
Abstract
The effects of supplemental ultraviolet-B (s-UV-B; 3.6 kJ m−2 day−1 above ambient) radiation on plant metabolite profile and free radical scavenging activities of Withania somnifera (an indigenous medicinal plant) under field conditions were investigated. The metabolite profiles of both leaves and roots were analyzed via GC–MS. The methanolic extracts were examined for their DPPH radical-, superoxide radical-, hydrogen peroxide-, hydroxyl radical-, and nitric oxide radical scavenging activities, ferrous ion chelating activity, and reducing power. GC–MS profile of leaves revealed increment in compounds such as isophytol (138.1 %), β-stigmasterol (183.05 %), trans-squalene (233.3 %), and Withaferin A (155.0 %), while compounds such as eugenol, β-carotene, lycopene, and vitamin E were detected in s-UV-B-treated leaves only. In roots, compounds such as ledol, neophytadiene, palmitic acid, retinol, sitosteryl oleate, and campesterol registered their presence only under s-UV-B. Methanolic extracts of treated plant organs were found to be more potent as free radical scavengers (their EC50 values being lower than those of control extracts). Anomalies were observed for nitric oxide radical scavenging in both leaves and roots. The present study indicates that s-UV-B alters the composition and contents of plant metabolites leading to an increase in their free radical scavenging activities. Hence, s-UV-B-treated plant organs might be more effective in combating oxidative stress as well as from a nutritional and health perspective.
Similar content being viewed by others


Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
Withania somnifera is an indigenous medicinal plant. It is found in the wilds in parts of northern India, western Himalayas, and forests of Madhya Pradesh and cultivated in Madhya Pradesh, Rajasthan, and comparatively drier parts of the country (Joy et al. 2001). The whole plant possesses innumerable medicinal properties with applications in the treatment of neurological disorders, Parkinson’s disease, Alzheimer’s disease, osteoarthritis, and paralysis to mention a few (Mirjalili et al. 2009; Sharma et al. 2011; Alam et al. 2012). The medicinal properties of W. somnifera are primarily attributed to alkaloids and withanolides (steroidal lactones) (Mirjalili et al. 2009). Other phyto-constituents important from plant development and/or human consumption perspective include carbohydrates/reducing sugars, proteins/amino acids, phenols, flavonoids, tannins, saponins, terpenoids, glycosides, and coumarins (Shahriar et al. 2012; Panchawat 2012; Uddin et al. 2013). The variations in the contents of these phyto-compounds depend upon a number of factors such as geographical location, climatic conditions, types of cultivation practices (including effects of fertilizers, soil type, and irrigation pattern), genetic and biochemical variations within plant species, plant part used, age of the plant, collection period, drying methods, storage conditions, extraction method, and solvent(s) used for extraction (Wendakoon et al. 2012).
Various biotic and abiotic stress factors alter the amount and/or composition of various plant metabolites (by channelizing the photosynthates towards the production of more secondary metabolites and compromising on growth, for instance). Ultraviolet-B radiation is one such abiotic stress factor which, due to anthropogenic production of ozone-depleting substances (ODSs), changing climatic conditions, and altered land-use patterns (Ballaré et al. 2011; Anderson et al. 2012; Laube et al. 2014) is reaching the Earth in increased quantities. Since sunlight is inevitable for the survival of photosynthetic organisms, they are invariably exposed to enhanced UV-B levels also. Supplementary ultraviolet B (s-UV-B) has been known to cause morphological and physiological level changes (Choudhary and Agrawal 2015) as well as altered composition and contents of pharmacologically important constituents in some medicinal plants (Zhang and Björn 2009; Ning et al. 2012) including Withania somnifera (Takshak and Agrawal 2014). Enhanced production of reactive oxygen species (ROS) and reactive nitrogen species (RNS) is another definitive response of plants to s-UV-B. They are capable of causing extensive cellular damage by reacting with membrane lipids, proteins, enzymes, nucleic acids, and carbohydrates (Mackerness et al. 2001). In humans, too, these reactive species have been known to cause a number of diseases including AIDS, cancer, Alzheimer’s- and Parkinson’s disease (Bandhopadhyay et al. 1999). Antioxidants protect cells by scavenging free radicals. Many phyto-compounds such as flavonoids, phenols, alkaloids, α-tocopherol, carotenoids, peptides, and vitamins serve as antioxidants (Formica and Regelson 1995; Rice-Evans et al. 1997) due to their redox potential which allows them to serve as reducing agents/hydrogen donors, singlet oxygen quenchers, and metal chelators (Rice-Evans et al. 1997). Thus, it follows that phyto-extracts rich in these constituents are likely to show high free radical scavenging capabilities.
Preliminary phytochemical analysis of plant extracts provides knowledge about important bioactive constituents. The latter can be further quantified to determine their commercial availability, and depending upon the type(s) of phyto-constituents present, a cursory appraisal of the medicinal properties of the plant can be made. As of today, majority of studies on medicinal plants have focused on the metabolite profile to the extent of detailing them in different tissues of the plant (whether it is at the preliminary level or detailed analysis via GC–MS). No study, to the best of our knowledge, has as yet reported the alterations in metabolite profile of the organic (methanolic) extracts of W. somnifera roots and leaves under s-UV-B stress conditions. In our previous study (Takshak and Agrawal 2014) we had quantified various groups of secondary metabolites (alkaloids, anthocyanins, carotenoids, flavonoids, phytosterols, saponins, and tannins) in the leaves and roots of W. somnifera. The present study, however, was conducted with the following objectives: (1) carry out preliminary phytochemical screening to determine the type and nature of compounds present in the different extracts of leaf and root tissue of the test plant, (2) further analyze elaborate metabolite profile (to identify the key individual components) of plant methanolic extracts of leaf and root tissue via gas chromatography/mass spectrometry (GC–MS) and analyze the changes in their relative concentrations under s-UV-B based on their retention peak area and (3) study the properties of the methanolic extracts prepared from control and s-UV-B-treated plant organs in terms of determining their free radical scavenging capacities.
Materials and methods
Experimental design and s-UV-B treatment
The experiments were conducted in Botanical Garden, Department of Botany, Banaras Hindu University, Varanasi (25°80′N, 82°03′E, and 76 m above mean sea level), situated in the Eastern Gangetic Plains of India. The experimental period extended from end of March to mid-July 2012 with average maximum temperature being 37.8 °C and the average minimum temperature being 24.4 °C. Relative humidity varied between 41.9 and 61.3 %. 1-month-old W. somnifera plants were obtained from nursery and transplanted in experimental plots of 1 m × 1 m (12 plants per plot; 3 rows with 4 plants in each row). The plots were prepared in triplicate for control and well as s-UV-B treatment. Plants were watered regularly as per the requirements.
Once the plants were established in the field, they were exposed to s-UV-B radiation using UV-B lamps (Q Panel UV-B 313 40 W fluorescent lamps, Q panel Inc., Cleveland, OH, USA) fitted on steel frames at equal distances directly above the plant canopy. The control plants received only ambient UV-B dose (9.6 kJ m−2 day−1) while treated plants received ambient +3.6 kJ m−2 day−1 s-UV-B dose (simulating 10 % ozone depletion at Varanasi) as weighted by Caldwell (1971) generalized plant action spectrum normalized at 300 nm. For the purpose UV-B lamps were wrapped with 0.13 mm cellulose diacetate filters (Cadillac Plastic Co., Baltimore, MD, USA; transmission down to 280 nm); the filters were changed every week due to their photo-degradation by UV-B. The distance between plant canopy and UV-B lamps was kept constant throughout the experiment. s-UV-B treatment was given to the plants for 3 h (11:00 to 14:00 h) during the solar noon period. UV-B irradiance was measured using Ultraviolet Intensity Meter (UVP Inc., San Gabriel, CA, USA) and biologically effective UVB (UV-BBE) using Spectropowermeter (Scientech, Boulder, USA).
Preliminary phytochemical screening
Sample preparation
The plants were harvested at 100 DAT, shade dried, and the leaves and roots were powdered. The coarse powder was passed through a double layer of cheese cloth to obtain a uniform fine powder. The samples were prepared as per the protocol of Paramakrishnan et al. (2012). The powdered plant material (10 g) was extracted with different solvents (100 ml each; methanol, ethanol, petroleum ether, acetone, chloroform, and water). The powder: solvent ratio was kept at 1:10 to ensure complete extraction. Extraction was carried out via cold maceration for 72 h with intermittent shaking of the flasks; cold maceration was preferred to prevent the evaporation of volatile compounds. After extraction, solutions were filtered through Whattmann filter paper (No. 4), extracts collected and stored at 4 °C for further use.
Sample analysis
The parameters were analyzed following the methods outlined in Savithramma et al. (2011), Vijayalakshmi and Ravindhran (2012), and Sawant and Godghate (2013). The extracts were tested for the presence of alkaloids, amino acids/proteins, anthocyanins, anthraquinones, cellulose, coumarins, emodins, flavonoids, glycosides, leucoanthocyanins, lignin, phenols, phlobatannins, reducing sugars, resins/gums/mucilages, saponins, steroids, tannins, and terpenes.
GC–MS analysis and identification of compounds
Sample preparation
10 g powdered sample was soaked in 100 ml methanol for 72 h with intermittent shaking. The mixture was centrifuged at 3000g and the supernatant decanted. The rest of the protocol was followed as per Sharma et al. (2013) with some modifications. 10 ml supernatant was treated with 0.5 N H2SO4 (20 ml) and a few drops of NH4OH were added to make it alkaline. 10 ml of chloroform was added to the extract and shaken for 10 min to ensure thorough mixing. The solution was kept at room temperature for 24 h for incubation, after which the upper layer was discarded. 10 ml solution from the lower layer (now containing the majority of the phytoconstituents) was concentrated to 2 ml which was used for analysis by GC–MS. Analyses were performed using three replicates for each treatment.
GC–MS conditions and specifications
The prepared extracts of both leaves and roots of the control and treated test plants were subjected to gas chromatography–mass spectroscopy (GC–MS) analysis. The samples were analyzed on GC–MS QP-2010 Plus (Shimadzu) equipped with Rtx-5 MS (30 m × 0.25 mm i.d. × 0.25 μm film thickness) column. 1.0 μl of the sample was injected into the system at 250 °C with helium as carrier gas with a flow rate of 1.21 ml min−1. The column oven temperature was initially maintained at 100 °C for 4 min, then increased to 250 °C at the rate of 6 °C min−1 with a hold time of 5 min and again raised up to 310 °C at the rate of 15 °C with a hold time of 12 min, at which it was maintained till the end of the program. Other GC conditions such as pressure, linear velocity, purge flow, and split ratio were maintained at 90.4 kPa, 40.9 cm s−1, 3.0 ml min−1, and 1:10, respectively. The MS operating conditions were as follows: interface temperature: 260 °C; ion source temperature: 230 °C; mass scan (m/z): 40–600 amu for 0.5 s; solvent cut time: 4.5 min; scan speed: 1250 amu s−1; MS running time: 44.99 min; threshold: 1000.
Identification of compounds
The identification of the components of the extracts was based on a comparison of their relative retention indices and mass spectra fragmentation patterns with the available WILEY and NIST libraries (WILEY8.LIB and NIST08.LIB) as well as with those of authentic reference compounds previously reported.
Antioxidant activity determinations
Sample preparation
1 g powdered sample was extracted in 10 ml methanol via cold maceration for 72 h with intermittent shaking. The mixture was centrifuged at 3000 g and supernatant decanted. From this core solution (105 μg ml−1), stock solution of 1000 μg ml−1 was prepared from which a range of dilutions (50, 100, 150, 200, 250, 300, 350, 400, 450, and 500 μg ml−1) were further prepared. Ascorbic acid (AsA) (for all assays) and butylated hydroxyl-toluene (BHT) (for DPPH scavenging and reducing power assays), prepared in same concentrations, served as reference compounds against which the scavenging activities of the extracts were determined. All protocols were performed with five replicates.
Free radical scavenging activities
DPPH radical scavenging activity (Blois 1958): To 3 ml of plant methanolic extract, 1 ml of DPPH (0.2 mM in methanol) was added. The solution was incubated in dark for 30 min at room temperature. The absorbance was recorded at 517 nm. DPPH in methanol, without plant extracts, served as control, while methanol served as blank. The radical scavenging activity was calculated as follows:
A0: absorbance of control; A1: absorbance of sample
Superoxide radical scavenging activity (McCord and Fridovich 1969): The total reaction volume for the assay comprised of 0.2 ml EDTA (6.6 mM containing 3 μg NaCN), 0.1 ml NBT (50 μM), 0.05 ml riboflavin (2 μM), 2.63 ml phosphate buffer (67 mM, pH 7.8), and 0.02 ml various concentrations of plant extracts. All the solutions were vortexed and initial absorbance recorded at 560 nm. These were then subjected to illumination with incandescent lamps (15 W) for 30 min, and again absorbance was recorded at 560 nm. The difference in absorbance before and after illumination indicated superoxide radical scavenging activity. Phosphate buffer served as blank. The radical scavenging activity was calculated as follows:
where ΔA = absorbance after illumination − absorbance before illumination
Hydrogen peroxide scavenging activity (Ruch et al., 1989): To 2.4 ml of phosphate buffer (0.1 M, pH 7.4), 0.6 ml H2O2 (40 mM) was added. 1 ml of different concentrations of various extracts were added to the solution and incubated at room temperature for 10 min. Absorbance was recorded at 230 nm. Controls were prepared without plant extract while blank comprised of only phosphate buffer without H2O2. H2O2 scavenging activity was calculated as follows:
A0: absorbance of control; A1: absorbance of the sample
Hydroxyl radical scavenging activity (Kunchandy and Rao 1990): Final volume of 1 ml of reaction solution comprised of 0.1 ml deoxyribose (2.8 mM), 0.1 ml FeCl3 (0.1 mM), 0.1 ml EDTA (0.1 mM), 0.1 ml H2O2 (1 mM), 0.1 ml ascorbate (0.1 mM), 0.48 ml KH2PO4-KOH buffer (20 mM, pH 7.4), and 20 μl of plant extracts. The solutions were incubated at 37 °C for 1 h and 1 ml 1 % TBA was added to them after incubation. The solutions were heated at 95 °C for 20 min to develop color, allowed to cool and TBARS formation was measured spectrophotometrically at 532 nm. The controls were prepared without the extracts and KH2PO4-KOH buffer was used as blank. Hydroxyl radical scavenging activity was calculated as follows:
A0: absorbance of control; A1: absorbance of the sample
Nitric oxide radical scavenging activity (Green et al. 1982): 2 ml sodium nitroprusside (100 mM), 0.5 ml phosphate buffer saline (PBS; 0.025 M, pH 7.4), and 0.5 ml of plant extracts were mixed and incubated at 25 °C for 30 min. 0.5 ml Griess reagent (1 % sulphanilamide, 2 % O-phosphoric acid, and 0.1 % naphthylethylene diamine dihydrochloride) was added and the solution incubated for another 30 min. The absorbance of the resulting pink color was measured at 546 nm. Blank comprised of PBS. Nitric oxide radical scavenging activity was calculated as follows:
A0: absorbance of control; A1: absorbance of the sample
Ferrous ion chelating activity (Dinis et al. 1994): 0.5 ml of various concentrations of the extracts were added to 1.5 ml distilled water and 0.5 ml FeCl2 (2 mM). The reaction was started by the addition of 0.1 ml ferrozine (5 mM). The solution was vortexed for 1 min to ensure thorough mixing and allowed to stand for 10 min at room temperature. Thereafter, its absorbance was recorded at 562 nm. Distilled water was used as blank. Ferrous ion chelating activity was calculated as follows:
A0: absorbance of control; A1: absorbance of the sample
Reducing power assay (Oyaizu 1986): to 2.5 ml phosphate buffer (0.2 M, pH 6.6), 2.5 ml potassium ferricyanide (1 %) was added followed by 1 ml of extracts. This solution was incubated at 50 °C for 30 min, allowed to cool, and 2.5 ml 10 % TCA was added to it. The solution was centrifuged at 3000 rpm for 10 min. To 2.5 ml of supernatant, an equal volume of distilled water and 0.5 ml FeCl3 (1 %) were added. The solution was allowed to stand for 10 min and its absorbance recorded at 700 nm. Phosphate buffer served as blank. The reducing power of plant extracts was reported in terms of absorbance at 700 nm.
Determination of EC50 values
The EC (‘effective’ concentration) of any drug/toxicant/extract that produces a response half-way between the baseline and maximal response is defined as EC50. The EC50 values were calculated to determine the potency of the control and treated plants’ methanolic extracts of leaves and roots in terms of radical scavenging activities. Lower the EC50 value, higher the potency of the extract.
Statistical analysis
Means and standard errors of the samples were calculated. The means of values obtained via GC–MS are reported as the average of three replicates while the free radical scavenging capacities are reported as mean of five replicates. Mean differences were determined by applying Student’s t test. All statistical analyses were performed using the SPSS software version 16.
Results
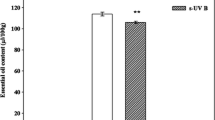
The compounds that were tested for their presence in phytoextracts of leaves and roots of W. somnifera are outlined in Table 1. Maximum number of phytoconstituents was identified in methanolic plant extracts. Anthraquinones, emodins, and resins/gums were absent in all extracts of both organs. The metabolite profile of phytoextracts showed changes in composition and concentration of various metabolites. The foliar extracts of W. somnifera identified a total of 43 and 83 compounds in control and treated plants, respectively. Similarly, in control root extracts 34 components were identified compared to a total of 59 in s-UV-B-treated roots (Figs. 1, 2). Alterations in the content of some these compounds due to s-UV-B are given in Tables 2 and 3.

Comparative changes in the chromatographic profiles of leaf extracts of Withania somnifera

Comparative changes in the chromatographic profiles of root extracts of Withania somnifera
In s-UV-B-treated leaves, some of the phytoconstituents accumulated in higher percentage; for instance, isophytol (138.10 %), methyl stearate (65.00 %), ethyl stearate (120.00 %), nonacosane (107.69 %), trans-squalene (233.33 %), β-stigmasterol (183.05 %), and Withaferin A (155.00 %). On the other hand, compounds such as crinosterol and cholesterol were found to be appreciably reduced by 44.19 and 21.38 %, respectively. Some other important medicinal compounds that were detected in s-UV-B-treated leaves included eugenol, oleyl alcohol, geranylgeraniol, solanesol, dihydrobrassicasterol, β-carotene, brassicasterol, vitamin E, and lycopene (Table 2).
In roots, more variable profile was observed compared to leaves. Compounds like neophytadiene showed an increment of 17.14 % while some other alcohols and esters showed decreased concentrations such as 2,4-Ditert-butyl phenol (61.89 %), phytol (4.65 %), methyl palmitate (76.04 %), ethyl palmitate (88.48 %), methyl stearate (82.57 %), and methyl behenate (67.66 %). Some other medicinal compounds that were detected after s-UV-B treatment of roots include ledol, dibutyl phthalate, palmitic acid, stigmasterol acetate, and retinol, while some other compounds such as podocarpic acid, ethyl oleate, ethyl stearate, methyl oleate, and α-selinene were reduced to non-detectable limits (Table 3).
As per the GC–MS results, it is clearly indicated that s-UV-B alters the metabolite profile of plants. As such it can be safely assumed that altered plant extract composition is also likely to alter the free radical scavenging capacity of these extracts. The free radical scavenging capabilities of methanolic extracts of control and treated W. somnifera leaves and roots are compared on the basis of their EC50 values and are outlined in Table 4.
The free radical scavenging activities increased with increasing concentrations of the extracts (Supplementary Fig. 1). On comparing the leaf extracts for their scavenging activities, it was found that s-UV-B-treated plant extracts were more potent in scavenging DPPH, ·O2 −, H2O2, and ·OH radicals (as depicted by their lower EC50 values, Table 4) compared to the control plants, though the difference was non-significant in case of the ·O2 − scavenging activity (EC50 = 300.01 and 291.79 μg ml−1 for control treatment, respectively). NO scavenging activity was significantly higher in case of control leaves (EC50 = 225.86 μg ml−1) compared to the treated leaves (EC50 = 258.94 μg ml−1). Fe2+ chelating activity and reducing power were again significantly higher for s-UV-B-treated leaves (Table 4). Methanolic extracts of the roots also depicted a similar trend with DPPH ·O2 −, H2O2, and ·OH scavenging activities being higher for s-UV-B-treated roots (the difference was not significant for DPPH radical scavenging activity, Table 4). Similar to leaves, NO scavenging activity was higher for control roots (EC50 = 271.11 μg ml−1 as against EC50 = 320.19 μg ml−1 for s-UV-B-treated roots). Fe2+ chelating activity and reducing power were found to be greater for s-UV-B-treated plant roots (Table 4).
Discussion
The leaf and root methanolic extracts of W. somnifera contained maximum number of pharmacologically important phyto-constituents (alkaloids, anthocyanins, coumarins, flavonoids, glycosides, phenols, quinones, saponins, steroids, tannins, and terpenes). Some of these (alkaloids, anthocyanins, flavonoids, phenols, saponins, steroids, and tannins) have been quantified and found to be increased under s-UV-B stress (Takshak and Agrawal 2014). All these phyto-constituents are important for normal plant development and functioning, to protect plants against various biotic and abiotic stresses (e.g., anthocyanins, Olsson et al. 1999; flavonoids, Agati et al. 2012; phenolics, Dai and Mumper 2010; phytosterols, Posé et al. 2009; and lignin, Boudet 2000), as well as from human health and nutraceutical perspectives (e.g., alkaloids, Sharma et al. 2011; anthocyanins, Ross and Kasum 2002; coumarins, Riverio et al. 2010; flavonoids, Nambiar et al. 2010; glycosides, Mithen et al. 2000; phenolics, Willis et al. 2009; quinones, Babula et al. 2009; saponins, Man et al. 2010; phytosterols, Woyengo et al. 2009; tannins, Cai et al. 2004; and terpenes, Paduch et al. 2007).
Data regarding s-UV-B induced changes in phytoextract composition of W. somnifera have not been reported to date; so it is not possible to compare our results with other studies. However, investigations on individual compounds respecting s-UV-B help substantiate our findings. Pharmacological importance of some of the compounds is detailed in Supplementary Table 1. Chang et al. (2009) found enhanced levels of eugenol in the essential oil of basil plants under s-UV-B similar to our results (where eugenol was detected in s-UV-B-treated leaves indicating its enhanced concentration). Trans-squalene (an isoprenoid) is an intermediate in the biosynthesis of cholesterol. Though it increased considerably under s-UV-B treatment in leaves (233.33 %), cholesterol was found to be reduced (by 21.38 %). This might probably indicate the plant’s adaptation to stress by directing the precursors towards enhanced production of secondary metabolites (overexpression of squalene synthase, and hence squalene, has been shown to result in enhanced withanolide production in W. somnifera; Grover et al. 2013). Gil et al. (2012) speculated that squalene overproduction in UV-B-treated grape plants increases phytosterol and triterpene biosynthesis. Studies by Jäpelt et al. (2011) indicate that in all of their test plants except Solanum glaucophyllum, cholesterol concentrations were found to be reduced under UV treatment.
Lycopene and β-carotene were detected in s-UV-B-treated W. somnifera leaves (indicating their increased concentrations compared to the control). Their concentrations were found to be reduced in tomato fruit skin under UV-B excluded conditions (Becatti et al. 2009). Vitamin E (α-tocopherol) was also detected in s-UV-B-treated leaves of the test plant. s-UV-B treatment has been reported to promote the synthesis of α-tocopherol in Anabaena doliolum (Bhargava et al. 2007), and Vitis vinifera (Gil et al. 2012). Withaferin A, a steroidal lactone, is one of the important withanolides present of W. somnifera. Above-ground plant parts of W. somnifera recorded an overall increase of 12.4 % in withaferin A concentration as determined by HPLC analysis (Takshak and Agrawal 2014). Phytol (a diterpene alcohol) declined under s-UV-B in the present study; it was found to be increased in grapes exposed to high UV-B irradiation (Gil et al. 2012), contrary to our results. Other compounds such as ledol (sesquiterpene alcohol), and retinol (diterpene alcohol) were found to be present only in s-UV-B-treated roots (indicating their increased concentrations) which is significant from a nutritional perspective as these compounds have been known to effective against cancer and dermatological disorders (Spiridonov et al. 2005; Huang et al. 2009). However, s-UV-B negatively affected podocarpic acid concentration which is an important anti-cancer, oestrogenic, anti-inflammatory, cardio-protective, and cholesterol-lowering agent (Liu et al. 2005; McKee et al. 2014).
Scavenging of free radicals is beneficial for the plant to survive various abiotic and biotic stresses as well as from human health perspective (epidemiological studies have demonstrated phyto-nutrients/extracts of a number of medicinal plants as dietary supplements are capable of preventing/ameliorating oxidative damage and related diseases; Radimer et al. 2004). In this study, we demonstrated the antioxidant properties of methanolic extracts of W. somnifera roots and leaves (both under control conditions as well as under s-UV-B treatment). Again, to the best of our knowledge, no previous studies describing the effects of abiotic stresses on the free radical scavenging capacities of phytoextracts are available to help corroborate our findings. However, as is evident from their EC50 values, the extracts of the treated organs showed stronger DPPH radical, superoxide anion scavenging, hydrogen peroxide scavenging, hydroxyl radical scavenging, reducing power and metal chelating activities when compared to their control counterparts. It has already been mentioned that many groups of phyto-constituents as well as individual components detected in W. somnifera extracts possess antioxidative properties (e.g., alkaloids, anthocyanins, carotenoids, flavonoids, steroids, and tannins), and increase in their concentrations in s-UV-B-treated plant organs (Takshak and Agrawal 2014) might be responsible for their enhanced scavenging activities in extracts. The differences in the scavenging activities of the extracts might be attributed, not only to the concentration of the scavenging components, but also to the structural individuality of these components (e.g., the number and position of the hydroxyl groups), which in turn determine their electron donating abilities (Ebrahimzadeh et al. 2009). Lower NO scavenging activity of s-UV-B-treated plant extracts obtained in our results indicates their higher concentrations under UV-B stress. Though detrimental to cells at high concentrations, NO also serves as a signaling molecule in plant; it has been shown to induce the upregulation of chalcone synthase and chalcone isomerase genes which are vital entities of the phenylpropanoid pathway (Tossi et al. 2011; Zhang et al. 2011). The flavonoids derived from this pathway protect the plants from damage induced by s-UV-B. The enhanced chalcone isomerase activity in leaves and roots of W. somnifera might be, in part, due to enhanced concentrations of NO produced under s-UV-B in these organs (Takshak and Agrawal 2014).
Conclusions
The present study leads to the conclusion that s-UV-B radiation alters the number as well as the composition of the metabolites found in the leaves and roots of W. somnifera. The extracts of the treated plant organs are more potent in scavenging free radicals which indicate the plant’s ability to adapt to oxidative stress. This also denotes that pharmacological preparations from treated plant organs might be more effective in the treatment of oxidative stress and related disorders in humans. However, our study is the first attempt to determine the alterations in plant metabolite profile under s-UV-B stress and consequent alterations in their scavenging capacities. Future work could be directed at the quantitative analysis of individual metabolites to determine their role in overall plant performance as well as from nutraceutical perspective. Moreover, since the plant metabolite profile is dependent upon a number of environmental conditions and extraction factors, further studies on microbial and animal models are required with a view to optimizing their medicinal potential.
Author contribution statement
All the experimental analysis as well as drafting of the manuscript with statistical analysis was performed by Swabha Takshak under the supervision of Prof. S. B. Agrawal. Both the authors read and approved the final manuscript.
Abbreviations
- AsA:
-
Ascorbic acid
- BHT:
-
Butylated hydroxytoluene
- DAT:
-
Days after transplantation
- DPPH:
-
1,1-Diphenyl-2-picrylhydrazyl
- EC:
-
Effective concentration
- GC–MS:
-
Gas chromatography mass spectroscopy
- MDA:
-
Malondialdehyde
- ODSs:
-
Ozone depleting substances
- RNS:
-
Reactive nitrogen species
- ROS:
-
Reactive oxygen species
- UV:
-
Ultraviolet
References
Agati G, Azzarello E, Pollastri S, Tattini M (2012) Flavonoids as antioxidants in plants: location and functional significance. Plant Sci 196:67–76
Alam N, Hossain M, Khalil MI, Moniruzzaman M, Sulaiman SA, Gan SA (2012) Recent advances in elucidating the biological properties of Withania somnifera and its potential role in health benefits. Phytochem Rev 11:97–112
Anderson JG, Wilmouth DM, Smith JB, Sayers DS (2012) UV dosage levels in summer: increased risk of ozone loss from convectively injected water vapour. Science 337:835–839
Babula P, Adam V, Havel L, Kizek R (2009) Noteworthy secondary metabolites naphthoquinones- their occurrence, pharmacological properties and analysis. Cuur Pharm Anal 5:47–58
Ballaré CL, Caldwell MM, Flint SD, Robinson AA, Bornman JF (2011) Effects of solar ultraviolet radiation on terrestrial ecosystems. Patterns, mechanisms, and interactions with climate change. Photochem Photobiol Sci 10:226–241
Bandhopadhyay U, Das A, Bannerjee RK (1999) Reactive oxygen species: oxygen damage and pathogenesis. Curr Sci 77:658–666
Becatti E, Petroni K, Giuntini D, Castagna A, Calvenzani V, Serra G, Mensuali-Sodi A, Tonelli C, Ranieri A (2009) Solar UV-B radiation influences carotenoid accumulation of tomato fruit through both ethylene-dependent and -independent mechanisms. J Agric Food Chem 57:10979–10989
Bhargava P, Atri N, Srivastava AK, Rai LC (2007) Cadmium mitigates ultraviolet-B stress in Anabaena doliolum: enzymatic and non-enzymatic antioxidants. Biol Plant 51:546–550
Blois MS (1958) Antioxidants determination by the use of a stable free radical. Nature 4617:1199–1200
Boudet AM (2000) Lignins and lignification: selected issues. Plant Physiol Biochem 38:81–96
Cai YZ, Luo Q, Sun M, Corke H (2004) Antioxidant activity and phenolic compounds of 112 traditional Chinese medicinal plants associated with anticancer. Life Sci 74:2157–2184
Caldwell MM (1971) Solar ultraviolet radiation and the growth and development of higher plants. In: Giese AC (ed) Phytophysiology. Academic Press, New York, pp 131–177
Chang X, Alderson PG, Wright CJ (2009) Enhanced UV-B radiation alters basil (Ocimum basilicum L.) growth and stimulates the synthesis of volatile oils. J Hortic For 1:27–31
Choudhary KK, Agrawal SB (2015) Effect of elevated ultraviolet-B on four tropical soybean cultivars: quantitative and qualitative aspects with special emphasis on gas exchange, chlorophyll fluorescence, biomass and yield. Acta Physiol Plant. doi:10.1007/s11738-015-1780-4
Dai J, Mumper RJ (2010) Plant phenolics: extraction, analysis and their antioxidant and anticancer properties. Molecules 15:7313–7352
Dinis TCP, Madeira VMC, Almeida LM (1994) Action of phenolic derivatives (acetoaminophen, salycilate, and 5-aminosalycilate) as inhibitors of membrane lipid peroxidation and as peroxyl radical scavengers. Arch Biochem Biophys 315:161–169
Ebrahimzadeh MA, Nabavi SF, Nabavi SM (2009) Antioxidant activities of methanol extract of Sambucus ebulus L. flower. Pak J Biol Sci 12:447–450
Formica JV, Regelson W (1995) Review of the biology of quercetin and related bioflavonoids. Food Chem Toxicol 33:1061–1080
Gil M, Pontin M, Berli F, Bottini R, Piccoli P (2012) Metabolism of terpenes in the response of grape (Vitis vinifera L.) leaf tissues to UV-B radiation. Phytochem 77:89–98
Green LC, Wagner DA, Glogowski J, Skipper PL, Wishnok JK, Tannenbaum SR (1982) Analysis of nitrate and 15N in biological fluids. Anal Biochem 126:131–136
Grover A, Samuel G, Bisaria VS, Sundar D (2013) Enhanced withanolide production by overexpression of squalene synthase in Withania somnifera. J Biosci Bioeng 115:680–685
Huang Z, Lin Y, Fang J (2009) Biological and pharmacological activities of squalene and related compounds: potential uses in cosmetic dermatology. Molecules 14:540–554
Jäpelt RB, Silvestro D, Smedsgaard J, Jensen PE, Jakobsen J (2011) LC-MS/MS with atmospheric pressure chemical ionisation to study the effect of UV treatment on the formation of vitamin D3 and sterols in plants. Food Chem 129:217–225
Joy PP, Thomas J, Mathew S, Skaria BP (2001) Medicinal plants. In: Bose TK, Kabir J, Das P, Joy PP (eds) Tropical hortic, vol 2. Naya Prokash, Calcutta, pp 449–632
Kunchandy E, Rao MNA (1990) Oxygen radical scavenging activity of curcumin. Int J Pharm 58:237–240
Laube JC, Newland MJ, Hogan C, Brenninkmeijer CAM, Fraser PJ, Martineire P, Oram DE, Reeves CE, Röckmann T, Schwander J, Witrant E, Sturges WT (2014) Newly detected ozone-depleting substances in the atmosphere. Nat Geosci 7:266–269
Liu W, Chen S, Dropinski J, Colwell L, Robins M, Szymonifka M, Hayes N, Sharma N, MacNaul K, Hernandez M, Burton C, Sparrow CP, Menke JG, Singh SB (2005) Design, synthesis, and structure–activity relationship of podocarpic acid amides as liver X receptor agonists for potential treatment of atherosclerosis. Bioorg Med Chem Lett 15:4574–4578
Mackerness AHS, Fred JC, Jordan B, Thomas B (2001) Early signaling components in ultraviolet-B responses: distinct roles for reactive oxygen species and nitric oxide. FEBS Lett 489:237–242
Man S, Gao W, Zhang Y, Huang L, Liu C (2010) Chemical study and medical application of saponins as anti-cancer agents. Fitoterapia 81:703–714
McCord JM, Fridovich I (1969) Superoxide dismutase. An enzymic function for erythrocuprein (hemocuprein). J Biol Chem 244:6049–6055
McKee NA, Bradford S, Parish DE, Neely WC, Parish EJ (2014) Diversity-oriented synthesis of a library of podocarpic acid derivatives. Austin J Bioorg Org Chem 1:7–13
Mirjalili MH, Moyano E, Bonfill M, Cusido RM, Palazón J (2009) Steroidal lactones from Withania somnifera, an ancient plant for novel medicine. Molecules 14:2373–2393
Mithen RF, Dekker M, Verkerk R, Rabot S, Johnson IT (2000) The nutritional significance, biosynthesis and bioavailability of glucosinolates in human foods. J Sci Food Agric 80:967–984
Nambiar VS, Daniel M, Guin P (2010) Characterization of polyphenols from coriander leaves (Coriandrum sativum), red amaranthus (A. paniculatus) and green amaranthus (A. frumentaceus) using paper chromatography: and their health implications. J Herb Med Toxicol 4:173–177
Ning W, Peng X, Ma L, Cui L, Lu X, Wang J, Tian J, Li X, Wang W, Zhang L (2012) Enhanced secondary metabolites production and antioxidant activity in post-harvest Lonicera japonica Thunb. in response to UV radiation. Innov Food Sci Emerg Technol 13:231–243
Olsson LC, Veit M, Borman JF (1999) Epidermal transmittance and phenolic composition in leaves of atrazine-tolerant and atrazine-sensitive cultivars of Brassica napus grown under enhanced UV-B radiation. Physiol Plant 107:259–266
Oyaizu M (1986) Studies on products of browning reaction. Antioxidative activities of products of browning reaction prepared from glucosamine. Jan J Nutr Diet 44:307–315
Paduch R, Kandefer-Szersze M, Trytek M, Fiedurek J (2007) Terpenes: substances useful in human healthcare. Arch Immunolog et Ther Exp (Warsz) 55:315–327
Panchawat S (2012) Pharmacological evaluation of Withania somnifera leaf. Asian J Biochem Pharma Res 2:153–158
Paramakrishnan N, Ahuja J, Suresh J, Khan MN, Sebastian M (2012) Evaluation of acute oral toxicity of aerial parts of Artemisia parviflora Roxb. in Swiss albino mice. Der Pharm Sin 3:99–103
Posé D, Castanedo I, Borsani O, Nieto B, Rosado A, Taconnat L, Ferrer A, Dolan L, Valpuesta V, Botella MA (2009) Identification of the Arabidopsis dry2/sqe1-5 mutant reveals a central role for sterols in drought tolerance and regulation of reactive oxygen species. Plant J 59:63–76
Radimer K, Bindewald B, Hughes J, Ervin B, Swanson C, Picciano MF (2004) Dietary supplement use by US adults: data from the National Health and Nutrition Examination Survey, 1999–2000. Am J Epidemiol 160:339–349
Rice-Evans CA, Miller NJ, Paganga G (1997) Antioxidant properties of phenolic compounds. Trends Plant Sci 2:152–159
Riverio ME, De Kimpe N, Moglioni A, Vazquez R, Monczor F, Shayo C, Davio C (2010) Coumarins: old compounds with novel therapeutic perspectives. Curr Med Chem 17:1325–1338
Ross JA, Kasum CM (2002) Dietary flavonoids: bioavailability, metabolic effects, and safety. Ann Rev Nutr 22:19–34
Ruch RJ, Cheng SJ, Klaunig JE (1989) Prevention of cytotoxicity and inhibition of intracellular communication by antioxidant catechins isolated from Chinese green tea. Carcinogenesis 10:1003–1008
Savithramma N, Linga Rao M, Suhrulatha D (2011) Screening of medicinal plants for secondary metabolites. Middle-East J Sci Res 8:579–584
Sawant RS, Godghate AG (2013) Comparitive studies of phytochemical screening of Carissa carandus Linn. Asian J Plant Sci Res 3:21–25
Shahriar M, Bahar A, Hossain MI, Akhter S, Haque MA, Bhuiyan MA (2012) Preliminary phytochemical screening, in vitro antioxidant, and cytotoxic activity of five different extracts of Withania somnifera root. Int J Pharm 2:450–453
Sharma V, Sharma S, Paliwal R (2011) Withania somnifera: a rejuvenating ayurvedic medicinal herb for the treatment of various human ailments. Int J PharmTech Res 3:187–192
Sharma RA, Goswami M, Yadav A (2013) GC-MS screening of alkaloids of Withania somnifera L. in vivo and in vitro. Indian J Appl Res 3:63–66
Spiridonov NA, Konovalov DA, Arkhipov VA (2005) Cytotoxicity of some Russian ethnomedicinal plants and plant compounds. Phytother Res 19:428–432
Takshak S, Agrawal SB (2014) Secondary metabolites and phenylpropanoid pathway enzymes as influenced under supplemental ultraviolet-B radiation in Withania somnifera Dunal, an indigenous medicinal plant. J Photochem Photobiol B: Biol 140:332–343
Tossi V, Amenta M, Lamattina L, Cassia R (2011) Nitric oxide enhances plant ultraviolet-B protection up-regulating gene expression of the phenylpropanoid biosynthetic pathway. Plant Cell Environ 34:909–924
Uddin G, Gul S, Rauf A (2013) Preliminary phytochemical screening, in vitro antimicrobial and antioxidant evaluation of Withania somnifera Dunal. World Appl Sci J 27:562–565
Vijayalakshmi R, Ravindhran R (2012) Preliminary comparative phytochemical screening of root extracts of Diospyrus ferrea (Wild.) Bakh and Aerva lanata (L.) Juss. Ex Schultes. Asian J Plant Sci Res 2:581–587
Wendakoon C, Calderon P, Gagnon D (2012) Evaluation of selected medicinal plants extracted in different ethanol concentrations for antibacterial activity against human pathogens. J Med Act Plants 1:60–68
Willis LM, Shukitt-Hale B, Joseph JA (2009) Recent advances in berry supplementation and age-related cognitive decline. Curr Opin Clin Nutr Metab Care 12:91–94
Woyengo TA, Ramprasath VR, Jones PJH (2009) Anticancer effects of phytosterols. Eur J Clin Nutr 63:813–820
Zhang WJ, Björn LO (2009) The effect of ultraviolet-B radiation on the accumulation of medicinal compounds in plants. Fitoterapia 80:207–218
Zhang M, Dong JF, Jin HH, Sun LN, Xu MJ (2011) Ultraviolet-B induced flavonoid accumulation in Betula pendula leaves is dependent upon nitrate reductase-mediated nitric oxide signalling. Tree Physiol 31:798–807
Acknowledgments
The authors are thankful to the Head, Department of Botany and Coordinator, Centre of Advanced Study in Botany, Banaras Hindu University, for providing laboratory facilities, and to the University Grants Commission (UGC), New Delhi, for financial assistance. The authors also extend their thanks to Dr. Ajai Kumar, Advanced Instrumentation Research Facility, Jawaharlal Nehru University, New Delhi, for GC–MS analysis.
Author information
Authors and Affiliations
Corresponding author
Additional information
Communicated by J. Gao.
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
About this article

Cite this article
Takshak, S., Agrawal, S.B. Alterations in metabolite profile and free radical scavenging activities of Withania somnifera leaf and root extracts under supplemental ultraviolet-B radiation. Acta Physiol Plant 37, 260 (2015). https://doi.org/10.1007/s11738-015-2014-5
Received:
Revised:
Accepted:
Published:
DOI: https://doi.org/10.1007/s11738-015-2014-5