) were found to grow in some areas in Kyushu (Fig. 1) (Satake et al. 1982). Although trichomes have various functions, preliminary observations suggested that the trichomes of PLL seemed to act as a defensive trait against herbivores (Fig. 1; A. Yamawo, personal observation).

Glabrous and hairy types of Persicaria lapathifolia var. lapathifolia naturally growing in a field and lower leaf surfaces of both types (scales: 1.5 cm). Leaves of the glabrous individuals were infested by herbivores in the field (photos taken by Akira Yamawo)
We first conducted a cultivation experiment to confirm whether the dimorphism in trichome production is under genetic control or due to phenotypic plasticity. Then, we examined the effects of trichomes against feeding, development, and reproduction of Galerucella grisescens (Joannis) (Coleoptera: Chrysomelidae), a major herbivore of PLL (e.g. Abe et al. 2007; Tokuda 2015). We also quantified chemical defensive compounds in both types of PLL leaves because some plants are known to adopt multiple defensive traits against herbivores (Koricheva et al. 2004; Yamawo et al. 2014).
Materials and methods
Plant species
Persicaria lapathifolia var. lapathifolia is an annual herb growing to a height of 50–150 cm and is distributed in roadsides, waterways, and croplands of the North Temperate Zone (Hotta et al. 1989). As mentioned earlier, this species generally has no trichomes on its leaves (glabrous type) in many parts of Japan, but individuals bearing dense trichomes (hairy type) are known to grow in some areas in Kyushu. Persicaria species are generally thought to be self-compatible (Stanford 1925; Simmons 1945).
Insect species
Galerucella grisescens is a multivoltine leaf beetle with two to three generations per year in cool areas and approximately five in warm areas (Takahashi 1957). This species is distributed widely in the Palaearctic region (Suzuki 1987; Kimoto and Takizawa 1994) and feeds mainly on the leaves of plants of Polygonaceae, but sometimes utilizes plants of Rosaceae (Suzuki 1985, 1987). Adults oviposit their egg mass (average 12–22 eggs, ranging from 8 to 27) on the underside of host leaves (Lee 1990). In our census fields, G. grisescens is a major herbivore of PLL, together with Chaetocnema granulosa (Coleoptera: Chrysomelidae) and Calybites phasianipennella (Lepidoptera: Gracillariidae) in the summer season (Tokuda 2015).
A laboratory-reared population of G. grisescens originated from adults and larvae collected from Rumex japonicus Houtt. (Polygonaceae) in Suehirobashi, Saga City in April and May 2013 was used in this study. They were reared in an incubator (BioTRON LH-220/350S, NKsystem, Osaka) in the laboratory at 25 °C with 16L:8D photoperiodic conditions. Approximately 10–20 adults were reared in each plastic container (11.5 cm long, 17 cm wide, and 5 cm high) with wet tissues (Kimwipe, Crecia Co. Ltd., Tokyo) in the bottom. A few fresh leaves of R. japonicus were offered to the beetles every 2–3 days. When egg masses were laid on leaves, the leaves were moved to petri dishes (9 cm diameter and 2 cm high) with a piece of wet filter paper (9 cm diameter) and maintained in an incubator at 25 °C with 16L:8D photoperiodic conditions. After eclosion, larvae were supplied fresh leaves of R. japonicus until pupation. Emerged adults were placed into plastic containers for continuous rearing.
Preliminary examination of offspring phenotype of P. lapathifolia lapathifolia
Preliminary experiments were conducted to confirm the phenotype of offspring of glabrous and hairy type individuals. Seeds of PLL used for the experiments were collected in October 2011 from a single individual of each type growing in Honjo, Saga, Kyushu, Japan (33°15′N, 130°18′E). Approximately 50 seeds were sown in plastic containers (8 cm long, 7 cm wide, and 5 cm high) with wet red soil (3 cm deep). The plastic containers (8 cm long, 7 cm wide, and 5 cm high) were covered with plastic wrap and put into a refrigerator (5 °C) for one month for dormancy-breaking (Watanabe and Hirokawa 1974). Following this, the containers were put into an incubator (BioTRON LH-220/350S, NKsystem, Osaka) for 5 days at 20 °C with 12L:12D photoperiodic conditions for germination. Then, emerged seedlings (approximately 3 cm; cotyledon stage) were individually planted using forceps into plastic cups (215 ml volume; 6.5 cm diameter and 9 cm high) containing 70% red soil and 30% humus, and were continuously maintained in the incubator for 2 weeks under the same conditions. The seedlings were watered sufficiently every other day. After this, 30 seedlings of the respective types were transferred to a greenhouse at Saga University and maintained for 1 month, after which they were replanted into plastic pots (17 cm diameter and 20 cm high) containing 70% red soil and 30% humus. The plants were sufficiently watered twice (6:00 and 18:00) each day. The presence or absence of leaf trichomes was determined one month after replanting.
Feeding experiment in the laboratory
Choice and no-choice feeding experiments were performed in the laboratory to confirm the effects of trichomes on feeding by G. grisescens. Seeds of PLL were collected from several individuals of each phenotype in Honjo and cultivated for two months in the greenhouse at Saga University using the method mentioned above. Their apical third and fourth leaves were used in the experiments.
In the choice test, an adult G. grisescens was offered two square leaf pieces (1 cm × 1 cm) of glabrous and hairy types of PLL for 48 h (25 °C, 16L: 8D). The assay container was a petri dish with an absorbent cotton placed on the bottom. The leaf pieces were placed near the centre of the petri dish and spaced 1 cm apart from each other. In the no-choice test, two leaf pieces of the same plant type were supplied for an adult. All other procedures were the same as in the choice test.
The leaf pieces remaining in the petri dishes were scanned (scanner, MP950, Canon), and leaf area consumed by G. grisescens was calculated using a free software, “Lia32” (produced by K. Yamamoto of Nagoya University, Nagoya, Japan). Twenty and ten replications were performed in the choice and no-choice tests, respectively.
To exclude the effect of trichomes on the consumption of glabrous and hairy type leaves by G. grisescens, another laboratory experiment using powdered leaves was conducted. Both types of PLL leaves growing nearby were collected from several individuals of each phenotype in Honjo in September 2014, dried in silica gel for a day, and pulverized by an electric mixer (A10, IKA Labortechinik, Staufen, Germany). Then, 0.5 g of leaf powder was mixed with 1.0 ml of water. Approximately 3.0 mg of the mixture was given to an adult of G. grisescens in a 1.5 ml microtube for 24 h (25 °C, 16L: 8D). Each adult was used either in glabrous or hairy type treatment. By measuring the weight of the mixture before and after the exposure using an electronic balance, the amount of mixture consumed by the adult was calculated. Eight replications were performed for each treatment in males and 12 replications for each in females, respectively.
Effects of trichomes on the larval developmental performance of G. grisescens
A laboratory experiment was conducted to confirm the effects of trichomes on the larval developmental performance of G. grisescens. An egg mass, selected randomly in a continuous rearing container, was moved to a petri dish with a piece of wet filter paper and maintained in the incubator at 25 °C with 16L: 8D photoperiodic conditions. After eclosion, 15 larvae were moved into another petri dish with a piece of wet absorbent cotton. Then, the larvae were supplied with undamaged, fresh glabrous or hairy type host leaf at the third or fourth position from the shoot tip until pupation to examine the larval duration. Both types of PLL leaves were collected from Honjo. The leaf piece was exchanged every 2 days and absorbent cotton was watered at that opportunity. Emerged adults were sexed and their fresh body weights were measured within 24 h of emergence using an electric balance (BP211D, Sartorius, Tokyo). Three replications (45 larvae in total) were conducted, respectively, for the glabrous and hairy types of PLL treatments.
Effects of trichomes on adult survival and reproductive performance of G. grisescens
Larvae of G. grisescens were reared either with glabrous or hairy type leaves of PLL as in the preceding experiment, and male and female adults that emerged on the same day were paired and put into a petri dish with wet absorbent cotton. Thirty pairs of adults were prepared for individuals grown on glabrous and hairy type leaves of PLL, respectively. Among them, 16 and 13 pairs grown on glabrous and hairy type leaves were fed glabrous type leaves, and 14 and 17 pairs were fed with the hairy type throughout this experiment.
They were observed every 2 days and adult survival and the number of eggs and egg masses were recorded. The leaf piece was also exchanged every 2 days and absorbent cotton was watered at that opportunity.
Quantification of chemical defence compounds of glabrous and hairy type leaves
To compare the chemical defence compounds between the glabrous and hairy type leaves, total phenolics and condensed tannin contents were examined. Seeds of PLL were collected from several individuals of each phenotype in Honjo and were cultivated for 2 months in the greenhouse at Saga University using the method described above. Apical third and fourth leaves were sampled from PLL cultivated in the greenhouse. Collected leaves were dried and crushed by the electric mixer. Total phenolics and condensed tannins in a 20 mg leaf powder sample were extracted using 50% methanol (10 ml) for 1 h in a 40 °C ultrasonic bath. Then, total phenolics and condensed tannin concentrations were measured using Folin–Ciocalteu method (Julkunen-Tiitto 1985) and the butanol–HCL assay (Porter and Wooduffe 1984), respectively.
Statistical analyses
Ratios of offspring phenotypes in the glabrous and hairy types in the cultivation experiment were analysed using Fisher’s exact probability test. Consumed leaf areas in the choice and no-choice feeding experiments, amounts of powdered leaves consumed by G. grisescens adults, and larval duration of G. grisescens were compared using the Mann–Whitney U test, because the distribution of the residuals differed significantly from normality. Adult weight of G. grisescens and amounts of total phenolics and condensed tannins were analysed using the Welch’s t test. Adult life span and the total number of eggs were analysed by a two-factorial ANOVA followed by the Welch’s t-test to detect the effects of larval and adult foods.
Results
Phenotype of offspring
The ratios of offspring phenotypes in the glabrous and hairy PLL types were significantly different (Fisher’s exact probability test, P < 0.001). In total, 86.7% of glabrous type offspring (n = 30) and 96.7% of hairy type offspring (n = 30) exhibited the same types as their parents.
Effects of leaf trichomes on G. grisescens
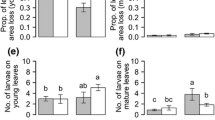
In both the choice and no-choice tests, glabrous type leaves were significantly more consumed by G. grisescens adults. The adults did not feed on the hairy type leaves at all (Fig. 2a, b). In the hairy leaf treatment, the larval duration of G. grisescens became significantly longer (Fig. 2c) and the adult body weight was significantly lower than in the glabrous leaf treatment (Fig. 2d, e). Larval survival rates were not different between the glabrous and hairy leaf treatments; 44 out of 45 larvae (97.8%,) successfully pupated in both treatments.

a, b Leaf areas of glabrous and hairy types of Persicaria lapathifolia var. lapathifolia (PLL) consumed by Galerucella grisescens adults in the choice (a n = 20) and no-choice (b n = 10) tests. Asterisks indicate significant differences between treatments (Mann–Whitney U-test, P < 0.001). c–e The larval duration (c n = 44) and the adult body weight of G. grisescens females (d n = 22) and males (e n = 22) growing on glabrous or hairy type leaves of PLL. Asterisks indicate significant differences between treatments (Mann–Whitney U-test, P < 0.05 for larval duration; Welch Two-Sample t-test, P < 0.001 for adult body weight). Abbreviations of treatments (foods) are as follows: G glabrous type, H hairy type, G→G glabrous type in larval and adult stages, H→G hairy type in larval and glabrous type in adult stages, G→H glabrous type in larval and hairy type in adult stages, and H→H hairy type in larval and adult stages. Bars indicate SD
Male adult life span was not significantly different between the glabrous and hairy leaf treatments (two-factorial ANOVA; df = 1, F = 1.14, P = 0.291 for adult food; df = 1, F = 0.001, P = 0.921 for larval food; df = 1, F = 0.001, P = 0.920 for adult food × larval food) (Fig. 3a). Female adult life span was significantly longer in the glabrous than in the hairy leaf treatment (two-factorial ANOVA; df = 1, F = 7.86, P = 0.007 for larval food; df = 1, F = 6.47, P = 0.014 for adult food; df = 1, F = 3.130, P = 0.082 for adult food × larval food) (Fig. 3b) and, within the glabrous leaf treatment, female adults reared on hairy type leaves during larval period lived longer than those reared on glabrous leaves (Welch’s t test; df = 26.922, t = 3.084, P < 0.01) (Fig. 3b). In addition, females laid a significantly larger number of eggs in the glabrous leaf treatment than in the hairy leaf treatment, regardless of the type of leaves fed (glabrous or hairy) in the larval period (df = 1, F = 114.33, P < 0.0001 for adult; df = 1, F = 0.002, P = 0.968 for larvae; df = 1, F = 0.194, P = 0.662 for adult × larval food) (Fig. 3c).

a–c The male (a; G→G: n = 16, H→G: n = 13, G→H: n = 14, H→H: n = 17) and female (b G→G: n = 13, H→G: n = 11, G→H: n = 14, H→H: n = 15) longevity of G. grisescens on glabrous or hairy type leaves of PLL. No significant differences were detected between foods in the adult stage (two-factorial ANOVA; P = 0.08287). Different letters indicate significant differences between foods in the larval stage (two-factorial ANOVA; P < 0.05). c The mean number of eggs laid by the females fed on glabrous and hairy type leaves of PLL (G→G: n = 16, H→G: n = 13, G→H: n = 14, H→H: n = 17). Different letters indicate significant differences between foods in the adult stage (two-factorial ANOVA; P < 0.001). Abbreviations of treatments (foods) are as follows: G glabrous type, H hairy type, G→G glabrous type in larval and adult stages, H→G hairy type in larval and glabrous type in adult stages, G→H glabrous type in larval and hairy type in adult stages, and H→H hairy type in larval and adult stages. Bars indicate SD
Laboratory feeding experiment using powdered leaves and defence compounds in both type of leaves
Amounts of powdered leaves consumed by G. grisescens were not significantly different between glabrous and hairy types (df = 1, F = 2.76, P = 0.11 for sex; df = 1, F = 0.12, P = 0.73 for leaf type; df = 1, F = 0.14, P = 0.71 for sex) (Fig. 4a).

a Leaf powder consumption by Galerucella grisescens adults. No significant differences were detected between treatments (n = 12 for female and n = 8 for male; two-factorial ANOVA; P = 0.726). b–c Total phenolics (b n = 5) and condensed tannin (c n = 5) concentrations in glabrous and hairy leaves of Persicaria lapathifolia var. lapathifolia. Asterisks indicate significant differences between treatments (Welch Two-Sample t-test; P < 0.001). Abbreviations of treatments are as follows: G glabrous type and H hairy type. Bars indicate SD
Significantly more total phenolics (Welch Two-Sample t test: t = 13.187, df = 9, P < 0.0001) and condensed tannins (Welch Two-Sample t test: t = 18.824, df = 9, P < 0.0001) were detected in the hairy than in the glabrous leaves of PLL (Fig. 4b, c).
Discussion
Under the same environmental conditions, most offspring of glabrous and hairy type individuals exhibited the same types as their parents. Thus, the variation in the leaf trichome production of PLL is not due to phenotypic plasticity; rather, it is under genetic control. However, the detailed mechanisms of phenotypic determination are a future study subject.
Adults of G. grisescens did not consume hairy type leaves in either the choice or no-choice experiments (Fig. 2). Moreover, adults of G. grisescens that developed on hairy type leaves during the larval period were smaller than those reared on glabrous type leaves. These results indicate that the leaf trichomes of PLL can reduce infestation by this leaf beetle, similar to other plant species (e.g. Sletvold et al. 2010; Yamawo et al. 2012; Sato and Kudoh 2015).
Agrawal and Spiller (2004) also discovered similar results in the small flea leaf beetle Chaetocnema brunnescens (Coleoptera) associated with green (glabrous) and silver (hairy) leaf phenotypes of Conocarpus erectus (Combretaceae). Adults of C. brunnescens consumed significantly larger leaf areas of the green type than the silver type, both in the choice and no-choice tests. In contrast, significant differences were shown only in the choice test for relatively large Biopsyche thoracica (Lepidoptera) larvae, suggesting that leaf trichomes of C. erectus are more effective against small herbivores (Agrawal and Spiller 2004). Sato et al. (2014) focused on the presence or absence of leaf trichomes in Arabidopsis halleri subsp. gemmifera (Brassicaceae) and an associated herbivore Phaedon brassicae Baly (Coleoptera: Chrysomelidae). Although in the choice test, adults of P. brassicae fed on significantly more glabrous leaves than hairy leaves, they consumed similar amounts of glabrous and hairy leaves in the no-choice test. Such a difference in the no-choice test may be related to feeding habits and rearing conditions of the insects used. Although G. grisescens are known to feed on Polygonaceae, Rosaceae, and some other plants, P. brassicae is fundamentally associated only with Brassicaceae. Thus, P. brassicae might be more likely to adapt to host physical defences. In addition, larval feeding may also affect the differences in the no-choice test. In the experiment by Sato et al. (2014), P. brassicae was reared on Brassica rapa L. var. glabra Regel, and in our study G. grisescens was on R. japonicus. Although we have no information on the leaf trichomes of Brassica rapa var. glabra used by Sato et al. (2014), R. japonicus, used in our study, had no visible trichomes on the leaf surfaces. Therefore, G. grisescens adults used in our no-choice test did not experience leaf trichomes during the larval period. This may more or less affect the clear result in the no-choice test. Even so, we can conclude that leaf trichomes of PLL are effective for G. grisescens because the leaf beetle reared on the hairy type achieved lower adult performance on the hairy type.
Interestingly, the larval survival rate of G. grisescens was not significantly different between individuals feeding on the glabrous and hairy type leaves. However, G. grisescens larvae feeding on hairy type leaves had significantly delayed development, and became smaller adults and shorter-lived males than those feeding on glabrous leaves (Figs. 2, 3). In the experiment by Sato et al. (2014), the weight of larvae at 1 and 4 days after release also showed no significant difference between the hairy and glabrous type leaves. However, similar to our results, the larval weight eight days after release on the hairy leaves was significantly lower than that on the glabrous leaves (Sato et al. 2014).
Galerucella grisescens females that fed on hairy type leaves during adulthood produced significantly fewer eggs than those that fed on glabrous leaves. Because G. grisescens is a multivoltine species and oviposits onto host leaves, the trichome production in PLL is effective not only to reduce the feeding area by G. grisescens, but also to increase predation and parasitism risks for G. grisescens by prolonging the larval duration and to decreasing the number of offspring by shortening male longevity and female fecundity. In general, female body size positively affects the fecundity. Even though G. grisescens individuals reared on hairy type leaves during the larval stage became smaller adults, the type of food during the larval stage did not affect the total number of eggs laid, but food during the adult stage did. In addition, on glabrous type leaves, female adult longevity grown on hairy type leaves during the larval stage was significantly longer than that grown on glabrous type leaves (Fig. 3). Although the reason is unclear, the longevity might be restricted in some extents by lifetime egg productions and slow egg production of small adults grown on hairy type leaves may result in their longer life span. Further experiments are needed to clarify the exact reason.
Based on our analysis, hairy type leaves contained more total phenolics and condensed tannins. This suggests that hairy type individuals allocate more resources, not only to the trichome production, but also to chemical defence against herbivores. However, consumptions of powdered leaves by G. grisescens were not significantly different between glabrous and hairy types. This implies that the chemical defensive compounds are not effective to prevent feeding by G. grisescens. Leaves of PLL contain the feeding deterrent, 3-hydroxy-5-methoxy-6,7-methylenedioxy flavanone, against the first instar of Gallerucida bifasciata, a close species of our focal leaf beetle, and possibly possess an insecticide or growth inhibitor to the second and third instars as well as the adult of G. bifasciata (Abe et al. 2007). Thus, G. grisescens seemed to have adapted this chemical defence to PLL, which may be effective for other herbivores including G. bifasciata.
In summary, we revealed that the leaf trichomes of PLL are an effective physical defence against larvae and adults of G. grisescens. Specifically, our study suggests that leaf trichomes have multiple defensive functions against herbivores, not only the direct reduction of feeding by larvae and adults of herbivores, but also the possible increase of mortality risks by delaying larval development and possible decrease of offspring generation by reducing adult longevity and fecundity. To clarify the maintaining mechanism of trichome dimorphism in PLL, further investigations are needed to clarify the effects of leaf trichomes and chemical compounds against other herbivores and the production cost of these defences in PLL.
References
Abe M, Niizeki M, Matsuda K (2007) A feeding deterrent from Persicaria lapathifolia (Polygonaceae) leaves to larvae of Gallerucida bifasciata (Coleoptera: Chrysomelidae). Appl Entomol Zool 49:449–456
Agrawal AA, Spiller DA (2004) Polymorphic buttonwood: effects of disturbance on resistance to herbivores in green and silver morphs of a bahamian shrub. Am J Bot 91:1990–1997
Benzing DH (1976) Bromeliad trichomes: structure, function and ecological significance. Selbyana 1:330–348
Handley R, Ekbom B, Agren J (2005) Variation in trichome density and resistance against a specialist insect herbivore in natural populations of Arabidopsis thaliana. Ecol Entomol 30:284–292
Hawthorne DJ, Shapiro JA, Tingey WM, Mutschler MA (1992) Trichome-borne and artificially applied acylsugars of wild tomato deter feeding and oviposition of the leafminer Liriomyza trifolii. Entomol Exp et Appl 65:65–73
Heinz KM, Zalom FG (1995) Variation in trichome-based resistance to Bemisia argentifolii (Homoptera: Aleyrodidae) oviposition on tomato. J Econ Entomol 88:1494–1502
Hess FD, Bayer DE, Falk RH (1974) Herbicide dispersal patterns: I. as a function of leaf surface. Weed Sci 22:394–401
Hotta M, Ogata K, Nitta A, Hoshikawa K, Yanagi M, Yamazaki K (1989) Useful plants of the world. Heibonsha, Tokyo (in Japanese)
Howe HF, Westley LC (1988) Ecological relationships of plants and animals. Oxford University Press, New York
Julkunen-Tiitto R (1985) Phenolic constituents in the leaves of northern willows: methods for the analysis of certain phenolics. J Agric Food Chem 33:213–217
Kimoto S, Takizawa H (1994) Leaf Beetles (Chrysomelidae) of Japan. Tokai University Press, Tokyo (in Japanese)
Koricheva J, Nykänen H, Gianoli E (2004) Meta-analysis of trade-offs among plant antiherbivore defenses: are plants jacks-of-all-trades, masters of all? Am Nat 163:E64–E75
Kruidhof HM, Allison JD, Hare JD (2012) Abiotic induction affects the costs and benefits of inducible herbivore defenses in Datura wrightii. J Chem Ecol 38:1215–1224
Lee JE (1990) Morphological studies on the immature stages of two Japanese species of the genus Galerucella (Coleoptera, Chrysomelidae). Jpn J Entomol 58:425–439
Porter LJ, Wooduffe J (1984) Haemanalysis: the relative astringency of proanthocyanidin polymers. Phytochemistry 23:1255–1256
Reymond P, Bodenhausen N, van Poecke RMP, Krishnamurthy V, Dicke M, Farmer EE (2004) A conserved transcript pattern in response to a specialist and a generalist herbivore. Plant Cell 16:3132–3147
Satake Y, Ohwi J, Kitamura S, Watari S, Tominari T (1982) Wild flowers of Japan Herbaceous plants Choripetalae. Heibonsha Ltd., Tokyo (in Japanese)
Sato Y, Kudoh H (2015) Tests of associational defence provided by hairy plants for glabrous plants of Arabidopsis halleri subsp. gemmifera against insect herbivores. Ecol Entomol 40:269–279
Sato Y, Kawagoe T, Sawada Y, Hirai MY, Kudoh H (2014) Frequency-dependent herbivory by a leaf beetle, Phaedon brassicae, on hairy and glabrous plants of Arabidopsis halleri subsp. gemmifera. Evol Ecol 28:545–559
Simmonds NW (1945) Polygonum persicaria L. J Ecol 33:121–131
Sletvold N, Huttunen P, Handley R, Karkkainen K, Agren J (2010) Cost of trichome production and resistance to a specialist insect herbivore in Arabidopsis lyrata. Evol Ecol 24:1307–1319
Southwood TRE (1986) Plants surface and insects—an overview. Insects and the plant surface. E. Arnold, London, pp 1–22
Stanford EE (1925) The inflorescence and flower-form in Polygonum, subgenus Persicaria. Rhodora 27:41–47
Suzuki N (1985) Habitat selection of three chrysomelid species associated with Rumex spp. Oecologia 66:187–193
Suzuki N (1987) Leaf beetle communities on Rumex japonicus. In: Kimoto S, Takeda H (eds) Insect communities in Japan: habitat segregation and diversity. Tokai University Press, Tokyo, pp 142–148 (in Japanese)
Takahashi Y (1957) Life histry and its control of Gallerucella vittaticollis. Agric Hortic 32:1782–1784 (in Japanese)
Tokuda M (2015) Host specificity and wing polymorphism in Galerucera grisescens (Coleoptera: Chrysomelidae). Nat Insects 50(12):8–11 (in Japanese)
Walters DR (2011) Plant defense: warding off attack by pathogens, herbivores and parasitic plants. Blackwell Publishing, Iowa
Watanabe Y, Hirokawa F (1974) Ecological studies on the germination and emergence of annual weeds, effects of temperatures on the dormancy-breaking in seeds of Chenopodium album, Echinochloa crusgalli var. praticola and Polygonum lapathifolia. J Weed Sci Tech 17:24–28 (in Japanese)
Yamawo A, Suzuki N, Tagawa J, Hada Y (2012) Leaf ageing promotes the shift in defence tactics in Mallotus japonicus from direct to indirect defence. J Ecol 100:802–809
Yamawo A, Tagawa J, Hada Y, Suzuki N (2014) Different combinations of multiple defence traits in an extrafloral nectary-bearing plant growing under various habitat conditions. J Ecol 102:238–247
Zvereva EL, Kozlov MV, Niemelä P (1998) Effects of leaf pubescence in Salix borealis on host-plant choice and feeding behaviour of the leaf beetle, Melasoma lapponica. Entomol Exp Appl 89:297–303
Acknowledgements
We are indebted to M. Kamo and K. Ogata for their kind support in the laboratory experiments, and to S. Kumashiro, K. Tanaka, Y. Shiomi, H. Ohashi, Y. Kubo, Y. Mochioka, S. Kai, R. Konishi, and T. Matsuo for their help during the course of this study. This study was supported in part by JSPS KAKENHI Grant Number 26870435 to MT.
Author information
Authors and Affiliations
Corresponding author
Additional information
Handling editor: Heikki Hokkanen.
Rights and permissions
About this article

Cite this article
Shirahama, S., Yamawo, A. & Tokuda, M. Dimorphism in trichome production of Persicaria lapathifolia var. lapathifolia and Its multiple effects on a leaf beetle. Arthropod-Plant Interactions 11, 683–690 (2017). https://doi.org/10.1007/s11829-017-9520-x
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11829-017-9520-x